 2023, Vol. 43
2023, Vol. 43文章信息
- 李建, 卢世君, 邱礼杉, 庄春晓, 杨芳芳, 赵娟娟
- LI Jian, LU Shijun, QIU Lishan, ZHUANG Chunxiao, YANG Fangfang, ZHAO Juanjuan
- 深圳市建成区入侵植物对草本植物种类及功能多样性的影响
- Effects of invasive plants on species and functional diversity of herbs in the built-up area of Shenzhen, China
- 生态学报. 2023, 43(18): 7507-7522
- Acta Ecologica Sinica. 2023, 43(18): 7507-7522
- http://dx.doi.org/10.20103/j.stxb.202206201740
-
文章历史
- 收稿日期: 2022-06-20
- 网络出版日期: 2023-05-08

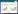




外来入侵物种(IAPs)指“在不同的栖息地和发生范围内, 个体在多个地点分散、生存和繁殖的引入物种”[1]。国家生态环境部2021年发布的《2020中国生态环境状况公报》[2]数据显示:全国已发现660多种外来入侵物种。已有大量研究表明, 入侵植物对被入侵植物群落的结构和生态功能造成了显著影响[3—6], 并且入侵植物对生物多样性的影响具有尺度依赖性, 入侵物种对生物多样性的影响在越大的空间尺度上越小[7]。在植物群落中, 外来入侵植物和其共存的本地植物发挥着不同的作用[8]。
外来入侵物种扩散至一个新的生态系统, 存在着从定殖到扩散逐步建立种群的过程, 在不同的阶段, 其相对丰度、盖度、株高、生物量等入侵特性皆存在明显的差异, 这导致了在入侵群落中表现出不同的入侵程度[9—10]。此外, 当一种入侵植物成功入侵后可以改变被入侵植物群落的环境条件, 从而帮助另一种入侵植物成功入侵其所在的生态系统。因此, 在同一个植物群落中会出现不同入侵者的共同入侵[11—14]。可见, 研究不同入侵程度的单独入侵和共同入侵如何影响群落物种多样性和功能多样性, 对于揭示植物入侵原理至关重要。
城市绿地是城市生物的重要栖息地, 绝大多数都受到人为干扰的影响, 形成了不同的生境类型, 显著增加了入侵植物成功入侵的可能性[15—16]。根据干扰假说和空余生态位假说[17—18], 当生态系统遭受强烈自然或人为干扰后, 本地植物会大量灭绝, 因而群落会产生空余生态位, 进而为外来植物入侵提供机会, 导致外来入侵物种的入侵成功率增加[19]。因此, 城市当中的绿地更容易被外来植物入侵。
物种多样性是生物多样性在物种上的表现形式, 也是生物多样性的关键, 它既体现了生物之间及环境之间的复杂关系, 又体现了生物资源的丰富性[20—21]。功能多样性是指影响群落或生态系统功能的物种性状值和范围, 是生物多样性的体现和重要组成部分, 在解释和预测生态系统结构和功能方面具有重要意义[22]。“植物功能性状”指对植物体定植、存活、生长和死亡存在潜在显著影响的一系列植物属性, 且这些属性能够单独或联合指示生态系统对环境变化的响应, 并且能够对生态系统过程产生强烈影响[23], 可以客观反映植物在异质环境下的适应能力[24]。
虽然已有许多研究对入侵植物的生态影响进行了评估, 但大多仅限于单一入侵植物的生态影响, 例如对加拿大一枝黄花[16, 25]、紫茎泽兰[26]、少花蒺藜草[27]、豚草[28]、喜旱莲子草[29]、黄顶菊[30]等入侵植物的研究中均发现该种植物入侵会改变植物群落物种多样性。城市生态系统中的景观异质性可能导致入侵过程的显著变化, Wang等人研究了加拿大一枝黄花在不同的九种城市生态中对植物群落的物种和功能多样性的影响[16]。对两种入侵植物的共同入侵对植物群落的复合作用及其机理研究较少, 有关学者对两种入侵植物共同入侵造成的群落物种多样性以及功能多样性进行研究, 以探寻两种入侵植物共同入侵时的相互作用[31—32]。
因此, 本研究基于深圳市建成区现场植被调查数据, 选取盖度和频数均排在前列的南美蟛蜞菊和鬼针草两种恶性入侵植物作为研究对象, 结合不同绿地类型, 调查分析南美蟛蜞菊和鬼针草不同程度的单独入侵和共同入侵对草本植物群落物种多样性和功能多样性的影响, 以期为揭示相关影响机制、防控城市入侵植物危害提供一定的科学依据。南美蟛蜞菊和鬼针草均属于我国入侵植物中种数最多的菊科[33]。根据《中国外来入侵植物名录》[34], 鬼针草入侵等级为1级, 即恶性入侵种;南美蟛蜞菊入侵等级为2级, 即严重入侵种。鬼针草(Bidens pilosa)隶属菊科(Asteraceae)鬼针草属(Bidens), 为一年生草本, 广泛分布于热带和亚热带区域, 常见于农田和路边。鬼针草原产于热带美洲, 其生命周期短、结实率高、萌发率高、繁殖速度快、生态适应性强等, 会对当地经济和生态效益造成巨大损失和严重影响[35—36]。南美蟛蜞菊(Sphagneticola trilobata)为菊科(Asteraceae)蟛蜞菊属(Sphagneticola)。原产于热带美洲, 其生长迅速, 无性繁殖能力极强, 易形成覆盖植被。且具有较强的化感作用, 可抑制其他一些作物和杂草的生长, 有很强的侵占性, 对生态系统的结构有重要影响[37]。
1 研究区域深圳市位于广东省南部, 介于113°43′—114°38′E, 22°24′—22°52′N之间, 珠江口东岸, 是粤港澳大湾区四大中心城市之一。深圳陆域整体形状为南北窄、东西宽的狭长形, 且地势由东南至西北逐渐降低, 由北向南分为谷底丘陵、山脉海岸和海湾半岛3个地貌带, 且地面坡度较为和缓, 地貌类型丰富[38]。其建成区面积为923.2km2, 属亚热带海洋性气候, 夏季高温多雨, 冬季干燥稍冷, 地带性植被属于南亚热带季风常绿阔叶林, 雨季集中在每年的4—9月份, 雨量充足, 年均降雨量1935.8mm;年平均气温23℃[39]。深圳市下辖福田区、罗湖区、南山区、盐田区、宝安区、龙岗区、坪山区、龙华区、光明区和大鹏新区[40]。本研究所涉及范围为深圳市的建成区。
2 研究方法 2.1 调查方法采用电脑随机抽样为主, 结合6个公园的判断抽样, 于2019年7—8月对深圳市建成区进行植物调查。首先, 用100m×100m等距网格覆盖深圳市的SPOT 5遥感图像, 再从单元格中电脑随机抽样。另外, 对于深圳市最重要和富有特色的6个公园, 基于面积成比例原则增加取样面积, 以提高调查结果的代表性。共调查1hm2样地600个。对样地内各绿地斑块分别进行编号和调查, 其中每个斑块作为一个乔灌样地, 对乔灌样地调查每个物种。在每个乔灌样地中按面积设置草本植物样方3—5个, 调查草本样方中所有草本植物种类。选取草本样方时, 主要选取斑块中物种较丰富的典型群落进行抽样调查, 调查样方量采用物种数累积法确定。当新样方调查到的物种数小于样地斑块中物种数的10%时, 达到足够的取样量[41]。对草本样方中的每个植物种测量株数、均高, 并估算盖度。入侵植物的确定参考《中国入侵植物名录》[34]。
2.2 数据处理 2.2.1 植物入侵样方选取基于调查数据, 分析了鬼针草和南美蟛蜞菊单独入侵或共同入侵对样方内其他植物的物种多样性及功能多样性的影响。基于前人多项研究所用等级划分方法的总结和改进[25, 42], 根据入侵植物在入侵植物群落的盖度, 将其入侵程度分为轻度(< 35%)、中度(35%—75%)和重度(>75%)。一共调查了1m×1m草本样方2178个, 其中鬼针草单独入侵的样方826个, 南美蟛蜞菊单独入侵的样方423个, 鬼针草和南美蟛蜞菊共同入侵的样方120个(图 1), 未受到鬼针草和南美蟛蜞菊入侵的样方809个。参考我国住房和城乡建设部《城市绿地分类标准(CJJ/T 85—2017)》, 把调查绿地划分为六种类型(表 1)。

|
| 图 1 鬼针草和南美蟛蜞菊的分布 Fig. 1 The distribution of Bidens pilosa and Sphagneticola trilobata |
| 序号 Number |
绿地类型 Green space type |
对应国家标准绿地类型[43] Green space type referenced to national standard[43] |
内容 Content |
绿地面积 Green space area/m2 |
样方数 Samples number |
| Ⅰ | 其他附属绿地 | 公共管理与功能服务设施用地附属绿地 | 公共管理与功能服务设施用地内的绿地 | 138509.2 | 282 |
| 商业服务业设施用地附属绿地 | 商业服务业设施用地内的属绿地 | ||||
| 工业用地附属绿地 | 工业用地内的绿地 | ||||
| 物流仓储用地附属绿地 | 物流仓储用地内的绿地 | ||||
| 公用设施用地附属绿地 | 公用设施用地内的绿地 | ||||
| Ⅱ | 道路绿地 | 道路与交通设施用地附属绿地 | 道路与交通设施用地内的绿地 | 328500.4 | 674 |
| Ⅲ | 公园绿地 | 公园绿地 | 向公众开放, 以游憩为主要功能, 兼具生态、景观、文教和应急避险等功能, 有一定游憩和服务设施的绿地 | 718187.8 | 821 |
| Ⅳ | 广场绿地 | 广场绿地 | 以游憩、纪念、集会和避险等功能为主的城市公共活动场地 | 19943.7 | 70 |
| Ⅴ | 荒地 | / | 没有明确用途, 荒废、弃置的土地 | 49104.0 | 130 |
| Ⅵ | 居住区绿地 | 居住区用地附属绿地 | 居住用地内的配套绿地 | 168762.9 | 201 |
本研究共选取8个功能性状特征[41](表 2), 包括植物的形态特征(株高、叶质、果实类型), 植物的生殖特征(花期、果期、传粉方式、种子的扩散方式), 植物生理特征(固氮类型)。蕨类植物繁殖方式特殊, 故本研究将所有以孢子繁殖的植物类型剔除。
| 功能特征 Functional traits |
数据类型 Data type |
数值 Value |
相关生理生态功能 Relative physiological and ecological functions |
确定方法 Determination method |
| 植株高度 Plant height |
数值数据 | 以厘米(cm)为单位的具体高度 | 表征物种间的竞争力, 物种对环境的适应能力[44], 对资源的利用速度和竞争力[45]。 | 野外调查测量 |
| 叶质 Leaf texture |
名称数据 | 1.革质;2.纸质;3.草质;4.膜质;5.肉质 | 表征物种的竞争能力, 对环境的适应能力[45]。 | 查阅《中国植物志》、文献期刊等获得;若未有明确记录, 根据物种图片确定。 |
| 果实类型 Fruit type |
名称数据 | 1.闭果;2.裂果;3.肉质果 | 表征资源的利用速率[46]。 | 查阅《中国植物志》、文献期刊等 |
| 花期 Florescence |
名称数据 | 1.短(4个月以下);2.中(4—8个月);3.长(8个月及以上) | 表征物种的竞争能力, 对环境的适应能力[45, 47]。 | 查阅《中国植物志》、文献期刊等 |
| 果期 Fruiting stage |
名称数据 | 1.短(4个月以下);2.中(4—8个月);3.长(8个月及以上) | 表征物种的竞争能力, 对环境的适应能力[45, 47]。 | 查阅《中国植物志》、文献期刊等 |
| 传粉方式 Pollination mode |
名称数据 | 1.风媒;2.虫媒 | 表征物种对环境的适应能力[45, 47]。 | 查阅《中国植物志》、文献期刊等获得。若未有明确记录, 依据花的特征确定。花被片明显, 花大而艳丽, 有香气的是虫媒花植物;花小, 雄蕊多数, 无香味的是风媒花植物[48]。 |
| 种子的扩散方式 Seeds dispersal mode |
名称数据 | 1.风力传播;2.动物传播;3.自体传播;4.混合传播 | 表征物种在空间和时间上的扩散能力[47]。 | 查阅文献期刊等获得。若未有明确记录, 根据果实性状特征(尤其是影响传播的典型特征)、所在科属与传播机制的关系确定。有2种及以上传播方式的植物, 列为混合传播型植物。 |
| 固氮类型 Nitrogen fixation type |
名称数据 | 1.固氮;2.不固氮 | 表征植物对氮的利用能力, 生态系统的恢复能力和抵抗能力。 | 查阅《中国植物志》、文献期刊等 |
(1)α多样性指数
通过统计分析获得每个草本样方的物种丰富度指数、生物多样性指数, 优势度指数和均匀度指数。用Margalef丰富度指数表征物种丰富度(F);用Shannon-Wiener多样性指数表征物种多样性(H′);用Simpson优势度指数表征物种优势度(1-D)用Pielou均匀度指数表征物种分布的均匀程度(J)[49—50]。公式如下:
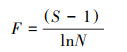
|
(1) |

|
(2) |

|
(3) |

|
(4) |
式中, S是调查样方内所有物种种类的数量。Pi为调查样方内物种i的相对盖度, 即Pi=ni/N, ni是调查样方内物种i的盖度, N是调查样方内所有物种的盖度。
(2) 物种多样性影响幅度
入侵植物对植物物种多样性影响幅度[Influence amplitude index of alien invasive plants on plant taxonomic diversity (PTD), IAIPTD]

|
(5) |
式中, IAIS表示入侵植物对植物种类多样性影响幅度指数[Influence amplitude index of alien invasive plants on species diversity (S), IAIS], IAIH′表示入侵植物对植物Shannon多样性影响幅度指数[Influence amplitude index of alien invasive plants on Shannon′s diversity (H′), IAIH′], IAID表示入侵植物对植物Simpson优势度影响幅度指数[Influence amplitude index of alien invasive plants on Simpson′s dominance (D), IAID], IAIJ表示入侵植物对植物Pielou均匀度影响幅度指数[Influence amplitude index of alien invasive plants on Pielou′s evenness (J), IAIJ], IAIF表示入侵植物对植物Margalef丰富度影响幅度指数[Influence amplitude index of alien invasive plants on Margalef′s richness (F), IAIF]。该指标表征了对物种多样性的影响幅度, 是植物物种多样性生态影响风险的一个评估指标。影响幅度越大, 其对生态系统的影响风险越严重, 越发需要进行防控, 具体计算方式参考王从彦等人的专利[51]与文章[52]。
(3) 功能多样性的计算方法
功能多样性主要通过功能丰富度指数(FRic, 公式6)、功能均匀度指数(FEve, 公式7)、二次熵指数(Rao′s quadratic entropy, FDQ, 公式10)[20]以及群落的特征加权平均值(CWMi, 公式12)[53]
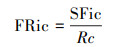
|
(6) |
式中, 物种在群落中所占的生态位用SFic表示, 即每个物种的不同功能性状;性状c在群落中的绝对特征值大小用Rc表示。

|
(7) |
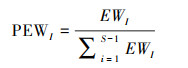
|
(8) |

|
(9) |
式中, EW表示均匀度, dist(i, j)表示物种i和j的欧式距离, wi表示i物种的数目。
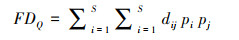
|
(10) |
式中, dij为物种i与物种j的功能特征距离, pi和pj分别是一个特定样方中植物物种i和j的相对丰度。
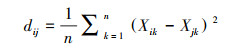
|
(11) |
式中, Xik和Xjk分别代表植物物种i和j在一个特定样方中的功能性状k, n代表功能性状的数量。
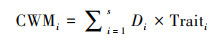
|
(12) |
式中, CWMi表示所测物种i功能性状的群落加权平均值, Di表示物种i的相对多度, Traiti表示物种i的平均功能性状值。
2.2.4 统计分析利用Excel 2019对数据进行整理, 采用SPSS 26.0对各指标进行统计分析, 并计算平均值和均值偏差(Mean±SD)。通过单因素方差分析(One-way ANOVA), 基于LSD(最小显著差数法)分析方法确定不同入侵情况下植物物种多样性和功能多样性的差异水平, 显著差异水平设置为P<0.05。物种多样性用PAST软件进行计算。使用R4.1.3计算功能多样性。线性拟合分析与作图均由Origin 2021完成。
3 结果与分析 3.1 物种多样性及功能多样性随入侵植物盖度的变化关系鬼针草和南美蟛蜞菊的单独及共同入侵都显著影响了物种多样性指数(P < 0.05), Simpson优势度指数、Shannon-Winner多样性指数、Margalef丰富度指数以及Pielou均匀度指数均随入侵程度增加而下降(图 2)。在相同的入侵程度下, 物种多样性指数是南美蟛蜞菊单独入侵 < 鬼针草单独入侵 < 两者共同入侵。物种多样性指数与入侵程度的线性拟合方程R2值均在0.5865以上, 最大值达到0.9356。各项物种多样性指数的拟合结果R2值, 都是各自单独入侵高于两者共同入侵, 且除了Margalef丰富度指数以外其他多样性指数均是鬼针草单独入侵高于南美蟛蜞菊单独入侵。

|
| 图 2 不同入侵植物盖度下草本植物群落物种多样性指数的变化趋势及线性拟合结果 Fig. 2 Trends and linear fitting results of species diversity indexes of herbaceous plant community under different coverage of invasive plants |
但是, 这两种植物的入侵对群落功能多样性的影响并非全部显著(图 3)。其中, 鬼针草单独入侵以及鬼针草和南美蟛蜞菊共同入侵显著降低了群落功能丰富度FEve(P < 0.05)。随着鬼针草入侵程度的增加, 功能丰富度指数FRic略有提高, 功能均匀度指数FEve下降, 功能离散度指数FDQ提高, 群落加权平均株高也显著增加(P < 0.05)。南美蟛蜞菊单独入侵的草本植物群落中, 功能多样性指数FRic、功能丰富度指数FEve以及功能离散度指数FDQ均与其入侵程度呈负相关(0.0666 < R2 < 0.2211), 而群落加权平均株高与其入侵程度呈正相关(R2=0.0772)。鬼针草和南美蟛蜞菊共同入侵的草本植物群落中, 功能丰富度指数FRic、功能离散度指数FDQ以及群落加权平均株高随着入侵程度的增加而提高, 功能均匀度指数FEve则随着入侵程度的增加而下降。

|
| 图 3 不同入侵植物盖度下草本植物群落功能多样性指数的变化趋势及线性拟合结果 Fig. 3 Trends and linear fitting results of functional diversity indexes of herbaceous plant community under different coverage of invasive plants |
入侵植物对群落植物分类多样性的影响幅度随其盖度增加显著提高(P < 0.05)(图 4)。且在相同盖度下, 影响幅度以南美蟛蜞菊单独入侵>鬼针草单独入侵>两者共同入侵。当影响幅度大于0时, 入侵植物开始对群落产生生态风险。但鬼针草和南美蟛蜞菊开始对物种多样性产生风险时的入侵程度(即盖度)不同(图 4), 且南美蟛蜞菊单独入侵更易对草本植物群落产生生态风险。鬼针草和南美蟛蜞菊两者对植物分类多样性影响幅度的最高值在0.35—0.75之间, 因此二者皆可归类为严重入侵植物, 其生态影响风险较高。

|
| 图 4 不同入侵情况对草本植物群落植物分类多样性的影响幅度及其线性拟合结果 Fig. 4 Influence amplitude of different invasion situations on taxonomic diversity of herbaceous plant communities and the linear fitting results |
利用单因素方差分析比较了不同入侵情况下本地草本植物群落物种多样性的差异性(图 5)。结果显示, 不同入侵情况下各物种多样性指数均为轻度入侵>中度入侵>重度入侵。轻度入侵下二者分别单独入侵或共同入侵均显著提高了物种多样性指数(P < 0.05), 只有南美蟛蜞菊轻度入侵对Margalef丰富度指数未达到显著水平。鬼针草和南美蟛蜞菊单独入侵的重度入侵显著降低了物种多样性指数(P < 0.05), 两者共同入侵在重度入侵时也降低了物种多样性指数, 但是仅Pielou指数达到显著性水平(P < 0.05)。各种入侵程度下, 共同入侵的物种多样性指数均高于单独入侵。

|
| 图 5 不同入侵程度对草本植物群落物种多样性的影响 Fig. 5 Effects of different invasion degrees on the species diversity of herbaceous plant community 不同入侵情况和未入侵比较, 不同入侵情况之间不进行比较; 不同小写字母表示差异显著 |
与未入侵样地相比, 鬼针草轻度入侵显著降低了功能丰富度指数FRic和功能离散度指数FDQ(P < 0.05), 显著提高了功能丰富度指数FEve(P < 0.05), 且功能丰富度指数FRic是轻度入侵>中度入侵>重度入侵。南美蟛蜞菊重度入侵显著降低了功能丰富度指数FEve(P < 0.05)。鬼针草和南美蟛蜞菊单独入侵或共同入侵下均显著提高了群落的加权平均株高(P < 0.05), 且群落加权平均株高是重度入侵>中度入侵>轻度入侵。其余结果均不显著(P>0.05)(图 6)。

|
| 图 6 不同入侵程度对草本植物群落功能多样性的影响 Fig. 6 Effects of different invasion degrees on the functional diversity of herbaceous plant communities 不同入侵情况和未入侵比较, 不同入侵情况之间不进行比较。不同小写字母表示差异显著 |
从表 3可以看出, 与未被入侵的群落相比, 各绿地类型中, 鬼针草单独入侵或两者共同入侵均显著提高了各物种多样性指数(P < 0.05)。南美蟛蜞菊入侵则降低了各绿地类型的各物种多样性指数, 但只在其他附属绿地和道路绿地达到显著水平(P < 0.05)。在未被入侵的植物群落中, 各绿地类型的物种多样性指数均以居住区绿地最高(表 3)。在鬼针草单独入侵的样方中, 各绿地类型的物种多样性指数均以广场绿地最低, 其他附属绿地最高(表 3)。结合以上分析表明鬼针草在广场绿地入侵程度较为严重, 在其他附属绿地入侵程度较轻。在南美蟛蜞菊入侵的样方中, 各绿地类型的物种多样性指数均以荒地为最高, 道路绿地为最低。结合以上分析表明南美蟛蜞菊在道路绿地入侵程度较为严重, 在荒地入侵程度较轻。在鬼针草和南美蟛蜞菊共同入侵的样方中, 荒地的物种多样性指数最高。
| 绿地类型 Type of green space |
入侵植物 Invasive plants |
Simpson 优势度指数 Simpson′s dominance index |
Shannon-Wiener 多样性指数 Shannon-Wiener′s diversity index |
Margalef 丰富度指数 Margalef′s richness index |
Pielou 均匀度指数 Pielou′s evenness index |
| 其他附属绿地 | 无 | 0.61±0.07a | 1.16±0.31a | 0.91±0.5a | 0.25±0.07a |
| Other ancillary green Spaces | 鬼针草 | 0.66±0.05b | 1.38±0.27b | 1.21±0.55b | 0.29±0.06b |
| 南美蟛蜞菊 | 0.58±0.06b | 1.00±0.26b | 0.65±0.42b | 0.2±0.06b | |
| 鬼针草+南美蟛蜞菊 | 0.66±0.05b | 1.32±0.24b | 1.05±0.41b | 0.27±0.07b | |
| 道路绿地 | 无 | 0.61±0.07a | 1.13±0.32a | 0.86±0.53a | 0.25±0.08a |
| Road green space | 鬼针草 | 0.64±0.06b | 1.29±0.32b | 1.08±0.58b | 0.27±0.07b |
| 南美蟛蜞菊 | 0.57±0.07b | 0.96±0.27b | 0.57±0.41b | 0.2±0.07b | |
| 鬼针草+南美蟛蜞菊 | 0.64±0.05b | 1.26±0.25b | 0.96±0.48b | 0.24±0.05b | |
| 公园绿地 | 无 | 0.6±0.07a | 1.08±0.31a | 0.79±0.51a | 0.22±0.07a |
| Park green space | 鬼针草 | 0.64±0.06b | 1.3±0.29b | 1.13±0.56b | 0.26±0.06b |
| 南美蟛蜞菊 | 0.59±0.06a | 1.03±0.24a | 0.64±0.38a | 0.21±0.06a | |
| 鬼针草+南美蟛蜞菊 | 0.63±0.06b | 1.22±0.27b | 0.93±0.46b | 0.25±0.07b | |
| 广场绿地 | 无 | 0.59±0.07a | 1.04±0.27a | 0.69±0.42a | 0.23±0.07a |
| Square green space | 鬼针草 | 0.64±0.05b | 1.26±0.25b | 0.94±0.36b | 0.26±0.06b |
| 南美蟛蜞菊 | — | — | — | — | |
| 鬼针草+南美蟛蜞菊 | — | — | — | — | |
| 荒地 Wasteland | 无 | 0.6±0.08a | 1.14±0.39a | 0.88±0.62a | 0.24±0.09a |
| 鬼针草 | 0.65±0.06b | 1.3±0.31b | 1.05±0.55b | 0.27±0.07b | |
| 南美蟛蜞菊 | 0.59±0.08a | 1.06±0.34a | 0.75±0.49a | 0.22±0.07a | |
| 鬼针草+南美蟛蜞菊 | 0.66±0.05b | 1.35±0.24b | 1.11±0.48b | 0.27±0.06b | |
| 居住区绿地 | 无 | 0.62±0.07a | 1.18±0.34a | 0.91±0.54a | 0.25±0.08a |
| Residential green space | 鬼针草 | 0.65±0.06b | 1.34±0.29b | 1.16±0.54b | 0.29±0.07b |
| 南美蟛蜞菊 | 0.57±0.1a | 0.97±0.4a | 0.58±0.52a | 0.21±0.11a | |
| 鬼针草+南美蟛蜞菊 | — | — | — | — | |
| 仅在各绿地类型中进行显著性比较, 同一列中, 不同小写字母表示在0.05水平上显著相关; “—”表示未在此绿地类型中调查到研究对象的样方, 或调查到该入侵植物的样方数太少不具有参考性 | |||||
从表 4可以看出, 鬼针草入侵只在其他附属绿地中显著降低了功能丰富度指数FRic和功能离散度指数FDQ(P < 0.05), 鬼针草和南美蟛蜞菊单独入侵分别在其他附属绿地和广场绿地中显著提高了功能丰富度指数FEve(P < 0.05), 其余均不显著(P>0.05)。在不同绿地类型中, 各种入侵情况的样地群落加权平均株高均比未入侵高, 其中广场绿地中鬼针草和公园绿地及道路绿地以外各种绿地类型中的南美蟛蜞菊入侵结果均不显著(P>0.05), 其余均显著(P < 0.05)。不同的入侵类型中, 道路绿地的功能丰富度指数FRic和Rao二次熵指数FDQ均为最高。这说明道路绿地群落中的物种数目较多, 功能性状分布的范围较广, 物种占据的功能生态位较大。
| 绿地类型 Type of green space |
入侵植物 Invasive plants |
功能丰富度指数 FRic Functional richness index |
功能均匀度指数 FEve Functional eveness index |
Rao 二次熵指数 Rao′s quadratic entropy index |
群落加权平均株高 Community-Weighted mean height |
| 其他附属绿地 | 无 | 1.57±4.23a | 0.67±0.16a | 1.26±2.07a | 17.54±8.94a |
| Other ancillary green spaces | 鬼针草 | 0.26±4.54b | 0.73±0.13b | 0.21±0.83b | 24.6±10.91b |
| 南美蟛蜞菊 | 0.7±4.13a | 0.67±0.23a | 1.06±2.11a | 18.4±2.47a | |
| 鬼针草+南美蟛蜞菊 | 1.3±4.49a | 0.72±0.16a | 1.27±2.03a | 25.09±7.23b | |
| 道路绿地 | 无 | 4.23±0.3a | 0.64±0.18a | 3.87±2.15a | 15.53±8.95a |
| Road green space | 鬼针草 | 4.54±0.32a | 0.65±0.17a | 4.14±2.18a | 24.55±11.88b |
| 南美蟛蜞菊 | 4.13±0.55a | 0.61±0.18a | 3.99±2.11a | 21.24±7.91b | |
| 鬼针草+南美蟛蜞菊 | 4.49±0.83a | 0.64±0.17a | 4.87±2.13a | 29.33±10.54b | |
| 公园绿地 | 无 | 0.3±0.49a | 0.68±0.18a | 0.29±1.16a | 17.43±11.23a |
| Park green space | 鬼针草 | 0.32±0.71a | 0.7±0.16a | 0.26±0.96a | 20.75±15.72b |
| 南美蟛蜞菊 | 0.55±0.04a | 0.72±0.17a | 0.51±1.4a | 25.43±14.82b | |
| 鬼针草+南美蟛蜞菊 | 0.83±0.09a | 0.7±0.16a | 0.65±1.95a | 20.29±8.76b | |
| 广场绿地 | 无 | 0.49±0.57a | 0.67±0.16a | 0.43±1.07a | 20.26±10.77a |
| Square green space | 鬼针草 | 0.71±0.4a | 0.61±0.22a | 0.37±0.9a | 22.58±9.98a |
| 南美蟛蜞菊 | — | — | — | — | |
| 鬼针草+南美蟛蜞菊 | — | — | — | — | |
| 荒地 Wasteland | 无 | 0.57±0.58a | 0.69±0.18a | 0.42±1.09a | 19.41±13.78a |
| 鬼针草 | 0.4±0.28a | 0.72±0.14a | 0.4±1.54a | 37.89±18.79b | |
| 南美蟛蜞菊 | 0.09±1.91a | 0.64±0.15a | 0.04±0.01a | 19.78±4.84a | |
| 鬼针草+南美蟛蜞菊 | 0.09±0a | 0.73±0.17a | 0.03±0.02a | 29.38±11.88b | |
| 居住区绿地 | 无 | 0.58±0.8a | 0.72±0.15a | 0.55±1.38a | 16.68±8.42a |
| Residential green space | 鬼针草 | 0.28±0.09a | 0.72±0.14a | 0.21±1.11a | 20.15±6.6b |
| 南美蟛蜞菊 | 1.91±0.1a | 0.76±0.17a | 1.73±2.41a | 16.93±2.19a | |
| 鬼针草+南美蟛蜞菊 | — | — | — | — | |
| 仅在各绿地类型中进行显著性比较, 同一列中, 不同小写字母表示在0.05水平上显著相关; “—”表示未在此绿地类型中调查到研究对象的样方, 或样方数太少不具有参考性 | |||||
本研究发现入侵植物对城市草本植物群落的物种多样性产生显著影响。鬼针草和南美蟛蜞菊分别单独入侵和共同入侵的三种状态下, 草本植物群落的四个物种多样性指数均与入侵植物盖度(即入侵程度)呈显著负相关(P < 0.05), 且入侵程度越高则多样性指数越低, 各物种多样性指数均表现为轻度入侵>中度入侵>重度入侵。Wang等人对加拿大一枝黄花(Solidago canadensis)的研究也发现, 随着被入侵植物群落中加拿大一枝黄花入侵程度的增加, 植物多样性、优势度、丰富度和群落稳定性显著下降[25]。张修玉等人对紫茎泽兰(Ageratina adenophora)(破坏草)的研究也得出了相一致的结论[26]。王坤芳等人也发现少花蒺藜草(Ageratina adenophora)入侵地植物多样性随入侵程度加重而降低[27]。可能因为入侵植物与其共存的植物相比具有更强的竞争力, 因此其成功定植后, 种群密度和生物量逐渐增加, 盖度逐渐增大, 使得共存的其他物种生存空间减小, 群落物种多样性开始持续下降[27—28]。
然而, 不同的入侵程度对草本植物群落的影响不同。与未被入侵的草本植物群落相比, 虽然鬼针草和南美蟛蜞菊单独重度入侵时导致草本植物群落物种多样性显著降低(P < 0.05), 但是轻度入侵反而使其显著升高(P < 0.05)。同样地, 两者共同入侵的群落中, 重度入侵降低了物种多样性指数, 但只有Pielou均匀度指数达到显著水平(P < 0.05), 而轻度入侵则显著提高了各物种多样性指数(P < 0.05)。王从彦等人对加拿大一枝黄花入侵群落的研究也报道了基本一致的结果[3, 25, 54]。柳晓燕等人同样发现豚草(Ambrosia artemisiifolia)入侵群落入侵初期具有较高的本地物种多样性, 随后随着入侵程度增大, 物种丰富度和Shannon-Wiener多样性指数开始降低[28]。另外, Wu等[55]发现, 喜旱莲子草(Alternanthera philoxeroides)轻度入侵会增加群落物种丰富度和均匀度, 而重度入侵则显著降低本地植物物种多样性。据分析, 轻度入侵增加物种多样性、丰富度、均匀度和优势度, 可能主要是由乘客效应(passenger effects)驱动的, 而不是入侵植物的存在所介导的驱动效应[3, 8, 56]。本文中共同和单独入侵对群落物种多样性指数的影响基本一致, 可能跟本文两种类型的入侵都采用入侵植物盖度来划分入侵程度有关。
植物入侵对物种多样性的影响还跟入侵植物的种类有关。在相同的入侵程度(即盖度)下, 物种多样性指数以南美蟛蜞菊单独入侵 < 鬼针草单独入侵 < 两者共同入侵, 且对物种多样性的影响幅度以两者共同入侵 < 鬼针草单独入侵 < 南美蟛蜞菊入侵。被这两种入侵植物入侵的样方比单独被鬼针草、南美蟛蜞菊入侵的地块有更高的物种多样性指数。这意味着鬼针草和南美蟛蜞菊的入侵影响不是相加的——相反, 当这两种入侵物种同时出现时, 对群落物种多样性的入侵影响似乎减少了。与两者单独入侵相比, 共同入侵对群落物种多样性的影响降低了。不同入侵植物种类共同入侵时, 它们之间的相互作用可能是协同作用, 亦或是拮抗作用[57]。协同作用即为两种入侵植物共同入侵的复合影响大于两者单一入侵的影响;而拮抗作用则是两种入侵植物共同入侵的复合影响小于两者单一入侵的影响[58]。该结果表明鬼针草和南美蟛蜞菊之间共同入侵产生的生态效应可能为拮抗作用, 这与之前对波斯胡桃和加拿大一枝黄花的共同入侵研究结果一致[59]。
不分入侵程度的入侵植物总体分析得到了不同的结果。本文在分不同绿地类型的入侵影响分析中, 未划分入侵程度, 将不同入侵程度的样方统一作为被入侵样方进行分析。结果发现, 鬼针草单独入侵和与南美蟛蜞菊两者共同入侵均显著提高了本地草本植物群落的物种多样性指数(P < 0.05), 而南美蟛蜞菊单独入侵则降低了各物种多样性指数。与分不同入侵程度时的结论有差异。其可能的原因是在所调查的绿地中, 鬼针草单独入侵主要处于轻度入侵阶段, 而南美蟛蜞菊大都处于中度至重度入侵。
另外, 植物群落功能多样性指数也与入侵程度有一定的相关关系, 但是线性拟合相关性低(R2 < 0.2262), 且不同功能多样性指标随入侵程度的变化规律不同。在三种不同的入侵模式下, 草本植物群落的功能均匀度指数FEve随着盖度增加而降低, 群落加群平均株高(CWMH)则相反。而群落功能丰富指数FRic和功能离散度指数FDQ随盖度的变化规律不同, 且结果均不显著(P>0.05)。功能均匀度反映了物种功能性状分布的均匀程度[60]。本研究中功能均匀度下降, 说明随着入侵植物盖度的增加, 群落中功能性状分布开始集中。群落加权平均株高的上升证实了这一结论。
研究结果显示, 不论是未划分入侵程度的各种绿地类型, 还是划分入侵程度的城市绿地, 鬼针草和南美蟛蜞菊单独入侵或共同入侵对群落功能丰富度指数(FRic)、功能均匀度指数(FEve)、二次熵指数(FDQ)影响结果大都不显著。入侵植物可能占据了某一生态位, 顶替了原本植物的生态功能, 所以对群落功能多样性影响不显著。然而入侵植物群落的加权平均株高在各种绿地类型中均高于其他植物, 且这种差距随着其入侵程度的增加而增大, 这与之前的研究结果一致[3, 61]。株高在植物资源获取和物种竞争能力方面起着至关重要的作用, 尤其是对有限的光资源竞争方面[62—63]。株高的差异可能反映了这一群体内竞争能力的差异, 并可能导致群落内的光照分配发生变化[64]。入侵植物与本地植物在群落加权平均株高上的这种差异可能是其成功入侵的重要驱动力之一[65]。
本研究主要研究鬼针草和南美蟛蜞菊这两种深圳市最主要的入侵植物对草本植物种类及功能多样性的影响作用, 因此没有计算其他入侵植物的影响。在未来的研究中可进一步研究其他入侵植物的影响, 及其跟本文两种入侵植物的联合影响作用。在功能性状的选取上考虑了跟入侵植物相关的部分生理、营养特征, 一定程度反映了入侵草本植物群落的生态功能多样性。主要选用的是分类数据。未来研究中可加入更多的数值型功能特征以更准确地反映入侵植物群落的功能多样性。例如, 叶长、叶宽、叶绿素浓度等数值型数据都和植物利用光的能力有关, 可以用于表征入侵植物的入侵能力[16]。
城市生态系统中各种因子及其相互关系错综复杂[66—67], 限于可操作性、时间和经费的限制、相关生态过程和机制的不确定性等, 目前尚无法在纳入所有影响因素的情况下, 计算入侵因子的影响作用, 以达到最高的准确性。因此参考前人的方法[68—69], 在无法考虑所有因素的情况下, 本文把重点放在关键入侵物种单独入侵和共同入侵可能带来的影响上面, 通过同时采用相关性分析和变化曲线来揭示其中的相关关系和变化规律, 并通过大范围、大面积的大样本量取样, 提高样本总体对整个城市各种人为影响情况的代表性, 各公园抽样调查面积不小于公园总面积的15%, 使样本总体的整体趋势能更好地代表目前全市人为影响现状下, 入侵植物与多样性间的相关关系, 改善了结果的可信性和说服力。未来的调查和研究中可进一步分析其他影响因素的影响作用, 通过不断探索各种影响因素的作用, 最终为最完整、最准确地揭示相关社会-经济-自然综合因素复杂影响机制提供参考。
5 结论本文调查分析了深圳市建成区不同绿地类型中鬼针草和南美蟛蜞菊分别单独入侵和共同入侵对草本植物群落物种多样性和功能多样性的影响。结果表明, 与未入侵的植物群落相比, 轻度入侵增加了物种多样性、优势度、丰富度和均匀度, 而重度入侵则相反。更重要的是, 随着入侵植物盖度(入侵程度)增加, 物种多样性、优势度、丰富度和均匀度均显著下降(P < 0.05)。且与鬼针草和南美蟛蜞菊单独入侵相比, 两者共存可能更有利于提高植物多样性。入侵程度的上升与群落功能多样性之间有一定的相关关系, 但不太显著。受到入侵的植物群落其群落株高加权平均值会随着入侵程度的增加而升高。这说明入侵植物的入侵程度对群落物种多样性起着至关重要的作用。同时, 植物的株高这一功能性状可能是其成功入侵的重要机制。本研究结果为进一步了解鬼针草和南美蟛蜞菊单独入侵及共同入侵对群落造成的影响提供一定的理论支撑, 为有效防治入侵提供一定的参考, 具有一定的理论和实践意义。
| [1] |
Blackburn T M, Pyšek P, Bacher S, Carlton J T, Duncan R P, Jarošík V, Wilson J R U, Richardson D M. A proposed unified framework for biological invasions. Trends in Ecology & Evolution, 2011, 26(7): 333-339. |
| [2] |
生态环境部. 2020年中国生态环境状况公报(摘录). 环境保护, 2021, 49(11): 47-68. DOI:10.14026/j.cnki.0253-9705.2021.11.010 |
| [3] |
Wang C Y, Jiang K, Liu J, Zhou J W, Wu B D. Moderate and heavy Solidago canadensis L. invasion are associated with decreased taxonomic diversity but increased functional diversity of plant communities in East China. Ecological Engineering, 2018, 112: 55-64. DOI:10.1016/j.ecoleng.2017.12.025 |
| [4] |
Martín-Forés I, Guerin G R, Lowe A J. Weed abundance is positively correlated with native plant diversity in grasslands of southern Australia. PLoS One, 2017, 12(6): e0178681. DOI:10.1371/journal.pone.0178681 |
| [5] |
Hejda M, Pyšek P, Jarošík V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. Journal of Ecology, 2009, 97(3): 393-403. DOI:10.1111/j.1365-2745.2009.01480.x |
| [6] |
Clay K. Non-native grass invasion alters native plant composition in experimental communities. Biological Invasions, 2010, 12(5): 1285-1294. DOI:10.1007/s10530-009-9546-9 |
| [7] |
Powell K I, Chase J M, Knight T M. Invasive plants have scale-dependent effects on diversity by altering species-area relationships. Science, 2013, 339(6117): 316-318. DOI:10.1126/science.1226817 |
| [8] |
Bernard-Verdier M, Hulme P E. Alien and native plant species play different roles in plant community structure. Journal of Ecology, 2015, 103(1): 143-152. DOI:10.1111/1365-2745.12341 |
| [9] |
Hooper D U, Dukes J S. Functional composition controls invasion success in a California serpentine grassland. Journal of Ecology, 2010, 98(4): 764-777. DOI:10.1111/j.1365-2745.2010.01673.x |
| [10] |
Wilson S D, Pinno B D. Environmentally-contingent behaviour of invasive plants as drivers or passengers. Oikos, 2013, 122(1): 129-135. DOI:10.1111/j.1600-0706.2012.20673.x |
| [11] |
Kuebbing S E, Classen A T, Simberloff D. Two co-occurring invasive woody shrubs alter soil properties and promote subdominant invasive species. Journal of Applied Ecology, 2014, 51(1): 124-133. DOI:10.1111/1365-2664.12161 |
| [12] |
Kuebbing S E, Patterson C M, Classen A T, Simberloff D. Co-occurring nonnative woody shrubs have additive and non-additive soil legacies. Ecological Applications, 2016, 26(6): 1896-1906. DOI:10.1890/15-1931.1 |
| [13] |
Wang C Y, Zhou J W, Liu J, Jiang K, Xiao H G, Du D L. Responses of the soil fungal communities to the co-invasion of two invasive species with different cover classes. Plant Biology: Stuttgart, Germany, 2018, 20(1): 151-159. DOI:10.1111/plb.12646 |
| [14] |
Wei M, Wang S, Xiao H, Wu B D, Jiang K, Wang C Y. Co-invasion of daisy fleabane and Canada goldenrod pose synergistic impacts on soil bacterial richness. Journal of Central South University, 2020, 27(6): 1790-1801. DOI:10.1007/s11771-020-4408-9 |
| [15] |
El-Barougy R F, Cadotte M W, Khedr A H A, Nada R M, Maclvor J S. Heterogeneity in patterns of survival of the invasive species Ipomoea carnea in urban habitats along the Egyptian Nile Delta. NeoBiota, 2017, 33: 1-17. DOI:10.3897/neobiota.33.9968 |
| [16] |
Wang C Y, Wu B D, Jiang K, Zhou J W, Du D L. Canada goldenrod invasion affect taxonomic and functional diversity of plant communities in heterogeneous landscapes in urban ecosystems in East China. Urban Forestry & Urban Greening, 2019, 38: 145-156. |
| [17] |
Hierro J L, Maron J L, Callaway R M. A biogeographical approach to plant invasions: the importance of studying exotics in their introduced and native range. Journal of Ecology, 2005, 93(1): 5-15. DOI:10.1111/j.0022-0477.2004.00953.x |
| [18] |
Sax D F, Gaines S D, Brown J H. Species invasions exceed extinctions on Islands worldwide: a comparative study of plants and birds. The American Naturalist, 2002, 160(6): 766-783. DOI:10.1086/343877 |
| [19] |
朱金方, 赵彩云, 李飞飞, 柳晓燕, 李俊生. 不同替代修复年限对飞机草种群和本土植物群落结构的影响. 环境科学研究, 2020, 33(12): 2848-2855. DOI:10.13198/j.issn.1001-6929.2020.06.24 |
| [20] |
刘旻霞, 南笑宁, 张国娟, 李博文, 徐璐, 穆若兰, 李亮, 于瑞新. 高寒草甸不同坡向植物群落物种多样性与功能多样性的关系. 生态学报, 2021, 41(13): 5398-5407. |
| [21] |
余轩, 王兴, 吴婷, 王启学, 马昀, 谢莉, 宋乃平. 围封对荒漠草原物种多样性和功能多样性的影响. 水土保持学报, 2021, 35(6): 243-250. DOI:10.13870/j.cnki.stbcxb.2021.06.033 |
| [22] |
韩涛涛, 唐玄, 任海, 王俊, 刘楠, 郭勤峰. 群落/生态系统功能多样性研究方法及展望. 生态学报, 2021, 41(8): 3286-3295. |
| [23] |
刘晓娟, 马克平. 植物功能性状研究进展. 中国科学: 生命科学, 2015, 45(4): 325-339. |
| [24] |
McIntyre S, Lavorel S, Landsberg J, Forbes T D A. Disturbance response in vegetation - towards a global perspective on functional traits. Journal of Vegetation Science, 1999, 10(5): 621-630. DOI:10.2307/3237077 |
| [25] |
Wang S, Wei M, Wu B D, Jiang K, Du D L, Wang C Y. Degree of invasion of Canada goldenrod (Solidago canadensis L.) plays an important role in the variation of plant taxonomic diversity and community stability in Eastern China. Ecological Research, 2019, 34(6): 782-789. DOI:10.1111/1440-1703.12049 |
| [26] |
张修玉, 许振成, 宋巍巍, 黎忠, 赵晓光, 胡习邦. 紫茎泽兰(Eupatorium adenophorum)入侵地的生物多样性. 生态环境学报, 2010, 19(7): 1525-1531. |
| [27] |
王坤芳, 纪明山, 彭爽. 不同入侵程度下少花蒺藜草对植物群落特征影响. 东北农业大学学报, 2015, 46(11): 16-21. DOI:10.19720/j.cnki.issn.1005-9369.2015.11.003 |
| [28] |
柳晓燕, 朱金方, 李飞飞, 赵彩云. 豚草入侵对新疆伊犁河谷林下本地草本植物群落结构的影响. 生态学报, 2021, 41(24): 9613-9620. |
| [29] |
张丽苗, 谭雪, 董智, 郑杰, 袁中勋, 李昌晓. 喜旱莲子草入侵对三峡库区重庆主城河岸带植物多样性的影响. 草业学报, 2022, 31(9): 13-25. |
| [30] |
祁小旭, 张思宇, 林峰, 张玲玲, 杨殿林, 皇甫超河, 王慧. 黄顶菊对不同入侵地植物群落及土壤微生物群落的影响. 生态学报, 2019, 39(22): 8472-8482. |
| [31] |
Wang C Y, Yu Y L, Cheng H Y, Du D L. Which factor contributes most to the invasion resistance of native plant communities under the co-invasion of two invasive plant species?. Science of the Total Environment, 2022, 813: 152628. DOI:10.1016/j.scitotenv.2021.152628 |
| [32] |
Wang C Y, Wei M, Wang S, Wu B D, Cheng H Y. Erigeron annuus (L.) Pers. and Solidago canadensis L. antagonistically affect community stability and community invasibility under the co-invasion condition. Science of the Total Environment, 2020, 716: 137128. DOI:10.1016/j.scitotenv.2020.137128 |
| [33] |
闫小玲, 刘全儒, 寿海洋, 曾宪锋, 张勇, 陈丽, 刘演, 马海英, 齐淑艳, 马金双. 中国外来入侵植物的等级划分与地理分布格局分析. 生物多样性, 2014, 22(5): 667-676. |
| [34] |
马金双, 李惠茹. 中国外来入侵植物名录. 北京: 高等教育出版社, 2018.
|
| [35] |
郝建华, 刘倩倩, 强胜. 菊科入侵植物三叶鬼针草的繁殖特征及其与入侵性的关系. 植物学报, 2009, 44(6): 656-665. |
| [36] |
王桔红, 史生晶, 陈文, 李云, 崔现亮. 鬼针草与含羞草化感作用及其入侵性的研究. 草业学报, 2020, 29(4): 81-91. |
| [37] |
吴彦琼, 胡玉佳, 廖富林. 从引进到潜在入侵的植物——南美蟛蜞菊. 广西植物, 2005, 25(5): 413-418. |
| [38] |
唐瑜聪. 深圳市极端气候时空特征及其灾害风险评估[D]. 北京: 中国地质大学(北京), 2017.
|
| [39] |
张曼琳, 潘妮, 赵娟娟, 李明娟, 江南. 城市花粉致敏植物种类构成、分布与潜在危害评估——以深圳市为例. 生态学报, 2021, 41(22): 8746-8757. |
| [40] |
何海珊, 赵宇豪, 吴健生. 低碳导向下土地覆被演变模拟——以深圳市为例. 生态学报, 2021, 41(21): 8352-8363. |
| [41] |
李明娟, 赵娟娟, 刘时彦, 宋晨晨, 江南. 山地城市公园植物群落功能多样性与物种多样性研究——以重庆市主城区为例. 中国园林, 2021, 37(2): 124-129. |
| [42] |
张羽, 蔡竟芳, 万云, 高海燕, 孙凯, 沈一栾, 阿斯哈, 李红丽. 凤眼莲入侵程度对金鱼藻和黑藻生长及种间关系的影响. 生态学报, 2022, 42(16): 6863-6873. |
| [43] |
中华人民共和国住房和城乡建设部, 《城市绿地分类标准》(CJJ/T85-2017), 2017b.
|
| [44] |
Zhang J T, Li M, Nie E B. Pattern of functional diversity along an altitudinal gradient in the Baihua Mountain Reserve of Beijing, China. Brazilian Journal of Botany, 2014, 37(1): 37-45. |
| [45] |
Venn S E, Green K, Pickering C M, Morgan J W. Using plant functional traits to explain community composition across a strong environmental filter in Australian alpine snowpatches. Plant Ecology, 2011, 212(9): 1491-1499. |
| [46] |
Flynn D F B, Gogol-Prokurat M, Nogeire T, Molinari N, Richers B T, Lin B B, Simpson N, Mayfield M M, DeClerck F. Loss of functional diversity under land use intensification across multiple taxa. Ecology Letters, 2009, 12(1): 22-33. |
| [47] |
Purschke O, Schmid B C, Sykes M T, Poschlod P, Michalski S G, Durka W, Kühn I, Winter M, Prentice H C. Contrasting changes in taxonomic, phylogenetic and functional diversity during a long-term succession: insights into assembly processes. Journal of Ecology, 2013, 101(4): 857-866. |
| [48] |
Friedman J, Barrett S C H. Wind of change: new insights on the ecology and evolution of pollination and mating in wind-pollinated plants. Annals of Botany, 2009, 103(9): 1515-1527. |
| [49] |
马克平, 刘玉明. 生物群落多样性的测度方法Ⅰα多样性的测度方法(下). 生物多样性, 1994, 2(4): 231-239. |
| [50] |
马克平. 生物群落多样性的测度方法Ⅰα多样性的测度方法(上). 生物多样性, 1994, 2(3): 231-239. |
| [51] |
王从彦, 杜道林, 黄萍, 孙见凡, 李健, 戴志聪, 李冠霖, 李俊生, 徐海耕, 韦梅, 王舒, 程慧源, 伍丙德, 于友礼. 入侵植物对植物物种多样性生态影响风险评估量化方法: 中国, 202011171687. 7. 2021-02-02. [2022-5-19]
|
| [52] |
Wang C Y, Cheng H Y, Wang S, Wei M, Du D L. Plant community and the influence of plant taxonomic diversity on community stability and invasibility: a case study based on Solidago canadensis L. Science of the Total Environment, 2021, 768: 144518. |
| [53] |
车应弟, 刘旻霞, 李俐蓉, 焦骄, 肖卫. 基于功能性状及系统发育的亚高寒草甸群落构建. 植物生态学报, 2017, 41(11): 1157-1167. |
| [54] |
Wang C Y, Cheng H Y, Wu B D, Jiang K, Wang S, Wei M, Du D L. The functional diversity of native ecosystems increases during the major invasion by the invasive alien species, Conyza canadensis. Ecological Engineering, 2021, 159: 106093. |
| [55] |
Wu H, Carrillo J, Ding J Q. Species diversity and environmental determinants of aquatic and terrestrial communities invaded by Alternanthera philoxeroides. Science of the Total Environment, 2017, 581/582: 666-675. |
| [56] |
Fenesi A, Geréd J, Meiners S J, Tóthmérész B, Török P, Ruprecht E. Does disturbance enhance the competitive effect of the invasive Solidago canadensis on the performance of two native grasses?. Biological Invasions, 2015, 17(11): 3303-3315. |
| [57] |
Zenni R D, da Cunha W L, Musso C, de Souza J V, Nardoto G B, Miranda H S. Synergistic impacts of co-occurring invasive grasses cause persistent effects in the soil-plant system after selective removal. Functional Ecology, 2020, 34(5): 1102-1112. |
| [58] |
Tekiela D R, Barney J N. Co-invasion of similar invaders results in analogous ecological impact niches and no synergies. Biological Invasions, 2017, 19(1): 147-159. |
| [59] |
Lenda M, Skórka P, Knops J, mihorski M, Gaj R, Moroń D, Woyciechowski M, Tryjanowski P. Multispecies invasion reduces the negative impact of single alien plant species on native flora. Diversity and Distributions, 2019, 25(6): 951-962. |
| [60] |
Villéger S, Mason N W H, Mouillot D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology, 2008, 89(8): 2290-2301. |
| [61] |
Wang C, Zhou J, Liu J, Xiao H G, Wang L. Differences in functional traits and reproductive allocations between native and invasive plants. Journal of Central South University, 2018, 25(3): 516-525. |
| [62] |
Gross N, Suding K N, Lavorel S, Roumet C. Complementarity as a mechanism of coexistence between functional groups of grasses. Journal of Ecology, 2007, 95(6): 1296-1305. |
| [63] |
Thomson F J, Moles A T, Auld T D, Kingsford R T. Seed dispersal distance is more strongly correlated with plant height than with seed mass. Journal of Ecology, 2011, 99(6): 1299-1307. |
| [64] |
Meng F Q, Cao R, Yang D M, Niklas K J, Sun S C. Trade-offs between light interception and leaf water shedding: a comparison of shade- and Sun-adapted species in a subtropical rainforest. Oecologia, 2014, 174(1): 13-22. |
| [65] |
Gross N, Suding K N, Lavorel S. Leaf dry matter content and lateral spread predict response to land use change for six subalpine grassland species. Journal of Vegetation Science, 2007, 18(2): 289-300. |
| [66] |
Pickett S T A, Cadenasso M L, Grove J M, Groffman P M, Band L E, Boone C G, Burch W R, Grimmond C S B, Hom J, Jenkins J C, Law N L, Nilon C H, Pouyat R V, Szlavecz K, Warren P S, Wilson M A. Beyond urban legends: an emerging framework of urban ecology, as illustrated by the Baltimore ecosystem study. BioScience, 2008, 58(2): 139-150. |
| [67] |
Pickett S T A, Cadenasso M L, Childers D L, McDonnell M J, Zhou W Q. Evolution and future of urban ecological science: ecology in, of, and for the city. Ecosystem Health and Sustainability, 2016, 2(7): e01229. |
| [68] |
Bradley B A, Laginhas B B, Whitlock R, Allen J M, Bates A E, Bernatchez G, Diez J M, Early R, Lenoir J, Vilà M, Sorte C J B. Disentangling the abundance-impact relationship for invasive species. Proceedings of the National Academy of Sciences of the United States of America, 2019, 116(20): 9919-9924. |
| [69] |
Duncan R P, Blackburn T M, Rossinelli S, Bacher S. Quantifying invasion risk: the relationship between establishment probability and founding population size. Methods in Ecology and Evolution, 2014, 5(11): 1255-1263. |