 2023, Vol. 43
2023, Vol. 43文章信息
- 仝川, 罗敏, 陈鹭真, 黄佳芳
- TONG Chuan, LUO Min, CHEN Luzhen, HUANG Jiafang
- 滨海蓝碳湿地碳汇速率测定方法及中国的研究现状和挑战
- Methods of carbon sink rate measurement of coastal blue carbon wetland ecosystems, current situation and challenges in China
- 生态学报. 2023, 43(17): 6937-6950
- Acta Ecologica Sinica. 2023, 43(17): 6937-6950
- http://dx.doi.org/10.20103/j.stxb.202208162357
-
文章历史
- 收稿日期: 2022-08-16
- 网络出版日期: 2023-04-28





2. 湿润亚热带生态地理过程教育部重点实验室, 福州 350117;
3. 福州大学环境与安全工程学院, 福州 350116;
4. 厦门大学环境与生态学院, 厦门 361102
2. Key Laboratory of Humid Sub-tropical Eco-geographical Process of Ministry of Education, Fuzhou 350117, China;
3. College of Environment and Safety Engineering, Fuzhou University, Fuzhou 350116, China;
4. College of the Environment and Ecology, Xiamen University, Xiamen 361102, China
生态系统碳汇是指在一定时空尺度内生态系统对大气中二氧化碳(CO2)的净吸收且长期储存的碳, 也就是文献中经常出现的生态系统固碳, 更延伸地讲, 还包括生态系统固碳的过程和机制。湿地生态系统对于大气中CO2的吸收主要是通过植物的光合作用。生态系统碳汇速率指生态系统在单位时间、单位面积净吸收并长期储存的碳(C)量(C g m-2 a-1), 也就是常说的固碳速率。
多数湿地植物均具有较高的总初级生产力, 因此也就具有很强的对于大气中CO2的吸收能力。滨海蓝碳湿地主要包括红树林、滨海盐沼和海草床生态系统。红树植物地上和地下部分均可较长时间地固持植物通过光合作用吸收的CO2。当然, 对于红树林、滨海盐沼和海草床等滨海蓝碳生态系统的土壤/沉积物更是其植物光合固碳的最终封存场所, 因此, 碳累积和碳埋藏等专业术语多出现在关于滨海湿地固碳、碳汇的相关研究文献中[1—3]。
不同于森林、草原以及内陆泥炭湿地和沼泽湿地等内陆生态系统, 滨海蓝碳湿地生态系统是一个“开放的”生态系统, 其植物光合吸收的碳, 不论是地上植株枯萎形成的碎屑, 还是埋藏到沉积物中的植物地上凋落物以及地下根系死亡分解形成的溶解性有机碳(DOC)等, 一部分均会随着潮水的流动和地下水的流动输送到海洋中[4]。因此, 在更大的空间尺度上, 滨海蓝碳湿地生态系统对于大气中CO2的长期碳固持主要体现在其地上生物量固碳(主要是红树林生态系统)、其自身系统垂直方向沉积物/土壤的碳埋藏, 以及以DOC、溶解性无机碳(DIC)和颗粒有机碳(POC)等形式水平方向输送入海洋, 之后或通过沉积生物泵(BP)机制埋藏固持于近海沉积物, 或通过微型生物碳泵(MCP)机制[5], 产生惰性溶解有机碳(RDOC), 进而在海水中储存数千年[6](图 1)。

|
| 图 1 滨海蓝碳湿地生态系统广义的碳汇过程 Fig. 1 Broad carbon sink process of coastal blue carbon wetlands ecosystem ER: 生态系统呼吸; GPP: 总初级生产力; NEP: 净生态系统产生力; DOC: 溶解有机碳; POC: 颗粒态有机碳; DIC: 溶解无机碳; RROC: 惰性溶解有机碳 |
唐剑武等[7]提出了一个广义的海岸带蓝碳的定义, 即滨海盐沼、红树林和海草床等海岸带高等植物以及浮游植物、藻类和贝类生物等, 在自身生长和微生物的共同作用下, 将大气中的CO2吸收、转化并长期保存到海岸带底泥中的碳, 以及其中一部分从海岸带向近海及大洋输出的有机碳。滨海盐沼生态系统常被认为是一个碳输出的溢出系统[8]。尽管, 滨海盐沼生态系统土壤中埋藏着来源于自身植物光合固定的碳[9], 但是, 滨海盐沼生态系统也是一个初级生产的泵, 它源源不断地向周边环境, 包括近海海域以碎屑和DOC的形式输出有机物质[10]。海草床生态系统植物光合固定的碳也有相当大的比例输出到“海草床生态系统”之外, 甚至有一定比例的海草植物叶片直接被草食性动物(草食性鱼类和海龟等)取食而“溢出”海草床生态系统[11—12]。Duarte等[13]研究发现海草床约50%—70%的净初级生产力“溢出”到其生态系统之外并埋藏在其它地方。也就是说, 滨海蓝碳湿地生态系统是一个开放的生态系统, 其自身生态系统通过光合作用吸收的碳并未全部原位固持在自身生态系统的沉积物/土壤中, 而是部分输出到其周边生态系统(包括海洋生态系统)的沉积物和土壤中并长期固持下来。另一方面, 对于河口蓝碳湿地生态系统, 同时还是一个收纳其入海河流流域水平碳通量输出的重要“聚集系统”, 河流流域上、中和下游的森林生态系统光合固持的有机碳随着水土流失和河流径流也将输出到河口盐沼和红树林生态系统, 并可长期埋藏在以上湿地的沉积物中。
滨海蓝碳湿地生态系统碳汇的定义可以分别在狭义和广义两个层面加以界定。狭义的滨海湿地蓝碳碳汇仅指某一滨海蓝碳湿地生态系统植物光合净吸收并长期固持在自身生态系统中的碳; 广义的滨海湿地碳汇指除了本生态系统植物光合作用净吸收并长期固持在自身生态系统中的碳以外, 还包括该系统植物光合作用固定但最终通过横向传输过程输送到海洋生态系统并长期固执在海洋沉积物和海水中的碳(图 1)。按照狭义的滨海蓝碳湿地碳汇的定义, 对于滨海盐沼生态系统和海草床生态系统, 考虑到其地上植物部分很难长时间封存其光合吸收的碳, 故在计量其碳汇速率时一般仅考虑滨海盐沼植物和海草床植物光合固定并最终埋藏在其自身生态系统土壤中的碳, 图 2为在狭义滨海蓝碳湿地碳汇的框架下, 3个滨海蓝碳生态系统植物光合固碳后的形成碳汇部分及非碳汇部分(图 2)。

|
| 图 2 滨海蓝碳湿地生态系统狭义碳汇定义下的植物光合固碳的碳汇和非碳汇去向 Fig. 2 Directions of carbon sink and non-carbon sink of photosynthetic carbon fixation by coastal blue carbon wetland ecosystem plants under narrow carbon sink definition |
滨海蓝碳湿地生态系统碳汇速率的测定虽已有文献报道, 也提出了包括沉积物碳累积+植被净初级生产力法以及生态系统碳通量收支法[1—2, 14]。但是由于滨海蓝碳湿地生态系统的复杂性, 目前关于滨海蓝碳湿地生态系统碳汇速率的测定, 不论是在方法学上, 还是具体的测定技术等方面仍均面临着较大的挑战。下面分别阐述目前滨海蓝碳湿地生态系统碳汇速率测定的常用方法, 并分析讨论各方法面临的主要问题。
2.1 生态系统碳通量法湿地生态系统碳汇速率测定中不可回避的一个重要指标是净生态系统生产力(NEP)。NEP反映了湿地生态系统两大通量(光合作用和呼吸作用)间的差值, 即湿地植被总光合作用速率减去湿地生态系统呼吸作用消耗的光合作用产物的速率(这里的呼吸作用包括自养呼吸和异养呼吸), 是生态系统尺度上光合作用和呼吸作用平衡的结果, 即常说的湿地生态系统固碳速率, 也可以认为是碳汇速率, 即单位面积湿地生态系统在单位时间内对大气CO2的净固持量。然而, 鉴于测定技术的限制, 目前还很难准确测定这个指标。研究者们更多是采用近似的方法加以估测, 其中运用最多的就是湿地生态系统年尺度净生态系统二氧化碳交换(NEE), 并以此表征湿地生态系统年尺度NEP。NEE是气象学家的定义, 正值代表生态系系统净排放CO2, 负值代表净吸收CO2。当然, 测算湿地生态系统碳汇速率(C g m-2 a-1)还应该同时测定和扣除湿地生态系统年尺度甲烷(CH4)排放通量。Gallagher等[1]认为湿地生态系统碳通量法是测定滨海蓝碳湿地生态系统碳汇速率的一个“间接”方法。开放的滨海盐沼生态系统和海草床生态系统植物光合固碳的一部分最终以无机碳和有机碳的形式通过潮水和地下水的横向传输输送到周边近海海域[15—16], 因而没有能够固持在自身生态系统中, 而碳通量法测定的结果没有扣除该部分碳通量, 因此, 通过生态系统碳通量法测定的滨海蓝碳湿地生态系统NEE所表征的碳汇速率, 严格讲属于的滨海蓝碳湿地生态系统广义碳汇速率。目前主要的技术支持途径包括涡度协方差通量塔法和箱式法。
2.1.1 涡度协方差通量塔法目前涡度协方差通量塔法被广泛应用于自然生态系统NEE的测定。该方法基于微气象学原理, 通过安装在通量塔上的开路式(或闭路式)红外气体分析仪和三维超声风速仪直接测定和计算一定范围(数m2到数km2)内的生态系统NEE, 是高时间分辨率测定生态系统与大气间净CO2通量的主流方法[17]。虽然已发表的一些文献的题目并未涉及到碳汇速率, 但这些研究运用涡度协方差通量塔法测定了滨海盐沼生态系统和红树林生态系统年尺度上的NEE[18—22], 也就是测定了滨海湿地生态系统碳汇速率, 但是, 目前均较少同步测定CH4排放通量并加以扣除。Heilman等[18]较早运用涡度协方差通量塔法研究了美国纽卡斯河三角洲滨海沼泽湿地生态系统NEE, 研究表明: 潮水水淹增加可促进滨海沼泽湿地NEE, 潮水水淹阶段日尺度为碳源(0.16 mol m-2 d-1), 潮水退去湿地变干时为碳汇(-0.14 mol m-2 d-1)。涡度协方差通量塔法具有高时间分辨率(如每半小时)测定生态系统NEE的优点, 并可以捕获极端天气事件(如气象干旱等)对于NEE的影响, 特别是对于研究者难以高频率到达的滨海蓝碳湿地, 涡度协方差通量塔法具有明显的优势。但是由于涡度协方差通量塔建设和后期管理成本较高, 在一定程度上限制了其广泛的应用。
2.1.2 箱式法在没有安装涡度协方差通量塔的滨海盐沼湿地, 透明箱以及黑布遮盖+红外气体分析仪相结合的方法在测定碳收支和碳汇速率方面也得到较广泛的应用[23—26]。该方法成本较低, 相对简单易行, 其优势在于可以在一个河口区的多个滨海沼泽湿地同步开展测定, 特别是对于有多影响因子处理和重复(如氮负荷增加处理、增温处理或海平面上升处理等)的滨海盐沼湿地研究样地, 该方法具有很大的优势。该方法的具体测定步骤如下:
(1) 瞬时NEE、生态系统呼吸(ER)和CH4排放通量的测定
每月选择2—3个晴天, 在中午光照较高时段使用不同遮光率布匹遮住透明静态箱以模拟不同光照强度, 运用CO2/H2O气体分析仪, 分别测定不同透光率情景下静态箱内CO2浓度。在有光情景测定结束后, 最后用透光率为0%的黑布遮住静态箱, 测定黑暗情景下静态箱内CO2浓度。滨海沼泽湿地瞬时NEE和ER根据静态箱内CO2浓度的变化率计算得出[19, 24]。NEE和ER测定结束后, 利用静态暗箱或采集气样实验室利用气相色谱仪测定CH4气体浓度, 或原位运用CH4/CO2/H2O气体分析仪直接测定静态箱内CH4浓度, 最后根据静态箱内CH4浓度的变化率计算CH4排放通量[27—28]。
(2) 年尺度上总光合作用吸收量、NEE和ER估算
总光合作用吸收量表征指标包括总初级生产力(GPP)[29—30], 总光合产物(GPP)[31]、总生态系统交铁(GEE)[24, 32—33], 这些指标均表示生态系统通过光合作用对与大气中CO2的总吸收量。这里以GEE为例介绍由GEE和ER推算NEE的方法。GEE为负值, 表示生态系统光合作用吸收大气中的CO2[32]。基于不同光照情景下测定的瞬时NEE和ER, 根据GEE与NEE和ER的关系式[32, 24]计算生态系统GEE, 然后建立GEE与光合有效辐射值(PAR)的拟合关系模型, 同时建立ER与气温的拟合关系。
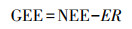
|
(1) |
GEE与PAR的直角双曲线拟合:
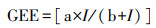
|
(2) |
式中, GEE: μmol CO2 m-2 s-1, I为PAR(μmol m-2 s-1), a, b为常数[24]。
每5 min或10 min 1次的PAR数据由测定样地附近的气象站读取, 根据公式2计算每5 min或10 min 1次的GEE值, 将每天内5 min或10 min间隔的GEE值求和得出日尺度GEE累计值, 然后尺度上移可计算月尺度GEE累计值和年尺度GEE累计值(g CO2 m-2 a-1)。
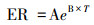
|
(3) |
式中, ER为生态系统呼吸速率(μmol CO2 m-2 s-1), T为气温(℃), A, B为经验常数[34]。
每5 min或10 min 1次的气温数据由测定样地附近的气象站读取, 利用公式3得出每5 min或10 min 1次的ER值, 将每天内5 min间隔的ER值求和得出日尺度ER累计值, 然后尺度上移可计算月尺度ER累计值和年尺度ER累计值(g CO2 m-2 a-1)。NEE月累计值通过GEE月累计值与ER月累计值由公式(1)计算得出, 各月NEE数值求和便得到年尺度的NEE(g CO2 m-2 a-1)。
(3) 滨海盐沼湿地生态系统年尺度蓝碳碳汇的估算
根据每月测定的CH4排放通量, 估算滨海盐沼湿地生态系统年CH4排放通量, 并根据百年尺度CH4全球变暖潜力值(GWP)25[35], 或百年尺度CH4持续增温潜势值(SGWPs)45[36—37], 将年CH4排放通量转化为CO2当量排放通量。
滨海盐沼湿地生态系统蓝碳碳汇速率可由NEE与CH4排放通量的代数和计算得出, 公式如下:
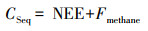
|
(4) |
式中, CSeq: 湿地生态系统年尺度碳汇速率(g CO2-eq m-2 a-1); NEE: 湿地生态系统年尺度CO2净交换量(g CO2 m-2 a-1); Fmethane: 湿地生态系统年尺度CH4排放通量的CO2当量通量值(g CO2-eq m-2 a-1)。
2.2 沉积物碳累积+植被净初级生产力结合法沉积物/土壤碳积累速率可表征滨海湿地生态系统净碳平衡, 它是湿地生态系统碳输入(包括植物光合作用碳输入和外源碳输入)和碳损失(CO2和CH4排放及地表和地下水中DOC的输出)的平衡值[2, 38]。通过测定红树林生态系统沉积物碳累积速率, 同时测定其植被净初级生产力(NPP), 最后将两者相加可得出红树林生态系统碳汇速率。目前关于滨海湿地沉积物碳积累的测定方法较为成熟。此外, 年尺度沉积物碳积累测定被认为是“扣除”滨海湿地生态系统碳收支中的横向碳输出量, 而运用涡度协方差通量塔法或箱式法未“扣除”横向碳输出量[39]。
以下分别介绍滨海湿地沉积物碳累积的测定方法以及植物部分碳汇速率计算方法。
2.2.1 沉积物碳累积速率的测定当湿地植物生物量处于一个较长期的稳定状态, 其固定的大气中的碳的大部分以较快的速度埋藏进入沉积物中[40]。因此, 测定滨海湿地沉积物/土壤的碳累积、或称碳埋藏一般被认为是测定滨海湿地生态系统碳汇速率的直接方法[1, 2]。目前采用的技术方法主要包括土芯+同位素定年方法及地表高程变化法。
(1) 土芯+同位素定年结合法
该方法通过采集滨海蓝碳湿地一定深度的沉积物土芯(如0—150 cm)并分层(如2 cm分层), 实验室测定各土层有机碳含量和容重, 最终计算滨海蓝碳湿地单位面积、一定深度沉积物的碳储量, 同时通过210Pb和137Cs测年, 评估沉积物不同分层的沉积年限, 最终确定滨海蓝碳湿地单位面积、一定时间段(较长期: 50—100年, 或短期<50年)沉积物碳累积速率、或称碳埋藏速率、或称碳垂直沉积速率[41—43], 并由此计算年尺度碳汇速率(C g m-2 a-1)。因为210Pb和137Cs测年需要沉积物样完全未受扰动才能反映真实沉积状况, 考虑到滨海蓝碳湿地生态系统沉积物多受到人类活动的干扰和多动力因素的影响, 有时需要采取包括人工放射性核素(如239+240 Pu)在内的多种方法加以定年[44]。
(2) 地表高程变化法
该方法运用水平标志层[45]或地表高程-标志层监测体系(SET)[46]测定滨海蓝碳湿地当年沉积物沉积速率[46—47]。采集当年沉积物样品, 室内测定沉积物有机碳含量和土壤容重, 计算滨海蓝碳湿地单位面积当年沉积形成的碳储量, 最终计算当年沉积物碳累积速率, 即年碳汇速率(C g m-2 a-1)。虽然地表高程变化可能受到地表沉积和侵蚀等因素的影响, 但由于该方法具有高精度和反复测量的优势, 目前已在全球滨海蓝碳湿地垂直沉积速率测量中得到广泛应用[48], 成为滨海盐沼和红树林生态系统地表高程变化的标准测量方法。
2.2.2 植被NPP的测定植被净初级生产力(NPP)是一个生态系统中植被部分单位时间、单位面积通过光合作用固定的CO2量减去植物自养呼吸后的剩余部分, 包括植被地上净初级生产力和植被地下净初级生产力。对于红树林生态系统, 在计量其碳汇速率时, 需要测定红树植物地上和地下部分年尺度NPP的变化。利用红树植物地上生物量估测模型(异速生长方程)估算红树植物在年初和年末的地上生物量, 并结合红树植物碳含量的测定, 最终估算红树植被地上NPP和红树植物地上部分碳汇速率(g C m-2 a-1)是常用的方法。不同红树林树种的异速生长方程存在差异, 地域的不同也有可能导致同种红树植物异速生长方程存在差异[49], 因此, 急需建立不同红树树种以及同一树种在不同区域的异速生长方程以满足红树林生态系统碳汇速率计量的需求。如何测定红树植被地下NPP, 进而估算红树植物地下部分碳汇速率, 却是一个很大的挑战。目前, 较多应用的方法是运用内生长土芯法[50], 该方法可以测定在一定时间尺度、一定土体新生长的红树植物根系的生物量。近年来, 微根管技术也开始在原位测定红树植物地下根系的长度、直径、密度及生命长度中应用[51]。滨海盐沼植被地上部分NPP的测定可运用样方收获法。海草床海草植被地上部分NPP的测定较为困难, 一般根据不同的海草生长型, 灵活采用叶片针孔标记法或剪叶标记法。叶片针孔标记法适用于叶片更替型海草植物, 这类海草植物种类较多, 如鳗草属(Zostera)、海菖蒲属(Enhalus)、虾形草属(Phyllospadix)、泰来草属(Thalassia)等; 剪叶标记法适用于非叶片更替型海草, 主要是喜盐草属(Halophila)海草[52]。
此外, 运用遥感技术, 特别是近年来出现的激光雷达数据以及高光谱数据, 结合地面红树林植物地上生物量的测定和光谱值的测定, 建立红树林植物地上生物量光谱估测模型, 并结合红树植物碳含量的测定, 最终估算红树林植被年尺度地上生物量的变化[53], 也可估算红树植物地上部分碳汇速率。
2.2.3 沉积物碳累积+植被NPP结合法的思考滨海蓝碳湿地沉积物碳积累与滨海蓝碳湿地生态系统碳汇的关系较为复杂。Gallagher等[1]认为滨海蓝碳湿地沉积物碳积累速率应减去外源碳的初始输入速率。此外, 在估算滨海蓝碳湿地沉积物对于通过植物光合净吸收并固持的大气中的C量时, 滨海蓝碳湿地沉积中原有的惰性碳(如黑炭), 在理论上应该从沉积物碳积累速率中扣除[54]。滨海蓝碳湿地沉积物/土壤中微塑料碳也被错误地统计到滨海蓝碳生态系统碳汇速率中[55], 而微塑料碳的外来性显然不应该属于滨海蓝碳湿地生态系统碳汇计量的范围。沉积物C: N同位素比值、沉积物有机碳δ13C值等均可用于辨识沉积物中外源和自身碳[14, 56]。一些学者认为区分滨海蓝碳湿地生态系统沉积物中自身和外源碳在其沉积物固持有机碳中的比例至关重要, 因为外源有机碳并不一定来自滨海蓝碳湿地生态系统对于大气CO2的净吸收和固持。一些研究表明: 对于滨海盐沼和海草床生态系统, 非自身产生的外源有机碳在其沉积物长期固持的有机碳中占据了较大的比例, 只有部分长期固持的有机碳来自滨海湿地生态系统植被凋落物分解和根系死亡后的碳输入[14, 57]。
如何评估滨海蓝碳湿地植被NPP与碳汇速率的关系也面临许多问题。目前中国关于滨海蓝碳湿地生态系统碳汇速率测定的行业和地方标准均采用滨海湿地生态系统植被NPP和沉积物碳累积相结合法。对于红树林生态系统, 红树植物地上茎生物量在较长时间尺度存在, 可以认为是对大气中CO2的长期固持, 故可以被计量到红树林生态系统碳汇速率中。但是, 对于滨海盐沼生态系统和海草床生态系统, 在估算其生态系统碳汇速率时, 地上植被NPP是否考虑?如从广义碳汇的角度出发加以考虑, 其对于滨海盐沼生态系统和海草床生态系统碳汇速率的贡献率是多少?盐沼植物和海草床植物地上部分凋落物既可原位分解并固持碳在湿地土壤中, 也有异位分解(由于潮水的作用, 最终发生分解过程的空间位置已偏离盐沼湿地, 或被直接冲入近海海域)。海草床生态系统一定比例的叶片还可被草食性动物(草食性鱼类和海龟等)取食, 此部分约占海草叶片(地上部分)生产力的10%[11—12]。由于滨海湿地年尺度植被地下NPP的测定较为困难, 如何将红树林、滨海盐沼和海草床生态系统年尺度地下植被NPP计量到其碳汇速率中也是一个难题。表 1比较了滨海蓝碳湿地生态系统碳汇速率不同测定方法的优点、存在的问题和挑战。
| 测定方法 Methods |
优点 Strengths |
存在的问题 Problems |
|
| 生态系统碳通量法 Ecosystem carbon flux observation |
涡度相关法 | 可实现精细时间尺度滨海蓝碳湿地生态系统NEE的长期连续测定。 | (1)无法兼顾小尺度范围内呈镶嵌状异质性分布的不同滨海湿地植物群落碳汇速率的测定; (2)无法运用于滨海湿地生态系统小尺度范围增温、施氮、盐水入侵等试验样地碳汇速率的测定; (3)采集的数据易受台风等异常气象事件的影响而发生缺失和失真; (4)由于滨海蓝碳湿地的开放性, 生态系统碳通量法无法兼顾滨海蓝碳湿地的横向碳输出, 测定的是广义碳汇速率。 |
| 箱式法 | 简单易行、成本较低, 特别是对于多点、多个环境因子、多重复的处理(如施氮、增温等)下滨海蓝碳湿地试验样地NEE的测定。 | (1)虽然该方法NEE通过模型的方法可以满足精细时间尺度的计算, 但是由于箱体无法长时间连续放置在滨海湿地, 故高时间分辨率的甲烷排放通量的测定较为困难; (2)尽管采集气样的时间间隔可设置较短, 但箱体的封闭环境仍可造成箱内气压和温度的微小变化, 并可能导致测定结果的偏差; (3)很难运用于高大红树林生态系统碳汇速率的测定; (4)由于透明静态箱的透光率很难达到100%, 故测定的NEE值可能被低估; (5)测定的是广义碳汇速率。 | |
| 沉积物碳累积+植被NPP法 Sediment carbon accumulation+vegetation NPP |
土芯+同位素定年结合法 | 可以测定滨海蓝碳湿地长时间尺度(<100年)内各年份沉积物碳累积速率。 | (1)湿地沉积物芯采集样地最好未受到人类活动的干扰和多动力因素的影响, 但在实际采样中较为困难; (2)估算碳汇速率时, 应扣除外源有机碳, 包括惰性有机碳(如黑炭等)和微塑料有机碳; (3)测定的是狭义碳汇速率。 |
| 地表高程变化法 | 可以测定滨海蓝碳湿地沉积物当年碳累积速率。 | (1)地表高程可能受到地表沉积和侵蚀等因素的影响; (2)在估算碳汇速率时, 应扣除外源有机碳, 包括惰性有机碳(如黑炭等)和微塑料有机碳; (3)估算河口蓝碳湿地碳汇速率时, 应扣除河流流域输入到河口湿地的外源有机碳沉积速率; (4)测定的是狭义碳汇速率。 | |
| 植被NPP | 测定红树林生态系统碳汇速率时, 可通过测定红树植物年尺度地上部分NPP的变化估算其植被部分碳汇速率 | (1)对于滨海盐沼湿地和海草床湿地, 在估算其生态系统碳汇速率时, 地上植被NPP是否考虑?如考虑, 它们对于盐沼生态系统和海草床生态系统碳汇速率的贡献率是多少仍未清晰。 | |
| NPP:净初级生产力Net primary productivity;NEE:净生态系统交换Net ecosystem exchange | |||
目前严格运用沉积物碳累积+植被NPP法或生态系统碳通量法测定中国滨海盐沼和海草床生态系统碳汇速率的研究极少。已有的研究只是单独测定滨海盐沼植被NPP, 并将盐沼植被NPP等同于盐沼植被固碳能力。梅雪英和张修峰[58]通过收割法测定长江口崇明东滩芦苇(Phragmites australis)沼泽秋季地上生物量, 并依据地下生产力与地上生产力的比例, 推算出崇明东滩芦苇植被NPP为1110—2410 g C m-2 a-1; 索安宁等[59]运用同样方法测定辽河三角洲芦苇植被NPP为4212 g C m-2 a-1, 翅碱蓬(Suaeda heteroptera)植被NPP为1296 g C m-2 a-1; 张绪良等[60]通过收割法测定黄河三角洲芦苇湿地、盐地碱蓬(Suaeda salsa)湿地秋季植被NPP, 两者分别为1230—1950 g C m-2 a-1和660—780 g C m-2 a-1。考虑到中国南方滨海盐沼湿地植物(如芦苇、互花米草(Spartina alterniflora))等在一年四季均生长, 用一年夏秋季某一月份地上、地下生物量表征其初级生产力是否合适?是测定我国南方滨海盐沼湿地植被NPP面临的一个主要问题。张林海[61]根据“增重累积法”测定闽江河口互花米草植被NPP最高(1401.4 g C m-2 a-1), 而芦苇、短叶茳芏(Cyperus malaccensis)沼泽植被NPP相对较低, 分别为1056.8和1035.1 g C m-2 a-1。
此外, 一些研究报道了利用涡度协方差通量塔法测定黄河口盐沼生态系统生长季或水淹情景下的NEE[6, 29, 62], 但是未见年尺度NEE的报道。Yan等[20]运用涡度协方差通量塔法测定长江口互花米草群落、芦苇群落和藨草(Scirpus)群落镶嵌体年尺度NEE, 研究结果表明3个植物群落镶嵌体在年尺度上为碳汇, NEE日均值为-1.75 g C m-2 day-1, 换算为年尺度NEE累积值为-638.8 g C m-2 a-1。但是, 以上研究未同时测定CH4排放通量, 故并没有从NEE中扣除CH4排放通量并计算其碳汇速率。
林晓雪等[63]运用生态系统碳通量箱式法, 同步测定了闽江河口芦苇沼泽湿地年尺度NEE和CH4排放通量, 根据100年尺度单位CH4的全球变暖潜势(GWP), 得出芦苇沼泽湿地年尺度CH4排放通量为(210.6±32.0) g CO2-eq m-2, 并最终估算出闽江河口芦苇沼泽湿地碳汇速率为(5371.5±337.0) g CO2-eq m-2 a-1, 换算成碳为(1464.9±91.9) g C m-2 a-1, 其中, CH4排放通量抵消芦苇沼泽湿地辐射冷却效益的3.8%。侯雪景等[42]运用采集土芯+ 210Pb和137Cs测年方法测定得出胶州湾大沽河口芦苇沼泽湿地沉积物碳累积速率为93.5 g C m-2 a-1。高亚平等[64]运用叶片针孔标记法测定了山东荣成桑沟湾大叶藻(Zostera marine)地上部分NPP及大叶藻叶片碳含量, 得出大叶藻植被碳汇速率为543 g C m-2 a-1, 低于根据大型海草地上部分NPP估算的碳汇速率(375 g C m-2 a-1)[11]。Huang等[65]研究表明: 中国东沙群岛分布的丝粉藻属(Cymodocea)、二药藻属(Halodule)、泰来藻(Thalassia hemprichii)等地上叶片和地下部分最终的碳埋藏速率为34.5 g C m-2 a-1, 低于全球海草生态系统平均碳埋藏速率(83 g C m-2 a-1)[66]。遗憾的是, 目前未见利用沉积物碳累积+植被NPP法测定中国滨海盐沼和海草床生态系统年尺度碳汇速率的报道。
3.2 中国红树林生态系统碳汇速率同样, 目前严格运用沉积物碳累积+植被NPP法测定中国红树林生态系统碳汇速率的研究也很少。张莉等[67]基于中国不同滨海地区红树林群落NPP值(干重)为1149—3640 g m-2 a-1, 以及红树林土壤呼吸消耗约1/3的NPP, 推算出中国滨海红树林生态系统碳汇速率为209—661 g C m-2 a-1。毛子龙等[68]通过生物量测定、凋落物测定以及土壤CO2和CH4通量测定, 基于碳收支法计算得出深圳福田秋茄红树林生态系统为大气中CO2的汇, 碳汇速率为648—2384 g C m-2 a-1。
孙明等[22]运用涡度协方差通量塔法测定广西北海以白骨壤(Aricennia marina) 群落为主, 混生少量秋茄(Kandelia obovata) 和桐花树(Aegiceras corniculata)的红树林生态系统年尺度NEE为-386.7 g C m-2 a -1。陈卉[69]运用涡度协方差通量塔法测定福建云霄红树林生态系统和广东高桥红树林生态系统(主要优势红树植物均为桐花树、秋茄和白骨壤等)2009—2012年的NEE, 2个红树林生态系统3年NEE的均值分别为-683、-721 g C m-2 a-1。Guo等[19]运用涡度协方差通量塔法测定长江口崇明岛互花米草群落、芦苇群落和藨草群落镶嵌体的NEE, 研究结果表明: 河口潮汐沼泽NEE受到潮汐的强烈影响, 高潮摊潮汐沼泽湿地具有更高的碳汇速率, 潮汐沼泽湿地NEE呈现出以10—20天为周期、明显受潮汐驱动的时间格局。Liu等[21]利用涡度协方差通量塔法连续3年测定香港米铺自然保护区秋茄红树林生态系统年尺度NEE和CH4排放通量, 秋茄红树林年尺度NEE为-891—-690 g C m-2·a-1, 年尺度CH4排放通量均值为(11.7± 0.4)g CH4 m-2 a-1, 最终根据20年和100年尺度CH4持续增温潜势(SGWPs)[36—37], 计算得出秋茄红树林生态系统在20年和100年时间尺度的碳汇速率为1376—2169 g CO2-eq m-2 a-1。
3.3 中国国家尺度滨海蓝碳湿地生态系统碳汇规模近年来, 中国科学家利用沉积物碳埋藏法估算中国滨海蓝碳湿地生态系统年碳汇规模, 结果汇总见表 2。这些研究均表明中国滨海蓝碳湿地生态系统是重要的碳汇, 具有较大的沉积物碳埋藏量, 但是不同研究的估算结果存在一定的差异。在3个滨海蓝碳湿地生态系统类型中, 滨海盐沼碳埋藏量最大, 占滨海蓝碳湿地碳埋藏总量的76%—91%, 其次是红树林生态系统, 海草床生态系统沉积物碳埋藏量最低, 占比仅为2%—5%。Fu等[41]和周晨昊等[70]估算的中国滨海蓝碳湿地滨海湿地碳埋藏量分别0.21、0.35—0.8 Tg C/a, 远低于Wang等[71]基于美国滨海湿地碳沉积数据和湿地调查数据估算的美国滨海蓝碳湿地碳埋藏量(4.2—5.0 Tg C/a)。
| 蓝碳生态系统类型 Blue ecosystem types |
碳埋藏速率 Carbon burial rates/(g C m-2a-1) |
碳埋藏量 Carbon burial/(Tg C/a) |
占碳埋藏总量 的百分比/% |
参考文献 References |
|
| 中国China | |||||
| 1 | 红树林 | 226±39 | 0.074 | 12 | 周晨昊等, 2016[70] |
| 滨海盐沼 | 218±24 | 0.263—0.749 | 86 | ||
| 海草床 | 138±38 | 0.012 | 2 | ||
| 滨海蓝碳生态系统 | 0.349—0.835 | 100 | |||
| 2 | 红树林 | 124 | 0.04±0.02 | 19 | Fu等, 2021[41] |
| 滨海盐沼 | 154 | 0.16±0.06 | 76 | ||
| 海草床 | 43 | 0.01±0.05 | 5 | ||
| 滨海蓝碳生态系统 | 209 | 0.21 | 100 | ||
| 3 | 红树林 | 160 | 0.05 | 4 | Wang等, 2021[71] |
| 滨海盐沼 | 168 | 1.19 | 96 | ||
| 滨海蓝碳生态系统 | 1.24 | 100 | |||
| 4 | 红树林 | 160 | 0.05 | 9 | 王法明等, 2021[72] |
| 滨海盐沼 | 160 | 0.50 | 91 | ||
| 滨海蓝碳生态系统 | 0.55 | 100 | |||
| 5 | 红树林 | 0.06 | 陈鹭真等, 2021[48] | ||
| 全球Global | |||||
| 1 | 红树林 | 139 | 23.6 | 21.2 | Chmura等, 2003[3]和 |
| 滨海盐沼 | 151 | 60.4 | 54.2 | Duarte等, 2005[66] | |
| 海草床 | 83 | 27.4 | 24.6 | Duarte等, 2005[66] | |
| 滨海蓝碳生态系统 | 111.4 | 100 | Duarte等, 2005[66] | ||
| 2 | 红树林 滨海盐沼 |
226±39 218±24 |
34.4±5.4 87.2±9.6 |
17.1 43.3 |
Duarte等2005[66]和 Mcleod等, 2011[43] |
| 海草床 | 138±38 | 48—112 | 39.6 | Mcleod等, 2011[43] | |
| 滨海蓝碳生态系统 | 100 | ||||
| 3 | 红树林 | 22.5—24.9 | 陈鹭真等, 2021[48] | ||
| 中国: 1中在计算滨海盐沼碳埋藏量占碳总埋藏量的百分比时, 碳埋藏速率取其范围的中值; Fu等中的数据不包括中国港澳台数据; 全球: 3中在计算海草床碳埋藏量占碳总埋藏量的百分比时, 碳埋藏速率取其范围的中值 | |||||
周晨昊等[70]的研究中, 滨海盐沼、红树林和海草床生态系统沉积物碳年埋藏速率是直接利用全球的均值, 而Fu等[41]根据自己采集的红树林、盐沼和海草床的土芯测得的沉积物年碳埋藏速率数据以及已发表的中国滨海蓝碳湿地73个土芯测年数据得出中国国家尺度滨海蓝碳湿地生态系统沉积物年碳埋藏速率。以上研究结果存在差异的主要原因除了各个滨海蓝碳湿地生态系统碳汇速率的不同, 各研究中采用的中国滨海盐沼、红树林的面积也不同。滨海蓝碳生态系统由于其面积相对较小, 因此, 目前在区域和国家尺度估算森林、草原生态系统碳汇规模的生态系统过程模型模拟法以及大气反演法均十分不适合在滨海蓝碳生态系统碳汇规模估算中应用。此外, 由于中国滨海蓝碳湿地还未建立完善的涡度协方差通量塔观测网, 因此, 也未见运用涡度协方差通量塔网测定和估算的国家尺度滨海蓝碳生态系统NEE数据, 以及由其表征的国家或区域尺度碳汇规模的报道。
全球尺度滨海盐沼和红树林生态系统的碳汇速率十分接近, 两者的碳汇速率几乎是海草床的2倍, 滨海盐沼的碳埋藏量最大, 明显高于其它2个滨海蓝碳湿地生态系统类型(表 2)。Duarte等[66]同时还估算了基于生态系统NEP表征的全球尺度滨海蓝碳湿地碳汇速率和碳汇规模, 红树林、滨海盐沼和海草床的碳汇速率分别221, 1585、1211 g C m-2 a-1, 碳汇规模分别为44634 Tg C/a和400 Tg C/a, 其中滨海盐沼和海草床碳汇速率明显高于这两个生态系统基于沉积物碳埋藏表征的碳汇速率, 几乎是后者的10倍, 这也表明: 滨海盐沼和海草床生态系统均为输出碳的“溢出系统”, 其植物光合作用固定的碳绝大部分均随着潮水和地下水输送到海洋生态系统, 而并未固持埋藏在本生态系统沉积物/土壤中。
4 展望与挑战在中国“双碳战略”实践中, 清查法、生态系统过程模型模拟法、大气反演法等方法在估算国家尺度和区域尺度森林生态系统、草原生态系统碳汇规模方面已经得到较好的应用[73]。然而, 对于众多面积小, 零散分布的滨海蓝碳湿地生态系统碳汇速率测定和碳汇规模的估算, 不论是在碳汇速率定义的界定, 还是测定和估算方法等方面仍均面临着较多的问题。周晨昊等[70]对中国海岸带蓝碳生态系统碳汇潜力进行了文献整理和初步分析, Fu等[41]结合自己采集的土芯以及已发表的中国滨海蓝碳湿地土芯测年数据估算了中国国家尺度滨海蓝碳湿地沉积物年碳埋藏速率。但是中国目前还没有在国家尺度上运用沉积物碳累积+植被NPP法, 或碳通量收支法估算的滨海蓝碳湿地碳汇速率及碳汇规模的数据。Macreadie等[74]在总结滨海蓝碳研究的10个重要科学问题时, 将“如何减少蓝碳估算中的不确定性”作为重要问题之一加以提出。滨海湿地蓝碳碳汇速率的准确测定是区域尺度蓝碳碳汇规模估算的重要基础。目前, 我国滨海湿地生态恢复与生态建设如火如荼, 如何准确、快速地计量滨海湿地生态恢复与建设中蓝碳湿地生态系统新增碳汇规模, 为开展碳贸易工作提供碳汇计量方法学与具体技术的支撑, 是一项急迫的任务。
首先要明确滨海蓝碳湿地生态系统碳汇速率的定义, 无论是采用狭义还是广义的滨海蓝碳湿地碳汇速率的定义, 在使用前均要加以说明。目前我国已出台的关于滨海蓝碳湿地生态系统碳汇速率计量的部门和行业标准, 全部采用的是沉积物碳累积+植被净初级生产力法。滨海盐沼和海草床生态系统由于其明显的开放性, 其地上植被NPP并未被计量到碳汇速率中, 这显然是基于滨海蓝碳碳汇速率的狭义定义的计量方法。
申霞等[75]通过对碳通量原位测定、碳收支过程测定、土壤碳储量变化测定的综述, 分析和讨论了滨海盐沼碳汇速率不同测定方法的优缺点, 陈鹭真[76]综述了地表高程变化法在滨海蓝碳碳收支评估中的应用, 以上文献对于在中国开展滨海蓝碳湿地碳汇速率的计量具有很好的指导意义。目前由于滨海湿地蓝碳碳汇速率计量方法并未统一, 且3个主要滨海蓝碳湿地生态系统结构和生态过程各具特点, 给滨海蓝碳湿地碳汇速率的测定结果带来了较大的确定性, 因此如何减少滨海湿地蓝碳碳汇速率测定的不确定性, 是目前滨海湿地蓝碳碳汇速率测定和区域尺度碳汇规模估算的一个重要任务与挑战。
展望未来, 中国在滨海蓝碳湿地碳汇速率和规模计量中须加强以下几个方面的基础研究。
(1) 对于中国滨海蓝碳湿地生态系统的主要类型(红树林、盐沼和海草床), 选取各自的优势植物群落类型和典型的生境, 分别运用沉积物碳累积+植被NPP法、生态系统碳通量法测定其碳汇速率, 分别建立严格的碳汇速率计量方法, 并通过比较研究, 量化运用以上2种不同计量方法计量的碳汇速率可能的差值范围。
(2) Wang等[77]利用美国滨海湿地碳沉积数据和美国湿地调查数据, 系统估算了美国国家尺度滨海湿地固碳速率和规模, 并对未来美国滨海湿地碳汇功能进行模拟预测。中国今后也需要在已有的约30个地表高程监测样点[76]的基础上, 在滨海蓝碳湿地生态系统不同类型、同一生态系统类型不同植物群落类型、同一植物群落类型不同生境(水文梯度、盐度梯度和养分状况等)建立更具系统性和代表性的中国滨海蓝碳湿地地表高程测定系统网络, 获取中国滨海蓝碳湿地沉积物沉积速率和土壤碳含量的系统数据, 为运用沉积物碳累积+植被NPP法测算中国国家尺度滨海蓝碳湿地的碳汇速率和规模奠定基础。
(3) 相对于森林生态系统和草原生态系统, 目前中国滨海蓝碳湿地生态系统涡度协方差碳通量塔网络的建设严重滞后, 一方面影响了基于生态系统碳通量收支法对中国滨海蓝碳湿地生态系统碳汇速率估算的精度; 另一方面, 也未能很好地跟踪滨海/河口潮汐过程以及极端天气事件过程(如风暴潮、台风和气象干旱等)对于中国滨海蓝碳湿地生态系统碳汇速率和过程的影响, 建议建立更具系统性和代表性的中国滨海蓝碳湿地涡度协方差碳通量塔网络。
(4) 虽然中国学者对于红树林树种异速生长方程的建立已有一些报道[78—80], 但是远远不足以支撑中国国家尺度红树林植被地上NPP的估算工作, 今后仍需要开展更多的相关研究。此外, 中国滨海盐沼湿地主要土著优势种(芦苇、藨草、短叶茳芏、茳芏)和外来入侵种互花米草等的异速生长方程的建立更鲜见文献报道[81—82], 今后同样需开展该领域的系统研究。同时, 运用激光雷达数据和高光谱数据开展红树林和滨海盐沼湿地地上生物量估测的研究也十分必要。
(5) 运用地表高程测定系统结合沉积物有机碳含量和容重的测定可估算滨海湿地当年新沉积物层碳埋藏速率, 但是, 对于滨海蓝碳湿地生态系统, 其新沉积层以下深度(如5—50 cm)土壤由于植物根系周转死亡形成的稳定性有机碳碳汇的速率如何计量?最近一些研究甚至表明土壤微生物残体碳在生态系统地下碳固持中占有较大的比例[83], 如何将滨海蓝碳湿地土壤微生物残体碳产生的新增碳汇计量到其碳汇速率中, 均将是一个新的挑战。
(6) 考虑到滨海盐沼生态系统和海草床生态系统均为开放的生态系统, 在计量其生态系统狭义碳汇速率时, 其地上部分植被NPP对于生态系统碳汇速率的贡献率是多少?滨海盐沼湿地地上NPP有多大比例原位分解并长期固持在其土壤中?有多大比例溢出本系统而输出到近海海域?不同河口段、不同潮滩位置、不同优势植物群落的贡献率差异如何?均需要开展系统和扎实的野外研究工作。此外, 如何将红树林、滨海盐沼和海草床生态系统年尺度地下NPP计量到其碳汇速率中也是一个需要深入研究的难题。
(7) 考虑到滨海蓝碳湿地生态系统沉积物中存在的惰性碳(如黑炭等)和微塑料碳的外源性, 运用沉积物碳累积+植被NPP法测定其碳汇速率时应加以扣除, 因此, 开展我国滨海蓝碳湿地生态系统沉积物黑炭和微塑料碳含量的测定十分必要。
| [1] |
Gallagher J B, Zhang K, Chuan C H. A re-evaluation of wetland carbon sink mitigation concepts and measurements: a diagenetic solution. Wetlands, 2022, 42(3): 23. DOI:10.1007/s13157-022-01539-5 |
| [2] |
Forbrich I, Giblin A, Hopkinson C. Constraining marsh carbon budgets using long-term C burial and contemporary atmospheric CO2 fluxes. Journal of Geophysical Research: Biogeosciences, 2018, 123: 867-878. DOI:10.1002/2017JG004336 |
| [3] |
Chmura G L, Anisfeld S C, Cahoon D R, Lynch J C. Global carbon sequestration in tidal, saline wetland soils. Global Biogeochemical Cycles, 2003, 17(4): 1111. |
| [4] |
Artigas F, Shin J Y, Hobble C, Marti-Donati A, Schäfer K V R, Pechmann I. Long term carbon storage potential and CO2 sink strength of a restored salt marsh in New Jersey. Agricultural and Forest Meteorology, 2015, 200: 313-321. DOI:10.1016/j.agrformet.2014.09.012 |
| [5] |
焦念志, 张传伦, 李超, 王晓雪, 党宏月, 曾庆璐, 张锐, 张瑶, 汤凯, 张子莲, 徐大鹏. 海洋微型生物碳泵储碳机制及气候效应. 中国科学: 地球科学, 2013, 43(1): 1-18. |
| [6] |
Hansell D A, Carlson C, Repeta D, Schlitzer R. Dissolved organic matter in the ocean: a controversy stimulates new insights. Oceanography, 2009, 22: 202-211. DOI:10.5670/oceanog.2009.109 |
| [7] |
唐剑武, 叶属峰, 陈雪初, 杨华蕾, 孙晓红, 王法明, 温泉, 陈少波. 海岸带蓝碳的科学概念、研究方法以及在生态恢复中的应用. 中国科学: 地球科学, 2018, 48(6): 661-670. |
| [8] |
Mitsch W J, Gosselink J G. Wetlands. Wiley, 2015.
|
| [9] |
Allen J R L. Morphodynamics of Holocene salt marshes: a review sketch from the Atlantic and Southern North Sea coasts of Europe. Quaternary Science Reviews, 2000, 19(12): 1155-1231. DOI:10.1016/S0277-3791(99)00034-7 |
| [10] |
Odum E P. A research challenge: evaluating the productivity of coastal and estuarine water. Proceedings of the second sea grant conference. University of Rhode Island Newport, RI, 1968: 63-64.
|
| [11] |
Larkum A, Orth R, Duarte C. Seagrasses: biology, ecology and conservation. Springer Netherlands, 2007. |
| [12] |
van Montfrans J, Wetzel R L, Orth R J. Epiphyte-grazer relationships in seagrass meadows: consequences for seagrass growth and production. Estuaries, 1984, 7(4): 289-309. DOI:10.2307/1351615 |
| [13] |
Duarte C M, Kennedy H, Marbà N, Hendriks I. Assessing the capacity of seagrass meadows for carbon burial: current limitations and future strategies. Ocean & Coastal Management, 2013, 83: 32-38. |
| [14] |
Mueller P, Ladiges N, Jack A, Schmiedl G, Kutzbach L, Jensen K, Nolte S. Assessing the long-term carbon-sequestration potential of the semi-natural salt marshes in the European Wadden Sea. Ecosphere, 2019, 10(1): 1-15. |
| [15] |
Chapin F S, Woodwell G M, Randerson J T, Rastetter E B, Lovett G M, Baldocchi D D, Clark D A, Harmon M E, Schimel D S, Valentini R, Wirth C, Aber J D, Cole J J, Goulden M L, Harden J W, Heimann M, Howarth R W, Matson P A, McGuire A D, Melillo J M, Mooney H A, Neff J C, Houghton R A, Pace M L, Ryan M G, Running S W, Sala O E, Schlesinger W H, -D Schulze E. Reconciling carbon-cycle concepts, terminology, and methods. Ecosystems, 2006, 9(7): 1041-1050. DOI:10.1007/s10021-005-0105-7 |
| [16] |
Cai W J. Estuarine and coastal ocean carbon paradox: CO2 sinks or sites of terrestrial carbon incineration?. Annual Review of Marine Science, 2011, 3: 123-145. DOI:10.1146/annurev-marine-120709-142723 |
| [17] |
Baldocchi D D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: past, present and future. Global Change Biology, 2003, 9(4): 479-492. DOI:10.1046/j.1365-2486.2003.00629.x |
| [18] |
Heilman J L, Cobos D R, Heinsch F A, Campbell C S, McInnes K J. Tower-based conditional sampling for measuring ecosystem-scale carbon dioxide exchange in coastal wetlands. Estuaries, 1999, 22(3): 584-591. DOI:10.2307/1353046 |
| [19] |
Guo H Q, Noormets A, Zhao B, Chen J Q, Sun G, Gu Y J, Li B, Chen J K. Tidal effects on net ecosystem exchange of carbon in an estuarine wetland. Agricultural and Forest Meteorology, 2009, 149(11): 1820-1828. DOI:10.1016/j.agrformet.2009.06.010 |
| [20] |
Yan Y E, Guo H Q, Gao Y, Zhao B, Chen J Q, Li B, Chen J K. Variations of net ecosystem CO2 exchange in a tidal inundated wetland: coupling MODIS and tower-based fluxes. Journal of Geophysical Research, 2010, 115(D15): D15102. DOI:10.1029/2009JD012838 |
| [21] |
Liu J G, Zhou Y L, Valach A, Shortt R, Kasak K, Rey-Sanchez C, Hemes K S, Baldocchi D, Lai D Y F. Methane emissions reduce the radiative cooling effect of a subtropical estuarine mangrove wetland by half. Global Change Biology, 2020, 26(9): 4998-5016. DOI:10.1111/gcb.15247 |
| [22] |
孙明, 莫伟华, 谢敏, 陈燕丽, 潘良浩. 广西红树林净生态系统碳交换变化特征及影响因子研究. 生态与农村环境学报, 2021, 37(7): 909-916. DOI:10.19741/j.issn.1673-4831.2020.0779 |
| [23] |
Järveoja J, Nilsson M B, Gažovič M, Crill P M, Peichl M. Partitioning of the net CO2 exchange using an automated chamber system reveals plant phenology as key control of production and respiration fluxes in a boreal peatland. Global Change Biology, 2018, 24(8): 3436-3451. DOI:10.1111/gcb.14292 |
| [24] |
Wilson B J, Mortazavi B, Kiene R P. Spatial and temporal variability in carbon dioxide and methane exchange at three coastal marshes along a salinity gradient in a northern Gulf of Mexico estuary. Biogeochemistry, 2015, 123(3): 329-347. DOI:10.1007/s10533-015-0085-4 |
| [25] |
Golovatskaya E A, Dyukarev E A. Carbon budget of oligotrophic mire sites in the Southern Taiga of Western Siberia. Plant and Soil, 2009, 315(1): 19-34. |
| [26] |
Bäckstrand K, Crill P M, Jackowicz-Korczyñski M, Mastepanov M, Christensen T R, Bastviken D. Annual carbon gas budget for a subarctic peatland, Northern Sweden. Biogeosciences, 2010, 7(1): 95-108. DOI:10.5194/bg-7-95-2010 |
| [27] |
Hirota M, Tang Y H, Hu Q W, Hirata S, Kato T, Mo W, Cao G, Mariko S. Methane emissions from different vegetation zones in a Qinghai-Tibetan Plateau wetland. Soil Biology & Biochemistry, 2004, 36: 737-748. |
| [28] |
仝川, 姚顺, 王维奇, 黄佳芳, 张林海, 章文龙, 曾从盛. 中国东南沿海短叶茳芏潮汐沼泽湿地甲烷动态. 中国科学: 地球科学, 2012, 42(5): 723-735. |
| [29] |
邢庆会, 韩广轩, 于君宝, 吴立新, 杨利琼, 毛培利, 王光美, 谢宝华. 黄河口潮间盐沼湿地生长季净生态系统CO2交换特征及其影响因素. 生态学报, 2014, 34(17): 4966-4979. |
| [30] |
Han G X, Xing Q H, Yu J B, Luo Y Q, Li D J, Yang L Q, Wang G M, Mao P L, Xie B H, Mikle N. Agricultural reclamation effects on ecosystem CO2 exchange of a coastal wetland in the Yellow River Delta. Agriculture, Ecosystems & Environment, 2014, 196: 187-198. |
| [31] |
Lloyd C R. Annual carbon balance of a managed wetland meadow in the Somerset Levels, UK. Agricultural and Forest Meteorology, 2006, 138(1): 168-179. |
| [32] |
Malone S L, Starr G, Staudhammer C L, Ryan M G. Effects of simulated drought on the carbon balance of Everglades short-hydroperiod marsh. Global Change Biology, 2013, 19(8): 2511-2523. DOI:10.1111/gcb.12211 |
| [33] |
Urbanski S, Barford C, Wofsy S, Kucharik C, Pyle E, Budney J, McKain K, Fitzjarrald D, Czikowsky M, Munger J W. Factors controlling CO2 exchange on timescales from hourly to decadal at Harvard Forest. Journal of Geophysical Research, 2007, 112(G2): G02020. |
| [34] |
Miller W D, Neubauer S C, Anderson I C. Effects of sea level induced disturbances on high salt marsh metabolism. Estuaries, 2001, 24(3): 357-367. DOI:10.2307/1353238 |
| [35] |
IPCC. Climate change: the physical science basis contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change. Cambridge: Cambridge University Press, 2007.
|
| [36] |
Neubauer S C. Moving beyond global warming potentials to quantify the climatic role of ecosystems. Ecosystems, 2015, 18(6): 1000-1013. DOI:10.1007/s10021-015-9879-4 |
| [37] |
Neubauer S C, Megonigal J P. Correction to: Moving beyond global warming potentials to quantify the climatic role of ecosystems. Ecosystems, 2019, 22: 1931-1932. DOI:10.1007/s10021-019-00422-5 |
| [38] |
Bogard M, Bergamaschi B, Butman D, Anderson F, Knox S, Windham-Myers L. Hydrologic export is a major component of coastal wetland carbon budgets. Global Biogeochemical Cycles, 2020, 34: e06430. |
| [39] |
Maher D T, Call M, Santos I R, Sanders C J. Beyond burial: lateral exchange is a significant atmospheric carbon sink in mangrove forests. Biology Letters, 2018, 14(7): 20180200. DOI:10.1098/rsbl.2018.0200 |
| [40] |
Prairie Y T, Alm J, Beaulieu J, Barros N, Battin T, Cole J, del Giorgio P, DelSontro T, Guérin F, Harby A, Harrison J, Mercier-Blais S, Serça D, Sobek S, Vachon D. Greenhouse gas emissions from freshwater reservoirs: what does the atmosphere see?. Ecosystems, 2018, 21(5): 1058-1071. DOI:10.1007/s10021-017-0198-9 |
| [41] |
Fu C C, Li Y, Zeng L, Zhang H B, Tu C, Zhou Q, Xiong K X, Wu J P, Duarte C M, Christie P, Luo Y M. Stocks and losses of soil organic carbon from Chinese vegetated coastal habitats. Global Change Biology, 2021, 27(1): 202-214. DOI:10.1111/gcb.15348 |
| [42] |
侯雪景, 印萍, 丁旋, 张勇, 毕世普. 青岛胶州湾大沽河口滨海湿地的碳埋藏能力. 海洋地质前沿, 2012, 28(11): 17-26. |
| [43] |
McLeod E, Chmura G L, Bouillon S, Salm R, Björk M, Duarte C M, Lovelock C E, Schlesinger W H, Silliman B R. A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Frontiers in Ecology and the Environment, 2011, 9(10): 552-560. DOI:10.1890/110004 |
| [44] |
Santos I R, Maher D T, Larkin R, Webb J R, Sanders C J. Carbon outwelling and outgassing vs. burial in an estuarine tidal creek surrounded by mangrove and saltmarsh wetlands. Limnology and Oceanography, 2019, 64(3): 996-1013. DOI:10.1002/lno.11090 |
| [45] |
Thomas S, Ridd P V. Review of methods to measure short time scale sediment accumulation. Marine Geology, 2004, 207(1/2/3/4): 95-114. |
| [46] |
Lovelock C E, Adame M F, Bennion V, Hayes M, O'Mara J, Reef R, Santini N S. Contemporary rates of carbon sequestration through vertical accretion of sediments in mangrove forests and saltmarshes of south east Queensland, Australia. Estuaries and Coasts, 2014, 37(3): 763-771. DOI:10.1007/s12237-013-9702-4 |
| [47] |
Lynch J, Hensel P, Cahoon D. The surface elevation table and marker horizon technique: a protocol for monitoring wetland elevation dynamics. National Park Service, 2015. |
| [48] |
陈鹭真, 杨盛昌, 林光辉. 全球变化下的中国红树林. 厦门: 厦门大学出版社, 2021.
|
| [49] |
Ong J E, Gong W K, Wong C H. Allometry and partitioning of the mangrove, Rhizophora apiculata. Forest Ecology and Management, 2004, 188(1): 395-408. |
| [50] |
Vogt K A, Vogt D J, Bloomfield J. Analysis of some direct and indirect methods for estimating root biomass and production of forests at an ecosystem level. Plant and Soil, 1998, 200(1): 71-89. |
| [51] |
Arnaud M, Morris P J, Baird A J, Dang H, Nguyen T T. Fine root production in a chronosequence of mature reforested mangroves. The New Phytologist, 2021, 232(4): 1591-1602. |
| [52] |
Short F T, Duarte C M. Methods for the measurement of seagrass growth and production. Global Seagrass Research Methods. Amsterdam: Elsevier, 2001: 155-182.
|
| [53] |
黎夏, 叶嘉安, 王树功, 刘凯, 刘小平, 钱峻屏, 陈晓越, 何执兼, 覃朝锋. 红树林湿地植被生物量的雷达遥感估算. 遥感学报, 2006, 10(3): 387-396. |
| [54] |
Chew S T, Gallagher J B. Accounting for black carbon lowers estimates of blue carbon storage services. Scientific Reports, 2018, 8: 2553. |
| [55] |
Rillig M C. Microplastic disguising As soil carbon storage. Environmental Science & Technology, 2018, 52(11): 6079-6080. |
| [56] |
Leorri E, Zimmerman A R, Mitra S, Christian R R, Fatela F, Mallinson D J. Refractory organic matter in coastal salt marshes-effect on C sequestration calculations. Science of the Total Environment, 2018, 633: 391-398. |
| [57] |
Van de Broek M, Vandendriessche C, Poppelmonde D, Merckx R, Temmerman S, Govers G. Long-term organic carbon sequestration in tidal marsh sediments is dominated by old-aged allochthonous inputs in a macrotidal estuary. Global Change Biology, 2018, 24(6): 2498-2512. |
| [58] |
梅雪英, 张修峰. 长江口典型湿地植被储碳、固碳功能研究——以崇明东滩芦苇带为例. 中国生态农业学报, 2008, 16(2): 269-272. |
| [59] |
索安宁, 赵冬至, 张丰收. 我国北方河口湿地植被储碳、固碳功能研究——以辽河三角洲盘锦地区为例. 海洋学研究, 2010, 28(3): 67-71. |
| [60] |
张绪良, 张朝晖, 徐宗军, 侯雪景, 蔡庆芳. 黄河三角洲滨海湿地植被的碳储量和固碳能力. 安全与环境学报, 2012, 12(6): 145-149. |
| [61] |
张林海. 闽江河口湿地优势植物净初级生产力及N、P养分动态[D]. 福州: 福建师范大学, 2008.
|
| [62] |
杨利琼, 韩广轩, 于君宝, 吴立新, 朱敏, 邢庆会, 王光美, 毛培利. 黄河三角洲芦苇湿地生长季净生态系统CO2交换及其环境调控机制. 应用生态学报, 2013, 24(9): 2415-2422. |
| [63] |
林晓雪, 黄佳芳, 李慧, 仝川. 闽江河口芦苇沼泽和短叶茳芏沼泽生态系统含碳温室气体的年收支. 生态学报, 2022, 42(22): 9186-9198. |
| [64] |
高亚平, 方建光, 唐望, 张继红, 任黎华, 杜美荣. 桑沟湾大叶藻海草床生态系统碳汇扩增力的估算. 渔业科学进展, 2013, 34(1): 17-21. |
| [65] |
Huang Y H, Lee C L, Chung C Y, Hsiao S C, Lin H J. Carbon budgets of multispecies seagrass beds at Dongsha Island in the South China Sea. Marine Environmental Research, 2015, 106: 92-102. |
| [66] |
Duarte C M, Middelburg J J, Caraco N. Major role of marine vegetation on the oceanic carbon cycle. Biogeosciences, 2005, 2(1): 1-8. |
| [67] |
张莉, 郭志华, 李志勇. 红树林湿地碳储量及碳汇研究进展. 应用生态学报, 2013, 24(4): 1153-1159. |
| [68] |
毛子龙, 杨小毛, 赵振业, 赖梅东, 杨道运, 吴纯玲, 徐华林. 深圳福田秋茄红树林生态系统碳循环的初步研究. 生态环境学报, 2012, 21(7): 1189-1199. |
| [69] |
陈卉. 中国两种亚热带红树林生态系统的碳固定、凋落物分解及其同化过程[D]. 厦门: 厦门大学, 2013.
|
| [70] |
周晨昊, 毛覃愉, 徐晓, 方长明, 骆永明, 李博. 中国海岸带蓝碳生态系统碳汇潜力的初步分析. 中国科学: 生命科学, 2016, 46(4): 475-486. |
| [71] |
Wang F M, Sanders C J, Santos I R, Tang J W, Schuerch M, Kirwan M L, Kopp R E, Zhu K, Li X Z, Yuan J C, Liu W Z, Li Z A. Global blue carbon accumulation in tidal wetlands increases with climate change. National Science Review, 2021, 8(9): nwaa296. |
| [72] |
王法明, 唐剑武, 叶思源, 刘纪化. 中国滨海湿地的蓝色碳汇功能及碳中和对策. 中国科学院院刊, 2021, 36(3): 241-251. |
| [73] |
朴世龙, 何悦, 王旭辉, 陈发虎. 中国陆地生态系统碳汇估算: 方法、进展、展望. 中国科学: 地球科学, 2022, 52(6): 1010-1020. |
| [74] |
Macreadie P I, Anton A, Raven J A, Beaumont N, Connolly R M, Friess D A, Kelleway J J, Kennedy H, Kuwae T, Lavery P S, Lovelock C E, Smale D A, Apostolaki E T, Atwood T B, Baldock J, Bianchi T S, Chmura G L, Eyre B D. The future of Blue Carbon science. Nature Communications, Nature Publishing Group, 2019, 10(1): 1-13. |
| [75] |
申霞, 王鹏, 王为攀, 王胜艳, 谢瑞, 王永平, 姬昌辉. 滨海盐沼净碳汇能力研究方法综述. 生态学杂志, 2022, 41(4): 792-803. |
| [76] |
陈鹭真. 地表高程监测在滨海蓝碳碳收支评估中的应用. 海洋与湖沼, 2022, 53(2): 261-268. |
| [77] |
Wang F M, Lu X L, Sanders C J, Tang J W. Tidal wetland resilience to sea level rise increases their carbon sequestration capacity in United States. Nature Communications, 2019, 10: 5434. |
| [78] |
昝启杰, 王勇军, 廖宝文, 郑德璋. 无瓣海桑、海桑人工林的生物量及生产力研究. 武汉植物学研究, 2001, 19(5): 391-396. |
| [79] |
金川, 王金旺, 郑坚, 陈秋夏, 李俊清, 卢翔. 异速生长法计算秋茄红树林生物量. 生态学报, 2012, 32(11): 3414-3422. |
| [80] |
陈国贵, 李元跃, 蔡丽钦, 徐佳佳, 林沛然, 陈文城, 黄森磊. 红树植物外来种无瓣海桑对乡土种秋茄形态特征与生物量的影响. 海洋科学, 2017, 41(6): 26-33. |
| [81] |
潘良浩, 史小芳, 范航清. 茳芏(Cyperus malaccensis Lam.)生物量估测模型. 广西科学院学报, 2015, 31(04): 259-263. |
| [82] |
刘白贵, 仝川, 黄佳芳, 谭季, 李慧. 闽江河口感潮湿地短叶茳芏地上绿植株与立枯生物量估测模型. 生态学杂志, 2022. DOI:10.13292/j.1000-4890.202210.001 |
| [83] |
Liang C, Amelung W, Lehmann J, Kästner M. Quantitative assessment of microbial necromass contribution to soil organic matter. Global Change Biology, 2019, 25(11): 3578-3590. |