 2023, Vol. 43
2023, Vol. 43文章信息
- 李育森, 赵会宏, 王庆, 王大鹏, 罗永巨, 周磊, 黄世慧, 余梵冬
- LI Yusen, ZHAO Huihong, WANG Qing, WANG Dapeng, LUO Yongju, ZHOU Lei, HUANG Shihui, YU Fandong
- 大藤峡大坝蓄水对浔江鱼类群落的影响
- Effects of impoundment of Datengxia Dam on fish community of the Xunjiang River
- 生态学报. 2023, 43(17): 7140-7156
- Acta Ecologica Sinica. 2023, 43(17): 7140-7156
- http://dx.doi.org/10.20103/j.stxb.202209022499
-
文章历史
- 收稿日期: 2022-09-02
- 网络出版日期: 2023-04-28

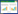




2. 广西水产科学研究院, 广西遗传育种与健康养殖重点实验室, 南宁 530021;
3. 中国水产科学研究院珠江水产研究所, 广州 510380
2. Guangxi Key Laboratory of Aquatic Genetic Breeding and Healthy Aquaculture, Guangxi Academy of Fishery Sciences, Nanning 530021, China;
3. Pearl River Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou 510380, China
鱼类作为水生生态系统中的重要类群, 可以通过下行效应影响整个水生态系统的生物与环境, 同时鱼类群落也会受到外界自然环境的影响[1]。基于鱼类群落对环境变化的响应也通常被用于生态系统完整性评价工作[2—4]。影响鱼类群落结构与资源量的因素可分为环境因素与人为因素, 环境因素又可以归类为生物因素与非生物因素两大类。唐衍力等[5]发现, 不同年龄的许氏平鮋由于饵料分布的差异, 以及存在着摄食偏好, 会分布于不同的环境以减少种内竞争;何雄波[6]研究表明, 不同鱼类可通过改变对相同食物的摄食比例来缓解种间竞争。鱼类有特定的水文需求, 水温会影响鱼类的生活史, 包括其分布、洄游、产卵和生长等[7]。在浊度较高的水域饵料生物的丰度相对更高, 鱼类饵料来源丰富, 更有利于鱼类种群的增长[8]。部分研究发现, pH与溶解氧是驱动鱼类群落结构变化的主要环境因子[9—11];也有研究发现盐度与底质也会影响鱼类群落结构[12—13]。
人为因素对鱼类群落的影响主要包括水利水电工程建设、过度捕捞、引进外来物种等[14—16]。近年来越来越多的水利水电工程不仅会对原有的鱼类群落产生影响, 还会导致栖息地的退化, 从而降低水域生态系统的生物多样性[17]。水利水电工程的运行改变了河流原本的水文动力、水质状况、初级生产力等进而影响鱼类群落[18]。大坝蓄水导致水体流速减缓, 大量泥沙、营养物质在库区滞留, 水体垂直交换受到限制, 水温与溶氧在垂直方向上分层[19]。有研究表明大坝降低了水体的搬运作用与水动力, 造成水中携带的泥沙与营养盐堆积, 虽然降低了水体的浊度与硬度等, 但有毒有害物质也随之沉降, 从而影响水质[20—21]。河流的连通性是保证鱼类洄游的前提, 拦河筑坝有可能会阻碍洄游性鱼类的索饵、生殖等洄游路线。葛洲坝截流后, 中华鲟数量锐减, 洄游到葛洲坝下产卵场的亲鱼减少, 自然产卵年际间由连续变成偶发[22];坝下江段白鲟与长江鲟的捕获数量也急剧减少[23]。除了直接阻断亲鱼洄游路线外, 鱼类繁殖还会受到大坝泄洪或者蓄水造成的水流速、水温等变化的影响。李伟等[24]对沅水干流四大家鱼的调查中发现, 大坝阻隔导致受精卵所需最小漂流距离不足从而影响了四大家鱼的繁殖, 同时水库下泄水温度偏低, 鱼类繁殖也会滞后。徐薇等[25]对银盘电站蓄水前后乌江下游繁殖鱼类调查发现, 流水生境繁殖类群减少、缓静水生境繁殖类群增加, 部分鱼类产卵规模与产卵场减小, 漂流性卵鱼类早期资源衰退。Rowell等[26]发现科罗拉河大坝的修建使得当地麦氏托头石首鱼幼鱼生长速率下降、性成熟年龄延缓。广西大藤峡枢纽蓄水后, 其坝下的浔江水文状况变化较大, 极有可能对浔江鱼类群落结构造成影响。本研究于浔江蓄水前后共计4年对其鱼类资源进行调查, 探究鱼类群落受到的影响, 为后续浔江的生态保护工作提供理论基础。
1 研究区域概况浔江位于广西壮族自治区东部, 属于珠江流域的西江干流中游, 自北向南的黔江与自西向东的郁江在桂平汇合称为浔江, 盛行亚热带季风气候。近年来关于浔江生态环境的研究报道越来越多[27], 陈晓鸿等[28]研究表明, 浔江已属于Ⅴ类水质, 水体总氮、总磷超标, 鱼体内锌、铬、砷同样存在超标现象, 同时, 张迎秋等[29]研究发现, 长洲水利枢纽改变了浔江桂平段的水文状况, 导致该江段鱼类优势种发生变化。广西大藤峡枢纽于2014年11月工程开工, 于2019年6月蓄水, 由黔江主坝与副坝、南木江副坝组成, 总库容34.79亿立方米, 防洪限制水位47.60 m, 防洪库容15亿立方米。根据全国大江大河实时水情记录(http://xxfb.mwr.cn/sq_djdh.html), 大坝蓄水前(2017年7月至2019年2月), 浔江大湟江口处流量为5963.64 m3/s, 蓄水后(2019年7月至2021年2月)流量有所减少, 为5709.63 m3/s;蓄水前平均水位为24.87 m, 蓄水后为22.78 m, 水位有所降低。
2 材料与方法 2.1 研究区域及样品采集调查区域为广西壮族自治区大藤峡枢纽坝下的浔江段(桂平至平南, 图 1), 本研究在大藤峡枢纽下游按从近到远设置了5个采样点, S1—S5距离大藤峡枢纽逐渐增大。每个采样点布置定制刺网2张(长度50 m, 高1.2 m, 网目尺寸3.5—6 cm), 虾笼2个(长15 m、高20 cm, 宽25 cm), 钓钩100个。作业时间为19时至次日7时。采集到的鱼类现场鉴定, 并测量和体长(精确到mm)、体重(精确到g)、尾数等数据, 随后放归浔江, 减少对物种资源的破坏。对于未能及时鉴定的物种则冷藏运送至实验室进一步鉴定并测量。

|
| 图 1 采样区域 Fig. 1 Sampling area S1—S5为距离大藤峡枢纽越来越远的5个采样点 |
调查时间为2017年7月至2021年2月, 调查频率为每月一次。7、8月为夏季, 9—11月为秋季, 12月至次年2月为冬季, 因此, 2017.07—2018.02, 2018.07—2019.02数据分别归为2017、2018年讨论, 后面的采样年度以此类推。大藤峡枢纽蓄水时间为2019年6月, 所以2017年、2018年表示蓄水前的情况, 2019年、2020年代表蓄水后情况。
2.2 鱼类丰度生物量比较(ABC)曲线与相似性聚类鱼类ABC曲线最初由Warwick[30]提出, 并已广泛应用于评估鱼类群落所受到的干扰水平[31]。从理论上讲, ABC曲线结合了物种丰度(数量)优势和生物量(质量)优势, 基于r—选择和K—选择的经典进化理论, 两种曲线可以产生3种可能的相对位置, 分别代表着受到轻度干扰, 中度干扰和强烈干扰的生物群落[30]。鱼类ABC曲线采用PRIMER 5.0软件绘制;此外, 采用相似性分析(SIMPER)进行组间相似性及主要贡献种的筛选[32]。采用非度量多维尺度分析(NMDS)对不同年份各采样点鱼类组成差异进行描述。
2.3 生态网络模型鱼类营养级数据取自世界鱼类数据库(https://www.fishbase.se/), 以鱼类为节点, 摄食关系为线, 通过营养级关系构建邻接矩阵, 最后将矩阵输入R语言转化为生态网络图。不考虑鱼类同类相食、寄生与腐蚀等关系, 假设两种鱼类营养级差值大于等于1时, 高营养级个体捕食低营养级个体[33]。
采用点度分布[p(k)] [34]、网络密度(D)[35]、平均路径长度(APL)[36]和聚类系数(C)[37]来评价生态网络结构的稳健性。采用累积概率分布[P(k)]来替代p(k)以便减少误差[38]。同时采用K-S检验网络的点度分布是否符合幂律分布[P(k)—k-α][39]。P>0.05则点度分布符合幂律分布, 此时的网络存在集散节点, 属于复杂网络。当α>3时, 网络中基本不存在离散节点;当3≥α>2时, 网络有少量集散节点, 为无标度网络;2≥α>1时, 拥有较多的集散节点。
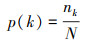
|
(1) |
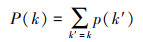
|
(2) |
式中, nk为点度值为k的数量, N为种类数, k和k′均表示某个点的点度。
网络密度(D)公式如下:
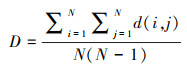
|
(3) |
式中, d(i, j)为种类i与j之间的关系量, N为种类数。
平均路径长度(APL):
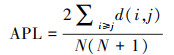
|
(4) |
式中, d(i, j)为种类i与j间的最短路径, N为种类数。
聚类系数(C):

|
(5) |
式中, N为种类数, Ei为种类i的k个邻节点之间实际存在的边数量, ki为种类i邻节点的数量。
生态网络图与相关参数的计算均通过R语言完成, 涉及igraph(https://igraph.org/)、reader(https://cran.r-project.org/web/packages/readr)、haven(https://cran.r-project.org/web/packages/haven/)、gglot2[40]安装包。
2.4 相对重要性指数与时间生态位相对重要性指数[41]:
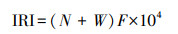
|
(6) |
式中, N为某一种类的数量占总尾数的百分比;W为某一种类的重量占总重量的百分比;F为某一种类的出现站数占调查总位点数的百分比。当IRI值大于500时为优势种, 500—100的为常见种, 100—10的为一般种, 10—1的为少见种, 小于1的为稀有种。本研究将IRI>10的鱼类作为主要鱼类进行研究。
时间生态位宽度采用Shannon-Wiener指数计算[42]:
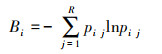
|
(7) |
式中, Bi为生态位宽度, Pij为种i在第j个站位下的个体数占该种所有个体数的比例;R为资源总数。Bi<1.00为窄生态位种, 1.00≤Bi≤2.00为中生态位种, Bi>2.00为广生态位种。
空间生态位重叠采用Pinaka指数计算[43]:
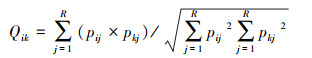
|
(8) |
式中, Qik为种i与种k的生态位重叠指数, 其值越大表示生态位重叠程度最高。其取值范围为0—1;Pij为种i在第j个资源状态下的个体数占该种所有个体数的比例;Pkj为种k在第j个资源状态下的个体数占该种所有个体数的比例;R为资源状态总数。其中, Qik>0.6时, 生态位显著重叠[44]。
2.5 总体相关性检验用Pianka生态重叠指数评估群落构建模式。
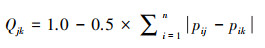
|
(9) |
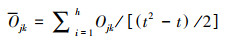
|
(10) |
式中, Qjk介于0(完全分离)和1(完全重叠)之间, 当竞争关系处于主导时, 实测值往往小于模拟值(P<0.05), 当生境过滤作用占主导时, 实测值一般大于模拟值(P<0.05), 两者无显著性差异表示随机作用占据主导。式中, Pij和Pik分别表示某一季节物种j和物种k在站位i的丰度与总丰度比值。Qjk表示某一季节各种对重叠指数(Qjk)平均值;t表示物种数目, h表示物种对数。计算过程均采用EcoSim700软件进行并采用随机化算法(RA3), (保留生态位宽度/重置零值, 迭代次数=1000, 随机数种子=10), 显著性水平定在p<0.05[45]。
标准化效应尺度(SES)
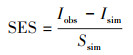
|
(11) |
式中, Iobs为群落C-score指数实测值, Isim为零模型群落值, Ssim为零模型群落标准离差。当标准化效应尺度值(SES)介于-2和2之间时, 说明群落处于随机分布格局(P>0.05); 当标准化效应尺度值绝对值大于2时(P<0.05), 说明群落为非随机分布, 正值说明群落处于分离格局, 而负值则说明群落处于集聚格局[45]。
2.6 相关数据的差异性分析使用SPSS 19.0软件与GraphPad Prism 9.0.0软件进行数据统计, 利用单因素方差分析(ANOVA)对近4年渔获物种类、数量、质量的季节、年度差异进行评价, 差异显著性阈值P=0.05。
3 结果与分析 3.1 鱼类群落物种组成和相似性浔江2017—2020年调查期间共采集到鱼类77种, 隶属7目19科63属。渔获重量与尾数如表 1所示。单因素方差分析结果显示, 蓄水前各季节种类数在21—55种, 日均渔获质量(8.20±3.46)kg至(12.03±2.78)kg, 日均渔获尾数在(89±42)尾至(326±127)尾之间;蓄水后各季节种类数在17—45种, 日均渔获质量(7.54±3.19)kg至(11.01±2.46) kg, 日均渔获尾数在(105±74)尾至(229±125)尾。蓄水前浔江鱼类种类数量和日均渔获重量优于蓄水后, 但未达到统计学意义上的显著差异;而日均渔获尾数在蓄水后有所上升。
| 年份 Year |
种类/尾 Species |
日均渔获重量/kg Catch mass per day |
日均渔获尾数/尾 Catch number per day |
|||||||||
| 夏 Summer |
秋 Autumn |
冬 Winter |
总和 Sum |
夏 Summer |
秋 Autumn |
冬 Winter |
夏 Summer |
秋 Autumn |
冬 Winter |
|||
| 2017 | 33 | 41 | 21 | 53 | 12.03±2.78 | 11.73±1.87 | 8.20±3.46 | 301±174 | 301±169 | 89±42 | ||
| 2018 | 38 | 24 | 35 | 55 | 8.85±2.27 | 9.59±1.76 | 9.89±0.62 | 326±127 | 131±28 | 162±7 | ||
| 2019 | 31 | 27 | 26 | 45 | 10.24±0.61 | 9.84±4.19 | 7.54±3.19 | 229±125 | 187±116 | 169±114 | ||
| 2020 | 23 | 22 | 17 | 34 | 8.89±2.46 | 11.01±2.46 | 10.78±0.98 | 105±74 | 120±17 | 202±25 | ||
采用非度量多维尺度分析(NMDS)对不同年份各采样点鱼类组成差异进行描述, 结果显示2017年与2018年各采样点之间相隔较远, 而2019年与2020年各采样点之间都相对聚集, 表明蓄水后各采样点间鱼类组成差异有所减小(图 2)。

|
| 图 2 浔江近4年鱼类群落结构的NMDS分析 Fig. 2 NMDS analysis for fish assemblages structure in Xunjiang river in recent 4 years NMDS:非度量多维尺度分析Non-metric multi-dimensional scaling |
各采样点鱼类种类差异如图 3所示:2017年与2018年鱼类种类数量呈现出上游采样点多于下游采样点的趋势;2019年与2020年有所不同, 下游采样点种类数高于上游采样点。于此同时, 优势种卷口鱼(Ptychidio jordani)、赤眼鳟(Squaliobarbus curriculus)、鲤(Cyprinus carpio)和粗唇鮠(Leiocassis crassilabris)中, 除了鲤随着时间推移在5个站点间始终分布较为均匀外, 其他优势种在各站点间均由原来的分布不均变得更加均匀(表 2)。

|
| 图 3 近4年浔江各采样点鱼类种类差异 Fig. 3 Fish species difference between different stations of Xunjiang river in recent 4 years |
| 年份Year 种名Species name |
2017 | 2018 | 2019 | 2020 | |||||||||||||||||||
| S1 | S2 | S3 | S4 | S5 | S1 | S2 | S3 | S4 | S5 | S1 | S2 | S3 | S4 | S5 | S1 | S2 | S3 | S4 | S5 | ||||
| 斑鱯Mystus guttatus | 2.35 | 1.95 | 2.58 | 1.20 | 1.38 | 2.61 | 2.22 | 4.05 | 0.52 | 1.24 | 1.97 | 2.75 | 2.16 | 2.69 | 1.64 | 1.44 | 0.76 | 1.36 | 1.19 | 1.36 | |||
| 草鱼Ctenopharyngodon idellus | 0.17 | 0.11 | 0.17 | 0.00 | 0.06 | 0.13 | 0.20 | 0.26 | 0.39 | 0.07 | 0.00 | 0.07 | 0.00 | 0.07 | 0.13 | 0.00 | 0.08 | 0.17 | 0.08 | 0.25 | |||
| 赤眼鳟Squaliobarbus curriculus | 0.86 | 0.57 | 3.56 | 0.75 | 0.17 | 1.31 | 1.70 | 3.20 | 0.26 | 0.13 | 0.92 | 0.79 | 0.98 | 0.85 | 0.46 | 0.85 | 1.27 | 1.10 | 0.51 | 1.19 | |||
| 粗唇鮠Leiocassis crassilabris | 6.66 | 4.59 | 3.44 | 4.25 | 2.64 | 3.92 | 1.57 | 3.66 | 1.31 | 0.78 | 4.91 | 4.33 | 3.34 | 4.72 | 4.06 | 5.27 | 4.67 | 3.91 | 5.69 | 4.33 | |||
| 大刺鳅Mastacembelus armatus | 0.34 | 0.46 | 0.92 | 0.06 | 0.06 | 0.26 | 0.20 | 0.91 | 0.20 | 0.07 | 0.66 | 0.46 | 0.79 | 0.92 | 0.66 | 0.34 | 0.25 | 0.42 | 0.25 | 0.51 | |||
| 大眼鳜Siniperca kneri | 0.00 | 0.00 | 0.11 | 0.00 | 0.00 | 0.07 | 0.00 | 0.07 | 0.13 | 0.00 | 0.20 | 0.33 | 0.20 | 0.39 | 0.26 | 0.34 | 0.42 | 0.25 | 0.34 | 0.25 | |||
| 鲫Carassius auratus | 1.15 | 0.46 | 0.17 | 0.29 | 0.00 | 4.18 | 1.96 | 3.27 | 0.91 | 0.20 | 0.13 | 0.20 | 0.26 | 0.00 | 0.20 | 0.00 | 0.00 | 0.00 | 0.00 | 0.08 | |||
| 间鱼骨Hemibarbus medius | 0.40 | 0.23 | 0.00 | 0.00 | 0.17 | 0.85 | 0.26 | 0.20 | 0.26 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.17 | 0.25 | 0.00 | |||
| 卷口鱼Ptychidio jordani | 0.75 | 1.15 | 0.98 | 3.73 | 0.57 | 2.48 | 3.33 | 1.57 | 4.96 | 3.85 | 3.41 | 5.96 | 4.19 | 4.98 | 6.95 | 6.88 | 7.82 | 6.54 | 9.60 | 11.47 | |||
| 鲤 Cyprinus carpio | 0.69 | 0.46 | 0.52 | 0.75 | 0.63 | 0.59 | 0.72 | 0.52 | 1.18 | 0.59 | 0.59 | 0.85 | 0.39 | 0.46 | 0.59 | 0.42 | 0.34 | 0.51 | 0.68 | 0.85 | |||
| 鲮Cirrhinus molitorella | 0.40 | 0.06 | 0.17 | 0.23 | 0.34 | 0.72 | 0.91 | 0.65 | 0.20 | 0.26 | 0.20 | 0.07 | 0.13 | 0.26 | 0.20 | 0.51 | 0.42 | 0.51 | 0.68 | 1.10 | |||
| 麦瑞加拉鲮Cirrhinus mrigala | 7.46 | 2.41 | 0.11 | 0.29 | 0.00 | 0.00 | 0.39 | 0.00 | 0.07 | 0.07 | 0.00 | 0.00 | 0.00 | 0.00 | 0.13 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |||
| 罗非鱼Tilapia | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.98 | 0.39 | 0.20 | 0.72 | 2.29 | 2.23 | 1.31 | 1.77 | 2.23 | 1.25 | 0.51 | 0.51 | 0.68 | 0.59 | 0.93 | |||
| 鲇Silurus asotus | 0.86 | 0.11 | 0.63 | 0.17 | 0.23 | 1.44 | 0.26 | 0.85 | 0.39 | 0.52 | 0.26 | 0.20 | 0.26 | 0.52 | 0.39 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |||
| 瓦氏黄颡鱼Pelteobagrus vachelli | 1.15 | 0.98 | 1.38 | 0.46 | 0.23 | 2.22 | 1.11 | 1.37 | 0.46 | 0.39 | 1.77 | 2.36 | 1.25 | 1.51 | 2.23 | 0.34 | 0.85 | 0.59 | 1.02 | 0.59 | |||
| 物种数量统计Statistics of species/种 | 33 | 26 | 31 | 25 | 20 | 36 | 32 | 29 | 26 | 21 | 23 | 25 | 21 | 26 | 34 | 17 | 15 | 17 | 20 | 26 | |||
| 仅展示丰度排序前15的经济鱼类; | |||||||||||||||||||||||
基于渔获种类与重量的相似性聚类分析显示, 各年度间鱼类群落相似度均在59.73%以上, 其中2018年与2019年鱼类群落相似性最高, 达75.42%, 2017年与2020年鱼类群落相似性最低, 仅为59.73%(图 4), 对相似性贡献最高的种类为卷口鱼、其次为赤眼鳟。

|
| 图 4 浔江鱼类群落相似性聚类 Fig. 4 Similarity clustering of fish communities in Xunjiang river 卷口鱼Ptychidio jordani;赤眼鳟Squaliobarbus curriculus;鲤 Cyprinus carpio;粗唇鮠Leiocassis crassilabris;斑鱯Mystus guttatus;鲮Cirrhinus molitorella;罗非鱼sp Tilapia;瓦氏黄颡鱼Pelteobagrus vachelli;鲇Silurus asotus;麦瑞加拉鲮Cirrhinus mrigala;草鱼Ctenopharyngodon idellus;大刺鳅Mastacembelus armatus;海南鮊Culter recurviceps;间鱼骨Hemibarbus medius;尖头塘鳢Eleotris oxycephala;大眼鳜Siniperca kneri;鲫Carassius auratus;银鯝Xenocypris argentea;东方墨头鱼Garra orientalis;短须鱊Acheilognathus barbatulus |
浔江近4年鱼类ABC曲线如图 5所示:大藤峡大坝蓄水前后, 曲线发生了明显的变化, W值在-0.004到0.092之间。累积生物量曲线起点呈连年上升趋势, 部分鱼类优势度有所上升。蓄水前生物量曲线位于丰度曲线上方, 鱼类群落内生物量比丰度更显优势, 鱼类群落受到外界干扰较小, 尤其是2018年鱼类群落最为稳定, 此前W值均大于0;蓄水后生物量曲线与丰度曲线交叉重叠现象更为严重, 丰度与生物量曲线分布优势难分, 鱼类群落受到的干扰较为严重, 其中2020年曲线W值为-0.004, 受到的干扰加剧, 干扰程度在大坝蓄水后随着时间的推移而加剧。

|
| 图 5 浔江鱼类ABC曲线 Fig. 5 Fish ABC curve of Xunjiang river ABC:鱼类丰度-生物量比较曲线Abundance biomass comparison curve |
参与生态网络构建的鱼类共有77种, 其中2017年53种, 2018年55种, 2019年45种, 2020年34种。营养级范围2—4.4。顶级物种(仅为捕食者, 营养级>3.4)有红鳍原鲌(Cultrichthys erythropterus)、鲶(Silurus asotus )、淡水石斑鱼、长吻鮠(Leiocassis longirostris)等序号范围在1—20的种类, 中间物种(同时作为捕食者与被捕食者, 3≤营养级≤3.4)有中间黄颡鱼(Pelteobagrus intermedius)、粗唇鮠、七丝鲚(Coilia grayii)等序号范围在21—58的种类, 带半刺光唇鱼(Acrossocheilus hemispinus cinctus)、麦瑞加拉鲮(Cirrhinus mrigala)、鲮(Cirrhinus molitorella)等序号范围59—77的为基础物种(仅被捕食, 营养级<3)(表 3)。
| 序号 Serial numbers |
种名 Species names |
营养级 Trophic levels |
序号 Serial numbers |
种名 Species names |
营养级 Trophic levels |
|
| 1 | 红鳍原鲌 Cultrichthys erythropterus | 4.4 | 40 | 大眼华鳊Sinibrama macrops | 3.2 | |
| 2 | 鲶Silurus asotus | 4.4 | 41 | 南方拟Pseudohemiculter dispar | 3.2 | |
| 3 | 淡水石斑鱼Cichlasoma managuense | 4 | 42 | 海南似鱎 Toxabramis houdemeri | 3.2 | |
| 4 | 长吻Leiocassis longirostris | 3.9 | 43 | 银鯝Xenocypris argentea | 3.2 | |
| 5 | 大眼鳜Siniperca kneri | 3.9 | 44 | 乐山小鳔鮈Microphysogobio fukiensis | 3.2 | |
| 6 | 斑鳜Siniperca scherzeri | 3.9 | 45 | 倒刺鲃Spinibarbus deniculatus | 3.2 | |
| 7 | 黑体塘鳢Eleotris melanosoma | 3.9 | 46 | 壮体沙鳅Botia robusta | 3.2 | |
| 8 | 革胡子鲶Clarias gariepinus | 3.8 | 47 | 泥鳅Misgurnus anguillicaudatus | 3.2 | |
| 9 | 叉尾斗鱼Macropodus opercularis | 3.8 | 48 | 大鳞副泥鳅Paramisgurnus dabryanus | 3.2 | |
| 10 | 丁鱥Tinca tinca | 3.7 | 49 | 小黄黝鱼Micropercops swinhonis | 3.2 | |
| 11 | 胡子鲶Clarias fuscus | 3.7 | 50 | 子陵吻虾虎鱼Rhinogobius giurinus | 3.2 | |
| 12 | 斑鱯Mystus guttatus | 3.7 | 51 | 拟细鲫Nicholsicypris normalis | 3.1 | |
| 13 | 尖头塘鳢Eleotris oxycephala | 3.7 | 52 | 细鳊Rasborinus lineatus | 3.1 | |
| 14 | 月鳢Channa asiatica | 3.7 | 53 | 纹唇鱼Osteochilus salsburyi | 3.1 | |
| 15 | 斑鳢Channa maculate | 3.7 | 54 | 鲤Cyprinus carpio | 3.1 | |
| 16 | 越南鲶Pterocryptis cochinchinensis | 3.5 | 55 | 卷口鱼Ptychidio jordani | 3.1 | |
| 17 | 西江鲶SilurusgilbertiHora | 3.5 | 56 | 麦穗鱼Pseudorasbora parva | 3.1 | |
| 18 | 黄颡鱼Pelteobagrus fulvidraco | 3.5 | 57 | 无斑南鳅Schistura incerta | 3 | |
| 19 | 瓦氏黄颡鱼Pelteobagrus vachelli | 3.5 | 58 | 横纹南鳅Schistura fasciolata | 3 | |
| 20 | 条纹鮠Leiocassis virgatus | 3.5 | 59 | Hemiculter leucisculus | 2.8 | |
| 21 | 中间黄颡鱼Pelteobagrus intermedius | 3.4 | 60 | 鳙Hypophthalmichthys nobilis | 2.8 | |
| 22 | 粗唇鮠Leiocassis crassilabris | 3.4 | 61 | 大鳞鲃 Barbus capito | 2.8 | |
| 23 | 七丝鲚Coilia grayii | 3.4 | 62 | 大刺鳅Mastacembelus armatus | 2.8 | |
| 24 | 鱤Elopichthys bambusa | 3.3 | 63 | 带半刺光唇鱼 Acrossocheilus hemispinus cinctus |
2.7 | |
| 25 | 广东鲂Megalobrama hoffmanni | 3.3 | 64 | 南方白甲鱼 Onychostoma gerlachi | 2.7 | |
| 26 | 间鱼骨Hemibarbus medius | 3.3 | 65 | 高体鰟鮍Rhodeus ocellatus | 2.5 | |
| 27 | 唇鱼骨Hemibarbus labeo | 3.3 | 66 | 鲫Carassius auratus | 2.5 | |
| 28 | 银鮈Squalidus argentatus | 3.3 | 67 | 东方墨头鱼Garra orientalis | 2.5 | |
| 29 | 蛇鮈Saurogobio dabryi | 3.3 | 68 | 短盖巨脂鲤Piaractus brachypomus | 2.5 | |
| 30 | 四须盘鮈Discogobiotetrabarbatus | 3.3 | 69 | 下口鲶Hypostomus plecostomus | 2.5 | |
| 31 | 大斑薄鳅Leptobotia pellegrini | 3.3 | 70 | 麦瑞加拉鲮Cirrhinus mrigala | 2.3 | |
| 32 | 花斑副沙鳅Parabotia fasciata | 3.3 | 71 | 越南鱊Acheilognathus tonkinensis | 2.2 | |
| 33 | 中华花鳅Cobitis sinensis | 3.3 | 72 | 短须鱊Acheilognathus barbatulus | 2.2 | |
| 34 | 弓斑东方鲀 Takifugu ocellatus | 3.3 | 73 | 草鱼Ctenopharyngodon idellus | 2 | |
| 35 | 杂交鲟 | 3.3 | 74 | 鳊Parabramis pekinensis | 2 | |
| 36 | 青鱼Mylopharyngodon piceus | 3.2 | 75 | 鲮Cirrhinus molitorella | 2 | |
| 37 | 赤眼鳟Squaliobarbus curriculus | 3.2 | 76 | 鲢Hypophthalmichthys molitrix | 2 | |
| 38 | 海南鮊Culter recurviceps | 3.2 | 77 | 罗非鱼sp Tilapia | 2 | |
| 39 | 飘鱼Pseudolaubuca sinensis | 3.2 | ||||
| 鱼类营养级数据取自世界鱼类数据库(https://www.fishbase.se/) | ||||||
浔江鱼类生态特性如表 4、图 6所示:2017年与2018年网络密度水平相当, 均为0.11, 2019年网络密度明显降低(0.09), 2020年网络密度又增至0.13。平均路径长度均为1。幂指数呈现递增趋势, 表示网络中的集散节点数在减少, 2017—2019年3≥α>2, 且p>0.05, 生态网络中有仅少量集散节点, 为无标度的复杂网络结构, 2020年α>3, 此时生态网络中基本不存在集散节点, 为简单网络, 于此同时聚类系数为0。
| 年份 Year |
网络密度 Network density(D) |
平均路径长度 Average path length(APL) |
聚类系数 Clustering coefficient(C) |
幂指数 Power exponent(α) |
P |
| 2017 | 0.11 | 1.00 | 0.11 | 2.41 | 0.59 |
| 2018 | 0.11 | 1.00 | 0.06 | 2.80 | 0.83 |
| 2019 | 0.09 | 1.00 | 0.06 | 2.81 | 0.70 |
| 2020 | 0.13 | 1.00 | 0.00 | 3.04 | 0.70 |

调查期间, 卷口鱼、赤眼鳟、鲤、粗唇鮠为优势种(IRI>500), IRI分别为2347.23、1317.83、1029.10、850.59;罗非鱼sp(Tilapia)、斑鱯(Mystus guttatus)等7种鱼类为常见种, IRI值在104.47—451.23之间;鲶、海南鲌(Culter recurviceps)、大眼鳜(Siniperca kneri)等9种鱼类为一般种。本文仅讨论IRI>10的经济鱼类, 大藤峡蓄水前后, 20种主要鱼类时间生态位宽度在0.81—2.47之间, 蓄水前时间生态位最宽的是鲤(2.47), 其次是赤眼鳟(2.46), 最窄的是纹唇鱼(Osteochilus salsburyi)(0.73)。蓄水后时间生态位最宽的是卷口鱼(2.42), 其次是鲤(3.35), 最窄是鲶(0.81)。蓄水前后始终为广生态位种(Bi>2.00)的有卷口鱼、赤眼鳟、鲤等;始终为中生态位种(1.00≤Bi≤2.00)的有罗非鱼sp、大眼鳜、鲫等;此外, 以东方墨头鱼为首的9种鱼类生态位宽度变宽;鲶为首的11种鱼类生态位宽度变窄。大藤峡枢纽蓄水后, 草鱼由广生态位种变为中生态位种;鲶由中生态位种变为窄生态位种;大刺鳅与海南鲌由中生态位种变为广生态位种;东方墨头鱼与纹唇鱼由窄生态位种变为中生态位种(表 5)。
| 序号 Order numbers |
种名 Species names |
IRI | 时间生态位宽度Bi Temporal niche width | ||
| 蓄水前 | 蓄水后 | 差值 | |||
| sp1 | 卷口鱼 | 2347.25 | 2.19 | 2.42 | -0.23 |
| sp2 | 赤眼鳟 | 1317.83 | 2.46 | 2.14 | 0.32 |
| sp3 | 鲤 | 1029.10 | 2.47 | 2.35 | 0.12 |
| sp4 | 粗唇鮠 | 850.59 | 2.23 | 2.31 | -0.08 |
| sp5 | 罗非鱼sp | 451.23 | 1.47 | 1.46 | 0.01 |
| sp6 | 斑鱯 | 411.22 | 2.22 | 2.19 | 0.03 |
| sp7 | 鲮 | 288.51 | 2.03 | 2.08 | -0.05 |
| sp8 | 瓦氏黄颡鱼 | 232.64 | 2.37 | 2.07 | 0.3 |
| sp9 | 间鱼骨 | 128.38 | 2.29 | 2.21 | 0.08 |
| sp10 | 草鱼 | 109.94 | 2.11 | 1.41 | 0.7 |
| sp11 | 大刺鳅 | 104.47 | 1.87 | 2.20 | -0.33 |
| sp12 | 鲶 | 95.75 | 1.66 | 0.81 | 0.85 |
| sp13 | 海南鲌 | 75.14 | 1.99 | 2.14 | -0.15 |
| sp14 | 大眼鳜 | 35.05 | 1.79 | 1.91 | -0.12 |
| sp15 | 鲫 | 31.59 | 1.01 | 1.41 | -0.4 |
| sp16 | 东方墨头鱼 | 24.88 | 0.87 | 1.96 | -1.09 |
| sp17 | 纹唇鱼 | 18.37 | 0.73 | 1.39 | -0.66 |
| sp18 | 银鲴 | 17.63 | 1.33 | 1.04 | 0.29 |
| sp19 | 南方拟 | 12.73 | 1.32 | 1.04 | 0.28 |
| sp20 | 七丝鲚 | 12.08 | 1.73 | 1.59 | 0.14 |
| 差值采用蓄水前该种时间生态位宽度减去蓄水后该种生态位宽度;IRI:相对重要性指数Index of relative importance | |||||
蓄水前后浔江主要鱼类时间生态位重叠值变化如表 6所示:共获得主要鱼类种对有380对, 时间生态位重叠值(Qik)范围为0—0.92。Qik>0.6, 即显著重叠的种对有83对, 占总对数的21.84%;0.6≥Qik≥ 0.3的种对有170对, 占总对数的44.74%;0.3>Qik的种对有127对, 占总对数的33.42%;瓦氏黄颡鱼-七丝鲚(sp8-sp20)的生态位重叠值最高(0.92), 其次是鲶-鲫(sp12—sp15)、间鱼骨-七丝鲚(sp9—sp20)重叠值分别为0.91、0.90, 均出现在蓄水后。而蓄水前生态位重叠最高的种对是鲤-草鱼(sp3—sp10), 为0.85;其次是赤眼鳟-大刺鳅(sp2—sp11)、间鱼骨-草鱼(sp9—sp10), 均为0.82, 这三个种对在蓄水后除了赤眼鳟-大刺鳅仍为显著重叠外, 另外两个种对重叠值均大幅度降低。蓄水前后, 显著重叠的种对有所下降, 由46对降为37对;0.6≥Qik≥0.3的种对有所上升, 76对上升为94对。
| sp1 | sp2 | sp3 | sp4 | sp5 | sp6 | sp7 | sp8 | sp9 | sp10 | sp11 | sp12 | sp13 | sp14 | sp15 | sp16 | sp17 | sp18 | sp19 | sp20 | |
| sp1 | 0.37 | 0.40 | 0.39 | 0.04 | 0.29 | 0.21 | 0.47 | 0.38 | 0.22 | 0.16 | 0.06 | 0.25 | 0.30 | 0.01 | 0.43 | 0.03 | 0.43 | 0.10 | 0.39 | |
| sp2 | 0.32 | 0.75 | 0.59 | 0.54 | 0.74 | 0.46 | 0.75 | 0.52 | 0.67 | 0.82 | 0.67 | 0.64 | 0.59 | 0.44 | 0.18 | 0.52 | 0.28 | 0.37 | 0.49 | |
| sp3 | 0.36 | 0.75 | 0.61 | 0.70 | 0.62 | 0.64 | 0.66 | 0.82 | 0.85 | 0.77 | 0.65 | 0.69 | 0.61 | 0.69 | 0.18 | 0.42 | 0.42 | 0.43 | 0.28 | |
| sp4 | 0.76 | 0.33 | 0.49 | 0.76 | 0.42 | 0.26 | 0.43 | 0.43 | 0.46 | 0.55 | 0.27 | 0.23 | 0.55 | 0.12 | 0.27 | 0.04 | 0.30 | 0.25 | 0.42 | |
| sp5 | 0.07 | 0.46 | 0.68 | 0.45 | 0.56 | 0.12 | 0.39 | 0.34 | 0.54 | 0.69 | 0.60 | 0.33 | 0.53 | 0.35 | 0.00 | 0.37 | 0.04 | 0.12 | 0.05 | |
| sp6 | 0.36 | 0.70 | 0.62 | 0.48 | 0.53 | 0.30 | 0.72 | 0.34 | 0.61 | 0.66 | 0.57 | 0.42 | 0.35 | 0.24 | 0.03 | 0.66 | 0.10 | 0.25 | 0.14 | |
| sp7 | 0.18 | 0.72 | 0.65 | 0.25 | 0.31 | 0.41 | 0.36 | 0.78 | 0.76 | 0.38 | 0.30 | 0.42 | 0.45 | 0.61 | 0.24 | 0.04 | 0.56 | 0.61 | 0.38 | |
| sp8 | 0.56 | 0.50 | 0.37 | 0.57 | 0.19 | 0.83 | 0.23 | 0.47 | 0.55 | 0.43 | 0.62 | 0.77 | 0.52 | 0.45 | 0.13 | 0.48 | 0.34 | 0.43 | 0.28 | |
| sp9 | 0.59 | 0.34 | 0.46 | 0.64 | 0.23 | 0.68 | 0.29 | 0.85 | 0.82 | 0.49 | 0.36 | 0.66 | 0.61 | 0.63 | 0.09 | 0.05 | 0.44 | 0.40 | 0.31 | |
| sp10 | 0.08 | 0.58 | 0.33 | 0.17 | 0.38 | 0.73 | 0.47 | 0.46 | 0.19 | 0.73 | 0.68 | 0.64 | 0.60 | 0.77 | 0.08 | 0.15 | 0.29 | 0.37 | 0.22 | |
| sp11 | 0.27 | 0.76 | 0.81 | 0.55 | 0.87 | 0.83 | 0.48 | 0.55 | 0.51 | 0.54 | 0.72 | 0.50 | 0.46 | 0.50 | 0.00 | 0.43 | 0.18 | 0.19 | 0.26 | |
| sp12 | 0.17 | 0.43 | 0.23 | 0.14 | 0.15 | 0.53 | 0.07 | 0.49 | 0.29 | 0.34 | 0.37 | 0.76 | 0.56 | 0.76 | 0.01 | 0.39 | 0.06 | 0.17 | 0.09 | |
| sp13 | 0.60 | 0.56 | 0.48 | 0.43 | 0.10 | 0.44 | 0.56 | 0.62 | 0.68 | 0.16 | 0.39 | 0.30 | 0.56 | 0.78 | 0.06 | 0.29 | 0.22 | 0.26 | 0.29 | |
| sp14 | 0.68 | 0.17 | 0.44 | 0.74 | 0.15 | 0.35 | 0.11 | 0.48 | 0.56 | 0.09 | 0.31 | 0.07 | 0.35 | 0.40 | 0.10 | 0.03 | 0.45 | 0.43 | 0.39 | |
| sp15 | 0.31 | 0.43 | 0.17 | 0.25 | 0.10 | 0.60 | 0.09 | 0.68 | 0.44 | 0.43 | 0.34 | 0.91 | 0.41 | 0.14 | 0.03 | 0.11 | 0.16 | 0.19 | 0.06 | |
| sp16 | 0.30 | 0.71 | 0.60 | 0.36 | 0.38 | 0.79 | 0.38 | 0.63 | 0.40 | 0.75 | 0.65 | 0.24 | 0.29 | 0.35 | 0.34 | 0.01 | 0.18 | 0.26 | 0.62 | |
| sp17 | 0.35 | 0.27 | 0.32 | 0.30 | 0.06 | 0.54 | 0.12 | 0.57 | 0.61 | 0.17 | 0.35 | 0.63 | 0.47 | 0.33 | 0.53 | 0.26 | 0.04 | 0.29 | 0.03 | |
| sp18 | 0.39 | 0.31 | 0.28 | 0.59 | 0.42 | 0.42 | 0.10 | 0.63 | 0.50 | 0.21 | 0.49 | 0.43 | 0.48 | 0.39 | 0.59 | 0.31 | 0.20 | 0.77 | 0.47 | |
| sp19 | 0.01 | 0.55 | 0.71 | 0.36 | 0.82 | 0.37 | 0.68 | 0.08 | 0.18 | 0.28 | 0.75 | 0.09 | 0.29 | 0.10 | 0.00 | 0.19 | 0.00 | 0.33 | 0.46 | |
| sp20 | 0.50 | 0.27 | 0.23 | 0.54 | 0.10 | 0.69 | 0.10 | 0.92 | 0.90 | 0.24 | 0.43 | 0.29 | 0.52 | 0.53 | 0.48 | 0.43 | 0.55 | 0.51 | 0.00 | |
| 对角线上方为蓄水前重叠值, 下方为蓄水后重叠值;编号对应的鱼类见表 5 | ||||||||||||||||||||
基于浔江鱼类时间生态位重叠构建的零模型结果显示:2017—2020年, Pianka指数实测平均值均大于模拟平均值, 表明近4年, 浔江鱼类群落的组成取决于环境因素, 生境过滤占主导作用。标准化效应尺度均大于2, 说明该群落为非随机分布, 处于分离格局。Pianka指数和资源利用模式的观测值与模拟值之间具有显著差异(p<0.05), 也表明浔江鱼类群落的构建是非随机占主导。但2020年标准化效应尺度降至2.70, 实测值更接近模拟值, 表明随着时间的推移, 浔江鱼类群落趋向于由竞争主导的随机分布发展格局, 栖息地的异质性有所降低(表 7)。
| 年份 Year |
实测值 Observe value |
模拟值 Simulated value |
P | 标准化效应尺度 Standardized effect size |
| 2017 | 0.31 | 0.24 | 0.001 | 9.43 |
| 2018 | 0.34 | 0.26 | 0.001 | 10.43 |
| 2019 | 0.35 | 0.28 | 0.001 | 9.23 |
| 2020 | 0.27 | 0.24 | 0.017 | 2.70 |
江河的自然流态通常被认为是水生生物多样性与生态完整性的要素之一[46], 大藤峡枢纽蓄水降低了河流连通性, 改变了河流原有的水文状况, 这也导致了鱼类群落发生了变化。比如, 大坝蓄水后浔江大湟江口处平均流量由5963.64 m3/s, 减少为5709.63 m3/s;平均水位由24.87 m显著降低为22.78 m。大坝蓄水后, 浔江两年度监测到的鱼类种类由蓄水前的69种降为51种, 物种数量有所下降, 这与前人研究结果相似[17]。大藤峡枢纽蓄水前, 鱼类ABC曲线揭示浔江鱼类群落呈现出好转的迹象, 仅受到轻微的外界干扰, 并且干扰主要来源于人为捕捞。究其原因, 可能是禁渔期制度的实施以及人工增殖放流和内陆捕捞准用渔具类型及渔具最小网目(或网囊)尺寸限制对鱼类资源的恢复产生了一定的效果。有研究表明不同类型刺网渔获物结果差异很大, 网目尺寸越小对渔获群落干扰越严重[47]。大藤峡枢纽蓄水后, 2019年ABC曲线显示, 生物量曲线与丰度曲线开始相交, 并且到2020年, 丰度曲线开始占优, W值已经小于0, 表明此时鱼类群落受到较为严重干扰[30, 32]。此外, 调查发现蓄水前浔江鱼类物种数量和日均渔获重量优于蓄水后, 且蓄水后鱼类群落以小规格鱼类为主, 这也表明了浔江鱼类群落结构在蓄水后发生了一定的改变。
2017—2019年浔江鱼类所构建的生态网络模型呈现为服从幂律分布的无标度复杂网络。吴康等[48]认为, 大多数网络连接集中于少数节点, 这是网络结构化和非随机性演化趋势的反映。本研究结果表明, 2017—2019年网络结构幂指数呈现递增趋势, 表示网络中的集散节点数在减少, 网络发生随机性演化, 结构趋向于简单化。聚类系数可以用来阐述种间相互作用, 聚类系数越低, 物种间的联系越相似[49], 本研究中聚类系数持续降低, 表明浔江鱼类的种间联系越来越相似, 网络复杂性降低。2020年的网络结构显示, 此时α>3, 网络中基本不存在离散节点, 并且聚类系数为0, 这可能会引起网络崩溃[33]。Dunne等[38]指出, 低连接度的食物网总体或者部分会服从幂律分布, 这与本文的研究结果类似。
浔江鱼类生态网络平均路径长度均为1, 表明生态系统中的能量可以通过摄食关系快速在不同营养级的鱼类间传递[50], 远低于海洋食物网路径长度的正常范围(1.3—3.7)[38]。大藤峡蓄水后, 随着时间推移, 鱼类种类数减少、网络中的节点数降低、网络密度呈减小趋势。鱼类群落结构主要受环境变化的影响[51], 浔江位于大藤峡枢纽的下游, 其水文状况会受到大坝的直接影响, 环境因子的改变会驱使不同类型鱼类的迁入与迁出, 从而会慢慢影响着鱼类的群落结构, 从而引起生态网络的响应。
4.2 大藤峡蓄水对浔江部分经济鱼类的影响本研究优势度排名前9的鱼类中, 除了罗非鱼sp始终为中生态位种外, 其他的均始终为广生态位种, 鱼类优势度与时间生态位宽度大小呈一定的正相关性。生态位宽度是物种对环境适应及种间竞争和分布范围评估的重要指标, 宽度值越大, 该物种对资源的利用和环境适应能力越好[52]。大藤峡蓄水后东方墨头鱼时间生态位宽度增加最多, 鲶下降最大, 表明东方墨头鱼对改变后的环境适应更强, 而鲶对当下环境适应度降低。大刺鳅与海南鲌时间生态位宽度发生了明显上升, 由中生态位种变为广生态位种;东方墨头鱼与纹唇鱼由窄生态位种变为中生态位种, 这些鱼类的时间生态位宽度增加可能是由于生存环境被压缩导致更容易被捕获所致。鲤为广布性鱼类, 对环境的适应性较强[53], 蓄水前后鲤的时间生态位宽度均保持在极高的水平, 表明在浔江的鱼类中, 鲤对环境资源的利用最强, 其出现的时间维度最宽, 更能适应季节变换引起的环境变化。Olsson等[54]指出, 外来入侵鱼类具有更大的生态位, 本研究中罗非鱼sp时间生态位宽度可以认为未发生变化, 表明它对环境的适应能力极强, 环境变化并不能让它的种群发生明显变动。
时间生态位重叠表示物种资源利用在时间上的相似程度, 重叠指数越高则资源利用的时间选择越相似[55], 反映物种间的竞争关系[56]。本研究发现, 时间生态位重叠值的变化主要集中在某些种对上, 总体上, 并无显著性差异。蓄水前时间生态位重叠最高的种对鲤-草鱼(0.85), 在蓄水后重叠值降低, 这是喜深水环境的草鱼向下游迁移所致, 这与本研究发现蓄水后45.45%的草鱼在S5采样点被捕获的结果一致。摄食习惯是导致种间竞争的重要因素之一[57], 蓄水后瓦氏黄颡鱼-七丝鲚的生态位重叠值升高至0.92, 二者均为肉食性鱼类, 会发生激烈的食物竞争, 可见蓄水后部分鱼类种间竞争加强。郑挺等[58]指出, 优势种群之间对环境的要求互补会表现出较高的生态位重叠度。本研究中时间生态位重叠较高的种对如鲶-鲫(蓄水后0.91)、赤眼鳟-大刺鳅(蓄水前0.82), 其食性方面差异较大, 可以判断其重叠值较高并非食物竞争引起, 有可能是食性及生存空间的互补引起。大坝蓄水后, 与南方拟连接的种对时间生态位重叠降幅最大, 如鲫-南方拟, 纹唇鱼-南方拟, 南方拟为中上层鱼类, 鲫与纹唇鱼均为中下层鱼类, 虽然水位降低会导致南方拟与另外两者之间发生空间竞争, 但南方拟为不喜欢急流水环境鱼类, 向下游静水区迁移从而减少竞争, 于此对应的是本研究发现蓄水后南方拟仅在S5采样点被捕获。Pratchett等[59]的研究也表明, 在种间竞争过程中, 占据优势的种类会优先选择饵料丰富、条件适宜的区域, 而处于劣势的种类只能通过食性分化、选择其他栖息地等来减少竞争。
4.3 大藤峡蓄水对浔江鱼类群落构建机制的影响Stegen等[60]认为生物群落构建可以分为两个确定过程(同质性选择、异质性选择)和两个随机过程(同质性扩散、具生态漂变的扩散限制)。本研究基于时间生态位重叠指数, 构建零模型, 发现浔江2017—2020年鱼类群落的构建取决于环境因素, 生境过滤占主导作用, 为非随机分布的分离格局, 处于群落构建的异质性选择过程。牛克昌等[61]提出, 在有限的资源条件下, 相似物种会通过生态位分离以减少种间竞争, 此时竞争排斥所造成的影响会超过生境过滤[62]。而2020年构建的零模型参数结果显示, 标准化效应尺度降至2.70, 实测值更接近模拟值, 表明随着时间的推移, 浔江鱼类群落趋向于由竞争主导的随机分布发展格局, 栖息地的异质性有所降低;此外, 不同年份各采样点鱼类NMDS分析发现蓄水后各采样点鱼类差异降低也支持这一观点。坝下鱼类群落所受的影响可能随着时间的推移而发生变化, 环境稳定后物种间的相互作用可能会变得更加重要[63]。本研究发现, 大藤峡枢纽蓄水后, 浔江鱼类群落格局正在快速发生变化, 虽然目前还处于非随机分布的分离格局, 但是已经有明显向由竞争主导的随机分布格局发展的趋势。
5 结论大藤峡蓄水后, 浔江鱼类种类数减少, 浔江鱼类ABC曲线揭示鱼类群落在蓄水后开始受到严重干扰, 并随着时间的推移受到的干扰愈加严重;与此同时浔江鱼类生态网络复杂性降低, 发生随机性演化。部分鱼类的时间生态位重叠值上升, 某些鱼类种对的种间竞争加强;浔江鱼类群落由生境过滤占主导作用的非随机分布的分离格局向竞争主导的随机分布发展格局转变。因此, 大藤峡蓄水已经对浔江鱼类群落造成较大负面影响, 需及时进行保护与修复。建议大藤峡枢纽在履行水力发电、防洪泄洪、储水灌溉等功能的同时, 也要兼顾保持河流的持续流动, 在鱼类繁殖期进行生态流量补偿促进鱼类繁殖, 此外, 保持过鱼通道的畅通和保持一定的流速, 以便洄游习性鱼类进行迁移。
| [1] |
帅方敏, 李新辉, 陈方灿, 李跃飞, 杨计平, 李捷, 武智. 淡水鱼类功能多样性及其研究方法. 生态学报, 2017, 37(15): 5228-5237. |
| [2] |
Schmölcke U, Ritchie K. A new method in palaeoecology: fish community structure indicates environmental changes. International Journal of Earth Sciences, 2010, 99(8): 1763-1772. DOI:10.1007/s00531-010-0524-3 |
| [3] |
黄宝荣, 欧阳志云, 郑华, 王效科, 苗鸿. 生态系统完整性内涵及评价方法研究综述. 应用生态学报, 2006, 17(11): 2196-2202. DOI:10.3321/j.issn:1001-9332.2006.11.039 |
| [4] |
Piet G J, Jennings S. Response of potential fish community indicators to fishing. ICES Journal of Marine Science, 2005, 62(2): 214-225. DOI:10.1016/j.icesjms.2004.09.007 |
| [5] |
唐衍力, 解涛, 于浩林, 虎琦, 刘岳, 张萌萌, 方光杰. 环境与摄食对山东省近海鱼礁区不同体长许氏平鲉分布的影响. 水产学报, 2020, 44(6): 924-935. |
| [6] |
何雄波. 中国近海带鱼科(Trichiuridae)常见种空间分布、种群结构与营养生态研究[D]. 厦门: 集美大学, 2019.
|
| [7] |
赵利. 环境流二元特性及其表征研究[D]. 北京: 中国水利水电科学研究院, 2015.
|
| [8] |
Cyrus D P, Blaber S J M. Turbidity and salinity in a tropical northern Australian Estuary and their influence on fish distribution. Estuarine, Coastal and Shelf Science, 1992, 35(6): 545-563. DOI:10.1016/S0272-7714(05)80038-1 |
| [9] |
刘鸿, 牛建功, 刘春池, 张涛, 米乃瓦尔·木依提, 陈韩飞, 蔡林钢. 新疆额敏河干流鱼类群落结构及其与环境因子的关系. 生态学杂志, 2017, 36(12): 3558-3563. |
| [10] |
刘燕山, 谷先坤, 唐晟凯, 李大命, 曾庆飞, 陆建明, 毛志刚, 殷稼雯, 陈辉辉, 沈冬冬, 何浩然, 张彤晴. 东太湖鱼类群落结构特征及其与环境因子的关系. 生态学报, 2021, 41(2): 769-780. |
| [11] |
马金, 黄金玲, 陈锦辉, 李拜, 赵静, 高春霞, 王学昉, 田思泉. 基于GAM的长江口鱼类资源时空分布及影响因素. 水产学报, 2020, 44(6): 936-946. |
| [12] |
史赟荣, 晁敏, 全为民, 唐峰华, 沈新强, 袁骐, 黄厚见. 2010年春季长江口鱼类群落空间分布特征. 中国水产科学, 2011, 18(5): 1141-1151. |
| [13] |
翟璐, 韩东燕, 傅道军, 张崇良, 薛莹. 胶州湾及其邻近海域鱼类群落结构及与环境因子的关系. 中国水产科学, 2014, 21(4): 810-821. |
| [14] |
Jennings S, Warr K J, Mackinson S. Use of size-based production and stable isotope analyses to predict trophic transfer efficiencies and predator-prey body mass ratios in food webs. Marine Ecology Progress Series, 2002, 240: 11-20. DOI:10.3354/meps240011 |
| [15] |
Power M, Attrill M J, Thomas R M. Environmental factors and interactions affecting the temporal abundance of juvenile flatfish in the Thames Estuary. Journal of Sea Research, 2000, 43(2): 135-149. DOI:10.1016/S1385-1101(00)00010-1 |
| [16] |
金显仕, 单秀娟, 郭学武, 李显森. 长江口及其邻近海域渔业生物的群落结构特征. 生态学报, 2009, 29(9): 4761-4772. DOI:10.3321/j.issn:1000-0933.2009.09.021 |
| [17] |
Chapin Ⅲ F S, Zavaleta E S, Eviner V T, Naylor R L, Vitousek P M, Reynolds H L, Hooper D U, Lavorel S, Sala O E, Hobbie S E, Mack M C, Díaz S. Consequences of changing biodiversity. Nature, 2000, 405(6783): 234-242. DOI:10.1038/35012241 |
| [18] |
线薇薇, 刘瑞玉, 罗秉征. 三峡水库蓄水前长江口生态与环境. 长江流域资源与环境, 2004, 13(2): 119-123. |
| [19] |
赵林林, 朱广伟, 陈元芳, 李未, 朱梦圆, 姚昕, 蔡琳琳. 太湖水体水温垂向分层特征及其影响因素. 水科学进展, 2011, 22(6): 844-850. |
| [20] |
刘丛强, 汪福顺, 王雨春, 王宝利. 河流筑坝拦截的水环境响应——来自地球化学的视角. 长江流域资源与环境, 2009, 18(4): 384-396. DOI:10.3969/j.issn.1004-8227.2009.04.015 |
| [21] |
廖文婷, 邓红兵, 李若男, 郑华. 水利工程对坝下径流的影响——以葛洲坝、三峡水利枢纽为例. 生态学报, 2018, 38(5): 1750-1757. |
| [22] |
吴金明, 王成友, 张书环, 张辉, 杜浩, 刘志刚, 危起伟. 从连续到偶发: 中华鲟在葛洲坝下发生小规模自然繁殖. 中国水产科学, 2017, 24(3): 425-431. |
| [23] |
Zhang H, Wei Q W, Du H, Li L X. Present status and risk for extinction of the Dabry's sturgeon (Acipenser dabryanus) in the Yangtze River watershed: a concern for intensified rehabilitation needs. Journal of Applied Ichthyology, 2011, 27(2): 181-185. DOI:10.1111/j.1439-0426.2011.01674.x |
| [24] |
李伟, 郭伟杰, 赵伟华, 赵良元, 唐见, 汤显强. 沅水干流水生态保护现状及问题分析. 长江技术经济, 2019, 3(4): 75-82. |
| [25] |
徐薇, 杨志, 万力, 唐会元, 陈小娟. 银盘电站蓄水前后乌江下游产漂流性卵鱼类自然繁殖变化. 水生态学杂志, 2019, 40(6): 8-15. |
| [26] |
Rowell K, Flessa K W, Dettman D L, Román M J, Gerber L R, Findley L T. Diverting the Colorado River leads to a dramatic life history shift in an endangered marine fish. Biological Conservation, 2008, 141(4): 1138-1148. |
| [27] |
黎小正, 庞燕飞, 曾辉, 吴祥庆, 马宁, 梁雪松. 广西桂平东塔河段产卵场水质综合评价. 湛江海洋大学学报, 2005, 25(3): 34-39. |
| [28] |
陈晓鸿, 李强, 喇优抓. 浔江水体和虾虎鱼体内常见污染指标分析. 化工设计通讯, 2020, 46(9): 169-171. |
| [29] |
张迎秋, 黄稻田, 李新辉, 刘乾甫, 李捷, 李跃飞, 杨计平, 朱书礼. 西江鱼类群落结构和环境影响分析. 南方水产科学, 2020, 16(1): 42-52. |
| [30] |
Warwick R M. A new method for detecting pollution effects on marine macrobenthic communities. Marine Biology, 1986, 92(4): 557-562. |
| [31] |
Bianchi G, Hamukuaya H, Alvheim O. On the dynamics of demersal fish assemblages off Namibia in the 1990s. South African Journal of Marine Science, 2001, 23(1): 419-428. |
| [32] |
李圣法. 以数量生物量比较曲线评价东海鱼类群落的状况. 中国水产科学, 2008, 15(1): 136-144. |
| [33] |
林坤, 麦广铭, 王力飞, 王学锋. 2015—2018年珠江口近岸海域鱼类群落结构及其稳定性. 水产学报, 2020, 44(11): 1841-1850. |
| [34] |
Dorogovtsev S N, Mendes J F, Samukhin A N. Size-dependent degree distribution of a scale-free growing network. Physical Review E, Statistical, Nonlinear, and Soft Matter Physics, 2001, 63(6 Pt 1): 062101. |
| [35] |
Sheather S J, Jones M C. A reliable data-based bandwidth selection method for kernel density estimation. Journal of the Royal Statistical Society: Series B: Methodological, 1991, 53(3): 683-690. |
| [36] |
Newman M E J. The structure and function of complex networks. SIAM Review, 2003, 45(2): 167-256. |
| [37] |
Holland P W, Leinhardt S. Transitivity in structural models of small groups. Comparative Group Studies, 1971, 2(2): 107-124. |
| [38] |
Dunne J A, Williams R J, Martinez N D. Food-web structure and network theory: the role of connectance and size. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99(20): 12917-12922. |
| [39] |
Newman M. Power laws, Pareto distributions and Zipf's law. Contemporary Physics, 2005, 46(5): 323-351. |
| [40] |
Wickham H. ggplot2. Springer International Publishing, 2016.
|
| [41] |
Pinkas L. Food habits of albacore, bluefin tuna and bonito in California waters. Fish Bull, 1971, 152: 1-105. |
| [42] |
Shannon E E, Weaver W. The mathematical theory of communication. Urbana, Chicago, London: University Illinois Press, 1963.
|
| [43] |
Pianka E R. The structure of lizard communities. Annual Review of Ecology and Systematics, 1973, 4: 53-74. |
| [44] |
Wathne J A, Haug T, Lydersen C. Prey preference and niche overlap of ringed seals Phoca hispida and harp seals P. groenlandica in the Barents Sea. Marine Ecology Progress Series, 2000, 194: 233-239. |
| [45] |
With K A, Gotelli N J, Graves G R. Null models in ecology. Ecology, 1997, 78(8): 2642. |
| [46] |
Lytle D A, Poff N L. Adaptation to natural flow regimes. Trends in Ecology & Evolution, 2004, 19(2): 94-100. |
| [47] |
徐国强, 李鹏飞, 张洪亮, 李德伟, 朱文斌. 基于渔获群落多样性指标的深水流刺网渔具作业性能分析. 海洋湖沼通报, 2019(2): 70-80. |
| [48] |
吴康, 方创琳, 赵渺希. 中国城市网络的空间组织及其复杂性结构特征. 地理研究, 2015, 34(4): 711-728. |
| [49] |
Marina T I, Salinas V, Cordone G, Campana G, Moreira E, Deregibus D, Torre L, Sahade R, Tatián M, Barrera Oro E, de Troch M, Doyle S, Quartino M L, Saravia L A, Momo F R. The Food Web of Potter Cove (Antarctica): complexity, structure and function. Estuarine, Coastal and Shelf Science, 2018, 200: 141-151. |
| [50] |
Coll M, Lotze H K, Romanuk T N. Structural degradation in Mediterranean Sea food webs: testing ecological hypotheses using stochastic and mass-balance modelling. Ecosystems, 2008, 11(6): 939-960. |
| [51] |
王雪辉, 邱永松, 杜飞雁, 林昭进, 孙典荣, 黄硕琳. 北部湾鱼类群落格局及其与环境因子的关系. 水产学报, 2010, 34(10): 1579-1586. |
| [52] |
Soberon J, Peterson A T. Interpretation of models of fundamental ecological niches and species' distributional areas. Biodiversity Informatics, 2005, 2: 1-10. |
| [53] |
王丽, 陈求稳, 陈凯, 刘祥. 淮河干流基于生态流量的目标鱼类选择研究. 环境科学学报, 2017, 37(6): 2379-2386. |
| [54] |
Olsson K, Stenroth P, Nyström P, Granéli W. Invasions and niche width: does niche width of an introduced crayfish differ from a native crayfish?. Freshwater Biology, 2009, 54(8): 1731-1740. |
| [55] |
郭天宇, 许荣满. 北京东灵山地区鼠类体外寄生虫时间生态位的研究. 中国媒介生物学及控制杂志, 2003, 14(1): 30-32. |
| [56] |
王桂明, 周庆强, 钟文勤. 内蒙古典型草原4种常见小哺乳动物的营养生态位及相互关系. 生态学报, 1996, 16(1): 71-76. |
| [57] |
Gaymer C F, Himmelman J H, Johnson L E. Effect of intra- and interspecific interactions on the feeding behavior of two subtidal sea stars. Marine Ecology Progress Series, 2002, 232: 149-162. |
| [58] |
郑挺, 林元烧, 曹文清, 张文静, 郑连明, 王宇杰, 杨位迪. 北部湾北部生态系统结构与功能——浮游动物空间生态位及其分化. 生态学报, 2014, 34(13): 3635-3649. |
| [59] |
Pratchett M S, Berumen M L. Interspecific variation in distributions and diets of coral reef butterflyfishes (Teleostei: Chaetodontidae). Journal of Fish Biology, 2008, 73(7): 1730-1747. |
| [60] |
Stegen J C, Lin X J, Fredrickson J K, Chen X Y, Kennedy D W, Murray C J, Rockhold M L, Konopka A. Quantifying community assembly processes and identifying features that impose them. The ISME Journal, 2013, 7(11): 2069-2079. |
| [61] |
牛克昌, 刘怿宁, 沈泽昊, 何芳良, 方精云. 群落构建的中性理论和生态位理论. 生物多样性, 2009, 17(6): 579-593. |
| [62] |
史赟荣, 沈新强, 王云龙. 海湾鱼类群落物种共存机制——以湄洲湾为例. 中国水产科学, 2016, 23(1): 169-176. |
| [63] |
Zhang C, Fujiwara M, Pawluk M, Liu H Z, Cao W X, Gao X. Changes in taxonomic and functional diversity of fish communities after catastrophic habitat alteration caused by construction of Three Gorges Dam. Ecology and Evolution, 2020, 10(12): 5829-5839. |