 2023, Vol. 43
2023, Vol. 43文章信息
- 王珍, 常顺利, 王冠正, 周小东, 张毓涛, 孙雪娇, 李翔
- WANG Zhen, CHANG Shunli, WANG Guanzheng, ZHOU Xiaodong, ZHANG Yutao, SUN Xuejiao, LI Xiang
- 天山中段北坡森林土壤有机碳库稳定性组分沿海拔的分异规律
- Altitudinal distribution of the forest soil organic carbon fractions on the northern slope of the middle Tianshan Mountains
- 生态学报. 2023, 43(18): 7390-7402
- Acta Ecologica Sinica. 2023, 43(18): 7390-7402
- http://dx.doi.org/10.20103/j.stxb.202210112893
-
文章历史
- 收稿日期: 2022-10-11
- 网络出版日期: 2023-05-08





2. 新疆天山森林生态系统国家定位观测研究站, 乌鲁木齐 830063;
3. 新疆林科院森林生态研究所, 乌鲁木齐 830063
2. Institute of Forest Ecology, Xinjiang Academy of Forestry, Urumqi 830063, China;
3. Tianshan Forest Ecosystem National Station, Urumqi 830063, China
土壤是陆地生态系统中最大的碳库[1], 森林土壤占全球土壤有机碳库的70%—73%[2], 森林土壤碳库微小变化可导致大气CO2浓度的显著变化, 进而影响全球碳循环与气候变暖[1]。土壤有机碳库由不同化学结构的碳组分构成, 各组分具有不同的稳定性和周转时间, 因此可将土壤有机碳库分为活性碳库(Ca)、缓效性碳库(Cs)和惰性碳库(Cp)[3]。其中, 活性碳对周围环境变化的响应较为敏感, 能及时反映土壤碳库的变化[4];缓效性碳和惰性碳一同稳定地储存在土壤中, 因其非常稳定且周转时间长, 是衡量土壤碳库积累与碳稳定性的主要因子[5]。活性碳中的微生物量碳最为活跃, 它直接影响土壤中的碳周转过程。全面系统地研究土壤活性碳、微生物量碳、缓效性碳和惰性碳等土壤有机碳库稳定性组分的分配比例, 能够更加深入地刻画土壤碳库对气候变化响应的机制和过程。
土壤有机碳库的稳定性组分可能会对气候变化的响应产生差异。随着全球气候的变暖, 在温度、水分、微生物的作用下, 储存在土壤中的碳可加快自身的周转速率, 使一部分不稳定的碳释放到大气中, 从而使全球气候加速变暖, 可能对气候变化产生正反馈[6]。相反那些较为稳定的碳组分一般被认为是碳汇的标志, 它可将碳固定在土壤中, 从而减缓土壤向大气中释放二氧化碳, 对气候变化产生负反馈作用[7]。高纬度高海拔区域土壤碳稳定性组分的配比关系对气候变暖呈正反馈还是负反馈效应, 目前尚无一致答案[8]。
海拔变化影响水热条件的再分配, 从而影响植被分布、土壤理化性质、土壤微生物构成[9—11], 因此也会影响森林土壤有机碳库以及碳组分的构成。有研究表明, 海拔升高显著增加长白山[12](针阔混交林、暗针云杉冷杉林、暗针叶云杉林、埃尔曼桦树林)和武夷山[13](亚热带常绿阔叶林、针叶林、亚高山矮林、高山草甸)土壤的活性碳含量, 显著增大鼎湖山森林[14](沟谷雨林、低地常绿阔叶林、山地常绿阔叶林)土壤惰性碳库占总碳库的比例。海拔变化对神农架[15](亚高山灌丛、亚高山暗针叶林、针阔混交林、落叶阔叶林、常绿阔叶林)表层土壤微生物量碳也具有显著的分异规律。综合来看, 以往对森林有机碳库对海拔梯度响应的研究主要集中在植被类型间的比较, 难以排除植被类型的干扰而只考虑水热配比变化条件下土壤有机碳库稳定性组分沿海拔的分异规律。
天山森林是以雪岭云杉(Picea Schrenkiana)为单优树种的山地针叶林, 在天山北坡中山带(海拔约1760—2800 m)呈垂直落差超过1000 m的带状斑块分布, 可作为排除植被类型差异影响而研究水热连续变化对土壤有机碳稳定性组分分异规律的理想实验地。因此, 本文提出科学问题为:(1)雪岭云杉森林土壤碳库稳定性组分是否随海拔变化呈现分异规律?(2)该分异规律对气候变暖如何响应?
1 材料与方法 1.1 研究区概况本研究主要依托中国森林生态系统研究网络(CFERN)新疆天山森林生态系统国家定位观测研究站(E87°07′—87°28′, N43°14′—43°26′)开展工作, 该站位于新疆维吾尔自治区乌鲁木齐市乌鲁木齐县板房沟林场, 属温带大陆性气候, 冬季最大积雪深度为65 cm, 年总辐射量达5.85×105 Jcm2/a, 林下土壤类型为灰褐色森林土。天山北坡受到中纬度西风带的控制, 来自大西洋和北冰洋的湿润气流从西方或西北方吹入, 受天山阻隔, 水热条件随地形和海拔重新分配, 垂直变化明显[16](表 1)。研究区是以雪岭云杉(Picea Schrenkiana)纯林为主的温帯针叶林, 主要灌木有小檗(Berberis heteropoda及Berberis nummularia)、栒子(Cotoneaster melanocarpus)、蔷薇(Rosa spinosissima)、忍冬(Lonicera hispida)、方枝柏(Juniperus pseudosabina)等, 林下草本主要由老鹘草(Geranium rotundifolium)、天山羽衣草(Alchemilla tianschanica)、羊角芹(Aegopodium Podagraria)等构成[17]。
| 海拔梯度 Elevation gradient /m |
平均树高 Mean height of trees/m |
平均胸径 Mean diameter at breast height/cm |
郁闭度 Crown density |
年均温 Annual average temperature/℃ |
年降水量 Annual average precipitation/mm |
| 1800—2000 | 2.12 | 7.56 | 0.50 | 14.29 | 437.44 |
| 2000—2200 | 9.96 | 18.48 | 0.20—0.90 | 13.09 | 443.99 |
| 2200—2400 | 12.31 | 16.70 | 0.20—0.80 | 11.89 | 453.26 |
| 2400—2600 | 12.33 | 25.41 | 0.20—0.70 | 10.69 | 460.04 |
| 2600—2800 | 6.36 | 9.93 | 0.20—0.70 | 9.49 | 477.65 |
2021年7月, 选取天山中段北坡山地森林(海拔1760—2800 m)为研究对象, 以200 m为一个海拔梯度, 分别在海拔1800—2000 m, 2000—2200 m, 2200—2400 m, 2400—2600 m, 2600—2800m进行采样, 且在同一海拔内取3个20 m×20 m样地, 每个样地至少相隔100 m以上, 同时在海拔1800 m处的裸地上设置3个20 m×20 m样地作为对照, 共设置18个样地。
在每个样地内按照“S”型的方法设置了3个采样点, 按照0—10 cm, 10—20 cm, 20—40 cm, 40—60 cm, 60—100 cm划分土层, 采用环刀法分层取样, 用以计算土壤容重;同时按上述土层采集各土层土样300g, 共计采集270个土样。
将土壤样品带回实验室进行处理, 去除其中的植物根系、石块等杂物, 将土样分成两等份:一份放置于-4℃冷藏, 用于测定微生物量碳含量;另一份放置阴凉通风处, 待风干过2 mm筛, 测其总有机碳, 活性碳, 缓效性碳以及惰性碳含量。
1.3 测试指标方法(1) 土壤总有机碳(Soil Organic Carbon, SOC):采用重铬酸钾氧化-外加热法。
(2) 土壤活性碳(Active carbon pool, Ca)、缓效性碳(Slow carbon pool, Cs)、惰性碳(Passive carbon pool, Cp)采用硫酸两步水解-重铬酸钾氧化滴定法[18]测定, 步骤如下:称取0.5g 2 mm风干土样加入离心管, 加20 mL2.5 mol/L H2SO4, 并水解30 min(105 ℃), 离心, 倾倒提取上清液于待测瓶。残渣加20 mL去离子水清洗, 后离心提取上清液再次转入待测瓶中, 供Ca使用;土壤残渣加去离子水清洗数次并在60 ℃下烘干, 加2 mL的13 mol/L H2SO4, 室温振荡过夜, 后加去离子水将H2SO4稀释成1 mol/L, 105 ℃水解3 h, 后续操作同前, 该上清液用于Cs测定;剩余残渣用去离子水清洗数次后在60 ℃下烘干, 研磨后使用重铬酸钾-浓硫酸-外加热法测定Cp的含量;Ca和Cs所提取的上清液分别使用总有机碳分析仪测定。
(3) 微生物量碳(Microbial Biomass Carbon, MBC):采用氯仿熏蒸-K2SO4浸提法[19]。即新鲜土样经氯仿熏蒸(24 h)后, 土壤微生物死亡细胞发生裂解, 释放出微生物量碳, 用一定体积的0.5 mol/L K2SO4溶液提取土壤, 借用有机碳自动分析仪测定微生物量碳含量。计算公式为:MBC=ΔEc/kc, 式中:ΔEc为熏蒸土壤有机碳与未熏蒸土壤有机碳的差值, kc(0.45)为微生物碳的浸提系数。
1.4 数据处理与分析 1.4.1 指标计算方法(1) 微生物熵(Microbe quotient, qMB)是土壤微生物量碳占土壤总有机碳的百分比, 即
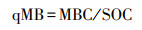
|
(1) |
式中, qMB为微生物熵, MBC为微生物量碳, SOC为土壤总有机碳
(2) 土壤碳库指数(CPI)的计算以裸地土壤碳库为对照, 参照Blair等[20]方法。
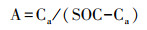
|
(2) |
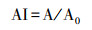
|
(3) |
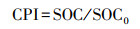
|
(4) |
式中, A为碳库活度, Ca为土壤活性碳库, SOC为土壤总有机碳库, SOC-Ca为土壤非活性碳库, AI为碳库活度指数, A0为参考土壤碳库活度, CPI为碳库指数, SOC0为参考土壤总有机碳库。
(3) 土壤各层的土壤碳密度(SOCD, kg/cm2)计算公式[21]为:
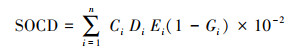
|
(5) |
式中, Ci为第i层土的有机碳含量(g/kg);Di为第i层土壤容重(g/cm3);Ei为第i层土壤厚度(cm);Gi为第i层>2 mm砾石含量(体积%);n为参与计算的土壤层次总数。
1.4.2 气象数据来源气象数据时间序列为1997—2012年, 由于研究区各海拔气象数据获取困难, 选择使用SWAT模型在乌鲁木齐河流域建模输出结果[22], 通过插值法来确定研究区域每个海拔的年降水数据, 温度数据来自小渠子气象站(海拔2160 m), 通过“海拔每升高100 m, 气温下降0.6 ℃”计算每个海拔的年温度数据。
1.4.3 数据处理所测数据使用Excel软件、SPSS 20.0软件进行统计分析, 用Origin2018、R 4.0.2作图;所有数据在方差分析前进行正态分布检验, 将不合要求的数据进行转换, 之后需要进行方差齐性检验和差异显著性检验。选择单因素方差(one-way ANOVA)中的Duncan法探讨相同海拔不同土层和相同土层不同海拔间土壤各碳组分含量、各碳库分配比、CPI的差异显著性(P < 0.05);用双因素方差(two-way ANOVA) 中的Duncan法探讨海拔和土层下土壤碳组分、各碳库分配比、CPI的差异显著性(P < 0.05)。利用R中的相关性矩阵图探讨海拔梯度上水热条件对土壤不同碳库的影响。
2 结果与分析 2.1 土壤有机碳的海拔分异规律天山中段北坡森林土壤各碳组分在海拔上的分异规律如图 1所示, 双因素方差分析表明海拔对SOC、Ca、Cs、Cp均具有极显著影响(P < 0.01), 土层对所有的土壤碳组分均有极显著影响(P < 0.001), 而海拔与土层的交互作用仅对Cs有显著影响(P < 0.01)。

|
| 图 1 海拔梯度上天山中段北坡森林土壤总有机碳、活性碳、缓效性碳、惰性碳的变化(平均值±标准误) Fig. 1 Dynamics of soil organic carbon (SOC)、active carbon (Ca)、slow carbon (Cs)、passive carbon (Cp) in forest soils on the northern slope of the middle Tianshan mountains along the elevation gradients (mean ± SE) EE:海拔, Elevation effect;HE:土层, Horizon effect;EE×HE:海拔和土层的交互作用, Interaction effect of elevation and horizon;不同大写字母表示不同海拔相同土层间差异显著(P < 0.05), 不同小写字母表示相同海拔不同土层间差异显著(P < 0.05);ns, 表示无显著性影响;*, **, ***分别代表P < 0.05, P < 0.01, P < 0.001 |
SOC含量处于17.12 —101.97 g/kg之间。从土层垂直剖面上来看, 除海拔2600—2800 m中各土层差异不显著外(P < 0.05), 其余海拔均随土层深度的增加显著减少(P < 0.05), 其中0—10 cm土层的SOC含量显著高于20—40, 40—60, 60—100 cm土层(P < 0.05)。在不同海拔相同土层中, SOC含量仅在0—10 cm土层显著增加后降低(P < 0.05), 其它土层均未表现出显著变化(P < 0.05), 而随着海拔升高, SOC含量整体呈现逐渐增加, 至2400—2600 m处达到最大值后又逐渐降低的趋势。
Ca含量处于2.90—17.62 g/kg之间, 从土层变化情况来看, 仅在海拔1800—2000 m、2200—2400 m的各土层有显著性差异(P < 0.05)。随着海拔升高, Ca含量大体呈现先增加至2400—2600 m处达到最大值后又减少的趋势。从不同海拔相同土层看, 除20—40 cm土层有显著性变化(P < 0.05), 其余土层在各海拔间均未表现出显著差异(P < 0.05)。
Cs含量处于3.98—60.20 g/kg之间, 除在海拔2600—2800 m的各土层中差异不显著外(P < 0.05), 其余海拔均随土层深度的增加显著减少(P < 0.05)。随着海拔的升高, Cs含量整体呈逐渐增加, 至2400—2600 m达到最大值后降低的趋势。在不同海拔相同土层中, 仅在0—10 cm土层显著增加后降低(P < 0.05), 其它土层未表现出显著变化(P < 0.05)。
Cp含量处于7.69—25.42 g/kg之间, 仅在海拔2000—2200 m与2200—2400 m中的土层间有显著差异(P < 0.05), 其余海拔均差异不显著(P < 0.05)。随着海拔升高, Cp含量整体呈逐渐增加, 至2000—2200 m处达到最大值后又逐渐降低的趋势。在不同海拔相同土层中, 除0—10 cm土层显著增加后降低外(P < 0.05), 其它土层均无显著变化(P < 0.05)。
2.2 土壤活性碳库分配比的海拔分异规律天山中段北坡森林土壤三种碳库在海拔梯度上的分配情况如图 2所示, A层中Cs、Cp为主要碳库, 和低、各中海拔相比, 森林上限Ca的分配比显著升高(P < 0.05), 含量分别为22.66%、16.44%、19.13%、17.19%、29.38%, Cs分配比显著减少, Cp先减少后显著升高(P < 0.05)。在B、C、D、E层中, 随着土层的加深, Ca、Cp是土壤中主要的碳库, 尤其在E层中两者的分配比分别为18.63%—30.34%, 40.71%—61.48%;且在B、C、D、E层中, Ca在高海拔处分配比显著升高, Cs无显著变化, Cp分配比显著减少(P < 0.05)。各海拔不同土层之间, 随着土层的加深, Ca分配比无显著变化, Cs显著减少(P < 0.05), Cp逐渐增加。双因素方差分析表明(表 2), 海拔对Ca、Cp有极显著影响(P < 0.01), 土层对Cs、Cp有极显著影响(P < 0.001), 海拔与土层的交互作用对Cs、Cp有显著影响(P < 0.05)。

|
| 图 2 海拔梯度上天山中段北坡森林土壤碳库三种稳定性组分的分配比变化(平均值±标准误) Fig. 2 Dynanmics in the distribution ratio of three stable components of the forest soil carbon pool on the northern slopes of the middle part of the Tian Shan along the elevation gradient (mean ± SE) Ca: 活性碳库(active carbon pool);Cs:缓效性碳库(slow carbon pool);Cp:惰性碳库(passive carbon pool);Horizon A:土层A(0—10cm);Horizon B:土层B(10—20cm);Horizon C:土层C(20—40cm);Horizon D:土层D(40—60cm);Horizon E: 土层E(60—100cm);不同大写字母表示不同海拔相同土层间差异显著(P < 0.05), 不同小写字母表示相同海拔不同土层间差异显著(P < 0.05) |
| 活性碳库 Active carbon pool Ca |
缓效性碳库 Slow carbon pool Cs |
惰性碳库 Passive carbon pool Cp |
||
| 双因素方差分析 Two-way ANOVA |
海拔 | 0.001** | 0.388 | 0.000*** |
| 土层 | 0.645 | 0.000*** | 0.000*** | |
| 海拔×土层 | 0.638 | 0.018* | 0.027* | |
| 海拔:Elevation effect;土层:Horizon effect;海拔与土层的交互作用:Interaction effect of elevation and horizon;表中数值为双因素方差的P值;*, **, ***分别代表P < 0.05, P < 0.01, P < 0.001 | ||||
天山中段北坡森林土壤微生物量碳、微生物熵在海拔上的分异规律如图 3所示, 海拔对MBC、qMB有极显著影响(P < 0.01), 土层仅对MBC有极显著影响(P < 0.001), 而海拔与土层的交互作用对MBC、qMB均无显著影响。

|
| 图 3 海拔梯度上土壤微生物量碳及微生物熵的变化(平均值±标准误) Fig. 3 Dynamics of microbial biomass carbon (MBC) and microbe quotient (qMB) along the elevation gradients (mean ± SE) EE:海拔, Elevation effect;HE:土层, Horizon effect;EE×HE:海拔和土层的交互作用, Interaction effect of elevation and horizon;不同大写字母表示不同海拔相同土层间差异显著(P < 0.05), 不同小写字母表示相同海拔不同土层间差异显著(P < 0.05);ns, 表示无显著性影响;*, **, ***分别代表P < 0.05, P < 0.01, P < 0.001 |
MBC的范围在96.26—668.83 mg/g之间。各海拔的MBC含量均随土层深度的加深不断减少;在海拔1800—2000 m, 2200—2400 m, 2400—2600 m的各土层间有显著性差异(P < 0.05)。随着海拔升高, MBC大体呈现出双峰曲线, 最大最小值分别出现在海拔2600—2800 m和1800—2000 m。在不同海拔相同土层中, 所有土层MBC含量在海拔2400—2600 m减少。
qMB处于0.59%—1.83%之间, 各海拔不同土层间均无显著性差异(P < 0.05)。随海拔梯度升高, qMB大体呈现出双峰曲线, 且最大最小值分别出现在海拔2600—2800 m和2400—2600 m。不同海拔相同土层中, 仅0—10 cm和10—20 cm土层有显著降低后增加(P < 0.05)。
2.4 土壤碳库质量的海拔分异规律天山中段北坡森林土壤碳密度及碳库质量在海拔梯度上的变化如表 3所示, 双因素方差分析表明, 海拔对A、AI、CPI、SOCD均有显著影响(P < 0.05), 土层对AI、SOCD有极显著影响(P < 0.01), 而海拔与土层的交互作用对A、AI、CPI、SOCD均无显著影响。
| 指标 Indexes |
海拔梯度 Elevation gradient/m |
土层深度Soil depth/cm | 全剖面 Whole profile |
Two-way ANOVA | ||||||
| 0—10 | 10—20 | 20—40 | 40—60 | 60—100 | 海拔 Elevation effect(EE) |
土层 Horizon effect(HE) |
EE×HE | |||
| 碳库活度 Carbon pool activity(A) |
裸地 | 0.38±0.03ABa | 0.45±0.11Aa | 0.38±0.04Aa | 0.27±0.06Aa | 0.27±0.04Aa | 0.34±0.03 | |||
| 1800—2000 | 0.29±0.00BCa | 0.24±0.03Ba | 0.21±0.03Ba | 0.30±0.09Aa | 0.36±0.05Aa | 0.30±0.04 | ||||
| 2000—2200 | 0.20±0.05Ca | 0.20±0.05Ba | 0.23±0.06Ba | 0.23±0.08Aa | 0.26±0.07Aa | 0.24±0.07 | ** | ns | ns | |
| 2200—2400 | 0.24±0.03Ca | 0.20±0.02Ba | 0.21±0.06Ba | 0.20±0.04Aa | 0.24±0.07Aa | 0.22±0.04 | ||||
| 2400—2600 | 0.21±0.02Ca | 0.43±0.05Aa | 0.39±0.02Aa | 0.37±0.14Aa | 0.45±0.09Aa | 0.39±0.07 | ||||
| 2600—2800 | 0.42±0.04Aa | 0.35±0.03ABa | 0.34±0.04ABa | 0.28±0.09Aa | 0.33±0.09Aa | 0.34±0.05 | ||||
| 碳库活度指数 Carbon pool Activity index(AI) |
1800—2000 | 0.79±0.07ABbc | 0.58±0.16Ac | 0.55±0.06Ac | 1.14±0.23Aab | 1.33±0.04Aa | 1.01±0.04 | |||
| 2000—2200 | 0.53±0.12Ba | 0.50±0.21Aa | 0.58±0.12Aa | 0.80±0.14Aa | 0.95±0.24Aa | 0.76±0.17 | ||||
| 2200—2400 | 0.63±0.07Ba | 0.49±0.10Aa | 0.59±0.21Aa | 0.95±0.44Aa | 0.97±0.34Aa | 0.81±0.25 | * | ** | ns | |
| 2400—2600 | 0.55±0.02Bb | 1.10±0.30Aab | 1.04±0.15Aab | 1.42±0.40Aab | 1.71±0.37Aa | 1.34±0.25 | ||||
| 2600—2800 | 1.14±0.22Aa | 0.84±0.19Aa | 0.92±0.14Aa | 1.45±0.89Aa | 1.38±0.57Aa | 1.23±0.40 | ||||
| 碳库指数 Carbon pool index(CPI) |
1800—2000 | 1.18±0.09BCa | 1.25±0.27Aa | 1.21±0.27Aa | 1.12±0.35Aa | 1.06±0.37Aa | 1.13±0.30 | |||
| 2000—2200 | 1.51±0.18ABa | 1.92±0.63Aa | 1.10±0.23Aa | 0.94±0.18Aa | 0.99±0.08Aa | 1.15±0.19 | ||||
| 2200—2400 | 1.54±0.11ABa | 1.46±0.24Aa | 0.99±0.29Aa | 0.89±0.21Aa | 0.82±0.26Aa | 1.01±0.21 | ** | ns | ns | |
| 2400—2600 | 1.97±0.33Aa | 2.03±0.73Aa | 2.28±0.90Aa | 1.82±0.85Aa | 1.38±0.53Aa | 1.77±0.64 | ||||
| 2600—2800 | 0.72±0.16Ca | 0.95±0.16Aa | 1.18±0.26Aa | 0.91±0.09Aa | 0.90±0.13Aa | 0.94±0.13 | ||||
| 土壤有机碳密度 Soil organic carbon density (SOCD, kg/cm2) |
裸地 | 4.22±0.63ABb | 2.99±0.52Ab | 5.34±1.17ABab | 4.60±1.09Ab | 8.61±1.71Aa | 25.77±8.73 | |||
| 1800—2000 | 4.86±0.55Aab | 3.51±0.14Ab | 5.56±0.21ABab | 4.48±0.39Aab | 7.02±1.56Aa | 25.43±2.44 | ||||
| 2000—2200 | 4.32±0.44ABb | 3.51±0.49Ab | 4.34±0.70ABb | 3.73±0.56Ab | 7.42±1.24Aa | 23.33±5.73 | ** | *** | ns | |
| 2200—2400 | 4.47±0.58ABa | 3.21±0.36Aa | 3.94±0.54Ba | 4.13±0.70Aa | 5.46±1.37Aa | 21.21±5.91 | ||||
| 2400—2600 | 5.31±0.49Aa | 3.48±0.75Aa | 8.35±2.26Aa | 7.25±2.94Aa | 10.57±3.56Aa | 34.95±15.18 | ||||
| 2600—2800 | 2.96±0.56Bab | 2.48±0.55Ab | 5.04±1.60ABab | 4.76±1.06Aab | 6.42±1.15Aa | 21.66±8.15 | ||||
| EE×HE:海拔和土层的交互作用, Interaction effect of elevation and horizon;表中数据为平均值±标准误;不同大写字母表示不同海拔相同土层间差异显著(P < 0.05), 不同小写字母表示相同海拔不同土层间差异显著(P < 0.05);*, **, ***分别代表P < 0.05, P < 0.01, P < 0.001 | ||||||||||
以裸地作为对照, 五个海拔的A、AI、CPI、SOCD值分别在0.20—0.45, 0.49—1.71, 0.72—2.28, 2.48—10.57之间, 从全剖面来看, 海拔2400—2600 m A、AI、CPI、SOCD值均最大。在不同海拔相同土层中, 随着海拔的升高, 0—10 cm土层的A值显著增大, 10—40 cm土层的A值在海拔2400—2600 m显著增大, AI值仅在0—10 cm土层显著增大(P < 0.05), CPI、SOCD值在海拔2400—2600 m达到最大。在相同海拔不同土层中, 随着土层的加深, A、AI、SOCD值均逐渐增大, 而CPI值逐渐减小。
3 讨论 3.1 海拔对土壤碳库及其稳定性组分的影响本研究区是天山雪岭云杉为单优树种的温带针叶林, 可避免受到其它植被的干扰, 因此土壤总有机碳库的变化主要与该区域的气候条件, 云杉的分布、长势相关。
本研究中, 随着海拔的升高, SOC与Ca含量均表现为先增后减少的趋势, 这与前人研究结果中随着海拔的升高, SOC表现为双峰特征[23], 土壤Ca未表现出明显的分异规律[7]不一致, 原因可能为本研究区随着海拔的升高水热梯度变化较为明显, 影响了雪岭云杉的生长及其凋落物厚度, 低海拔温度高降雨少, 云杉长势较好, 微生物较为活跃, 有机质分解强, 森林上限低温降雨多, 云杉少, 凋落物少, 而有机碳、活性碳主要来源于根系分泌物与凋落物[24], 因此SOC和Ca含量在森林上限处有所下降。
在高海拔地区, 土壤Cp含量降低, 这与吴玥等在戴云山黄山松林[9]的研究结果一致, 即土壤Cs、Cp随海拔上升呈先增加后减少的趋势, 导致此结果的原因可能是由于随海拔升高, SOC含量与气温逐步降低, 高海拔区域微生物活性增强, 从而加快了Cp的损失。
在天山中段北坡海拔1800—2800 m区间, 随海拔升高, 湿度增加, 雪岭云杉长势逐渐转好, 地表覆盖大量的枯枝落叶, 不利于有机质的分解, 因而SOC、Ca、Cs逐渐累积并在海拔2400—2600 m达到最大值。其次Ca含量主要来源于根系分泌物与凋落物[24], 由于气温较低, 表层堆积大量凋落物, 微生物活性弱(图 3), 不利于SOC分解, 因此大部分转化成性质最稳定的惰性碳固定于土壤中。SOC、Cs、Cp的最小值在海拔2600—2800 m, 与其同时此海拔也为MBC的峰值(图 3), 原因可能为海拔处于森林上限低温高湿的环境, 云杉长势稀疏且处于老龄化状态, 凋落物少, 因此SOC含量少[25]。再者地上的草本植物数量多, 微生物活性较强, 有利于SOC分解, 因此Cp含量较低。Cp的最大值在海拔2000—2200 m, 由于草本植物存活周期短, 及牛马等动物的啃食, 大多数死亡根系回归土壤, 增加了有机质的含量。但因为研究区植被单一, 微生物种群数量少, 导致分解的有机碳少, 因此转化为最稳定的惰性碳保留在土壤中。
3.2 土壤活性碳库沿海拔的配比关系及对气候变化的响应以相关性分析探讨海拔梯度上土壤三种碳库与气候要素、碳库活度之间的关系, 结果如图 4所示, 海拔与Cp仅在A层中呈极显著的负相关(P < 0.001), 其余土层均无显著性关系;除D层外, 其余4个土层均与碳库活度呈显著的正相关(P < 0.05)。就三种碳库的关系而言, A层中Ca与Cs、Cp均呈显著的正相关(P < 0.05), 并随土层的加深, 呈极显著的正相关(P < 0.01);在所有土层中, Cs与Cp均呈极显著的正相关(P < 0.01)。碳库活度与Ca在5个土层中均呈显著的正相关(P < 0.05);与Cp仅在A层、B层、C层中呈显著的负相关(P < 0.05);与Cs仅在A层、E层中呈显著的负相关(P < 0.05);除D层外, 碳库活度与AAT(Annual average temperature)、AAP(Annual average precipitation)分别呈显著的负相关、正相关(P < 0.05)。Cp分别与AAT、AAP仅在A层中呈极显著的正相关、负相关(P < 0.001)。

|
| 图 4 海拔梯度上天山中段北坡森林土壤碳库三种稳定性组分与气候要素、碳库活度之间的相关性 Fig. 4 Correlations between three stable components of the forest soil carbon pool and climatic elements and carbon pool activity on the northern slopes of the middle part of the Tian Shan along the elevation gradients Elevation: 海拔;Ca: 活性碳库(active carbon pool);Cs:缓效性碳库(slow carbon pool);Cp:惰性碳库(passive carbon pool);AAT:年均温(Annual average temperature);AAP:年降水量(Annual average precipitation);A: 碳库活度(carbon pool activity)Horizon A:土层A(0—10cm);Horizon B:土层B(10—20cm);Horizon C:土层C(20—40cm);Horizon D:土层D(40—60cm);Horizon E: 土层E(60—100cm);*, **, ***分别代表P < 0.05, P < 0.01, P < 0.001;对角线上显示的是各元素的分布图, 上三角矩阵中显示的是相关系数以及显著性水平, 下三角矩阵中具有拟合线的各元素之间的散点图 |
气候变化主要在两个方面影响土壤碳库, 一方面是温度、降水变化影响植物生产力速率和凋落速率;另一方面是气候变化影响微生物活性从而改变表层凋落物和土壤有机碳分解速率[26]。本研究中, 随着海拔梯度的升高, 碳库活度较高, 高海拔的Ca分配比较高, 在森林上限的A层中甚是如此, 而碳库活度与Ca呈显著的正相关关系, 且此区域温度较低, 降雨量增多, 微生物分解较快, 从而使得更多的惰性碳转化成活性碳。全球变暖主要发生在高海拔高纬度地区[27], 这意味着未来在全球变暖的情况下可能导致高海拔地区的活性碳库分解较快, 从而会有一部分土壤表层碳释放到大气中, 加速气候变暖, 同时导致了森林上限的碳库损失, 这与周焱等[28], 向成华等[29]研究一致。而低、中海拔的土层碳库中以Cs、Cp为主, 由于惰性碳分解慢, 周转时间长, 被认为是“碳汇”的标志, 即森林植被下的表层土壤将更多的碳固定在土壤中, 有利于减缓土壤碳向大气释放更多的CO2, 减缓气候变暖。不同海拔的土壤碳库动态变化主要发生表层, 这可能由于土壤微生物、土壤湿度、土壤温度等因子对表层土壤的影响较强[28]。总体上, 气候与各种干扰对表层土壤的影响较大, 正反馈也主要归因于表层土壤的碳库损失, 深层土壤中的碳库不太容易受到气候变暖的影响[30]。本研究结果, 忽略了碳组分的敏感性, 因碳组分的敏感性有所差异, 稳定性碳组分的温度敏感性高, 而易变碳组分低[31—32], 但是也并非如此[33]。因此后期实验条件允许的情况下, 建议增加对天山中段北坡各碳组分敏感性的测定。尽管如此, 通过建立气候因子和碳库活度之间的关系能为了解天山森林对全球气候变暖的响应提供参考。
3.3 土壤微生物量碳及碳库指数对土壤质量的表征作用土壤微生物量碳是土壤有机质中最为活跃的部分, 可更加敏感地反映土壤有机碳的变化, 也可及时准确地反映土壤质量[34]。本研究中, MBC的最小值在海拔1800—2000 m, 由于植物以灌木类居多, 高温低湿的环境限制了微生物的活动。王卫霞等[35]研究也表明土壤微生物较依赖湿度高的土壤环境。
土壤微生物熵是指土壤微生物量碳占土壤有机碳的百分比(MBC/SOC), 其比值越高, 表征土壤有机碳的活性程度越高, 土壤中有机碳向微生物量转化的速率越快, 微生物对有机碳的利用率越高[36]。本研究中, 除海拔2400—2600 m为土壤qMB最小值外, 高海拔的土壤qMB值显著高于低海拔(图 3), 即降水量多且温度适宜的高海拔土壤微生物在分解有机碳的过程中, 可将更多的有机碳转化为MBC, 且土壤微生物进一步分解活性碳以增加土壤微生物活性和数量[37], 从而使得土壤MBC的增加与土壤SOC含量的下降, qMB值增大, 可知此海拔(2600—2800 m)土壤质量较高;而温度较高的低海拔土壤碳转化为MBC的较少, 土壤养分有效性较低。
本研究中, 海拔2400—2600 m为qMB的最小值, 原因可能为此海拔低温高湿, 其次含有丰富的凋落物, 再者微生物活性较低, 对有机碳的利用较低。但此海拔的qMB垂直剖面值随着土层的加深, 大致呈升高的趋势, 表明土壤中的碳素处于积累状态, 且深层土壤中的碳积累强度高于表层土壤, SOC逐渐由表层向深层转移, 这一研究结果与易志刚等[14]的研究结果一致。土壤碳库的变化主要发生在活性碳库中, 活性有机碳直接参与土壤生物化学转化过程, 同时也是土壤微生物活动的能源和土壤氧化的驱动力, 对于活性有机碳的研究是森林土壤碳库动态及调控机理研究的重要方面[29]。土壤碳库活度(A)、碳库指数(CPI)越高, 则土壤有机碳的稳定性越差, 土壤碳库质量越高[38], 海拔2400—2600 m全剖面的A、AI、CPI、SOCD值均最大, 表明其土壤处于一个向良性发展的状态。
4 结论将土壤总有机碳库按照对气候变暖的稳定性分为Ca、Cs、Cp能更好地反映土壤碳库的海拔分异特征。本研究中, 土壤总有机碳库与三种碳库随海拔高度呈现出先增加后减少的规律, 温度、水分是影响碳库变化的重要因子。SOC、Ca、Cs均在海拔2400—2600 m处达到最大值, 土壤含量变化除0—10 cm土层变化规律不同, 其它土层均一致;Cp在海拔2000—2200 m达到最大值, 但其在各海拔的分异规律相差不大;表明海拔变化所引起的土壤碳库的变化是影响天山中段北坡森林土壤有机碳稳定性的重要因素。本研究区森林土壤Ca分配比例沿海拔升高而增加, 这意味着高海拔区域更大幅度的气候变暖将呈现正反馈效应。
| [1] |
Lal R. Soil carbon sequestration impacts on global climate change and food security. Science, 2004, 304(5677): 1623-1627. DOI:10.1126/science.1097396 |
| [2] |
Birdsey R A, Plantinga A J, Heath L S. Past and prospective carbon storage in United States forests. Forest Ecology and Management, 1993, 58(1/2): 33-40. |
| [3] |
Luo Z K, Viscarra Rossel R A, Shi Z. Distinct controls over the temporal dynamics of soil carbon fractions after land use change. Global Change Biology, 2020, 26(8): 4614-4625. DOI:10.1111/gcb.15157 |
| [4] |
Xu X F, Thornton P E, Post W M. A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Global Ecology and Biogeography, 2013, 22(6): 737-749. DOI:10.1111/geb.12029 |
| [5] |
Lavallee J M, Soong J L, Cotrufo M F. Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century. Global Change Biology, 2020, 26(1): 261-273. DOI:10.1111/gcb.14859 |
| [6] |
Davidson E A. Carbon dioxide loss from tropical soils increases on warming. Nature, 2020, 584(7820): 198-199. DOI:10.1038/d41586-020-02266-9 |
| [7] |
朱凌宇, 潘剑君, 张威. 祁连山不同海拔土壤有机碳库及分解特征研究. 环境科学, 2013, 34(2): 668-675. DOI:10.13227/j.hjkx.2013.02.037 |
| [8] |
Turetsky M R, Abbott B W, Jones M C, Anthony K W, Olefeldt D, Schuur E A G, Koven C, McGuire A D, Grosse G, Kuhry P, Hugelius G, Lawrence D M, Gibson C, Sannel A B K. Permafrost collapse is accelerating carbon release. Nature, 2019, 569(7754): 32-34. DOI:10.1038/d41586-019-01313-4 |
| [9] |
吴玥, 赵盼盼, 林开淼, 周嘉聪, 吕茂奎, 张秋芳, 郑蔚, 程蕾, 徐建国, 陈岳民. 戴云山黄山松林土壤碳组分的海拔变化特征及影响因素. 生态学报, 2020, 40(16): 5761-5770. |
| [10] |
Brevik E C. The potential impact of climate change on soil properties and processes and corresponding influence on food security. Agriculture, 2013, 3(3): 398-417. DOI:10.3390/agriculture3030398 |
| [11] |
Zhang G N, Chen Z H, Zhang A M, Chen L J, Wu Z J. Influence of climate warming and nitrogen deposition on soil phosphorus composition and phosphorus availability in a temperate grassland, China. Journal of Arid Land, 2014, 6(2): 156-163. DOI:10.1007/s40333-013-0241-4 |
| [12] |
Zhang M, Zhang X K, Liang W J, Jiang Y, Dai G H, Wang X G, Han S J. Distribution of soil organic carbon fractions along the altitudinal gradient in Changbai Mountain, China. Pedosphere, 2011, 21(5): 615-620. DOI:10.1016/S1002-0160(11)60163-X |
| [13] |
徐侠, 陈月琴, 汪家社, 方燕鸿, 权伟, 阮宏华, 徐自坤. 武夷山不同海拔高度土壤活性有机碳变化. 应用生态学报, 2008, 19(3): 539-544. DOI:10.13287/j.1001-9332.2008.0125 |
| [14] |
易志刚, 蚁伟民, 丁明懋, 周丽霞, 张德强, 王新明. 鼎湖山自然保护区土壤有机碳、微生物生物量碳和土壤CO2浓度垂直分布. 生态环境, 2006, 15(3): 611-615. DOI:10.3969/j.issn.1674-5906.2006.03.033 |
| [15] |
崔鸿侠, 潘磊, 黄志霖, 曾立雄, 付甜, 胡文杰. 神农架不同海拔巴山冷杉林土壤活性有机碳特征. 西南林业大学学报, 2017, 37(1): 110-115. |
| [16] |
孙雪娇, 常顺利, 张毓涛, 李吉玫. 天山森林植物功能性状与碳库沿海拔梯度的变化. 生态学报, 2018, 38(14): 4994-5005. |
| [17] |
仇瑶, 常顺利, 张毓涛, 王文栋, 何平, 王慧杰, 谢锦. 天山林区六种灌木生物量的建模及其器官分配的适应性. 生态学报, 2015, 35(23): 7842-7851. |
| [18] |
Rovira P, Vallejo V R. Labile and recalcitrant pools of carbon and nitrogen in organic matter decomposing at different depths in soil: an acid hydrolysis approach. Geoderma, 2002, 107(1/2): 109-141. |
| [19] |
Brookes P C, Powlson D S, Jenkinson D S. Measurement of microbial biomass phosphorus in soil. Soil Biology and Biochemistry, 1982, 14(4): 319-329. DOI:10.1016/0038-0717(82)90001-3 |
| [20] |
Blair G J, Lefroy R D B, Lisle L. Soil carbon fractions based on their degree of oxidation, and the development of a carbon management index for agricultural systems. Australian Journal of Agricultural Research, 1995, 46(7): 1459-1466. DOI:10.1071/AR9951459 |
| [21] |
徐艳, 张凤荣, 段增强, 张琳, 孔祥斌. 区域土壤有机碳密度及碳储量计算方法探讨. 土壤通报, 2005, 36(6): 836-839. |
| [22] |
董力轩, 常顺利, 张毓涛. SWAT模型在天山林区林冠截留过程中的改进应用. 生态学报, 2022, 42(18): 7630-7640. |
| [23] |
宫立, 刘国华, 李宗善, 叶鑫, 王浩. 川西卧龙岷江冷杉林土壤有机碳组分与氮素关系随海拔梯度的变化特征. 生态学报, 2017, 37(14): 4696-4705. |
| [24] |
赵元, 张伟, 胡培雷, 肖峻, 王克林. 桂西北喀斯特峰丛洼地不同植被恢复方式下土壤有机碳组分变化特征. 生态学报, 2021, 41(21): 8535-8544. |
| [25] |
阿米娜木·艾力, 常顺利, 张毓涛, 仇瑶, 何平. 天山云杉森林土壤有机碳沿海拔的分布规律及其影响因素. 生态学报, 2014, 34(7): 1626-1634. |
| [26] |
房蕊, 于镇华, 李彦生, 谢志煌, 刘俊杰, 王光华, 刘晓冰, 陈渊, 刘居东, 张少庆, 吴俊杰, Herbert S J, 金剑. 大气CO2浓度和温度升高对农田土壤碳库及微生物群落结构的影响. 中国农业科学, 2021, 54(17): 3666-3679. |
| [27] |
秦纪洪, 王琴, 孙辉. 川西亚高山-高山土壤表层有机碳及活性组分沿海拔梯度的变化. 生态学报, 2013, 33(18): 5858-5864. |
| [28] |
周焱, 徐宪根, 阮宏华, 汪家社, 方燕鸿, 吴焰玉, 徐自坤. 武夷山不同海拔高度土壤有机碳矿化速率的比较. 生态学杂志, 2008, 27(11): 1901-1907. |
| [29] |
向成华, 栾军伟, 骆宗诗, 宫渊波. 川西沿海拔梯度典型植被类型土壤活性有机碳分布. 生态学报, 2010, 30(4): 1025-1034. |
| [30] |
Wang M M, Guo X W, Zhang S, Xiao L J, Mishra U, Yang Y H, Zhu B, Wang G C, Mao X L, Qian T, Jiang T, Shi Z, Luo Z K. Global soil profiles indicate depth-dependent soil carbon losses under a warmer climate. Nature Communications, 2022, 13(1): 5514. |
| [31] |
Knorr W, Prentice I C, House J I, Holland E A. Long-term sensitivity of soil carbon turnover to warming. Nature, 2005, 433(7023): 298-301. |
| [32] |
Fang C M, Smith P, Moncrieff J B, Smith J U. Similar response of labile and resistant soil organic matter pools to changes in temperature. Nature, 2005, 433(7021): 57-59. |
| [33] |
Garcia-Pausas J, Casals P, Camarero L, Huguet C, Sebastià M T, Thompson R, Romanyà J. Soil organic carbon storage in mountain grasslands of the Pyrenees: effects of climate and topography. Biogeochemistry, 2007, 82(3): 279-289. |
| [34] |
刘放, 吴明辉, 魏培洁, 贾映兰, 陈生云. 疏勒河源高寒草甸土壤微生物生物量碳氮变化特征. 生态学报, 2020, 40(18): 6416-6426. |
| [35] |
王卫霞, 史作民, 罗达, 刘世荣, 卢立华. 南亚热带3种人工林土壤微生物生物量和微生物群落结构特征. 应用生态学报, 2013, 24(7): 1784-1792. |
| [36] |
吴秀芝, 刘秉儒, 阎欣, 刘任涛, 安慧. 荒漠草地土壤微生物生物量和微生物熵对沙漠化的响应. 应用生态学报, 2019, 30(8): 2691-2698. |
| [37] |
张于光, 张小全, 刘学端, 肖烨, Wu L Y. 不同林型土壤微生物有机碳降解基因的多样性. 生态学报, 2007, 27(4): 1412-1419. |
| [38] |
钱虹宇, 周宏鑫, 罗原骏, 蒲玉琳, 张世熔, 李婷, 贾永霞. 土壤活性有机碳及碳库管理指数对高寒湿地退化的响应. 生态学杂志, 2020, 39(7): 2273-2282. |