 2023, Vol. 43
2023, Vol. 43文章信息
- 刘文婷, 张金峰, 刘琪璟
- LIU Wenting, ZHANG Jinfeng, LIU Qijing
- 长白山东坡白桦-长白落叶松林优势种群结构与动态特征
- Dominant population structure and dynamic characteristics of Betula platyphylla and Larix olgensis forest on the eastern slope of Changbai Mountain, Northeast China
- 生态学报. 2023, 43(18): 7462-7473
- Acta Ecologica Sinica. 2023, 43(18): 7462-7473
- http://dx.doi.org/10.20103/j.stxb.202209272747
-
文章历史
- 收稿日期: 2022-09-27
- 网络出版日期: 2023-05-08

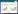




2. 北京林业大学 生态与自然保护学院, 北京 100083
2. College of Ecology and Nature Conservation, Beijing Forestry University, Beijing 100083, China
种群是同一个物种特定空间上的个体集合, 种群结构及其动态是其重要属性, 也是植物种群生态学研究的核心内容之一[1—2]。种群结构是指个体数量、高度、年龄及其空间分布情况。种群动态则是种群在整个生命过程中的数量变化情况[3—4]。种群统计是研究种群结构与动态的一种方法, 具体地是以生命表为基本信息, 结合年龄结构、存活曲线、生存分析、数量化动态分析、谱分析及时间序列预测等手段进行研究[5—6], 不仅可以揭示种群生存现状、受干扰情况, 还可以预测种群未来发展趋势, 对物种的科学保护与管理具有重要意义[7—8]。
长白山区植被资源丰富, 拥有欧亚大陆东部最典型的、保存最为完整的温带山地森林生态系统, 对该区气候调节、水土保持和维持生态系统平衡具有重要作用[9]。长白山地区在历史上经历多次火山爆发, 特别是东坡被大量火山喷发物覆盖, 对植被影响很大[10]。火山爆发的演替过程是先阳性树种, 后阴性或耐阴树种[11]。通常是原始林分遭到破坏后, 郁闭度下降, 先为阳性先锋树种的生长和更新创造条件, 恢复成次生林, 进而为阴性或耐阴树种提供生存条件, 最终发展为地带性顶极群落[10]。在整个演替过程中, 必然会出现许多过渡型森林植被类型, 了解这些过渡植被类型对群落演替及生态系统恢复具有重要意义。
白桦(Betula platyphylla)-长白落叶松(Larix olgensis)林是长白山林区暗针叶林林遭到采伐或火灾等干扰后形成的过渡性植被类型[12]。实地调查发现, 该阶段群落结构简单, 层次化分不明显, 其中白桦和长白落叶松几乎占据绝对优势[9—11]。先前的研究表明, 优势树种种群结构是否完整能够预示群落在未来一段时期内的发展趋势[9, 13]。目前关于白桦和长白落叶松天然更新的研究多集中于幼龄期, 认为该阶段决定了种群的演替过程及其在群落中的地位[14—15]。然而, 从种群结构来看, 大树、老树个体也不容忽视, 因为这些个体一般占据林内上层空间, 这可能会对林下幼苗及幼树产生影响[13, 16]。另一方面, 不同种群及同一种群内不同年龄个体的差异可能影响种内和种间竞争进而决定物种的生命周期[7, 17]。鉴于此, 本文以分布于长白山东坡的白桦和长白落叶松种群为研究对象, 利用种群统计学应用方法, 分析种群年龄结构与数量特征, 探讨演替过程中的种群动态规律, 以期为白桦-长白落叶松林的保护和暗针叶林的恢复提供理论依据。
1 研究区概况与方法 1.1 研究区概况研究区位于吉林省长白山东坡。长白山属温带大陆性山地气候, 受太平洋季风影响, 夏季温暖多雨, 冬季寒冷漫长;年平均气温-7—3℃;年平均降水700—1000 mm, 多集中在7—8月。土壤类型为土地暗棕色森林土, 土层厚度20—100 cm。该区受到火山爆发的影响, 而且历次爆发的影响范围不同[11], 生境条件复杂多样, 总体上地貌为火山台地。土壤是在不同程度风化的浮石(母质)上发育起来的。浮石作为成土母质, 风化比较快, 植物也容易定居。目前分布在东坡的大面积白桦-长白落叶松林就是在这种母质上发育起来的。
此次研究样地的乔木层除白桦和长白落叶松外, 还有臭冷杉(Abies nephrolepis)、鱼鳞云杉(Picea jezoensis var. komarovii)等;灌木层有长白蔷薇(Rosa koreana)、细叶杜香(Ledum palustre)、蓝靛果忍冬(Lonicera edulis)、毛脉黑忍冬(L. nigra var. barbinervis)、东北茶藨子(Ribes mandshuricum)和西伯利亚刺柏(Juniperus sibirica)等;草本层有林奈草(Linnaea borealis)、小叶章(Calamagrostis angustifolia)、舞鹤草(Maianthemum bifolium)、苔草(Carex spp.)和越橘(Vaccinium vitis-idaea)等。植物名称来自《东北植物检索表(第二版)》[18]。
1.2 研究方法 1.2.1 样地设置与调查参照CTFS (Center for Tropical Science)和中国森林生物多样性监测网络(CForBio)的大样地建立与监测技术规范[19], 于2010年在长白山国家级自然保护区东坡海拔1440—1450 m处结合野外环境建立了面积1 hm2的标准地(128.01°—128.25° E, 41.49°—42.05° N) (图 1)。2020年8—9月, 以固定样地西北角为原点, 利用全站仪将样地分成100个10 m×10 m的小样方, 测定并记录样方内木本个体的种类、胸径、数量和高度等。

|
| 图 1 研究样地位置示意图 Fig. 1 Schematic diagram of the location of the permanent sample plot |
采用“空间推时间”的方法, 以径级结构代替年龄结构分析种群变化特征[7]。根据白桦和长白落叶松种群的生长规律以及样地内树木直径分化情况, 参考胡尔查等[20]对白桦种群的划分标准, 将DBH≤2.5 cm的白桦幼苗划分为第Ⅰ龄级, 之后采用上限排外法, 以3 cm为径阶距进行划分, 将白桦种群分为7个龄级, 即Ⅱ级(2.5—5.5 cm)、Ⅲ级(5.5—8.5 cm)、Ⅳ级(8.5—11.5 cm)、Ⅴ级(11.5—14.5 cm)、Ⅵ级(14.5—17.5 cm)、Ⅶ级(>17.5 cm);参考张钦弟等[21]对落叶松种群结构的划分标准, 以5 cm为径阶距, 将长白落叶松划分为13个龄级, 即Ⅰ级(DBH≤5 cm)、Ⅱ级(5—10 cm)、Ⅲ级(10—15 cm)、Ⅳ级(15—20 cm)、Ⅴ级(20—25 cm)、Ⅵ级(25—30 cm)、Ⅶ级(30—35 cm)、Ⅷ级(35—40 cm)、Ⅸ级(40—45 cm)、Ⅹ级(45—50 cm)、Ⅺ级(50—55 cm)、Ⅻ级(55—60 cm)、ⅩⅢ级(>60 cm)。
1.2.3 编制静态生命表静态生命表又称为特定时间生命表, 反映了种群在某一特定时刻的数量剖面或廓线[22]。参考江洪的计算方法[23], 编制静态生命表。
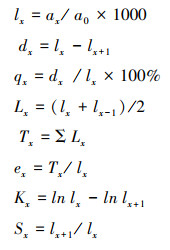
|
式中, x为龄级;ax为龄级内存活个体数, a0为ax的初始值;lx为x龄级标准化存活个体数;dx为x到x+1龄级的标准化后死亡个体数;qx为x到x+1龄级的树木个体死亡率;Lx为x到x+1龄级的个体平均寿命和;Tx为大于等于x龄级的全部个体的平均寿命和;ex为进入x龄级的存活个体的平均期望值;Kx为各年龄组的消失率, lnlx为lx的对数;Sx为种群存活率。
1.2.4 量化种群动态采用陈晓德[5]种群动态量化分析方法, 定量描述长白山东坡白桦和长白落叶松的种群动态, 计算公式如下:
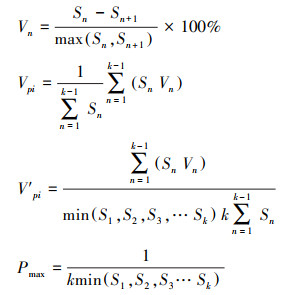
|
式中, Vn 代表种群内相邻两个龄级间个体数量变化;n为龄级;Sn和Sn+1分别是第n和第n+1龄级的个体数;Vpi代表忽略外部干扰因素时, 整个种群年龄结构变化动态指数;k表示种群的龄级数量;而当考虑未来外部干扰时, Vpi还与各龄级个体数(Sn)以及龄级数量(k)相关。V′pi为考虑外部干扰因素时, 整个种群年龄结构变化动态指数;P为种群承担的风险概率, P值最大时, 对种群的Vpi影响最大。Vn、Vpi、V′pi取正值、负值、零时分别反应种群相邻两个龄级个体数量的增长、衰退、稳定的动态关系。
1.2.5 拟合存活曲线、死亡率曲线和消失率曲线存活曲线是借助各个龄级存活个体的自然对数值与龄级作图描述种群动态变化的一种分析方法, 能够反映种群个体在各个龄级的存活状态[24]。为了更准确确定种群属于哪种存活曲线类型, 参考Hett和Loucks[3]的数学模型, 采用指数方程Nx = N0e-bx和幂函数方程Nx = N0e-b进行检验。其中, Nx为x龄级存活个体数;N0为种群初始个体数;b为死亡率。根据决定系数(R2)和F检验值判断拟合效果。
以不同龄级为横坐标, 死亡率qx和消失率Kx为纵坐标, 分别绘制死亡率曲线和消失率曲线。
1.2.6 生存分析根据种群生存率函数S(i)、累积死亡率函数F(i)、死亡密度函数f(ti)及危险率函数λ(ti)对种群的生存状况进行定量分析。具体计算公式见杨凤翔等[25]的研究。
1.2.7 谱分析谱分析能够用来探讨和揭示树木年龄更替过程的周期性和集中分布的波动性, 是Fourier级数的展开式, 通常写成正弦波形式[26]:
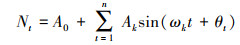
|
式中, Nt为t时刻种群的大小;A0为周期变化的平均值;t是种群各年龄分布的时间系列长度;n为系列总长度, 即龄级总数;Ak为各谐波的振幅(k=1, 2, 3, …, p;p=n/2, 为谐波总个数), 值的差异反映了各周期所起作用的大小;ωk及θk分别为谐波频率及相角。
Fourier分解的参数可以用以下公式进行估算:
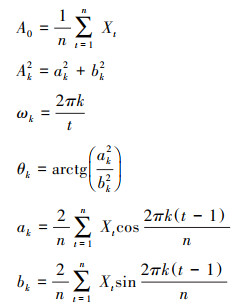
|
式中, ak、bk为参数估计值; Xt为t年龄序列的个体数, 考虑到样地中种群各龄级数量差异较大, 在计算前对Xt进行对数处理, 即用用ln(Xt + 1)代替Xt(该值与生命表中ax栏内数值相对应);利用谱分析的公式计算出各个波形的振幅Ak值。A1为基波, 表现了基本周期的波动, 其周期长度为种群本身所固有, 由种群波动特性决定。A2—Ak为谐波。每个谐波的周期分别是基本周期的1/2, 1/3, …, 1/p[27]。
1.2.8 时间序列模型采用时间序列预测中的一次平均推移法, 对白桦种群和长白落叶松种群的年龄结构进行模拟和预测。计算公式见肖宜安等[28]的研究。
1.3 数据处理本研究所有数据分析在Excel 2010和SPSS 20进行, 所有的图用Origin 2020软件绘制。
2 结果与分析 2.1 种群分布现状及结构特征本次共调查到白桦个体234株, 最大胸径19.30 cm;长白落叶松613株, 最大胸径为67.40 cm。由图 2可以看出, 白桦种群的龄级结构呈典型倒“J”型分布, 各龄级数量差异较大。其中, Ⅰ、Ⅱ龄级个体数为176株, 占个体总数的75.20%, 说明白桦幼苗和幼树的数量较多, 种群具有一定的增长潜力。长白落叶松的龄级结构呈现出近似的钟形结构。其中, 种群个体数在Ⅰ龄级最多(148株), 占总个体数的24.14%, 数量上与相邻的Ⅱ龄级几乎为两个极端, 说明林下大部分幼苗不能进入主林层;Ⅶ—Ⅸ龄级个体数占36.71%, 即种群内大树占优势。

|
|
图 2 白桦和长白落叶松种群龄级结构
Fig. 2 Age class structure of Betula platyphylla and Larix olgensis populations
根据胸径(DBD, Diameter at breast height)大小, 将白桦种群划分为7个龄级(Ⅰ: DBH≤2.5 cm; Ⅱ: 2.5 cm |
表 1显示, 随着龄级增加, 白桦种群的标准化存活率(lx)逐渐降低;长白落叶松种群的标准化存活率呈先降后增的连续波动, 在Ⅷ龄级达到峰值。白桦种群的生命期望(ex)总体呈先增后降的趋势, Ⅱ龄级表现出最高的生命期望, 说明该阶段白桦种群生长比较旺盛;长白落叶松种群的生命期望表现为先增加后降低的趋势, Ⅲ龄级生命期望值最高, 说明幼龄期过后, 存活下来的个体生存能力增强。
| 种群 Population |
龄级 Age class |
径级 Diameter class/cm |
ax | lx | lnlx | dx | qx | Lx | Tx | ex | Kx | Sx | |
| 白桦 | Ⅰ | < 2.5 | 123 | 1000.00 | 6.91 | 569.11 | 0.57 | 715.45 | 1386.18 | 1.39 | 0.84 | 0.43 | |
| Betula platyphylla | Ⅱ | 2.5—5.5 | 53 | 430.89 | 6.07 | 170.73 | 0.40 | 345.53 | 670.73 | 1.56 | 0.50 | 0.60 | |
| Ⅲ | 5.5—8.5 | 32 | 260.16 | 5.56 | 130.08 | 0.50 | 195.12 | 325.20 | 1.25 | 0.69 | 0.50 | ||
| Ⅳ | 8.5—11.5 | 16 | 130.08 | 4.87 | 73.17 | 0.56 | 93.50 | 130.08 | 1.00 | 0.83 | 0.44 | ||
| Ⅴ | 11.5—14.5 | 7 | 56.91 | 4.04 | 40.65 | 0.71 | 36.59 | 48.78 | 0.86 | 1.25 | 0.29 | ||
| Ⅵ | 14.5—17.5 | 2 | 16.26 | 2.79 | 8.13 | 0.50 | 12.20 | 16.26 | 1.00 | 0.69 | 0.50 | ||
| Ⅶ | >17.5 | 1 | 8.13 | 2.10 | 8.13 | 1.00 | 4.07 | 4.07 | — | 2.10 | - | ||
| 长白落叶松 | Ⅰ | <5 | 148 | 1000.00 | 6.91 | 837.84 | 0.84 | 581.08 | 2070.95 | 2.07 | 1.82 | 0.16 | |
| Larix olgensis | Ⅱ | 5—10 | 24 | 162.16 | 5.09 | 27.03 | 0.17 | 148.65 | 1489.86 | 9.19 | 0.18 | 0.83 | |
| Ⅲ | 10—15 | 20 | 135.14 | 4.91 | -141.89 | -1.05 | 206.08 | 1341.22 | 9.93 | -0.72 | 2.05 | ||
| Ⅳ | 15—20 | 41 | 277.03 | 5.62 | -81.08 | -0.29 | 317.57 | 1135.14 | 4.10 | -0.26 | 1.29 | ||
| Ⅴ | 20—25 | 53 | 358.11 | 5.88 | -20.27 | -0.06 | 368.24 | 817.57 | 2.28 | -0.06 | 1.06 | ||
| Ⅵ | 25—30 | 56 | 378.38 | 5.94 | -141.89 | -0.38 | 449.32 | 983.11 | 2.60 | -0.32 | 1.38 | ||
| Ⅶ | 30—35 | 77 | 520.27 | 6.25 | -27.03 | -0.05 | 533.78 | 1033.78 | 1.99 | -0.05 | 1.05 | ||
| Ⅷ | 35—40 | 81 | 547.30 | 6.31 | 94.59 | 0.17 | 500.00 | 797.30 | 1.46 | 0.19 | 0.83 | ||
| Ⅸ | 40—45 | 67 | 452.70 | 6.12 | 310.81 | 0.69 | 297.30 | 418.92 | 0.93 | 1.16 | 0.31 | ||
| Ⅹ | 45—50 | 21 | 141.89 | 4.95 | 40.54 | 0.29 | 121.62 | 195.95 | 1.38 | 0.34 | 0.71 | ||
| Ⅺ | 50—55 | 15 | 101.35 | 4.62 | 54.05 | 0.53 | 74.32 | 108.11 | 1.07 | 0.76 | 0.47 | ||
| Ⅻ | 55—60 | 7 | 47.30 | 3.86 | 27.03 | 0.57 | 33.78 | 43.92 | 0.93 | 0.85 | 0.43 | ||
| ⅩⅢ | >60 | 3 | 20.27 | 3.01 | 20.27 | 1.00 | 10.14 | 10.14 | 3.01 | - | |||
| ax: x龄级内存活个体数Actual survival number x age class; lx: x龄级的标准化存活个体数Logarithmic standard surviving number in x age; dx: 从x到x+1龄级的死亡个体数Standardized deaths from x to x + 1; qx: 从x到x+1龄级的种群个体死亡率Mortality rate from x to x+1; Lx: 从x到x+1龄级的平均存活个体数Survived individuals from x to x + 1; Tx: 大于等于x龄级的存活个体总数Total survival number of individuals over x age class; ex: 平均期望寿命Average life expectancy; lnlx: 标准化存活数对数The natural logarithm of lx; Kx: 种群消失率 Vanish rate; Sx: 种群存活率Survival rate | |||||||||||||
种群动态量化分析结果见表 2。白桦种群V1— V6大于0, 说明种群在不同龄级均呈增长趋势。长白落叶松种群V3—V7小于0, V1—V2、V8—V12大于0, 表明种群数量呈现出增长—衰退—增长的动态变化趋势。忽略外界干扰时, 白桦和长白落叶松种群的Vpi均为正值, 说明两个物种种群均为增长型。考虑外界干扰时, 白桦种群V′pi远高于长白落叶松种群(7.45%和0.69%), 且后者趋近于0。此外, 白桦种群随机干扰所承担的最大值Pmax大于长白落叶松种群(14.29和2.56), 这些结果说明, 受外界干扰时, 白桦种群表现为增长型, 而长白落叶松种群增长趋势不明显, 更倾向于稳定性。
| 动态指数级 Dynamic index class |
动态指数 Dynamic index value/% | 动态指数级 Dynamic index class |
动态指数 Dynamic index value/% | |||
| 白桦 Betula platyphylla | 落叶松 Larix olgensis | 白桦 Betula platyphylla | 落叶松 Larix olgensis | |||
| V1 | 56.91 | 83.78 | V9 | — | 68.66 | |
| V2 | 39.62 | 16.67 | V10 | — | 28.57 | |
| V3 | 50.00 | -51.22 | V11 | — | 53.33 | |
| V4 | 56.25 | -22.64 | V12 | — | 57.14 | |
| V5 | 71.43 | -5.36 | Vpi | 52.14 | 26.85 | |
| V6 | 50.00 | -27.27 | V′pi | 7.45 | 0.69 | |
| V7 | — | -4.94 | Pmax | 14.29 | 2.56 | |
| V8 | — | 17.28 | ||||
| V1—V12: 相邻龄级数量动态变化指数Quantitative dynamic index of adjacent age classes; Vpi: 忽略外部干扰时种群的数量动态变化指数Quantitative dynamic index of population without external interference; V′pi: 考虑外部干扰时种群的数量动态变化指数Quantitative dynamic index of population with external interference; Pmax: 种群承担的最大的风险概率The maximum probability of risk assumed by the population | ||||||
由图 3可知, 白桦种群在第Ⅰ—Ⅳ龄级下降缓慢, Ⅴ龄级下降趋势明显, 第Ⅵ龄级下降趋势又有所减缓, 呈现“凸-凹-凸”的变化特点;长白落叶松的存活曲线在整个龄级范围内波动较大。用2种数学模型方程拟合的结果显示, 白桦和长白落叶松的指数函数模型中R2和F值均大于幂函数模型, P值也更小(表 3), 说明指数模型的拟合效果更好, 两个种群存活曲线都符合Deevey-Ⅱ型。

|
| 图 3 白桦和长白落叶松种群存活曲线 Fig. 3 Survival curve of Betula platyphylla and Larix olgensis populations |
| 种群 Population | 方程 Equation | R2 | F | P | 存活曲线类型 Type |
| 白桦 | Nx=310.644e-0.804x | 0.987 | 384.267 | <0.001 | Deevey-Ⅱ |
| Betula platyphylla | Nx=193.047x-2.280 | 0.878 | 36.046 | 0.002 | |
| 长白落叶松 | Nx=105.445e-0.175x | 0.396 | 7.201 | 0.021 | Deevey-Ⅱ |
| Larix olgensis | Nx=110.976x-0.736 | 0.270 | 4.069 | 0.069 | |
| Nx: x龄级内存活数Actual survival number x age class | |||||
死亡率和消失率曲线反映了种群数量动态变化的过程。图 4显示, 白桦和长白落叶松种群个体死亡率(qx)和消失率(Kx)曲线变化趋势基本一致, 均表现为连续先降后增的动态变化趋势, 分别在第Ⅶ和第ⅩⅢ龄级出现消亡高峰。此外, 长白落叶松种群在Ⅲ—Ⅶ龄级的死亡率为负, 说明该种群幼龄阶段个体死亡严重。

|
| 图 4 白桦和长白落叶松种群死亡率和消失率曲线 Fig. 4 Mortality and vanish rate curve of Betula platyphylla and Larix olgensis populations |
图 5所示, 随着龄级增大, 白桦种群的生存率呈单调递减, 累计死亡率呈单调递增, 累计死亡率始终大于生存率。长白落叶松种群的生存率总体表现为先升后降的趋势, 累计死亡率为先降后升, Ⅰ—Ⅷ龄级之间, 生存率和死亡率存在较大波动, 可能与个体间的竞争和水热条件有关;Ⅷ龄级后累计死亡率大于生存率, 且逐渐趋近于0, 说明种群开始生理衰竭。

|
| 图 5 白桦和长白落叶松种群的生存率、累计死亡率、死亡密度、危险率函数曲线 Fig. 5 Functional curves of survival rate, cumulative mortality rate, death density and risk rate of Betula platyphylla and Larix olgensis populations |
白桦种群的死亡密度函数f(ti)和危险率函数λ(ti)整体表现出递减的趋势。其中, 死亡密度函数在Ⅰ—Ⅱ龄级急剧下降, 说明个体数量在这个阶段锐减;长白落叶松种群的死亡密度函数在第Ⅰ—Ⅲ龄级单调递减, 尔后变化趋势较为平缓, 而危险率函数在第Ⅰ—Ⅲ龄级骤降, 随后逐渐上升, 表明种群面临死亡风险。
2.5 谱分析白桦种群总的龄级为7级, 采用内插法补充为8级, 总的波序k=n/2=4;长白落叶松种群分为13级, 同样的方法补充为14级, 总的波序k=n/2=7。由表 4可以看出, 白桦和长白落叶松种群基波均最大(1.63和0.85), 说明两个物种种群的数量动态变化主要是物种的生物学特性控制的, 受天然更新的作用较大, 具有明显的大周期。从谐波的变化可以看出, 白桦种群表现为递减的变化趋势, 而长白落叶松种群在生长过程中受小周期波动的影响, 具体表现在1/4和1/5处(表 4), 对应空间序列Ⅶ、Ⅷ龄级, 可能是竞争加剧导致的波动, 这种小周期的波动有助于维持种群自身的稳定性。
| 种群 Population | A0 | A1 | A2 | A3 | A4 | A5 | A6 | A7 |
| 白桦 Betula platyphylla | 2.716 | 1.634 | 0.895 | 0.683 | 0.683 | - | - | - |
| 长白落叶松 Larix olgensis | 3.488 | 0.851 | 0.569 | 0.421 | 0.538 | 0.511 | 0.362 | 0.362 |
| A0: 周期变化的平均值Average value of periodic change; A1: 基波振幅Fundamental amplitude; A1—A7: 各谐波振幅Amplitude of each harmonic | ||||||||
时间序列预测的结果表明, 白桦种群在未来的2、4、6个龄级后, 各龄级的个体数量均呈现增加趋势, 并且趋势逐渐增大(表 5)。例如, 在第Ⅴ龄级, 个体存活数经过2、4个龄级之后, 分别增加了71.42%、285.74%;
| 种群 Population |
龄级 Age class |
初始个体数 Initial number of individuals |
Mt | |||
| M2 | M4 | M6 | M8 | |||
| 白桦 | Ⅰ | 123 | ||||
| Betula platyphylla | Ⅱ | 53 | 88 | |||
| Ⅲ | 32 | 43 | ||||
| Ⅳ | 16 | 24 | 56 | |||
| Ⅴ | 7 | 12 | 27 | |||
| Ⅵ | 2 | 5 | 14 | 39 | ||
| Ⅶ | 1 | 2 | 7 | 19 | ||
| 长白落叶松 | Ⅰ | 148 | ||||
| Larix olgensis | Ⅱ | 24 | 86 | |||
| Ⅲ | 20 | 22 | ||||
| Ⅳ | 41 | 31 | 58 | |||
| Ⅴ | 53 | 47 | 35 | |||
| Ⅵ | 56 | 55 | 43 | 57 | ||
| Ⅶ | 77 | 67 | 57 | 45 | ||
| Ⅷ | 81 | 79 | 67 | 55 | 63 | |
| Ⅸ | 67 | 74 | 70 | 63 | 52 | |
| Ⅹ | 21 | 44 | 62 | 59 | 52 | |
| Ⅺ | 15 | 18 | 46 | 53 | 51 | |
| Ⅻ | 7 | 11 | 28 | 45 | 47 | |
| ⅩⅢ | 3 | 5 | 12 | 32 | 41 | |
| Mt(1): 经过2、4、6、8个龄级时间后种群的大小the number of individuals in each age class after the population has developed after 2, 4, 6, and 8 age class in this study;(1): 移动平均法中的一次移动平均法标识Identification of one-time moving average method in moving average method | ||||||
在第Ⅶ龄级, 经过2、4、6个龄级后, 个体存活数增加了2、7、19倍, 说明白桦种群在今后一段时间的演替中占据数量优势地位。随着时间推移, 长白落叶松种群在Ⅱ—Ⅲ、Ⅹ—ⅩⅢ龄级个体数量总体呈增加趋势, Ⅳ—Ⅸ龄级个体数量总体呈减小趋势, 综合整个长白落叶松种群的发展来看, 在一定时间内仍可以维持更新, 但是随着老龄个体的增加, 未来会逐渐进入衰退期。
3 讨论 3.1 种群年龄结构特征白桦种群个体数量随着龄级的增加而减少, 呈明显的倒“J”型分布, 而长白落叶松种群个体数量呈近似的钟形结构, 且两个物种种群Ⅰ龄级个体数量占比较大(52.57%和24.14%), 说明两个物种的幼龄个体储备丰富, 具有较大的更新潜力;白桦和长白落叶松种群的动态变化指数Vpi均大于0, 说明两个物种均为增长型。然而, 在考虑外界干扰时, 长白落叶松种群的V′pi趋近于0, 说明其更趋向于稳定型。先前的研究显示, 长白山地区经历过多次火山爆发, 最近一次火山爆发前长白山东坡为长白落叶松纯林。此次爆发, 规模不大, 对植被产生的破坏较小[29]。因此, 在生境逐渐恢复且有种源的情况下, 长白落叶松因其耐干旱、喜光的特性得以继续生长更新[10]。野外调查时注意到, 该区林分具有间伐的痕迹, 伐木使得林分郁闭度减小, 为喜光树种提供一定的生存空间。白桦作为先锋树种, 因其萌芽力强、生命力旺盛及喜光的特性得以萌发[9]。此外, 该区土壤是火山爆发后形成的浮石裸地。据刘琪璟等报道[29]该类型土壤水分养分条件较好, 植被演替较快, 不需要其它植物做先导, 因此有利于先锋树种白桦的定居。
一般, 树木幼苗是比较脆弱的阶段[30]。本研究所论述的两个树种, 尽管Ⅰ龄级个体较多, 但存活率很低, 这与树种生物学特性及环境因子有关。长白落叶松种子体积较小、质量较轻, 随着传播距离的增加其发芽率会降低, 因此幼苗多聚集分布在母树周围[13];白桦种子小且带翅, 易分散, 但不耐荫, 其幼苗多在光照充足的个别斑块进行更新[31]。因此, 在白桦-长白落叶松林内, 白桦种群也呈聚集分布。然而, 随着林分内植株生长, 受密度制约效应的影响, 聚集分布的同种幼苗个体之间及与其他邻近植株个体间可能存在激烈的种内和种间竞争, 导致个体大量枯死[32]。此外, 长白山地区冬季寒冷漫长, 最低气温低至-20℃以下, 霜冻和寒冷也可能导致幼苗枯死[33]。由此可见, 白桦和长白落叶松种群幼龄个体要进入下一阶段, 必须经过竞争以及环境因子的多重筛选, 这反映了幼龄阶段是种群更新和发展的“瓶颈”。因此, 适时进行人工间苗和保护林内的腐殖质层, 对促进白桦和长白落叶松种子萌发及其幼苗生长会有积极作用。
3.2 种群动态结构植物种群某一时刻的数量剖面和生存现状以及对环境的适应度可以通过种群静态生命表、存活曲线、死亡率、消失率曲线来判断[21—22, 25]。本次用于编制静态生命表的数据是通过径级代替龄级的方式获得, 因此, 会出现死亡率为负值的情况, 大多数研究会采用匀滑的技术解决[23]。Proctor等[34]认为出现负值虽然与数学假设不符, 但是仍能够提供有用的生态学信息。张文辉等[35]在描述沙棘(Hippophae rhamnoides)种群变化做过类似的尝试, 结论是不进行匀滑处理更有利于反映种群的现实波动状况。本研究中, 白桦种群的生命期望值总体呈现下降的趋势, 在Ⅰ龄级最大, 说明此时种群生命力较旺盛, 对环境具有较强的适应力, 这与卢杰等[36]对高山松(Pinus densata)种群和彭辉等[37]对木荷(Schima superba)的研究结果类似。长白落叶松种群的生命期望随龄级增加总体表现为先增后降的趋势。其中, 在Ⅲ龄级最大, 说明种群渡过“瓶颈期”后, 生存质量变高;从第Ⅳ龄级开始, 生命期望值开始下降, 可能是由于种间、种内竞争导致的, 这与卫波宁等[38]对小花水柏枝(Myricaria wardii)种群和吴其超等[39]对五莲杨(Populus wulianensis)种群的研究结果一致。另外, 白桦和长白落叶松种群的均为Deevey-Ⅱ型, 这一结果与陈倩等[40]对木兰围场白桦种群以及张钦弟等[21]对华北地区落叶松种群的分析一致, 说明种群有相对稳定的死亡率。然而, 该研究结果与杨慧等[41]对北京东灵地区、胡尔查[20]对内蒙古乌拉山和靳静静[42]对山西七里峪白桦种群的存活曲线属于Deevey-Ⅰ型有所不同, 说明不同地区的生境和外界干扰会影响同类种群的生存状况。
生存函数可以进一步直观反映种群结构与动态变化情况, 具有较高的实际应用价值[27]。研究发现, 随着龄级增加, 白桦种群生存率递减, 累计死亡率递增, Ⅰ—Ⅱ龄级死亡密度和危险率骤降, 表现出前期大后期小的特点, 这与生命表分析的结果相对应。长白落叶松种群Ⅰ—Ⅲ龄级生存率和死亡率存在较大波动, 死亡密度递减, 危险率骤降, 可能与个体间竞争和水热条件有关;Ⅷ龄级后累计死亡率大于生存率, 种群逐渐往衰退的方向发展。4个生存函数曲线共同表明, 白桦种群在Ⅰ—Ⅳ龄级锐减, 在Ⅳ—Ⅶ龄级相对稳定;长白落叶松种群表现为前期(Ⅰ—Ⅲ龄级)增长, 中期(Ⅲ—Ⅸ龄级)稳定, 后期(Ⅸ—ⅩⅢ龄级)衰退的特点。
3.3 种群发展趋势谱分析中不同龄级个体数量的波动情况可以直观反映种群的更新动态[28]。谱分析显示, 白桦种群和长白落叶松种群均存在明显的周期性, 且受基波影响显著, 这与李艳丽等[33]对臭冷杉种群、宋萍等[43]对桫椤(Alsophila spinulosa)种群的研究相一致, 说明种群更新受生物学特性的影响较大。然而, 长白落叶松种群在天然更新中还存在小周期的波动。其中, 发生在Ⅷ龄级的波动较为明显, 这可能与种内竞争和生理衰老有关。这种大周期内有小周期的波动是种群本身存在的内调节机制, 能够促进种群的自然稀疏及其数量调节, 使得种群在更新过程中也能保持稳定[44]。
时间序列预测分析表明, 随着时间的推移, 白桦种群各龄级个体数量均逐渐增加, 说明该种群在未来发展良好。长白落叶松种群Ⅱ—Ⅳ龄级、Ⅹ—ⅩⅢ龄级个体数在未来均呈增加趋势, 推测种群未来以小树、老树占主导地位。另外, 在白桦-长白落叶松林中, 高龄级个体虽然少但占据较大的生存空间。因此, 从群落角度来看, 少数老龄个体的消亡可促使林下其他植物个体进入上层空间, 有利于群落发展[16]。时间序列分析虽能预测种群未来的发展趋势, 但其实现条件是以当前生境和种群结构为基础。一旦生境和种群结构遭到破坏, 种群增长趋势可能会中断甚至衰退[45]。因此, 要使得种群数量稳定增长, 必须加强白桦-长白落叶松林的保护。
4 结论长白山东坡白桦-长白落叶松林正处于暗针叶林演替的早期阶段。群落中白桦和长白落叶松种群年龄结构均为增长型, 但是二者幼龄个体死亡率均较高。从森林经营的角度, 建议对幼龄个体适当采取抚育间伐措施, 减少林木的环境压力, 促进其种群的生长发育及更新。此外, 受外界干扰时, 长白落叶松种群增长趋势缓慢, 因此采取人工保护措施、避免过度的人为干扰是维持种群稳定过渡和促进森林生态系统更新的关键。
致谢: 本研究的固定样地由徐倩倩、张国春、刘文慧、徐琼瑶于2010年建立。2020年参加复查的还有:徐振招、胡博、郑东升、高浩鑫、邓发昌。北京林业大学森林培育与保护教育部重点实验室为本研究提供实验条件。| [1] |
Frost I, Rydin H. Spatial pattern and size distribution of the animal-dispersed tree Quercus robur in two spruce-dominated forests. Écoscience, 2000, 7(1): 38-44. DOI:10.1080/11956860.2000.11682569 |
| [2] |
Gurevitch J, Scheiner S M, For G A. The ecology of plants. Sunderland, Massachusetts, USA: Sinauer Associates, Inc. 2002.
|
| [3] |
Hett J M, Loucks O L. Age structure models of balsam fir and eastern hemlock. The Journal of Ecology, 1976, 64(3): 1029. DOI:10.2307/2258822 |
| [4] |
Bakker J P, Grootjans A P, Hermy M, Poschlod P. How to define targets for ecological restoration?-Introduction. Applied Vegetation Science, 2000, 3(1): 3-6. DOI:10.1111/j.1654-109X.2000.tb00033.x |
| [5] |
陈晓德. 植物种群与群落结构动态量化分析方法研究. 生态学报, 1998, 18(2): 214-217. DOI:10.3321/j.issn:1000-0933.1998.02.017 |
| [6] |
Chhetri P K, Bista R, Cairns D M. Population structure and dynamics of Abies spectabilis at treeline ecotone of Barun Valley, Makalu Barun National Park, Nepa. Acta Ecologica Sinica, 2016, 36(4): 269-274. DOI:10.1016/j.chnaes.2016.05.003 |
| [7] |
吴承祯, 洪伟, 谢金寿, 吴继林. 珍稀濒危植物长苞铁杉种群生命表分析. 应用生态学报, 2000, 11(3): 333-336. DOI:10.3321/j.issn:1001-9332.2000.03.003 |
| [8] |
Kang D, Guo Y X, Ren C J, Zhao F Z, Feng Y Z, Han X H, Yang G H. Population structure and spatial pattern of main tree species in secondary Betula platyphylla forest in Ziwuling Mountains, China. Scientific Reports, 2014, 4: 6873. DOI:10.1038/srep06873 |
| [9] |
周以良, 赵光仪. 小兴安岭-长白山林区天然次生林的类型、分布及其演替规律. 东北林学院学报, 1964(0): 33-45. |
| [10] |
戴璐, 武耀祥, 韩士杰, 李玉文. 火山大爆发对长白山东坡历史植被演替的影响. 生态学杂志, 2008, 27(10): 1771-1778. DOI:10.13292/j.1000-4890.2008.0319 |
| [11] |
王绪高, 李秀珍, 贺红士, 冷文芳, 问青春. 大兴安岭北坡落叶松林火后植被演替过程研究. 生态学杂志, 2004, 23(5): 35-41. DOI:10.3321/j.issn:1000-4890.2004.05.007 |
| [12] |
徐文铎, 何兴元, 陈玮, 刘常富. 长白山植被类型特征与演替规律的研究. 生态学杂志, 2004, 23(5): 162-174. DOI:10.3321/j.issn:1000-4890.2004.05.031 |
| [13] |
张金峰, 葛树森, 梁金花, 李俊清. 长白山阔叶红松林红松种群年龄结构与数量动态特征. 植物生态学报, 2022, 46(6): 667-677. |
| [14] |
贾炜玮, 解希涛, 姜生伟, 李凤日. 大兴安岭新林林业局3种林分类型天然更新幼苗幼树的空间分布格局. 应用生态学报, 2017, 28(9): 2813-2822. DOI:10.13287/j.1001-9332.201709.014 |
| [15] |
刘足根, 朱教君, 袁小兰, 谭辉. 辽东山区长白落叶松天然更新调查. 林业科学, 2007, 43(1): 42-49. |
| [16] |
张金峰, 葛树森, 梁金花, 李俊清. 长白山阔叶红松林紫椴种群结构与动态特征. 生态学报, 2022, 42(13): 5381-5390. |
| [17] |
匡旭, 邢丁亮, 张昭臣, 宋厚娟, 王芸芸, 房帅, 原作强, 叶吉, 蔺菲, 王绪高, 郝占庆. 长白山北坡云冷杉林和落叶松林物种组成与群落结构. 应用生态学报, 2014, 25(8): 2149-2157. |
| [18] |
傅沛云, 李冀云. 东北植物检索表 (2版). 北京: 科学出版社, 1995.
|
| [19] |
Condit R. Research in large, long-term tropical forest plots. Trends in Ecology & Evolution, 1995, 10(1): 18-22. |
| [20] |
胡尔查, 王晓江, 张文军, 海龙, 张雷, 张胜利, 徐鹏雁. 乌拉山自然保护区白桦种群的年龄结构和点格局分析. 生态学报, 2013, 33(9): 2867-2876. |
| [21] |
张钦弟, 张金屯, 苏日古嘎, 张斌, 程佳佳. 庞泉沟自然保护区华北落叶松种群生命表与谱分析. 应用与环境生物学报, 2010, 16(1): 1-6. |
| [22] |
王巍, 刘灿然, 马克平, 于顺利. 东灵山两个落叶阔叶林中辽东栎种群结构和动态. 植物学报, 1999, 41(4): 85-92. |
| [23] |
江洪. 云杉种群生态学. 北京: 中国林业出版社, 1992.
|
| [24] |
Deevey E S Jr. Life tables for natural populations of animals. The Quarterly Review of Biology, 1947, 22(4): 283-314. |
| [25] |
杨凤翔, 王顺庆, 徐海根, 李邦庆. 生存分析理论及其在研究生命表中的应用. 生态学报, 1991, 11(2): 153-158. |
| [26] |
伍业钢, 韩进轩. 阔叶红松林红松种群动态的谱分析. 生态学杂志, 1988, 7(1): 19-23. |
| [27] |
吴明作, 刘玉萃. 栓皮栎种群数量动态的谱分析与稳定性. 生态学杂志, 2000, 19(4): 23-26, 37. |
| [28] |
肖宜安, 何平, 李晓红, 邓洪平. 濒危植物长柄双花木自然种群数量动态. 植物生态学报, 2004, 28(2): 252-257. |
| [29] |
刘琪璟, 王战, 王少先. 长白山近期火山爆发对高山亚高山植被的影响. 地理科学, 1993, 13(1): 57-61, 96. |
| [30] |
Silvertown J W. Introduction to Plant Population Ecology. London: Longman Group United Kingdom, 1982.
|
| [31] |
田慧霞, 李钧敏, 毕润成, 闫明, 乔秀红. 山西太岳山白桦种群结构和空间分布格局. 生态学杂志, 2017, 36(1): 1-10. |
| [32] |
陈国鹏, 鲜骏仁, 曹秀文, 刘锦乾, 杨永红, 雷炜. 林窗对岷江冷杉幼苗生存过程的影响. 生态学报, 2016, 36(20): 6475-6486. |
| [33] |
李艳丽, 杨华, 亢新刚, 邱实. 长白山云冷杉种群结构和动态分析. 北京林业大学学报, 2014, 36(3): 18-25. |
| [34] |
Proctor M C F, Wratten S D, Fry G L A. Field and laboratory exercises in ecology. The Journal of Ecology, 1981, 69(3): 1074. |
| [35] |
张文辉, 郭连金, 刘国彬. 黄土丘陵区不同生境沙棘种群数量动态分析. 西北植物学报, 2005, 25(4): 641-647. |
| [36] |
卢杰, 郭其强, 郑维列, 徐阿生. 藏东南高山松种群结构及动态特征. 林业科学, 2013, 49(8): 154-160. |
| [37] |
彭辉, 周红敏, 王宏翔, 张弓乔. 凤阳山常绿阔叶林优势种木荷的种群结构与动态. 林业科学研究, 2022, 35(3): 179-184. |
| [38] |
卫波宁, 常子惠, 张永娟, 王素娟, 德吉. 雅鲁藏布江三大支流流域小花水柏枝种群结构及动态特征. 生态学报, 2022, 42(24): 1-12. |
| [39] |
吴其超, 臧凤岐, 李呈呈, 马燕, 高燕, 郑勇奇, 臧德奎. 濒危树种五莲杨种群结构与动态特征. 生态学报, 2021, 41(12): 5016-5025. |
| [40] |
陈倩, 杨晔, 史琛媛, 徐学华. 木兰围场天然次生林优势种群的结构与动态. 应用生态学报, 2017, 28(1): 37-46. |
| [41] |
杨慧, 娄安如, 高益军, 宋宏涛. 北京东灵山地区白桦种群生活史特征与空间分布格局. 植物生态学报, 2007, 31(2): 272-282. |
| [42] |
靳静静, 张钦弟, 毕润成. 山西七里峪白桦种群生活史特征与点格局分析. 生态学杂志, 2014, 33(9): 2316-2321. |
| [43] |
宋萍, 洪伟, 吴承祯, 封磊, 范海兰. 珍稀濒危植物桫椤种群生命过程及谱分析. 应用生态学报, 2008, 19(12): 2577-2582. |
| [44] |
矢佳昱, 韩海荣, 程小琴, 董玲玲, 田慧霞, 蔡锰柯, 康峰峰. 河北辽河源自然保护区油松种群年龄结构和种群动态. 生态学杂志, 2017, 36(7): 1808-1814. |
| [45] |
王泳腾, 黄治昊, 王俊, 张童, 崔国发. 燕山山脉黄檗种群结构与动态特征. 生态学报, 2021, 41(07): 2826-2834. |