 2023, Vol. 43
2023, Vol. 43文章信息
- 方笛熙, 万霞, 毛婉琼, 张锋
- FANG Dixi, WAN Xia, MAO Wanqiong, ZHANG Feng
- 病原体感染对物种间资源竞争的影响——基于资源竞争理论
- Effect of pathogen infection on resource competition between species: a study based on resource competition theory
- 生态学报. 2023, 43(17): 7109-7117
- Acta Ecologica Sinica. 2023, 43(17): 7109-7117
- http://dx.doi.org/10.20103/j.stxb.202208162351
-
文章历史
- 收稿日期: 2022-08-16
- 网络出版日期: 2023-04-28





2. 中国科学院亚热带农业生态研究所, 亚热带农业生态过程重点实验室, 长沙 410125;
3. 云南财经大学统计与数学学院, 昆明 650221
2. Key Laboratory of Argo-ecological Processes in Subtropical Region, Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha 410125, China;
3. Statistics and Mathematics School, Yunnan University of Finance and Economics, Kunming 650221, China
种间互作是物种共存和生物多样性维持的重要力量[1—3]。物种共存研究通常关注物种间的直接竞争作用[4—5], 然而更高营养级的消费者带来的间接效应在促进或阻止物种共存上有着潜在作用[6—9]。寄生作为一种最常见的消费策略广泛地存在于各类生态系统中[10—11], 通常被视为生态系统中的负面因素, 是疾病的起因和不健康环境的标志[12]。然而越来越多的研究正在改变着这一观点, 寄生者可以通过间接作用给非宿主物种带来积极效应, 间接影响种间竞争和消费者与资源的相互作用, 这些影响会通过群落内各营养级进行传播, 最终影响到整个生态系统的物质循环和能量流动[10, 13]。因此, 在研究物种共存和群落构建时, 考虑寄生的间接效应带来的影响是必要的。
病原体是一类在宿主体内无性繁殖, 能够引起宿主患上疾病但通常不会立即致其死亡的传染性寄生物[3, 14]。许多病原体与宿主之间存在高度的特异性(对宿主的选择具有高度专一性), 它们的宿主范围仅局限在一个种或某一特定类型的物种[15—17]。实验和理论研究都有证据表明此类病原体会对物种间的竞争产生影响。Schall[18]发现加勒比海地区的两种蜥蜴齿龈阿诺利蜥(Anolis gingivinus),沃氏阿诺利蜥(Anolis wattsii)只有在疟原虫(Plasmodium azurophilum)存在时才能共存。这种疟原虫很少感染A. wattsi, 而在A. gingivinius群落中很常见。尽管A. gingivinius是一个比A. wattsi更强的竞争者, 但是这种病原体降低了宿主的竞争能力, 最终允许两个蜥蜴物种共存。Fenton等[15]利用竞争模型(Lotka-Volterra模型)证明了单宿主病原体可以减低优势竞争者的密度, 从而削弱其竞争力, 因此能够促进物种共存。这些研究结果强调了单宿主病原体在物种共存、宿主群落构建及生物多样性维持中的重要作用, 但都没有明确竞争者和资源的相互作用[6—8, 15, 18—20], 因此很难从中理解病原体对物种资源利用方式的改变以及其对种间竞争的影响。本文考虑宿主高度特异的寄生类型, 通过耦合资源竞争模型和疾病传播模型, 研究了病原体对宿主的感染如何影响宿主与其竞争对手间的资源竞争。
1 资源竞争与物种共存不同于Lotka-Volterra竞争模型通过竞争系数抽象地刻画种间竞争的限制作用, Tilman的资源竞争模型既描述了竞争物种如何通过利用资源而增殖的过程, 也考虑了资源被消耗而衰减的动态变化[21—23]。因此, 使用资源竞争模型可以更好地理解物种间是如何通过利用共同的有限资源而发生竞争的。假设两物种(比如两种植物或者浮游植物)都依赖两种必需资源(比如氮和磷)生存, 种群和资源的动态变化可以用微分方程描述:
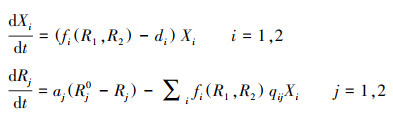
|
(1) |
式中, Xi为物种i的种群密度, Rj是资源j的可利用量;fi(R1, R2)表示物种i资源依赖的增殖率, qij表示物种i每单位繁殖对资源j的消耗量;aj是资源j的补给率, Rj0表示资源j独立于物种的固有平衡量(通常称供给点);di是物种i的死亡率。根据利比希最小因子定律, 可以设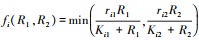
微分方程(1)的正平衡点满足代数方程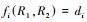
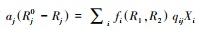
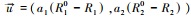
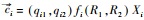
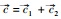
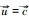
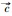

|
| 图 1 物种竞争两种基本资源的模式图解 Fig. 1 Graphic model of species competing for two essential resources ZNGI:零等倾线Zero net growth isocline;c1:物种1消费矢量;c2:物种1消费矢量;sp:资源供给点 |
解方程fiR1, R2=di得Rij*=Kijdi/rij-di, 表示能够支持物种i增长所需资源j的最低量, 即该物种对该资源的最低需求。由此可见, 竞争物种的零等倾线呈“L”型, 它们相交的条件是R11* < R21*且R12*>R22*, 或者R11*>R21*且R12* < R22*, 也就是说, 对一种资源的优势竞争者(R*更小)必是对另一种资源的弱势竞争者(R*更大)。平衡点是否稳定还取决于两物种对两种资源的相对消耗率。如果物种1对资源1有竞争优势(R11* < R21*), 物种2对资源2有竞争优势(R12*>R22*), 那么, 相对于资源1, 当物种1对资源2的消耗比物种2更快时, 即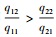
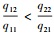
不失一般性, 假设病原体感染物种1(感染物种2结果类似), 其种群由易感者和感染者构成, 易感者可能被接触感染, 感染者由于病原体毒性使得死亡率增加。如果用S和I分别表示物种1的易感者和感染者密度, 基于方程(1), 可以用如下微分方程描述这个病原体胁迫下的竞争系统的种群动态:
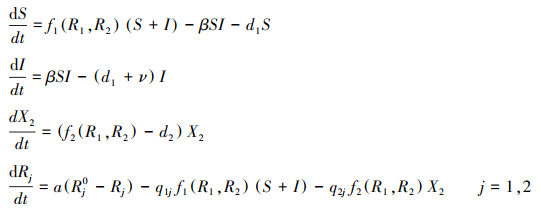
|
(2) |
式中, β为病原体的传播率, ν为疾病导致的死亡率(毒性), 其他符号和方程(1)相同。模型(2)耦合了物种1的感染动态和两物种的资源竞争过程, 可以用来研究病原体对物种竞争共存的影响。
病原体对物种竞争共存产生影响的前提是病原体能够入侵宿主种群, 这要求感染初期感染者的数量能够增加, 即dI/dt>0, 由此可得βS0/(d1+v)>1, 其中S0表示感染初期易感宿主的密度。值得注意的是, 不等式左边也是传染病动力学中的基本再生数(R0), 所以此条件实际上也表示了传染病爆发的条件, 即基本再生数大于1。让方程(2)的右边等于零, 可以计算该系统的平衡点。特别地, 由
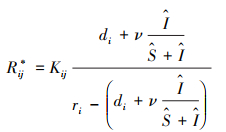
|
(3) |
式中, 帽子符号“∧”表示平衡点时的种群密度。由此可见, 病原体感染抬高了物种对资源的最低需求量, 使得宿主的零等倾线上移, 并且此时的最低需求量取决于感染者的比例(I/(S+I))和病原体的毒性(v)。这意味着病原体胁迫使得该种群对资源的竞争力减弱, 处于平衡态的种群密度更低。同时, 种群密度降低伴随着资源消耗的减少, 此时物种1的消费矢量为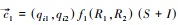
虽然感染导致宿主的零等倾线上移, 降低了宿主种群的密度, 但是研究发现病原体不会改变共存物种的共存状态(单宿主病原体不会导致宿主灭绝)。在两个竞争物种稳定共存的生态系统中(图 1中左图的深色区域), 病原体试图入侵宿主种群, 只会出现两种情况:1.病原体无法入侵(R0 < 1), 两共存物种的稳定续存不受影响;2.病原体入侵成功(R0>1), 此时描述两共存物种种群动态的方程从方程(1)变为方程(2)。令方程(2)中dI/dt=0可得易感者的平衡点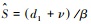
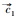
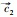

|
| 图 2 图解分析病原体感染对种间资源竞争的影响 Fig. 2 Graphical representation of infection affecting resource competition 灰色虚线为病原体入侵前物种1的零等倾线和两物种的消费矢量, 黑色实线为病原体入侵后物种的零等倾线和消费矢量 |

|
| 图 3 病原体感染对竞争动态的影响 Fig. 3 The effect of infection on competition dynamics 寄生相关参数取值:β=0.02, ν=0.1;左图资源供给点:R10=1, R20=0.8;右图资源供给点:R10=1, R20=1 |
接下来, 进一步探讨在资源供给使两个竞争物种发生竞争排斥的生态系统中, 病原体入侵后带来的影响。在没有病原体存在的情况下, 资源供给点sp2位于两物种消费矢量所在直线夹角外, 两物种无法共存, 物种2被竞争排斥。但是当病原体入侵宿主种群后, 感染使宿主零等倾线上移。如果物种1的零等倾线上移幅度足够大, sp2就能位于两物种消费矢量所在直线之间, 此时两物种可以共存(图 2右图)。根据感染后物种对资源的最低需求量(式3), 零等倾线上移幅度取决于感染者的比例和病原体的毒性, 这与生态系统的资源供给状况(资源供给点在资源平面上的位置)及宿主-病原体关系(病原体相关参数)有关, 它们共同决定病原体能否促进物种共存。
3 计算机模拟为了验证上述分析结果且更直观地展示病原体对种间资源竞争的影响, 借助软件Matlab R2021b, 基于方程(2)使用牛顿迭代法进行数值模拟, 每次模拟均使用了足够长的时间, 使得所有物种达到平衡。模拟的初值设定为:S=50, I=0, X2=50, R1=0.5, R2=0.5, 当竞争者数量达到平衡后引入少量的感染者, 以此来观察病原体对物种间资源竞争结果的影响。图(3)给出了两个典型的例子:在无感染时两物种可以共存的资源供给点, 病原体的入侵(倒三角形‘▼’指出入侵点)使宿主以更低的密度、非宿主竞争者以更高的密度共存, 感染不会改变共存结果(图 3左图);在某些无感染时非宿主竞争者被竞争排斥的资源供给点, 竞争劣势的物种2会灭绝, 但病原体感染物种1后降低了其竞争力, 物种2因此能入侵物种1种群(表现为图 3右图中物种2种群的逐渐恢复), 最终与物种1稳定共存。
进一步模拟了资源供给点和病原体相关参数(传播率β和毒性ν)对病原体胁迫下资源竞争的影响。首先, 研究在不同资源供给点上, 病原体感染对物种共存的最终影响。图 4中的左图给出没有病原体感染时模拟得出的资源平面图, 其结果与图 1中的左图一致, 只有在两物种消费矢量的所在直线之间(紫色区域)两物种才能共存。图 4中的右图是存在病原体入侵的模拟结果, 结果表明病原体的入侵增加了竞争物种在资源平面上的共存范围(紫色区域变大)。对比图 4中的两图, 进一步印证了上文的分析结果:对于两物种原本可以共存的资源供给区域(图 4左图的紫色区域), 病原体的入侵不会改变两物种的共存结果;对原本非宿主竞争者被竞争排斥的某些资源供给区域, 病原体的入侵改变了两物种的竞争结果, 促进了物种共存(图 4右图中增加的紫色区域即是符合图 2中‘sp2’的资源供给点)。然而病原体的存在对物种共存的促进不是必然的, 仍有很大一块资源供给区域(红色框内的橙色区域), 病原体能够成功入侵宿主种群, 但是两物种不能共存。在这块资源供给区域, 病原体对物种1的影响不足以让物种2摆脱竞争排斥(病原体导致宿主的零生长等倾线上移不足以满足产生图 2中‘sp2’点的条件)。

|
| 图 4 资源平面的数值模拟结果 Fig. 4 Numerical simulations of resources plane 黑色实线是无感染时物种的零等倾线; 黑色虚线是物种消费矢量的所在直线; 紫色区域是两物种能共存的资源供给区域, 橘黄色区域是仅物种1存在的资源供给区域, 深蓝色区域是仅物种2存在的资源供给区域; 红色框内区域代表病原体可以入侵的资源供给区域(满足R0>1); 右图中寄生相关参数取值:β=0.02, ν=0.1 |
其次, 病原体相关特征决定了疾病的传播和疾病对宿主的影响, 最终影响竞争结果。因此, 又在资源平面图上对不同的病原体相关参数进行模拟。根据R0=βS0/(d1+v), 可以发现传播率的增长有利于疾病的传播, 而毒性的增长不利于疾病的传播。图 5上半部分展示了不同传播率对竞争物种在资源平面上生存情况的影响, 传播率越大越有助于病原体的传播, 促进竞争物种共存的可能性越大。而随着毒性的增大, 病原体促进竞争物种共存的可能性先增后减(图 5下半部分)。这是因为:低毒性虽然有利于病原体的传播, 但是对宿主的影响较小(零等倾线上移程度不够), 不足以使物种2摆脱竞争排斥;高毒性不利于病原体的传播, 难以入侵宿主种群, 对宿主竞争的影响自然小。所以具有较大传播率且毒性中等的单宿主病原体最有可能促进物种共存。

|
| 图 5 不同传播率和毒性对宿主资源竞争的影响 Fig. 5 Effects of different transmission rate and virulence rate on resource competition |
病原体在生态系统中的作用往往被低估和轻视[10, 17], 但是病原体胁迫的影响可能是显著的, 它影响着宿主的种群动态、种间关系和生态系统中的物质循环和能量流动, 是生物多样性和食物网稳定性的重要驱动因素[7, 24—25]。病原体胁迫下的种间竞争动态早已在生态学领域中有所研究[7—8], 然而已有的理论研究大都基于现象模型[15, 19—20, 26]。此类现象模型忽略了资源利用过程, 以竞争系数这个抽象的参数来笼统地代表具体机制对物种共存的影响, 既没有具体说明物种在争夺什么, 也没有说明物种是如何竞争的[27]。由于资源竞争过程是通过竞争者利用共同的有限资源发生的, 寄生对种间资源竞争的影响很可能是因为改变了宿主的资源利用方式, 因此很难抛开竞争者-资源动态去理解寄生者对竞争系统是如何产生影响的。本文基于资源竞争理论, 耦合易感者-感染者(SI)传染病模型进行分析。分析表明, 单宿主病原体倾向于促进竞争物种共存, 这是通过病原体对宿主资源利用的影响而间接产生的。病原体降低了宿主对资源的消耗率, 抬高了对资源的最低需求, 使得宿主竞争力减弱。虽然这个过程威胁到宿主的生存, 但是单宿主病原体不会改变共存物种原本的共存状态。并且, 单宿主病原体可以使宿主的竞争对手更容易入侵, 形成共存局面, 极大地扩大了竞争物种共存的参数范围, 本质上促进了物种多样性维持。本文的另一研究结果表明单宿主病原体的传播率越大越有可能促进物种共存, 而中等强度毒性最能促进物种共存。
结果拓宽了有利于物种共存的条件, 这对入侵生物学来说可能有着重要的启迪作用。Torchin等[28]以及Mitchell和Power[29]对多种动植物的调查研究发现, 群落内的地方性疾病大多无法感染外来入侵种, 但会调节现有的本地宿主, 这就可能会引起天敌逃逸的发生。即使入侵者可能对新栖息地的适应能力较差、竞争能力较弱, 成功入侵也有可能发生, 反之, 如果没有这些地方性疾病, 入侵种就不可能成功入侵[30—33]。这在本质上是病原体对本地种的负面影响间接影响了两物种的竞争, 所以在生物入侵防控工作中考虑病原体的间接效应可能是必要的[32, 34]。
本文基于易感者-感染者疾病模型框架考虑了接触传播的单宿主寄生类型, 然而现实中病原体与宿主的相互作用非常复杂[35], 比如多宿主寄生、环境传播的寄生、易感者-感染者-恢复者(SIR)传播模式等, 这些复杂的疾病传播类型对生态系统结构和功能的影响无疑是未来生态学研究的巨大挑战。
| [1] |
莫冉, 宋卫信, 李群, 张锋. 代谢互养关系在维持微生物物种多样性中的作用. 生态学报, 2021, 41(16): 6506-6512. |
| [2] |
李远智, 肖俊丽, 刘翰伦, 王酉石, 储诚进. 生物间高阶相互作用研究进展. 生物多样性, 2020, 28(11): 1333-1344. |
| [3] |
Buck J C. Indirect effects explain the role of parasites in ecosystems. Trends in Parasitology, 2019, 35(10): 835-847. DOI:10.1016/j.pt.2019.07.007 |
| [4] |
储诚进, 王酉石, 刘宇, 蒋林, 何芳良. 物种共存理论研究进展. 生物多样性, 2017, 25(4): 345-354. |
| [5] |
Chesson P. Mechanisms of maintenance of species diversity. Annual Review of Ecology and Systematics, 2000, 31: 343-366. DOI:10.1146/annurev.ecolsys.31.1.343 |
| [6] |
Hudson P, Greenman J. Competition mediated by parasites: biological and theoretical progress. Trends in Ecology & Evolution, 1998, 13(10): 387-390. |
| [7] |
Hatcher M J, Dick J T A, Dunn A M. How parasites affect interactions between competitors and predators. Ecology Letters, 2006, 9(11): 1253-1271. DOI:10.1111/j.1461-0248.2006.00964.x |
| [8] |
Mordecai E A. Pathogen impacts on plant communities: unifying theory, concepts, and empirical work. Ecological Monographs, 2011, 81(3): 429-441. DOI:10.1890/10-2241.1 |
| [9] |
Grover J P. Stoichiometry, herbivory and competition for nutrients: simple models based on planktonic ecosystems. Journal of Theoretical Biology, 2002, 214(4): 599-618. DOI:10.1006/jtbi.2001.2488 |
| [10] |
Hatcher M J, Dick J T, Dunn A M. Diverse effects of parasites in ecosystems: linking interdependent processes. Frontiers in Ecology and the Environment, 2012, 10(4): 186-194. DOI:10.1890/110016 |
| [11] |
Lafferty K D, Allesina S, Arim M, Briggs C J, De Leo G, Dobson A P, Dunne J A, Johnson P T J, Kuris A M, Marcogliese D J, Martinez N D, Memmott J, Marquet P A, McLaughlin J P, Mordecai E A, Pascual M, Poulin R, Thieltges D W. Parasites in food webs: the ultimate missing links. Ecology Letters, 2008, 11(6): 533-546. DOI:10.1111/j.1461-0248.2008.01174.x |
| [12] |
Farrell M J, Stephens P R, Berrang-Ford L, Gittleman J L, Davies T J. The path to host extinction can lead to loss of generalist parasites. Journal of Animal Ecology, 2015, 84(4): 978-984. DOI:10.1111/1365-2656.12342 |
| [13] |
Hatcher M J, Dunn A M. Parasites in ecological communities: from interactions to ecosystems. Cambridge: Cambridge University Press, 2011. |
| [14] |
Lafferty K D, Kuris A M. Trophic strategies, animal diversity and body size. Trends in Ecology & Evolution, 2002, 17(11): 507-513. |
| [15] |
Fenton A, Brockhurst M A. The role of specialist parasites in structuring host communities. Ecological Research, 2008, 23(5): 795-804. DOI:10.1007/s11284-007-0440-6 |
| [16] |
Begon. M, Townsend C R, Harper J L. Ecology: From Individuals to Ecosystems. 4th ed. Oxford: Blackwell Publishing, 2005.
|
| [17] |
Hudson P J, Dobson A P, Lafferty K D. Is a healthy ecosystem one that is rich in parasites?. Trends in Ecology & Evolution, 2006, 21(7): 381-385. |
| [18] |
Schall J J. Parasite-mediated competition in Anolis lizards. Oecologia, 1992, 92(1): 58-64. DOI:10.1007/BF00317262 |
| [19] |
Kirk D, Shea D, Start D. Host traits and competitive ability jointly structure disease dynamics and community assembly. The Journal of Animal Ecology, 2019, 88(9): 1379-1391. DOI:10.1111/1365-2656.13028 |
| [20] |
Yan G Y. Parasite-mediated competition: a model of directly transmitted macroparasites. The American Naturalist, 1996, 148(6): 1089-1112. DOI:10.1086/285973 |
| [21] |
MacArthur R H. Geographical ecology: patterns in the distribution of species. Princeton: Princeton University Press, 1972.
|
| [22] |
Tilman D. Resources: a graphical-mechanistic approach to competition and predation. The American Naturalist, 1980, 116(3): 362-393. DOI:10.1086/283633 |
| [23] |
Tilman D. Resource competition and community structure. Princeton: Princeton university press, 1982.
|
| [24] |
Wood C L, Johnson P T. A world without parasites: exploring the hidden ecology of infection. Frontiers in Ecology and the Environment, 2015, 13(8): 425-434. DOI:10.1890/140368 |
| [25] |
Stringer A P, Linklater W. Everything in moderation: principles of parasite control for wildlife conservation. BioScience, 2014, 64(10): 932-937. DOI:10.1093/biosci/biu135 |
| [26] |
Greenman J V, Hudson P J. Parasite-mediated and direct competition in a two-host shared macroparasite system. Theoretical Population Biology, 2000, 57(1): 13-34. DOI:10.1006/tpbi.1999.1435 |
| [27] |
Miller T E, Burns J H, Munguia P, Walters E L, Kneitel J M, Richards P M, Mouquet N, Buckley H L. A critical review of twenty years & apos; use of the resource-ratio theory. The American Naturalist, 2005, 165(4): 439-448. DOI:10.1086/428681 |
| [28] |
Torchin M E, Lafferty K D, Dobson A P, McKenzie V J, Kuris A M. Introduced species and their missing parasites. Nature, 2003, 421(6923): 628-630. DOI:10.1038/nature01346 |
| [29] |
Mitchell C E, Power A G. Release of invasive plants from fungal and viral pathogens. Nature, 2003, 421(6923): 625-627. DOI:10.1038/nature01317 |
| [30] |
张黎华, 冯玉龙. 外来入侵杂草的生物防治及生防因子对本地非目标种的影响. 生态学报, 2007, 27(2): 802-809. |
| [31] |
Prenter J, MacNeil C, Dick J T A, Dunn A M. Roles of parasites in animal invasions. Trends in Ecology & Evolution, 2004, 19(7): 385-390. |
| [32] |
Dunn A M, Torchin M E, Hatcher M J, Kotanen P M, Blumenthal D M, Byers J E, Coon C A C, Frankel V M, Holt R D, Hufbauer R A, Kanarek A R, Schierenbeck K A, Wolfe L M, Perkins S E. Indirect effects of parasites in invasions. Functional Ecology, 2012, 26(6): 1262-1274. DOI:10.1111/j.1365-2435.2012.02041.x |
| [33] |
Chalkowski K, Lepczyk C A, Zohdy S. Parasite ecology of invasive species: conceptual framework and new hypotheses. Trends in Parasitology, 2018, 34(8): 655-663. DOI:10.1016/j.pt.2018.05.008 |
| [34] |
Hatcher M J, Dick J T A, Dunn A M. Parasites that change predator or prey behaviour can have keystone effects on community composition. Biology Letters, 2014, 10(1): 20130879. DOI:10.1098/rsbl.2013.0879 |
| [35] |
Keeling M J, Rohani P. Modeling infectious diseases in humans and animals. Princeton: Princeton University Press, 2008.
|