 2023, Vol. 43
2023, Vol. 43文章信息
- 蔡越, 邢万秋, 王卫光, 吴宇桐, 陈顼
- CAI Yue, XING Wanqiu, WANG Weiguang, WU Yutong, CHEN Xu
- 基于同位素技术的蒸散组分区分采样方案优化研究
- Optimization of sampling scheme for evapotranspiration partition based on isotope technology
- 生态学报. 2023, 43(19): 8076-8086
- Acta Ecologica Sinica. 2023, 43(19): 8076-8086
- http://dx.doi.org/10.20103/j.stxb.202210112894
-
文章历史
- 收稿日期: 2022-10-11
- 网络出版日期: 2023-03-01






2. 河海大学水文水资源学院, 南京 210098
2. College of Hydrology and Water Resources, Hohai University, Nanjing 210098, China
森林生态系统是水分调节和物质交换的重要场所[1], 土壤、植物和大气在地球表层生物圈形成的动态连续系统(soil-plant-atmosphere continuum, SPAC)是陆地水循环过程的重要环节, 水分在其内部的运移规律一直都是水文学等领域的研究热点[2]。植被蒸腾和土壤蒸发作为土壤、植被和大气之间进行水汽交换的两个主要途径, 是陆地生态系统蒸散发的关键组分, 对其量化是探究水分在森林内部水循环过程不同界面特征的基础[3], 植被蒸腾与生态系统蒸散发的比率(T/ET)是准确量化水分利用效率的关键参数[4]。近年来, 自然环境水分中的稳定氢氧同位素(16O和18O、1H和2H)被广泛应用于蒸散组分区分、植物水分溯源、土壤水分运动等研究中[5—6]。蒸散发过程中的水汽相变使同位素发生平衡分馏和动力学分馏[7], 造成土壤蒸发水汽同位素组成δE贫化, 植被蒸腾水汽同位素δT富集, 准确计算二者是基于同位素技术量化T/ET的关键。
根据同位素稳态(Isotope Steady-State, ISS)假设, 即植物体内水同位素在发生蒸腾作用之前不产生分馏, 其与外界水源同位素组成的差异可以反映内部与环境的物质及能量转化[8], 是进行蒸散组分区分相关研究的理论基础。然而越来越多的研究表明植物体内复杂的分馏机制并不完全符合同位素稳态假设[9—10], 各部位水同位素富集程度不一, 且植被之间生理生态特性各异, 对采样部位的选取要求更高。以往关于森林乔木蒸散组分区分研究大多采集植物木质部样品, 农田研究则大多取用作物茎秆部位。例如, 孙守家等[11]使用栓皮栎(Quercus variabilis Blume.)一年生枝条水同位素组成代替δT, 石俊杰等[12]在拆分农田蒸散发研究中则采用玉米(Zea mays Linnaeus.)地表以下茎秆部位。已有研究发现选取植物未发生明显分馏部位需要立足于土壤与植物内部水同位素组成的分布差异[13]。土壤水同位素组成会受降水、地表蒸发及土壤水分运动规律等因素的影响[14], 然而以往多数研究直接采集表层土壤用于计算δE[15—16], 未考虑取样深度对T/ET的影响。杨斌等[17]讨论了δE的主要影响因素, 并表明土壤蒸发前缘深度的选择显著影响δE的准确性。Hu等[18]得出土壤含水率垂向变化较大时不宜将表层土壤作为蒸发前缘。这些研究均表明, 选择土壤及植被的取样位置时应综合考虑当地气候特征、植被类型及土壤特性等因素, 但基于不同取样部位同位素构成特征对于复杂生态系统尤其是森林T/ET的影响来优化各水源采样方案的研究, 目前尚未见报导。
同位素观测技术的进步使得各类生态系统水同位素信息逐渐丰富, 基于同位素手段的蒸散组分区分研究得到发展, 但大多集中于干旱半干旱地区[19—20], 基于多水源采样方案的湿润区森林生态系统蒸散发组分区分研究相对较少。森林影响河流化学成分和水域环保功能, 对流域水源涵养功能有重要调节作用, 精确拆分其蒸散发是理解其SPAC系统水分传输与调控机制的基础, 有利于进一步明晰亚热带森林水文过程。我国人工林50%以上分布在南方亚热带地区[21], 该地区易受季风气候影响, 引发季节性洪涝和干旱事件[22]。新安江流域水资源丰富且水质较好, 流域内有丰乐水库、新安江水库两大蓄水工程, 对下游地区具有防洪、灌溉等综合效益[23]。源区属于亚热带典型湿润区, 植被类型及气候特征复杂, 使得区域水文过程相关研究较为困难。该区典型树种杉木是我国重要造林树种之一, 总面积约占全国林地8.17%, 在我国亚热带地区分布广泛[24]。了解源区森林的生态系统结构特征和水循环过程有助于探究其对下游河库及人类生活生产用水的影响。综上, 本研究将基于新安江源区杉木林多水源样品氢氧稳定同位素测试结果, 分析不同源水同位素组成特征差异, 并通过对比基于不同季节和不同采样部位的T/ET结果优化植被、土壤的采样方案, 为利用同位素技术进行蒸散发组分区分研究提供理论依据。
1 材料与方法 1.1 研究区概况新安江源区属钱塘江水系, 位于安徽省黄山市境内(117°38′E—118°29′E, 29°27′N—30°6′N), 流域控制面积2707.6 km2, 靠近中国东南沿海地区, 地处亚热带季风气候区, 年平均气温为16.5℃, 年平均相对湿度为78%, 降水年内分布不均, 多集中于夏季, 年平均降水量为1747.9 mm。地形以山地和丘陵为主, 地势西高东低, 主要土壤类型包括红壤土、黄壤土、水稻土等, 主要植被类型包括常绿针叶林、落叶阔叶林、混合林、作物地等。
1.2 研究方法 1.2.1 样品采集在研究区涡动通量观测塔(117 °59 ′31 ″E, 29 °40 ′4 ″N)附近选取一个20 m×20 m样地开展调查。样地主要树种为常绿乔木杉木(Cunninghamia lanceolata), 平均高度约8.83 m, 冠层面积约25.26 m2, 主要植物有狗脊(Woodwardia japonica)、毛竹(Phyllostachys heterocycla)等。研究区杉木生长季与非生长季分别为4至10月、11月至次年3月。样品采集时间为2021年9月1至3日和12月9日至11日。选择生长状况良好的杉木于每日不同时段(6:00、9:00、12:00、15:00、18:00)采集韧皮部(高1.5 m处)、木质部(一年生枝条)和叶, 并于样树附近采集分层土壤(深度范围0—1 m, 每10 cm取样一次)。采得样品均密封储存于冷冻环境(-18℃)中待后续水分抽提。采样期每日使用TR-WA5型水汽冷凝系统于0 m以及7.5 m高度进行3—4次大气水汽收集, 并于每日使用60 mL聚乙烯采样瓶采集河道水与山泉水样品2—3次。采样过程中液态水样品均及时经0.22 μm有机质滤头过滤并于低温环境(3—5℃)中冷藏保存。
1.2.2 样品处理与分析样品后续处理和测试工作在河海大学水文水资源与水利工程国家重点实验室中进行。使用LI-2100全自动真空冷凝抽提系统(北京理加联合科技有限公司)对样品水分进行低温真空抽提, 过滤后装入2 mL自动进样瓶中密封冷藏于低温环境(3—5℃)中待测。使用Picarro L2130-i激光同位素分析仪测定同位素组成, 结果用相对于维也纳标准平均海水(Vienna Standard Mean Ocean Water, VSMOW)的千分差值(‰)表示。结果计算如下:
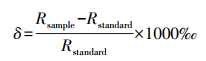
|
(1) |
式中, Rsample和Rstandard分别为水样和VSMOW标样中重同位素与轻同位素的比值, δ18O和δ2H分别为18O/16O和2H/1H。
过量氘(d)计算公式如下:
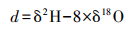
|
(2) |
计算T/ET的关键在于确定δE、δT和δET。其中, 土壤蒸发水汽稳定同位素组成δE的计算涉及多个参数, 包括土壤水同位素组成、土壤温度等, 选取适宜的土壤取样深度对δE计算结果的准确性起决定性作用。杉木林直径2—5 mm的根系有90%分布在0—40 cm土壤层中, 53%总干重的根系分布在10—30 cm土壤层中[24], 因此土壤取样位置对δE的计算十分重要。T/ET由下式基于同位素质量守恒原理计算得出:
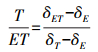
|
(3) |
式中, δE、δT、δET分别为土壤蒸发水汽稳定同位素组成、植物蒸腾水汽同位素组成和蒸散水汽同位素组成, ‰。
土壤蒸发水汽稳定同位素组成(δE)使用Craig-Gordon模型计算:
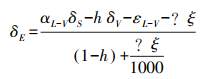
|
(4) |
式中, δS为土壤水同位素组成, ‰;h为土壤温度对应大气相对湿度;δV为大气水同位素组成, ‰;αL-V为水汽相变平衡分馏系数, αL-V=1/α+;;εL-V为平衡分馏效应, Δξ为同位素动力扩散系数。各参数计算公式分别如下:
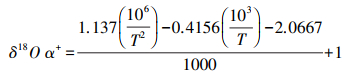
|
(5) |
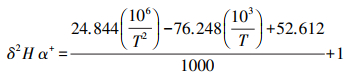
|
(6) |
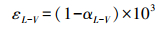
|
(7) |
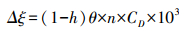
|
(8) |
式中, T为对应深度的土壤温度, K;θ为分子扩散分馏系数与总扩散分馏系数之比, 在土壤蒸发中取1;n描述分子扩散阻力与分子扩散系数的相关性, 在土壤蒸发中取1;CD是描述分子扩散效率的参数, 取常数28.5‰(δ18O)和25.1‰(δ2H)。
根据同位素稳态假设, 本研究植物蒸腾水汽同位素组成δT使用植物不发生明显分馏部位的水同位素组成代替。
Keeling Plot模型描述了大气水汽同位素组成与其浓度倒数之间的线性关系, 利用其拟合得到的曲线在y轴上的截距即为δET:
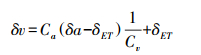
|
(9) |
式中, δV和δa分别为生态系统边界层(距地面7.5 m处)和背景大气(距地面20 m处)水汽同位素组成, ‰;CV和Ca为对应大气水汽浓度。
2 结果 2.1 土壤-植物-大气连续体(SPAC)中不同来源水分稳定同位素组成特征 2.1.1 不同来源水分同位素组成的整体分布特征图 1展示了生长季和非生长季不同来源样品水同位素组成的整体分布范围。各源水δ18O均值从大到小依次为:叶片、木质部、韧皮部、山泉、河道、土壤、大气。根据δ2H排序的结果略有差异:叶片、木质部、韧皮部、土壤、山泉、河道、大气, 可见δ18O和δ2H的变化规律并不完全一致。各源水δ18O和δ2H均呈现非生长季高、生长季低的特点。河道水、山泉水同位素组成的均值较为接近, 二者变幅(9月:山泉水氧同位素组成δ18Os:-7.402‰—-6.818‰;河道水氧同位素组成δ18Og:-7.292‰—-6.786‰;12月:δ18Os:-6.818‰—-6.393‰;δ18Og:-6.510‰—-5.919‰)较小且在土壤水变化范围(9月:土壤水氧同位素组成δ18Osoil:-10.509‰—-3.533‰;12月:δ18Osoil:-9.735‰—-0.795‰)之内。此外, 大气水汽的δ18O和δ2H较其他来源水分明显更为贫化。

|
| 图 1 新安江源区SPAC中不同来源水分稳定同位素组成分布 Fig. 1 Stable isotope compositions of water from different sources in SPAC in the source area of Xin′an River SPAC: 土壤-植物-大气多源水样 |
由表 1得知, 杉木韧皮部与土壤的水同位素组成均值最为接近, 整体分布重合范围比例较高。由非生长季杉木韧皮部与土壤之间水同位素组成均值的差异(Δδ18O=1.403‰, Δδ2H=0.350‰)可见, 这一特征在非生长季尤其明显, 说明水分从土壤被根系吸收到根茎部位的过程中几乎不发生分馏。然而, 当水分到达叶片时, 强烈的蒸腾作用使得动力分馏加剧, 因此叶片水重同位素较其他部位更为富集。木质部水δ18O和δ2H的分布范围介于叶片与韧皮部之间, 说明此处水同位素已产生明显分馏。根据同位素稳态假设, 结合杉木各部位水同位素组成分布特征, 得出本研究区杉木未发生明显分馏的适宜采样部位为韧皮部。
| 研究 Research |
植物 Plant species |
采样期 Sampling period |
部位 Parts |
氧同位素组成(δ18O) Oxygen isotope composition |
氢同位素组成(δ2H) Hydrogen isotopic composition |
18O富集程度(Δ18OL) Oxygen isotope enrichment |
2H富集程度(Δ2HL) Hydrogen isotope enrichment |
过量氘(d) Excess deuterium |
| 本研究This study | 杉木C.lanceolata | 生长季 | 韧皮部 | -5.266±1.470 | -54.174±6.269 | 12.091±3.772 | 39.669±11.214 | -12.044±6.550 |
| 叶片 | 6.825±3.906 | -14.506±10.503 | -69.103±22.769 | |||||
| 非生长季 | 韧皮部 | -3.869±1.366 | -43.011±6.618 | 15.347±4.666 | 48.954±10.942 | -12.061±6.124 | ||
| 叶片 | 11.479±5.035 | 5.942±11.616 | -85.886±29.378 | |||||
| 李龙等[30], 2020 | 竹柏P. nagi | 雨季 | 木质部 | -3.9 | -47.8 | 27.9 | 69.1 | -16.6 |
| 叶片 | 24.0 | 21.3 | -170.5 | |||||
| 旱季 | 木质部 | -3.3 | -31.6 | 12.6 | 38.1 | -5.2 | ||
| 叶片 | 9.3 | 6.5 | -70.5 | |||||
| 王锐等[29], 2021 | 樟树C. camphora | 湿润期 | 茎秆 | -5.66±1.54 | 13.12±5.47 | |||
| 叶片 | 7.46±5.47 | |||||||
| 干旱期 | 茎秆 | -6.34±0.37 | 11.48±5.13 | |||||
| 叶片 | 5.14±5.16 | |||||||
| δ18O:氧同位素组成Hydrogen isotopic composition;δ2H:氢同位素组成Hydrogen isotopic composition;Δ18OL:18O富集程度Oxygen isotope enrichment;Δ2HL:2H富集程度Hydrogen isotope enrichment;d:过量氘Excess deuterium | ||||||||
土壤水和韧皮部水δ18O-δ2H分布在大气水线GMWL(global meteoric water line of Craig)右下侧, 而河道水、山泉水和大气水则相反(图 2)。河道水和山泉水δ18O-δ2H分布集中于GMWL附近, 表明采样期地表水和地下水主要通过降水补给, 同位素较为稳定, 反映出水源受环境影响较小。12月山泉水蒸发线更靠近GMWL, 且斜率(ΔL9月=4.301;ΔL12月=6.921)大于河道水蒸发线(ΔL9月=2.352;ΔL12月=4.173), 非生长季结果的斜率明显大于生长季, 反映了不同水体和季节的蒸发分馏效应强弱。土壤水蒸发线(δ2Hsoil= 5.913δ18Osoil-13.207, R2= 0.868)的斜率和截距均小于GMWL, 表示降水入渗过程中产生了一定程度的蒸发分馏效应。通过比较木质部、叶片、韧皮部三者水分蒸腾线(δ2Hb= 3.360δ18Ob- 32.141, R2= 0.819;δ2HL= 2.717δ18OL-29.474, R2= 0.824;δ2Hphloem= 4.803δ18Ophloem- 26.844, R2= 0.798)的斜率, 得出杉木体内蒸腾作用和动力学分馏效应由强到弱依次为:叶片、木质部、韧皮部。非生长季各源水δ18O-δ2H分布较生长季更接近GMWL, 表明非生长季蒸发蒸腾过程中同位素动力学分馏效应弱于生长季。大气水汽δ18O-δ2H(δ2Hv= 6.393δ18Ov-6.087, R2=0.449)的斜率和截距与GMWL均有明显差距, 说明研究区大气水汽的主要来源除了降水还有植被蒸散发水汽和区域外大气水汽。

|
| 图 2 土壤水、韧皮部水、木质部水、河道水、山泉水、大气水同位素组成不同季节δ18O-δ2H特征 Fig. 2 Characteristics of δ18O-δ2H of soil water, phloem water, xylem water, ground water, spring water and atmospheric water vapor in different seasons |
生长季采样期多层土壤水δ18O和δ2H的日变幅较小(图 3), 9月1日各层土壤水同位素组成整体均值为-8.366‰(δ18Osoil)和-63.462‰(δ2Hsoil), 9月3日为-8.380‰(δ18Osoil)和-63.496‰(δ2Hsoil), 均值差异较小且分布范围接近, 9月3日结果分布较为均匀。非生长季采样期土壤水δ18O和δ2H变幅和均值明显大于9月, 三日δ18Osoil均值分别为-5.149‰、-7.409‰、-7.620‰, δ2Hsoil均值分别为-52.956‰、-57.617‰、-54.089‰。土壤水同位素在较长时间尺度上受降水影响, 结合12月降水量远低于9月的情况可知, 在季节尺度上当降水量越大时, 土壤水同位素组成越负。12月11日发生降水事件, 土壤水同位素却更加富集, 短时间内降水量较小时对深层土壤水同位素组成的影响不明显。

|
| 图 3 土壤水稳定同位素组成δ18O和δ2H日变化特征及采样期日降水量 Fig. 3 Diurnal variation characteristics of stable isotopic compositions δ18O and δ2H of soil water and daily precipitation during the sampling period |
研究区生长季与非生长季土壤水δ18O和δ2H的剖面变化规律见如图 4, 二者变化具有一致性, 深层土壤水同位素组成较浅层明显偏负。土壤水δ18O和δ2H每日整体变化规律不一, 呈随深度增加而持续减小、先增大后减小及先减小后增大三种形式。其中, 9月1日土壤水同位素组成于10—70 cm处缓慢下降, 经80 cm处拐点后至100 cm过程中大幅上升。9月3日土壤水同位素组成于10—50 cm深度呈稳定上升趋势, 拐点位于50—60 cm。12月9日土壤水同位素组成整体随深度增加而下降, 30—90 cm变幅较广。12月10日土壤水同位素组成变化波动较大, 经多处拐点, 于20—30 cm和90—100 cm范围内上升, 其余均呈下降趋势。12月11日土壤水δ18O和δ2H随深度增加呈先下降后小幅上升的趋势不同季节浅层土壤水同位素组成分布差异较大, 变化量达4.989‰(δ18Osoil)和34.264‰(δ2Hsoil), 随着深度的增加, 差距逐渐减小并趋于稳定, 变化量仅为2.032‰(δ18Osoil)和11.188‰(δ2Hsoil)。观察到浅层变化范围明显大于深层, 深层土壤水同位素组成变化具有一定滞后性。

|
| 图 4 不同深度(0—100 cm)土壤水稳定同位素组成δ18O和δ2H日变化特征 Fig. 4 Diurnal variation of stable isotopic compositions δ18O and δ2H in soil water at different depths (0—100 cm) |
本研究选取0—10 cm、20—30 cm和40—50 cm三种深度土壤样品测定数据对δE和T/ET进行计算, 结果如图 5所示。在土壤垂向物理特性变化方面, 9月采样期0—50 cm土壤平均温度为25.31℃, 随土壤深度的增加而递减。12月采样期土壤平均温度为11.35℃, 随土壤深度的增加呈递增趋势。9月土壤含水率略高于12月, 且垂向变化差异更明显。韧皮部水同位素组成与各层土壤水同位素组成差距不一(图 5), 主要原因为杉木根部吸收水分来源于不同深度的土壤。9月基于三种深度土壤样品计算所得δE整体变化量为29.207‰(δ18Osoil)和226.028‰(δ2Hsoil), 12月结果明显高于9月(12月水同位素组成整体变化量Δδ18Osoil =42.629‰;Δδ2Hsoil =302.881‰)。由图 5明显看出, δE随土壤深度增加而减小, T/ET结果则相反。9月基于各层土壤样品所得T/ET均值(δ18Osoil和δ2Hsoil的结果均值)为83.35%, 高于12月结果78.20%。9月T/ET的整体变化量为8.82%(δ18Osoil)和18.09%(δ2Hsoil), 低于12月结果19.75%(δ18Osoil)和23.08%(δ2Hsoil), 说明在寒冷干燥的气候条件下, 土壤取样深度的选择对δE和T/ET的计算结果影响更大。

|
| 图 5 各源水同位素组成(土壤水、韧皮部水及土壤蒸发水汽)以及基于不同土壤深度的T/ET结果 Fig. 5 Water isotopic composition of different water sources (soil water, phloem water and soil evaporation vapor) and T/ET results based on different soil depths T/ET: 植被蒸腾与生态系统蒸散发的比例 |
研究区SPAC系统各源水δ18O和δ2H的整体分布存在明显季节性变化, 呈现非生长季偏正、生长季偏负的规律, 这可能是亚热带季风气候区各个季节之间的水汽来源及蒸发条件不同所导致[25]。由于非生长季降水较少, 大气水汽同位素在运输过程中贫化程度较弱, 所以较生长季明显富集。森林水分主要来自大气降水和区域之间的水汽交互, 因此降水同位素组成的季节性变化能够反映其他源水同位素组成的规律[26]。已有研究表明, 我国东南沿海地区降水的δ18O与降水量存在明显负相关关系, 符合研究区多雨生长季SPAC系统的水同位素组成较少雨的非生长季更偏负的特点[27]。
在季节尺度上, 植物水分δ18O和δ2H的整体范围与降水存在关联, 然而其分馏机制主要受环境影响, 气候及下垫面等条件的差异会导致植物叶片水同位素富集程度不同。表2展示了不同植物体内的水同位素组成和分馏程度。对比发现, 本研究中生长季土壤水和韧皮部水同位素组成均较非生长季偏负, 其他研究中植物水分同位素组成普遍在湿润期更为富集。在季节尺度上, 湿润期充足的水分条件使得叶片水与大气水之间的交换更为频繁, 因此韧皮部与木质部之间的水同位素差距Δ18OL和Δ2HL更高[28]。王锐等[29]在亚热带针阔混交林水分同位素组成研究中发现, 季节变化上, Δ18OL与气温无显著相关关系, 而与相对湿度、太阳辐射等相关性较高。上述研究与本研究区域气候特征相似且叶片水富集程度(湿润期Δ18OL=13.12‰, 干旱期Δ18OL=11.48‰)差异较小。本研究生长季的植物叶片水Δ18OL和Δ2HL(Δ18OL=12.091‰;Δ2HL=39.669‰)略低于非生长季(Δ18OL=15.347‰;Δ2HL=48.954‰), 两季采样期之间平均温差较大, 高达17.31℃, 平均相对湿度仅相差1.46%。李龙等[30]的亚热带常绿阔叶林同位素组成特征研究中, 植物叶片水δ18O和δ2H富集程度在雨季(Δ18OL=27.9‰;Δ2HL=69.1‰)和旱季(Δ18OL=12.6‰;Δ2HL=38.1‰)均强于本研究。说明除环境因素外, 植被生理生态特征如叶片性状等也会影响叶片水的蒸腾速率和分馏强度[31—32], 这些因素对植物叶片水同位素组成富集程度的影响机制仍有待进一步研究。
3.2 不同研究土壤水同位素组成垂向特征大气降水、土壤水分运动等因素均会对土壤水同位素产生影响。徐庆等[33]在四川高山暗针叶林土壤水同位素组成研究中, 发现δ2H随深度增加呈先降低后增加的趋势, 垂向变化拐点位于5—20 cm, 与本研究的拐点分布范围50—90 cm差异明显, 原因与采样期降水条件、当地土壤性状和植物根系分布等均有一定关系。侯士彬等[14]在太行山土壤水同位素组成规律研究中发现, 刺槐林和侧柏林土壤水同位素组成垂向变化的拐点在50 cm左右, 荒草地则为30 cm左右。本研究土壤水垂向分布整体规律呈现三种类型, 随深度增加而减小、先增大后减小或先减小后增大, 整体呈现深层土壤水同位素组成较浅层更偏负的特征。土壤水同位素组成的变化与土壤水层响应快慢有关, 体现了降水入渗和蒸散发共同作用的复杂影响机制。浅层的土壤水同位素组成的变幅明显大于深层, 表明浅层土壤水更容易受大气降水和土壤蒸发等外界因素影响。降水同位素组成在经过下渗过程中的蒸发作用后会更加富集, 而本研究中的深层土壤水同位素组成明显较浅层更为贫化, 对降水的响应较弱, 由此推断出采样期内深层土壤水接受降水的补给量较少。深层土壤水富集程度明显不如浅层土壤水, 并且与山泉水同位素组成较为接近, 表明深层土壤水蒸发作用较弱, 接受相对贫化的地下水补给量较多, 体现了深层土壤水、地下水对一个地区SPAC系统内水分同位素组成的变化的缓冲作用[34]。
3.3 不同研究蒸散发组分区分研究中土壤取样深度比较土壤水的蒸发作用是其与大气进行水汽交换的主要途径之一[35], 土壤蒸发水汽同位素组成δE是量化T/ET的关键, 土壤取样位置决定了δE结果的准确性。当土壤垂向性状差异较大时, 基于不同深度土壤样品的计算结果差异较大[18], 亟需对土壤采样方案进行优化。大气及土壤中的水同位素组成作为模型输入要素, 其大小对结果产生直接影响。气象因子和土壤特性则通过影响对应土壤层的标准化相对湿度和分馏系数间接影响结果。研究表明, 平衡分馏系数对δE的影响不显著, 而相对湿度对δE结果影响较大[17]。本研究区非生长季的土壤特性垂向变化差异较大, 直接导致标准到土壤温度的大气相对湿度产生明显差距, 且土壤水同位素组成跨度较大, 使得基于不同采样深度的结果产生较大差异。同时, 降水入渗过程和土壤水蒸发过程共同影响下的各层土壤水同位素易产生剧烈变化[14], 导致T/ET结果产生差异。以往研究通常选取表层土壤样, 而研究结果表明, 当土壤含水率垂向跨度较大时, 选取土壤浅层作为取样位置易低估T/ET。当取样深度增大时, 所得δE结果贫化程度加剧, 导致T/ET过于趋近于1, 从而影响T/ET结果的准确性, 因此深层土壤不宜作为取样位置。本研究结果显示, 40—50 cm土壤层δE过偏负, 使T/ET结果过高。20—30 cm土壤水同位素组成与植物韧皮部水同位素组成接近, 因此可以推断研究区基于同位素进行蒸散组分区分研究的最佳土壤取样深度为20—30 cm。不同区域的土壤蒸发前缘位置选择有所差异, 下垫面均一的玉米田蒸散组分区分研究中选择5 cm作为取样深度[12]。黑河上游森林蒸散组分区分研究中, 区域气候干燥, 土层较薄, 选取0—10 cm土壤水同位素组成均值进行δE的计算[36]。Hu等[18]在内蒙古半干旱草地蒸散组分区分研究中, 得出适合该研究区蒸散组分区分计算的土壤取样深度为15 cm。基于同位素手段的蒸散组分区分研究应选取适宜土壤深度, 尤其是当土壤垂向特性变化较大以及降水事件后各层土壤水同位素组成发生复杂变化的情况下, 准确选取采样位置能够降低模型中各个参数的不确定性, 避免计算误差。
4 结论新安江源区SPAC系统多源水δ18O和δ2H存在明显季节性差异, 非生长季各来源水分同位素组成较生长季更富集, 且δ18O-δ2H更接近GMWL。通过比较蒸腾线斜率得出, 生长季植物分馏效应和蒸腾作用明显强于非生长季。水分在土壤-植物体内各部位运移过程中同位素组成逐渐富集, 韧皮部水和土壤水同位素组成分布差异最小, 说明韧皮部部位的水同位素分馏程度最符合稳态假设, 基于此选择韧皮部部位水同位素组成作为δT。不同季节的土壤水同位素组成变化规律差异较小, 深层土壤水同位素组成更加贫化。土壤水同位素组成垂向变化的拐点主要位于50—90 cm, 剖面变化规律不一, 主要有呈随深度增加而减小、先增大后减小或先减小后增大三种形式。深层土壤水同位素组成的变化具有滞后性, 变幅明显小于浅层土壤水。比较基于三种深度土壤样品的δE和T/ET结果, 发现研究区处于寒冷干燥的非生长季时, 土壤深度的不同选择使结果产生较大的差异, 通过对比木质部与土壤水分同位素构成优化采样方案, 最终选择20—30 cm作为拆分研究区典型针叶林生态系统蒸散组分的土壤适宜取样位置。
| [1] |
赵同谦, 欧阳志云, 郑华, 王效科, 苗鸿. 中国森林生态系统服务功能及其价值评价. 自然资源学报, 2004, 19(4): 480-491. DOI:10.3321/j.issn:1000-3037.2004.04.010 |
| [2] |
Zhang C Z, Zhang J B, Zhao B Z, Zhang H, Huang P. Stable isotope studies of crop carbon and water relations: a review. Agricultural Sciences in China, 2009, 8(5): 578-590. DOI:10.1016/S1671-2927(08)60249-7 |
| [3] |
Wang K C, Dickinson R E. A review of global terrestrial evapotranspiration: observation, modeling, climatology, and climatic variability. Reviews of Geophysics, 2012, 50(2): Rg2005. |
| [4] |
Beringer J, Hutley L B, Hacker J M, Neininger B, Paw U K T. Patterns and processes of carbon, water and energy cycles across northern Australian landscapes: from point to region. Agricultural and Forest Meteorology, 2011, 151(11): 1409-1416. DOI:10.1016/j.agrformet.2011.05.003 |
| [5] |
张玉翠, 孙宏勇, 沈彦俊, 齐永青. 氢氧稳定同位素技术在生态系统水分耗散中的应用研究进展. 地理科学, 2012, 32(3): 289-293. |
| [6] |
Wang P, Yamanaka T, Li X Y, Wei Z W. Partitioning evapotranspiration in a temperate grassland ecosystem: numerical modeling with isotopic tracers. Agricultural and Forest Meteorology, 2015, 208: 16-31. DOI:10.1016/j.agrformet.2015.04.006 |
| [7] |
Craig, H., Gordon, L.I. Deuterium and oxygen 18 variation in the ocean and the marine atmosphere. In Proceedings of the Conference on Stable Isotopes in Oceanographic Studies and Paleotemperatures, 1965: 9-130.
|
| [8] |
孙双峰, 黄建辉, 林光辉, 赵威, 韩兴国. 稳定同位素技术在植物水分利用研究中的应用. 生态学报, 2005, 25(9): 2362-2371. DOI:10.3321/j.issn:1000-0933.2005.09.032 |
| [9] |
Lee X H, Kim K, Smith R. Temporal variations of the 18O/16O signal of the whole-canopy transpiration in a temperate forest. Global Biogeochemical Cycles, 2007, 21(3): GB3013. |
| [10] |
Farquhar G D, Cernusak L A. On the isotopic composition of leaf water in the non-steady state. Functional Plant Biology: FPB, 2005, 32(4): 293-303. DOI:10.1071/FP04232 |
| [11] |
孙守家, 孟平, 张劲松, 何春霞, 郑宁. 华北低丘山区栓皮栎生态系统氧同位素日变化及蒸散定量区分. 生态学报, 2015, 35(8): 2592-2601. |
| [12] |
石俊杰, 龚道枝, 梅旭荣, 马孝义, 郝卫平, 胡笑涛. 稳定同位素法和涡度-微型蒸渗仪区分玉米田蒸散组分的比较. 农业工程学报, 2012, 28(20): 114-120. |
| [13] |
温学发, 张世春, 孙晓敏, 于贵瑞. 叶片水H218O富集的研究进展. 植物生态学报, 2008, 32(4): 961-966. DOI:10.3773/j.issn.1005-264x.2008.04.026 |
| [14] |
侯士彬, 宋献方, 于静洁, 刘鑫, 张广英. 太行山区典型植被下降水入渗的稳定同位素特征分析. 资源科学, 2008, 30(1): 86-92. DOI:10.3321/j.issn:1007-7588.2008.01.013 |
| [15] |
王渝淞, 贾国栋, 张永娥, 郑鹏飞, 朱栩辉, 孙乐乐, 余新晓. 北京山区侧柏林蒸散拆分研究. 水土保持学报, 2019, 33(2): 272-278. DOI:10.13870/j.cnki.stbcxb.2019.02.042 |
| [16] |
Aron P G, Poulsen C J, Fiorella R P, Matheny A M, Veverica T J. An isotopic approach to partition evapotranspiration in a mixed deciduous forest. Ecohydrology, 2020, 13(6): e2229. DOI:10.1002/eco.2229 |
| [17] |
杨斌, 谢甫绨, 温学发, 孙晓敏, 王建林. 华北平原农田土壤蒸发δ18O的日变化特征及其影响因素. 植物生态学报, 2012, 36(6): 539-549. |
| [18] |
Hu Z M, Wen X F, Sun X M, Li L H, Yu G R, Lee X H, Li S G. Partitioning of evapotranspiration through oxygen isotopic measurements of water pools and fluxes in a temperate grassland. Journal of Geophysical Research: Biogeosciences, 2014, 119(3): 358-372. DOI:10.1002/2013JG002367 |
| [19] |
Fatichi S, Pappas C. Constrained variability of modeled T: ET ratio across biomes. Geophysical Research Letters, 2017, 44(13): 6795-6803. DOI:10.1002/2017GL074041 |
| [20] |
Zhou S, Yu B F, Zhang Y, Huang Y F, Wang G Q. Partitioning evapotranspiration based on the concept of underlying water use efficiency. Water Resources Research, 2016, 52: 1160-1175. |
| [21] |
Sun X M, Wen X F, Yu G R, Liu Y F, Liu Q J. Seasonal drought effects on carbon sequestration of a mid-subtropical planted forest of southeastern China. Science in China Series D: Earth Sciences, 2006, 49(2): 110-118. |
| [22] |
Tang Y K, Wen X F, Sun X M, Wang H M. Interannual variation of the Bowen ratio in a subtropical coniferous plantation in southeast China, 2003-2012. PLoS One, 2014, 9(2): e88267. |
| [23] |
杨迪虎. 新安江流域安徽省地区水环境状况分析. 水资源保护, 2006, 22(5): 77-80. |
| [24] |
王兵, 马向前, 郭浩, 王燕, 冷泠. 中国杉木林的生态系统服务价值评估. 林业科学, 2009, 45(4): 124-130. |
| [25] |
卫克勤, 林瑞芬. 论季风气候对我国雨水同位素组成的影响. 地球化学, 1994, 23(1): 32-41. |
| [26] |
邓文平, 余新晓, 贾国栋, 李亚军, 刘玉洁. 北京西山鹫峰地区氢氧稳定同位素特征分析. 水科学进展, 2013, 24(5): 642-650. |
| [27] |
章新平, 姚檀栋. 我国降水中δ18O的分布特点. 地理学报, 1998, 53(4): 70-78. |
| [28] |
Butt S, Ali M, Fazil M, Latif Z. Seasonal variations in the isotopic composition of leaf and stem water from an arid region of Southeast Asia. Hydrological Sciences Journal, 2010, 55(5): 844-848. |
| [29] |
王锐, 章新平, 戴军杰, 罗紫东, 贺新光, 关华德. 亚热带针阔混交林土壤-植物-大气连续体(SPAC)中水稳定同位素特征. 生态环境学报, 2021, 30(6): 1148-1157. |
| [30] |
李龙, 唐常源, 曹英杰. 亚热带地区常绿阔叶林SPAC系统水分的氢氧稳定同位素特征. 应用生态学报, 2020, 31(9): 2875-2884. |
| [31] |
罗伦, 余武生, 万诗敏, 周平. 植物叶片水稳定同位素研究进展. 生态学报, 2013, 33(4): 1031-1041. |
| [32] |
李嘉竹, 刘贤赵. 氢氧稳定同位素在SPAC水分循环中的应用研究进展. 中国沙漠, 2008, 28(4): 787-794. |
| [33] |
徐庆, 刘世荣, 安树青, 蒋有绪, 林光辉. 四川卧龙亚高山暗针叶林土壤水的氢稳定同位素特征. 林业科学, 2007, 43(1): 8-14. |
| [34] |
田立德, 姚檀栋, M TSUJIMURA, 孙维贞. 青藏高原中部土壤水中稳定同位素变化. 土壤学报, 2002, 39(3): 289-295. |
| [35] |
Gat J R. The isotopic composition of evaporating waters-review of the historical evolution leading up to the Craig-Gordon model. Isotopes in Environmental and Health Studies, 2008, 44(1): 5-9. |
| [36] |
彭文丽, 赵良菊, 谢聪, 董玺莹, 刘全玉, 李瑞峰, 潘昭烨. 黑河上游青海云杉森林生态系统蒸散发分割. 冰川冻土, 2020, 42(2): 629-640. |