 2023, Vol. 43
2023, Vol. 43文章信息
- 游巍斌, 蔡新瑜, 王英姿, 王瑞, 张锦琳, 陈莹, 邱晓月, 方冰, 汤绍圣, 坚文倩
- YOU Weibin, CAI Xinyu, WANG Yingzi, WANG Rui, ZHANG Jinlin, CHEN Ying, QIU Xiaoyue, FANG Bing, TANG Shaosheng, JIAN Wenqian
- 基于不同网格尺度福州主城区鸟类多样性与景观特征的关系研究
- Relationship between bird diversity and landscape characteristics in the core area of Fuzhou city at two scales based on citizen science data
- 生态学报. 2023, 43(18): 7670-7681
- Acta Ecologica Sinica. 2023, 43(18): 7670-7681
- http://dx.doi.org/10.20103/j.stxb.202306071214
-
文章历史
- 收稿日期: 2023-06-07
- 采用日期: 2023-08-30





2. 福建农林大学林学院, 福州 350002;
3. 福建省南方森林资源与环境工程技术研究中心, 福州 350002;
4. 福州市自然保护地规划发展中心, 福州 354300;
5. 福州市林学会, 福州 354300;
6. 中国农业大学资源与环境学院, 北京 100193;
7. 福州市晋安河公园管理处, 福州 350011;
8. 福州市闽江公园管理处, 福州 350206
2. College of Forestry, Fujian Agricultural and Forestry University, Fuzhou 350002, China;
3. Fujian Southern Forest Resources and Environmental Engineering Technology Research Center, Fuzhou 350002, China;
4. Fuzhou Nature Reserve Planning and Development Center, Fuzhou 354300, China;
5. Fuzhou Forestry Society, Fuzhou 354300, China;
6. College of Resources Environmented Sciences, China Agricultural University, Beijing 100193, China;
7. Fuzhou Jin'an River Park Management Office, Fuzhou 350011, China;
8. Fuzhou Minjiang Park Management Office, Fuzhou 350206, China
快速城市化极大地改变了城市原有的自然景观格局, 人工地表的蔓延使得野生动物的栖息地进一步破碎化, 城市内部物种多样性受到严重威胁[1—2]。生物多样性的自然本底属性决定了其与城市生态安全的关系[3], 多样性水平降低通过影响到城市生态系统功能有效发挥进而给居民福祉的获得构成威胁[4]。在城市生物多样性相关研究中, 鸟类多样性水平深刻影响着城市居民身心健康[5];鸟类在城市内对破碎化的栖息环境适应能力强, 鸟种繁多, 对其多样性的调查技术也较为成熟[4, 6];城市鸟类常被作为城市生物多样性水平的指示物种[7];鸟类多样性研究备受关注[5, 8]。
过去20年, 国内外学者围绕不同尺度(梯度)下城市鸟类生态分布、多样性水平、群落组成、动态变化及其影响因素等领域开展了积极探索[6, 9—13]。在研究方法的选择上, 国内研究主要采用样点法和样线法等传统监测方法居多[4, 14—15];由于观鸟活动的较长发展历史和普及(如圣诞鸟类调查、eBird大型公民观鸟数据平台), 国外研究者使用公民科学数据来探讨鸟类多样性方面更加广泛[2, 16—19]。公民科学是指非专业人士的科学调查研究;其常活跃于生物多样性的保护与监测中, 较为常见则是观鸟活动[20—21];鸟类具有易于观察、分布广泛、鸟种丰富且色彩多样的特点, 能够吸引爱好者对其进行观察[22]。我国的公民观鸟活动虽起步较晚, 但近20年来发展迅速[20]。公民观鸟作为一种监测物种多样性的新手段, 已有研究表明其所提供的数据与专家提供的数据集同样具有价值, 亦为研究者提供开展鸟类研究的新途径[23—26]。
当前城市鸟类多样性与景观因素关系研究主要集中在植被组成、绿地面积、建筑密度、建筑高度等二维或三维结构特征因子的影响方面, 而刻画景观功能性特征方面的因素较少被纳入考虑。我们对不同尺度下基于公民数据分析得到的城市鸟类多样性与影响因子作用关系及其影响程度方面的知识仍知之甚少[21, 27—28]。本研究选择福建省会省福州市的主城区为研究区, 近年来经过快速城市化的福州市大力推进森林城市的建设, 特别是在主城区通过添景增绿工程、串珠公园建设等系列行动改善城市内河水系、提升了主城区绿化覆盖率, 这为开展本研究提供了理想场所。因此, 本研究在爬取的研究区公众观鸟数据的基础上, 探究两种尺度下景观特征因子与城市鸟类多样性的关系, 试图回答以下问题:(1)不同尺度下福州主城区鸟类组成及多样性特征;(2)影响研究区鸟类多样性的关键景观特征因素及其影响程度;旨在为城市鸟类生物多样性保护和生态城市建设提供参考。
1 研究区概况与研究方法 1.1 研究区概况福州市为福建省省会, 地处亚热带, 气候暖湿、年相对湿度约为77%、森林茂密、自然生态资源丰富、素有滨江滨海生态园林城市之称。其地形地貌为典型的河口盆地, 主要地形为山地和丘陵(占全市土地面积的72.68%)。2014年福州市获批创建国家森林城市, 开展以内河水系为纽带的串珠公园建设等一系列国土绿化行动;至2020年城区绿化覆盖率已达到了44.93%, 晋安区北部、东部、鼓楼区北部植被覆盖度相对较高。同时, 随着福州城市化的快速发展, 2020年全市城镇化水平已经达到72.5%;福州主城区人口密集且城镇化率接近百分之百。考虑福州市区域范围大(下辖6个区、6个县和1县级市)且城郊区域城市公民数据匮乏, 故本文仅选择城市化历史最早、程度最高的福州市三环道路范围内的主城区为研究对象, 涉及鼓楼区、台江区、仓山区及其晋安区的部分区域(图 1)。

|
| 图 1 研究区概况 Fig. 1 Overview of the study area |
(1) 公民观鸟数据的采集:鸟类观测数据来自中国观鸟记录中心(www.birdreport.cn), 选取研究区范围内2020年1月—12月公众在该平台上发布的436观鸟报告(累计观测6496只), 提取信息包括:观鸟时间、观测点经纬度坐标、鸟种(目、科和种)及其相应数量。参照《中国鸟类分类与分布(第三版)》《国家重点保护野生动物名录》(2021年版)和世界自然保护联盟IUCN(2022)对鸟类居留型、保护等级和受威胁等级进行分类。(2)不同尺度下鸟类多样性指数计算:鉴于当前广泛使用的生态地理数据的中、低分辨率多为30 m和1000 m, 结合鸟类在城市中的活动特征(杨刚等, 2015)本研究选择300 m(30的整倍数)和1000 m两个尺度探究不同尺度对研究结果的影响。采用ArcGIS工具的fishnet工具将研究区划分为300 m和1000 m两个尺度空间网格, 分别计算不同尺度下各网格中表征鸟类多样性特征的3个指数:丰富度(Richness)指数(S)、Shannon-Wiener多样性指数(H)和Simpson多样性指数(D)。同时记录每个网格中公众的观鸟报告数量(Number of report, NR)。上述多样性指数公式如下:
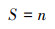
|
(1) |

|
(2) |
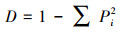
|
(3) |
式中, n为每个网格记录到的物种数;Pi为第i个物种个体数占每个网格总个体数的比例。
1.2.2 城市景观结构特征的识别与指标量化(1) 城市二维土地利用类型的分类与提取:将研究区土地利用划分为城市绿地、水体、居民区和城市道路4大类;为探究不同城市绿地类型对鸟类多样性的影响, 参考《城市绿地分类标准》(CJJ/T85—2017)将城市绿地进一步细分为:公园绿地、防护绿地、广场绿地、附属绿地和区域绿地。在此基础上, 分别计算两个尺度网格单元中绿地总面积占比(GA)、公园绿地面积占比(G1)、防护绿地面积占比(G2)、广场绿地面积占比(G3)、附属绿地面积占比(XG)、区域绿地面积占比(EG)、水体面积占比(WA)、居住区面积占比(RA)、城市道路面积占比(RD)等潜在景观二维结构特征因子。土地利用类型识别通过整合研究区worldview影像(0.5 m×0.5 m)及其零星地块的无人机航飞影像开展人工目视解译实现。(2)城市景观建筑三维特征的提取:在提取的研究区居住区单个建筑高度值的基础上, 采用ArcGIS分区统计工具计算不同尺度下各格网中的建筑平均高度(HT, m)、建筑群高度变异程度(H_SD, m)和建筑体积(BV, 万m3)等3个景观三维结构特征指标。
1.2.3 城市景观功能特征的识别与指标量化遥感生态指数(Remote Sensing Ecological Index, RSEI)是由徐涵秋[29]提出的用来表征区域生态环境质量的指数;其由湿度分量(WET), 归一化植被指数(normalized difference vegetation index, NDVI), 干度指数(normalized difference built-up and soil index, NDBSI)和地表温度(Land Surface Temperature, LST)所构成, 指标来源完全基于遥感信息, 具有获取简单, 计算客观的特点, 因而表征出来的结果也更为可靠, 其值越接近1时代表生态环境质量越好。因此, 由于RSEI指数集成了4个分量的综合信息, 本研究假定RSEI指数在宏观上能够表征城市景观分析网格中的平均生态功能质量, 故将RSEI作为表征景观网格生态功能的代替指标。RSEI指标信息中包含NDVI指数, 因此在使用时应当采用植物生长季节的地表反射率数据[30], 为了避免季节原因对RSEI造成的影响, 选取同年6到10月的遥感影像进行RSEI指数计算。采用Google Earth Engine平台计算了研究区内不同尺度下各网格中RSEI的平均值。遥感生态指数的具体计算过程参见文献29。
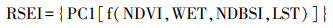
|
(4) |
式中, RSEI为生态环境质量指数, PC1表示第一主成分, NDVI是归一化植被指数, WET是湿度分量, NDBSI是干度指数, LST是地表温度。
1.2.4 统计分析方法采用Mann-Whitney U检验两个尺度下3个鸟类多样性指数的差异性。采样Moran′s I指数检测鸟类多样性指数的空间自相关特征。为探究影响鸟类多样性指数(S, H和D)的关键景观因子, 首先, 采用Spearman偏相关系数计算在控制观鸟报告数量NR条件下与鸟类多样性指数显著相关的主要景观因子(P<0.05), 进而剔除网格中观测报告数量NR可能对结果的影响, 最终获得GA、RD、RA、WA、HT、H_SD、BV和RSEI等8个因子纳入后续回归分析;其次, 采用广义线性模型(generalize linear model, GLM)分别分析3个多样性指数(因变量)与8个景观因子(自变量)的关联性。根据因变量数据分布特点, 采用伪泊松分布link函数拟合S指数, 采用高斯分布link函数拟合H和D指数, 并在拟合前使用方差膨胀因子(剔除VIF>5的变量)诊断自变量间的多重共线性。同时,为分析量纲不同的自变量对因变量的影响程度, 在执行广义线性模型多元回归之前, 将每个自变量标准化(均值0和标准差1), 以便通过比较标准化的回归系数来确定自变量影响的强度差异[31], 进而确定对城市鸟类多样性指数有显著影响的关键景观因子及其影响程度。采用R 4.4.2进行统计分析和制图。
2 结果与分析 2.1 基于公民数据的城市鸟类基础特征 2.1.1 鸟类组成与濒危保护鸟种空间分布2020年研究区范围内公众共观测到鸟类242种, 隶属19目59科, 共6496只;其中以雀形目鸟类最多(158种, 4968只), 占鸟类总数量的76.48%;其次为鹈形目(8种, 442只, 占比为6.80%);数量最少的为犀鸟目(1种, 2只)。此外, 鸽形目、鹰形目、佛法僧目、鹤形目、鸻形目、形目、鹃形目数量占比低于5%;鸡形目、目、鸮形目、雁形目、咬鹃目、雨燕目、隼形目、夜鹰目、鲣鸟目、犀鸟目数量占比均低于1%。
从鸟种来看, 观测数量排名前10的鸟种累计占比28.45%, 分别是:白头鹎(Pycnonotus sinensis, 4.28%)、珠颈斑鸠(Spilopelia chinensis, 3.33%)、暗绿绣眼鸟(Zosterops japonicus, 3.29%)、乌鸫(Turdus merula, 3.19%)、栗背短脚鹎(Hemixos castanonotus, 2.72%)、鹊鸲(Copsychus saularis, 2.59%)、远东山雀(Parus minor, 2.46%)、叉尾太阳鸟(Aethopyga latouchii, 2.39%)、北红尾鸲(Phoenicurus auroreus, 2.22%)、红耳鹎(Pycnonotus jocosus, 1.99%)。
从季节上看, 春冬两季观测到的鸟类种类与数量比夏秋两季多;春、夏、秋、冬四个季节观测到的鸟类种类分别为185种(2578只)、77种(352只)、106种(561只)和194种(3005只);四季中鸟类数量占全年总数量的比例分别为39.69%、5.42%、8.64%和46.26%。
从重点保护和受胁迫鸟类来看, 公众观测到Ⅰ级重点保护野生鸟类1种, 为中华秋沙鸭(Mergus squamatus)(1只), Ⅱ级重点保护野生鸟类26种(共378只)。根据IUCN判定标准, 观测到6种鸟类受到胁迫, 其中胁迫等级为极危(CR)的1种, 为细纹苇莺(Acrocephalus sorghophilus)(2只);濒危(EN)1种, 为中华秋沙鸭;易危(VU)鸟类2种:白腰燕鸥(Sterna aleutica)(2只)和白喉林鹟(Rhinomyias brunneatus)(6只);近危(NT)2种:紫寿带(Terpsiphone atrocaudata)(9只)和白眉山鹧鸪(Arborophila gingica)(8只)。公众观测到的受胁迫鸟类及重点保护野生鸟类主要分布在晋安区外围地带、鼓楼区和仓山区沿闽江一侧(图 2)。

|
| 图 2 受胁迫鸟类和国家重点保护野生鸟类空间分布 Fig. 2 Spatial distribution of threatened birds and state key protected wild birds |
从鸟类居留型来看, 研究区留鸟种类和数量最多(图 3), 共116种4127只(63.53%)、夏候鸟种类和数量最少, 为20种255只(3.93%)、冬候鸟73种共1678只(25.83%)、旅鸟33种共436只(6.71%)。其中旅鸟主要鸟种为远东山雀, 共160只(36.70%)、家燕(Hirundo rustica), 共102只(23.39%)、矶鹬(Actitis hypoleucos), 共35只(8.03%), 其观测到的数量在秋季占比最大。冬候鸟主要鸟种为白头鹎, 共278只(16.57%)、北红尾鸲(144只, 8.58%)、绿背姬鹟(Ficedula elisae), 共126只(7.51%)、黄腰柳莺(Phylloscopus proregulus), 共99只(5.90%), 主要在夏冬两季观测到的数量较多。夏候鸟主要鸟种为池鹭(Ardeola bacchus), 共124只(48.63%)、紫啸鸫(Myophonus caeruleus), 共39只(15.29%), 灰卷尾(Dicrurus leucophaeus), 共16只(6.27%)、八声杜鹃(Cacomantis merulinus), 共14只(5.49%), 其被观测到的数量在夏季占比最大;留鸟大部分鸟种数量较为接近, 其观测数量在春季中占比较大, 主要鸟种为暗绿绣眼鸟(216只, 5.23%)、珠颈斑鸠(214只, 5.19%)、乌鸫(207只, 5.02%)。

|
| 图 3 不同居留型鸟类数量占比的四季变化 Fig. 3 Seasonal changes in the proportion of migratory bird species |
从全域范围看, 丰富度指数S介于1至222之间, H介于0—0.67之间, D介于0—0.99之间, 上述3个多样性指数的Moran′s I系数均接近0(-0.04至0.08)(图 4)。300 m和1000 m尺度下S均值分别为28.2和36.2, H均值分别为3.78和4.21, D均值分别为0.83和0.87。Mann-Whitney U检验表明两个尺度下3个多样性指数中的S和H指数存在一定差异(0.05<P<0.1), 总体上表现出在更大尺度下多样性指数的均值更高(图 5)。由图 4所示, S较低(值介于1—40之间)的格网数量占比较大(300 m: 79.52%;1000 m: 68.42%);S较高(值介于73—222之间)的格网数量占比较小(300 m: 9.46%;1000 m: 10.53%)。H在中高值区(值介于3.37—5.70)格网数量占比较大(300 m:61.45%;1000 m:68.42%), 主要分布于鼓楼区(32.53%)和晋安区(15.66%)。D较高(值介于0.81—0.99之间)的格网数量占比较大(300 m:77.11%;1000 m:82.46%)。

|
| 图 4 两个尺度下鸟类多样性指数空间分布 Fig. 4 Spatial distribution of bird species diversity indices across two scales 3个多样性指数按自然断点法分为5级:低值区Ⅰ、较低值区Ⅱ、中值区Ⅲ、较高值区Ⅳ和高值区Ⅴ; 柱状图展示了不同颜色分级下的格网数量 |

|
| 图 5 两个尺度下3个鸟类多样性指数的差异性 Fig. 5 Differences of bird species diversity indices at two scales |
如图 6所示, GLM模型结果表明在300m尺度上各景观因子对S、H和D指数影响均不显著(P>0.05);而在1000 m尺度上S、H和D指数受到不同组合景观因子的显著影响(P<0.05)。在1000 m尺度上, 丰富度指数S受GA(标准化回归系数=1.56)、RD(标准化回归系数=-1.82)和BV(标准化回归系数=1.80)显著影响, 其中RD具有最大的负效应, 而BV和GA具有正效应;GA(正效应)和HT(负效应)是显著影响H指的景观因子, 两者作用程度相当但方向相反;而D指数仅受GA的正向影响。3个多样性指数均受城市二维结构特征GA的正向影响(P<0.05);城市三维结构特征(HT和BV)亦会影响城市鸟类多样性(S和H指数)。此外, 在考虑多个景观因子的综合作用后, WA, RSEI, RA和H_SD在两个尺度上均不存在显著影响(P>0.05)。

|
| 图 6 基于广义线性的两个尺度下鸟类多样性指数与景观因子回归结果 Fig. 6 Regression results of bird species diversity indices and landscape factors using generalized linear models at two scales 图中的方点表示标准化回归系数的均值, 误差棒表示标准误差;实心方点表示对多样性指数产生显著影响的景观变量(P < 0.05);*表示GLM回归方程显著(P < 0.05);GA:总绿地面积占比(%);RA:居住区面积占比(%);RD:道路面积占比(%);WA:水域面积占比(%);HT:建筑高度(m);H_SD: 建筑高度变异程度(m);BV:建筑体积(万m3);RSEI:遥感生态指数 |
在300 m和1000 m尺度下, 3个鸟类多样性指数均存在明显空间异质性。结合图 1和图 2来看, 观鸟者观察到重点保护鸟类的地点主要集中在鼓楼区;作为研究区最早的城市中心, 鼓楼区形成以小块绿地、水系和低矮建筑混杂的镶嵌格局;而观鸟者观察到的受胁迫鸟类主要分布在晋安区城区外围地带, 晋安区作为早期主城区的城郊地带, 是主城区建筑群与东北部成片大面积森林的接壤区。从国家重点保护野生鸟类和受胁迫鸟类所在的网格区域内鸟类多样性和8个偏相关分析筛选出的景观因子的均值可以看出, 这些网格区域中鸟类多样性指数均高于全域平均水平(表 1)。网格内GA和RSEI均值高于全域均值, 建筑群较少且无城市主干道路(RD)的景观特征(表 1)。相较于重点保护野生鸟类, 受胁迫鸟类所在网格中与鸟类多样性指数正相关的因子(GA和RSEI)的均值比全域平均水平要高, 而与鸟类多样性指数负相关的因子(RD、RA、HT、H_SD、BV)的值均普遍较全域均值更低。这预示着受胁迫鸟类对栖息环境的自然度和人工构筑物的敏感性比重点保护鸟类很可能更高。研究主要是在单个年度观鸟数据基础上开展的基于网格单元对城市鸟类多样性与景观特征关系的探索, 这些国家重点保护野生鸟类和受胁迫鸟类网格空间内及其跨越网格空间的活动行为(如鸟类的繁殖行为、取食行为和社群行为)对结果的影响并未被考虑。
| 网格尺度/m Grid scale |
保护类型 Protection type |
S | H | D | GA | RD | RA | WA | HT | H_SD | BV | RSEI |
| 300 | 受胁迫鸟类 Threatened birds |
67.92 (81.3%) | 5.41 (26.5%) | 0.97 (9.8%) | 0.56 (60.8%) | 0 (-71.1%) | 0.15 (-40.5%) | 0.01 (-87.0%) | 12.09 (-58.3%) | 6.02 (-68.1%) | 0 (-96.3%) | 0.62 (17.8%) |
| 保护鸟类 Protected birds |
46.73 (24.8%) | 4.64 (8.5%) | 0.90 (1.7%) | 0.38 (9.0%) | 0 (8.3%) | 0.26 (4.1%) | 0.14 (59.0%) | 16.61 (-42.7%) | 6.92 (-63.3%) | 0 (-93.5%) | 0.55 (2.4%) | |
| 1000 | 受胁迫鸟类 Threatened birds |
82.89 (121.3%) | 5.61 (31.3%) | 0.97 (9.9%) | 0.46 (32.8%) | 0 (-63.1%) | 0.15 (-40.7%) | 0.01 (-85.1%) | 10.41 (-64.1%) | 5.75 (-69.6%) | 0 (-54.6%) | 0.63 (18.5%) |
| 保护鸟类 Protected birds |
57.23 (52.8%) | 5.20 (21.7%) | 0.96 (8.9%) | 0.38 (10.9%) | 0 (-7.0%) | 0.22 (-8.7%) | 0.11 (22.2%) | 27.09 (-6.6%) | 17.78 (-5.8%) | 0 (-4.3%) | 0.55 (3.0%) | |
| 标注下划线的景观因子为偏相关系数为正值的变量; 括号上方数值代表各景观因子在对应尺度网格下的平均值; 括号中的数值代表各景观因子偏离全域均值的程度, 即:(保护鸟种所在网格各因子实际值-全域所有网格均值)/全域所有网格均值×100%;S:丰富度指数Richness index;H:Shannon-Wiener多样性指数Shannon-Wiener diversity index;D:Simpson′s多样性指数Simpson′s diversity index; GA:总绿地面积占比(%);RD:道路面积占比(%);RA:居住区面积占比(%);WA:水域面积占比(%);HT:建筑高度(m);H_SD: 建筑高度变异程度(m);BV:建筑体积(万m3);RSEI:遥感生态指数 | ||||||||||||
本研究发现在两个分析尺度上能够检测到的影响鸟类多样性的景观因子具有明显尺度依赖性。当考虑多个景观因子的综合作用时, 在300 m尺度上并未检测到影响鸟类多样性的关键因子。随着网格尺度增至1000 m, 回归分析样本单元也从300 m尺度的91个减少至61个, 景观因子与鸟类多样性指数却呈现显著关联性。本研究发现城市绿地总面积占比GA是对3个鸟类多样性的指标均具有促进作用的重要景观变量, 这与前人研究结果一致[4, 7, 18]。本研究进一步发现除了表征城市土地覆盖的二维特征GA外, 城市三维结构亦产生关键作用。在城市主城区景观基质中, 更少的城市道路(RD), 更多的绿地覆盖(GA)、更大的建筑群体积(BV), 有利于增加城市鸟类丰富度(S);而在更多的绿地覆盖区, 较低的建筑高度(HT)亦能增加H指数。已有研究表明:过高的建筑高度会增加鸟类撞窗的碰撞率[32], 进而增加鸟类的生存风险。在偏相关回归分析中, 建筑体积BV和居住区面积占比RA两个主要表征构筑物三维和二维结构特征的因子与3个多样性指数均负相关, 但综合了其他景观因素的影响后, RA在GLM回归中变得不显著(P>0.05), BV却产生较大的正向作用;BV的正效应与RD的负效应接近。这意味着在假设其他景观变量固定的情况下, 网格空间中更大的建筑群体积有利于城市鸟类高多样性的维持。本研究推测这可能是与城市形态的紧凑度有关[33], 一些研究已指出大型绿地得到保护的更密集紧凑的城市形态有利于鸟类的保护和维持[34]。尽管本文选择的建筑体积BV并不是唯一决定城市紧凑性的因素, 但在一定程度上, 较大的城市建筑群体积意味着城市向垂直方向的扩展, 增加土地利用效率, 更多的建筑物被放置在相对较小的建设土地区域内, 减少了城市分散及由此引起的道路通勤强度的增加[33, 35], 为保留和增加城市绿地覆盖空间提供更大的可能性。此外, 本文尝试了用RSEI作为表征生态功能的替代指标, 虽然在偏相关分析中RSEI与3个鸟类多样性指数正相关(偏相关系数范围0.33—0.40)且在重要保护鸟类所在网格分析中RSEI均值均较高, 但是在综合其他结构因素的共同作用后RSEI对3个多样性指数的影响并不显著;这可能是由于RSEI作为集成了干度、湿地和地表温度和植被信息的指数[29], 其对鸟类多样性的作用仍来源于城市景观中复杂的自然与人工系统形成的高度异质结构特征。尽管城市三维特征对鸟类多样性维持和作用机理有待进一步验证, 在未来生态城市规划和建设中, 城市管理者和规划师不能仅考虑增加城市绿色空间, 更要关注城市三维结构形态对城市生物多样性保护和维持的积极作用[36—38]。
尽管目前城市景观研究数据分辨率日趋提高, 为了能与已被广泛使用的大尺度且低成本1000 m分辨率生态地理数据结合应用, 结合本文研究结果建议研究人员可选择1000 m或1000 m整倍数的更大分析网格尺度开展此类研究。
4.3 公民观鸟数据的不确定性本研究在1000 m尺度上从城市景观特征方面筛选的二维和三维景观因子, 基本包括了前人类似研究中常考虑的景观变量, 但它们对S、H和D多样性指数的解释率并不高(R2=0.17—0.27, 图 6)。已有研究表明在每个网格中参与观鸟的观鸟者越多, 对城市鸟类生物多样性的监测就越充分, 评估结果亦越准确[39]。本研究在潜在景观因子初筛时, 已试图通过偏相关分析控制观测数量的影响, 然而, 由于客观因素限制, 观鸟者个人主观因素(观鸟方法、公众是否接受过培训)[40]和地理空间上选择偏好[39]对公民观鸟数据的质量影响未能获得较好评估, 均给本研究带来了一定不确定性;这亦可能是本研究R2偏低的原因之一。不可否认, 公民观鸟数据的确可以增加数据采集的效率和规模, 特别是在大尺度时空范围研究中优势明显[41], 为城市地区生物多样性的保护提供了重要数据来源。未来, 鼓励更多的公众参与到有组织数据的观鸟活动的同时更要注重提升参与者的专业性, 改善依托公民科学数据开展城市鸟类多样性研究的质量, 降低这类数据应用中的不确定性[42]。
5 结论福州主城区公民观鸟数据中观鸟者观测到鸟类242种且以雀形目为优势种。在3个鸟类多样性指数(S、H和D)中, 在更大的1000 m分析网格尺度下得到的城市鸟类多样性指数(S和H)显著高于300 m尺度。两个尺度中仅在1000 m尺度上检测到景观特征对城市鸟类多样性指数的显著影响。在1000 m尺度下城市景观二维(GA和RD)和三维(BV和HT)结构特征对城市鸟类多样性产生关键作用;其中GA和BV起促进作用, RD和HT则产生抑制作用。道路更少, 绿地覆盖更高且更紧凑的城市景观很可能更有利于城市鸟类多样性的保护和维持。
| [1] |
Foley J A, DeFries R, Asner G P, Barford C, Bonan G, Carpenter S R, Chapin F S, Coe M T, Daily G C, Gibbs H K, Helkowski J H, Holloway T, Howard E A, Kucharik C J, Monfreda C, Patz J A, Prentice I C, Ramankutty N, Snyder P K. Global consequences of land use. Science, 2005, 309(5734): 570-574. DOI:10.1126/science.1111772 |
| [2] |
Callaghan C T, Bino G, Major R E, Martin J M, Lyons M B, Kingsford R T. Heterogeneous urban green areas are bird diversity hotspots: insights using continental-scale citizen science data. Landscape Ecology, 2019, 34(6): 1231-1246. DOI:10.1007/s10980-019-00851-6 |
| [3] |
Aronson M F, Lepczyk C A, Evans K L, Goddard M A, Lerman S B, MacIvor J S, Nilon C H, Vargo T. Biodiversity in the city: key challenges for urban green space management. Frontiers in Ecology and the Environment, 2017, 15(4): 189-196. DOI:10.1002/fee.1480 |
| [4] |
郭诗怡, 斋藤馨, 夏原由博, 曹洋. 城市建成环境对鸟类多样性的影响机制研究评述及展望. 中国园林, 2022, 38(2): 71-76. |
| [5] |
Cardinale B J, Duffy J E, Gonzalez A, Hooper D U, Perrings C, Venail P, Narwani A, Mace G M, Tilman D, Wardle D A, Kinzig A P, Daily G C, Loreau M, Grace J B, Larigauderie A, Srivastava D S, Naeem S. Biodiversity loss and its impact on humanity. Nature, 2012, 486(7401): 59-67. DOI:10.1038/nature11148 |
| [6] |
张征恺, 黄甘霖. 中国城市鸟类学研究进展. 生态学报, 2018, 38(10): 3357-3367. |
| [7] |
Rodrigues A G, Borges-Martins M, Zilio F. Bird diversity in an urban ecosystem: the role of local habitats in understanding the effects of urbanization. Iheringia Série Zoologia, 2018, 108. |
| [8] |
陈水华, 丁平, 郑光美, 诸葛阳. 城市鸟类群落生态学研究展望. 动物学研究, 2000, 21(2): 165-169. |
| [9] |
Tilghman N G. Characteristics of urban woodlands affecting breeding bird diversity and abundance. Landscape and Urban Planning, 1987, 14: 481-495. DOI:10.1016/0169-2046(87)90061-2 |
| [10] |
Melles S, Glenn S M, Martin K. Urban bird diversity and landscape complexity: species-environment associations along a multiscale habitat gradient. Conservation Ecology, 2003, 7: art5. |
| [11] |
Reis E, López-Iborra G M, Pinheiro R T. Changes in bird species richness through different levels of urbanization: implications for biodiversity conservation and garden design in Central Brazil. Landscape and Urban Planning, 2012, 107(1): 31-42. DOI:10.1016/j.landurbplan.2012.04.009 |
| [12] |
Hughes A C, Orr M C, Lei F M, Yang Q M, Qiao H J. Understanding drivers of global urban bird diversity. Global Environmental Change, 2022, 76: 102588. DOI:10.1016/j.gloenvcha.2022.102588 |
| [13] |
王茜, 刘智存, 刘天宇, 万冬梅, 蒋一婷. 城市鸟类多样性研究中的多维度探索研究进展. 生态学杂志, 2022, 41(10): 2058-2063. |
| [14] |
许龙, 张正旺, 丁长青. 样线法在鸟类数量调查中的运用. 生态学杂志, 2003, 22(5): 127-130. |
| [15] |
杨刚, 许洁, 王勇, 丁由中, 袁晓, 裴恩乐, 马波, 王小明, 王正寰. 城市公园植被特征对陆生鸟类集团的影响. 生态学报, 2015, 35(14): 4824-4835. |
| [16] |
Coxen C L, Frey J K, Carleton S A, Collins D P. Species distribution models for a migratory bird based on citizen science and satellite tracking data. Global Ecology and Conservation, 2017, 11: 298-311. DOI:10.1016/j.gecco.2017.08.001 |
| [17] |
Feng M L E, Che-Castaldo J. Comparing the reliability of relative bird abundance indices from standardized surveys and community science data at finer resolutions. PLoS One, 2021, 16(9): e0257226. DOI:10.1371/journal.pone.0257226 |
| [18] |
Reif J, Vermouzek Z, Voříšek P, Romportl D, Morelli F. Birds' ecological characteristics differ among habitats: an analysis based on national citizen science data. Community Ecology, 2022, 23(2): 173-186. DOI:10.1007/s42974-022-00089-4 |
| [19] |
Echeverry-Galvis M A, Lozano Ramírez P, Amaya-Espinel J D. Long-term Christmas Bird Counts describe neotropical urban bird diversity. PLoS One, 2023, 18(2): e0272754. DOI:10.1371/journal.pone.0272754 |
| [20] |
斯幸峰, 丁平. 欧美陆地鸟类监测的历史、现状与我国的对策. 生物多样性, 2011, 19(3): 303-310. |
| [21] |
Li X Y, Liang L, Gong P, Liu Y, Liang F F. Bird watching in China reveals bird distribution changes. Chinese Science Bulletin, 2013, 58(6): 649-656. DOI:10.1007/s11434-012-5458-7 |
| [22] |
Kati V I, Sekercioglu C H. Diversity, ecological structure, and conservation of the landbird community of Dadia reserve, Greece. Diversity and Distributions, 2006, 12(5): 620-629. DOI:10.1111/j.1366-9516.2006.00288.x |
| [23] |
Silvertown J. A new dawn for citizen science. Trends in Ecology & Evolution, 2009, 24(9): 467-471. |
| [24] |
La Sorte F A, Lepczyk C, Burnett J, Hurlbert A, Tingley M, Zuckerberg B. Opportunities and challenges for big data ornithology. The Condor: Ornithological Applica-tions, 2018, 120(2): 414-426. DOI:10.1650/CONDOR-17-206.1 |
| [25] |
Stuber E F, Robinson O J, Bjerre E R, Otto M C, Millsap B A, Zimmerman G S, Brasher M G, Ringelman K M, Fournier A M V, Yetter A, Isola J E, Ruiz-Gutierrez V. The potential of semi-structured citizen science data as a supplement for conservation decision-making: Validating the performance of eBird against targeted avian monitoring efforts. Biological Conservation, 2022, 270: 109556. DOI:10.1016/j.biocon.2022.109556 |
| [26] |
Jäckel D, Mortega K G, Darwin S, Brockmeyer U, Sturm U, Lasseck M, Moczek N, Lehmann G U C, Voigt-Heucke S L. Community engagement and data quality: best practices and lessons learned from a citizen science project on birdsong. Journal of Ornithology, 2023, 164(1): 233-244. |
| [27] |
刘萌萌, 张曼玉, 韩茜, 武大伟, 王思路, 鲁长虎. 公众观鸟和传统样线法调查应用于鸟类多样性监测的比较: 以南京老山为例. 生态与农村环境学报, 2022, 38(9): 1-16. |
| [28] |
张敏, 田春坡, 车先丽, 赵岩岩, 陈什旺, 周霞, 邹发生. 广东省鸟类新记录及其与自然和社会经济因素的关联性. 生物多样性, 2022, 30(5): 37-47. |
| [29] |
徐涵秋. 城市遥感生态指数的创建及其应用. 生态学报, 2013, 33(24): 7853-7862. |
| [30] |
徐涵秋, 邓文慧. MRSEI指数的合理性分析及其与RSEI指数的区别. 遥感技术与应用, 2022, 37(1): 1-7. |
| [31] |
Ding Y, Zang R G. Determinants of aboveground biomass in forests across three climatic zones in China. Forest Ecology and Management, 2021, 482: 118805. |
| [32] |
Loss S R, Will T, Loss S S, Marra P P. Bird-building collisions in the United States: estimates of annual mortality and species vulnerability. The Condor, 2014, 116(1): 8-23. |
| [33] |
Artmann M, Inostroza L, Fan P L. Urban sprawl, compact urban development and green cities. How much do we know, how much do we agree?. Ecological Indicators, 2019, 96: 3-9. |
| [34] |
Concepción E D, Obrist M K, Moretti M, Altermatt F, Baur B, Nobis M P. Impacts of urban sprawl on species richness of plants, butterflies, gastropods and birds: not only built-up area matters. Urban Ecosystems, 2016, 19(1): 225-242. |
| [35] |
Amaya-Espinel J D, Hostetler M, Henríquez C, Bonacic C. The influence of building density on Neotropical bird communities found in small urban parks. Landscape and Urban Planning, 2019, 190: 103578. |
| [36] |
Artmann M, Kohler M, Meinel G, Gan J, Ioja I C. How smart growth and green infrastructure can mutually support each other—a conceptual framework for compact and green cities. Ecological Indicators, 2019, 96: 10-22. |
| [37] |
Liu Z H, Huang Q D, Tang G P. Identification of urban flight corridors for migratory birds in the coastal regions of Shenzhen city based on three-dimensional landscapes. Landscape Ecology, 2021, 36(7): 2043-2057. |
| [38] |
Casalegno S, Anderson K, Cox D T C, Hancock S, Gaston K J. Ecological connectivity in the three-dimensional urban green volume using waveform airborne lidar. Scientific Reports, 2017, 7: 45571. |
| [39] |
Duan H, Xia S, Yu X, Liu Y, Teng J, Dou Y. Using citizen science data to inform the relative sensitivity of waterbirds to natural versus human‐dominated landscapes in China. Ecology and Evolution, 2020, 10(14): 7233-7241. |
| [40] |
Callaghan C, Watson J, Lyons M, Cornwell W, Fuller R. Conservation birding: a quantitative conceptual framework for prioritizing citizen science observations. Biological Conservation, 2021, 253: 108912. |
| [41] |
Randler C. Users of a citizen science platform for bird data collection differ from other birdwatchers in knowledge and degree of specialization. Global Ecology and Conservation, 2021, 27: e01580. |
| [42] |
Boakes E H, McGowan P J K, Fuller R A, Ding C Q, Clark N E, O'Connor K, Mace G M. Distorted views of biodiversity: spatial and temporal bias in species occurrence data. PLoS Biology, 2010, 8(6): e1000385. |