 2023, Vol. 43
2023, Vol. 43文章信息
- 韦冰, 豆鹏鹏, 王芳, 姚婧梅, 林敦梅, 智悦, 何强, 黄屿玥, 史思捷, 冯莹, 王洪锐, 裴文杰
- WEI Bing, DOU Pengpeng, WANG Fang, YAO Jingmei, LIN Dunmei, ZHI Yue, HE Qiang, HUANG Yuyue, SHI Sijie, FENG Ying, WANG Hongrui, PEI Wenjie
- 底栖分解类群对金佛山森林溪流凋落物混合分解的影响
- Effects of benthic decomposers on the decomposition of litter mixtures in a forest headwater stream, Jinfo Mountain stream
- 生态学报. 2023, 43(17): 7317-7330
- Acta Ecologica Sinica. 2023, 43(17): 7317-7330
- http://dx.doi.org/10.20103/j.stxb.202207202082
-
文章历史
- 收稿日期: 2022-07-20
- 网络出版日期: 2023-04-28





2. 中国农业大学, 草业科学与技术学院, 北京 100193
2. College of Grassland Science and Technology, China Agricultural University, Beijing 100193, China
凋落物输入是高荫蔽度、低初级生产力的源头溪流中能量的主要来源[1—2], 其分解过程对于维持溪流生态系统碳和养分平衡有着重要的意义[3—4]。凋落物在自然界往往以混合物的形式存在, 受到不同分解者类群(微生物和大、小型底栖动物)的共同影响[5]。当今人类活动显著降低了植物的多样性, 混合凋落物的分解受到越来越多学者的重视, 相关研究在欧美起步较早, 在区域上比较局限, 以其所在的温带溪流为主, 形成了以橡树(Quercus robur)和桤木(Alnus glutinosa)为代表性凋落叶种的系统性研究, 而对热带亚热带溪流的研究较少[6—7]。国内的凋落物分解研究主要集中在陆地生态系统, 关于溪流生态系统的研究起步较晚[8]。研究表明, 不同区域内气候不同, 地带性植被类型不同, 凋落物的分解速率差异明显。相较于温带溪流, 热带地区凋落物分解速率较快, 微生物的分解活动更加强烈[9]。中国的亚热带地区约占全国国土面积的1/4, 是中国代表性气候带之一。但混合凋落物在亚热带的相关研究鲜见, 凋落物分解混合效应的机制不明, 特别是不同分解者对混合凋落物分解的贡献。这极大地制约了对亚热带溪流生态系统结构和功能的理解和认识。
混合凋落物的分解与单种叶片分解的简单加和不同, 可能出现正或负的非加和效应(协同或者拮抗), 即质量损失实际值高于或者低于以单种叶片平均分解速率推测的预期值[10]。这是由于随着混合凋落物种类的增多, 分解者可利用的资源增加, 凋落物分解加快[11];具有特殊性状的优势凋落叶种促进或减慢了其他物种的分解[12]。此外, 当不同种类凋落物混合时, 养分或者某些次生代谢物质可能在凋落物间发生转移, 从而影响分解过程中分解者的交互作用[13]。因此, 不同分解者类群参与的混合分解更为复杂。
本研究选取了以中国重庆市南川区金佛山源头溪流为代表的亚热带溪流, 以河岸3种常见乔木叶凋落物(八角枫(Alangium chinense)、缺萼枫香(Liquidambar acalycina)、薄叶润楠(Machilus leptophylla))为对象, 进行原位分解实验, 分析不同分解者类群参与叶凋落物在源头河流中的混合分解效应, 以期更好地阐明亚热带源头溪流凋落叶分解的特征与变化规律, 为我国亚热带水系, 乃至全球类似水系的生态修复提供参考及理论依据。
1 研究区概况与方法 1.1 研究区概况金佛山溪流位于金佛山国家级自然保护区内(28°46′—29°38′N, 106°54′—107°27′E), 发源于金佛山东北侧, 为大溪河右岸一级河流。金佛山地属亚热带湿润季风气候, 年均气温8.3—12.4℃, 年均降水量1164—1382mm, 年均日照1091.6h。河岸带植被主要有方竹(Chimonobambusa quadrangularis)、川黔鹅耳枥(Carpinus fangiana)、缺萼枫香(Liquidambar acalycina)、簇叶新木姜子(Neolitsea confertifolia)、香桂(Neolitsea confertifolia)、灯台树(Cornus controversa)八角枫(Alangium chinense)、水青冈(Fagus longipetiolata)等[13]。本实验地点海拔约1000m, 几乎不受人为活动干扰, 水质良好(表 1)。
| 研究地 Site |
水温/℃ Temperature |
溶解氧 Dissolved oxygen/ (mg/L) |
pH | 总氮 Total nitrogen/ (mg/L) |
总磷 Total phosphorus/ (mg/L) |
NH4+ Ammonia nitrogen/ (mg/L) | NO3- Nitrate/ (mg/L) | PO43- Phosphate/ (mg/L) | 电导率 Conductivity/ (μS/cm) |
| 金佛山 | 10.2±0.13 | 9.7±0.32 | 7.9±0.18 | 1.9±0.13 | 0.14±0.01 | 0.2±0.07 | 0.8±0.48 | 0.04±0.01 | 181±8.4 |
于2017年8—9月收集金佛山溪流河岸带3种理化性质差异显著的常见树种叶片凋落物(八角枫、缺萼枫香和薄叶润楠), 去除破损和病虫害的叶片, 在60℃烘箱中干燥48h备用。
1.2.2 凋落物原位分解实验称取烘干后的叶凋落物4.2g(精确到0.1g)装入分解袋内(15cm×10cm)中, 共计126个分解袋(7种凋落物处理×3种孔径×6个重复)。分解袋有3种孔径, 分别是小孔径0.05mm(只允许微生物进出分解袋)、中孔径0.25mm(允许微生物和小型底栖动物进出)和大孔径2mm(允许所有分解者(微生物、小型底栖动物和大型底栖动物)进出)。7种凋落物处理中包括单种凋落叶3组(即A:八角枫4.2g;B:缺萼枫香4.2g;C:薄叶润楠4.2g)和4组混合物种凋落叶(即D组合:八角枫和缺萼枫香各2.1g;E组合:八角枫和薄叶润楠各2.1g;F组合:缺萼枫香和薄叶润楠各2.1g;G组合:3个物种各1.4g)。将分解袋绑在尼龙网上, 并用石块将尼龙网随机固定在河床上。于2017年11月17日放置分解袋, 2018年1月12日回收, 用自封袋收集每一个分解袋, 低温保存运回实验室。取样同时测定了溪水的理化指标, 并采集了水样和底栖动物样品。
1.2.3 水体理化性质与凋落物初始性状测试野外使用多参数传感器(YSI Professional plus, Yellow Spring, 美国)测定溪水的温度、pH值、电导率和溶解氧并收集水样冷藏运回实验室。
室内利用碱性高锰酸钾法测定总氮(TN)和总磷(TP)、纳氏试剂光度法测定氨氮、紫外分光光度法测定硝酸盐及钼锑抗分光光度法测定磷酸盐等[14]。测定的凋落叶初始理化性质, 使用范氏洗涤法测定木质素、纤维素含量[15];使用拉力计(ZP-50, 艾固, 中国香港)测定韧性, 采用撕裂单位宽度叶片所用的力作为韧性值;使用TOC-TN分析仪(岛津, 日本)高温灼烧测定有机碳含量;TN、TP先用浓硫酸-过氧化氢消解粉碎的样品, 然后用TOC-TN测定氮含量, 使用钼锑抗比色法测定磷含量。根据测定结果计算碳氮比(C/N)、碳磷比(C/P)、氮磷比(N/P)与木质素/N。
1.2.4 底栖动物的分离与鉴定将两种孔径的索博网(30cm×30cm)以小孔径(0.05mm)在外、大孔径(0.25mm)在内的方式组装, 在各实验点内分别随机收集3个沉积物平行样本[16]。样品在75%的酒精中保存, 运回实验室进行分离、鉴定和计数, 分类鉴定到尽可能低的分类水平。根据采样面积, 换算出每种底栖动物的密度(ind/m2)。根据Tachet等[17]的分类方法将大型底栖动物划分为不同的摄食功能类群:食碎屑者、撕食者、刮食者、滤食者、钻食者、捕食者和寄生者。本研究按照体长, 以Giere等[18]的分类研究为依据划分了大型和小型底栖动物, 将能通过0.25mm但不能通过0.05mm孔径筛网的底栖动物划分为小型底栖动物, 将能通过2mm但不能通过0.25mm孔径筛网的为大型底栖动物。
1.2.5 数据分析根据凋落物分解前后的质量差异, 计算凋落物的质量损失率:
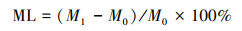
|
(1) |
式中, ML为质量损失率;M0和M1分别为凋落物分解前后的重量。混合凋落物的期望质量损失率由平均单种物种的凋落物质量损失率算出, 与其实际质量损失率计算凋落物分解的混合效应, 从而探究组成物种能否预测混合凋落物分解的质量损失[19]。
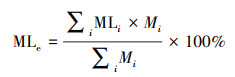
|
(2) |
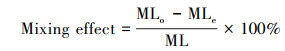
|
(3) |
式中, MLe为混合凋落物质量损失期望值;MLi为物种i的质量损失;Mi为组合中物种i的重量;MLo为混合凋落物实际的质量损失。Seastedt等[20]研究量化了微生物、小型和大型底栖动物对叶片重量损失的平均贡献:
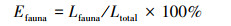
|
(4) |
式中, Efauna(%)是某种分解者对叶凋落物重量损失的影响;Lfauna是某种分解者造成的重量损失百分比, 不同孔径分解袋中凋落物重量损失的差异得出的;Ltotal是大孔分解袋中重量损失百分比, 是由微生物、底栖动物和其他效应导致的叶片分解。
选用混合凋落物性状的群落加权平均值(CWM)作为相应指数[21]:
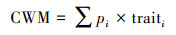
|
(5) |
式中, pi是物种i在混合凋落物中所占的比例;traiti是物种i的性状值。
混合凋落物的性状多样性由功能分散指数(FDis)表示, 能描述多维特征空间中组成物种到混合物种质心的平均距离的功能多样性分量[22], 用R软件中“FD”包计算。
混合凋落物物种丰富度对质量损失的影响可使用非参数组间差异的Kruskal-Wallis检验;配对t检验探究混合凋落物处理中的混合效应是否显著;混合凋落物处理与分解袋孔径及其交互作用对质量损失和混合效应的影响使用双因素方差分析探究, 并对质量损失进行事后检验(Tukey HSD检验)。对于混合凋落物的选择效应和互补效应, 使用Loreau等[23]的公式计算。加权性状、性状多样性等解释变量对质量损失、混合效应等响应变量的影响采用“lme4”包中的线性混合效应模型分析[24]。所有分析均在R软件(R version 3.4.2)中完成, P<0.05定为统计上差异显著。
2 结果 2.1 凋落叶的理化性质三种叶凋落物的初始理化性质差异显著(表 2):薄叶润楠的P含量最低, C含量及韧性最高, 同时, 其N/P、木质素/N比值最高, 在三种叶片中基质质量最低, 可降解性最差;缺萼枫香叶的C、P、木质素含量与韧性, 以及C/P、N/P介于其它两个物种之间, 叶片基质质量和可降解性居中;八角枫的N、P含量最高, 而结构性化合物(木质素和纤维素)含量最低, 其叶片基质质量较高, 更容易分解。
| 性状 Traits |
八角枫 Alangium chinense |
缺萼枫香 Liquidambar acalycina |
薄叶润楠 Machilus leptophylla |
| C/(mg/g) | 364.86±2.53b | 363.42±3.64b | 400.85±2.20a |
| N/(mg/g) | 15.52±0.64a | 9.90±0.17c | 12.23±0.29b |
| P/(mg/g) | 1.89±0.02a | 0.99±0.06b | 0.96±0.04b |
| 木质素Lignin/(mg/g) | 38.08±.52c | 101.99±12.90b | 255.21±9.95a |
| 纤维素Cellulose/(mg/g) | 89.37±9.04b | 177.93±17.88a | 176.10±8.62a |
| 韧性Toughness/(N/mm) | 0.24±0.02b | 0.26±0.02b | 0.35±0.03a |
| 木质素/N Lignin/N | 2.46±0.10c | 10.29±1.24b | 20.89±1.00a |
| 碳氮比C/N | 23.60±1.14c | 36.74±1.00a | 32.82±0.94b |
| 碳磷比C/P | 193.29±2.94c | 370.10±20.25b | 419.15±15.52a |
| 氮磷比N/P | 8.22±0.29c | 10.10±0.70b | 12.82±0.86a |
| C:碳含量;N:氮含量;P: 磷含量; 每行的不同字母表示差异显著 | |||
对4个混合处理凋落物的各初始加权性状进行主成分分析(图 1), 前两轴分别解释了总体变异的79.1%和20.1%, 二者累计贡献率为99.2%。第一轴与凋落叶N、P含量呈负相关, 与C含量、纤维素含量、木质素含量、木质素/N、C/N、和C/P正相关。第二轴与凋落叶C含量、N含量、P含量、木质素含量、韧性、木质素/N、N/P呈负相关, 与纤维素含量、C/N、C/P呈正相关。

|
| 图 1 四个混合处理初始加权性状的主成分分析 Fig. 1 Principal component analysis of initial community trait of four mixture treatments CWM:性状群落加权均值CommunityWeighted Mean;C:碳含量;N:氮含量;P:磷含量;AL:八角枫+缺萼枫香Alangium chinense+Liquidambar acalycina;AM:八角枫+薄叶润楠Alangium chinense+Machilus leptophylla;LM:缺萼枫香+薄叶润楠Liquidambar acalycina+Machilus leptophylla;ALM:八角枫+缺萼枫香+薄叶润楠Alangium chinense+Liquidambar acalycina+Machilus leptophylla |
凋落物分解的质量损失受凋落叶物种组成(F3, 36= 30.61, P<0.0001)和分解者类群(F2, 36=35.68, P<0.0001)的显著影响, 其中物种组成的解释度最大(表 3)。凋落物的混合效应受凋落叶物种组成(F2, 36= 3.43, P<0.05)和分解者类群(F3, 36= 36.95, P<0.0001)的显著影响(表 4)。
| 项目 Projects |
离差平方和 Sum of squares |
自由度 df |
均方 Mean-square |
F | 占变异的百分比/% % of total variation |
P |
| 物种组成Species composition | 3750 | 3 | 1250 | 30.61 | 44.4 | <0.0001 |
| 分解袋孔径Mesh size | 2915 | 2 | 1458 | 35.68 | 34.51 | <0.0001 |
| 交互作用Interaction | 311.2 | 6 | 1270 | 1.270 | 3.684 | 0.2955 |
| 误差Error | 1470 | 36 | 40.85 |
| 项目 Projects |
离差平方和 Sum of squares |
自由度 df |
均方 Mean-square |
F | 占变异的百分比/% % of total variation |
P |
| 物种组成Species composition | 2144 | 3 | 714.5 | 3.430 | 7.913 | <0.05 |
| 分解袋孔径Mesh size | 15395 | 2 | 7697 | 36.95 | 56.83 | <0.0001 |
| 交互作用Interaction | 2052 | 6 | 341.9 | 1.641 | 7.573 | 0.1642 |
| 误差Error | 7500 | 36 | 208.3 |
对于单个物种而言, 凋落物的质量损失在3个物种间存在显著差异, 八角枫(Alangium chinense, 质量损失为53.05%)>缺萼枫香(Liquidambar acalycina, 质量损失为30.00%)>薄叶润楠(Machilus leptophylla, 质量损失为12.63%)(图 2)。3种叶凋落物的质量损失随孔径的增加而普遍呈上升趋势, 其中大孔分解袋中的质量损失(28.10%—88.35%)显著高于小孔分解袋(14.78%—64.08%)(图 2)。

|
| 图 2 凋落物在不同处理和分解袋孔径下的质量损失 Fig. 2 Mass loss of litter under different treatments and mesh sizes A:八角枫Alangium chinense;B:缺萼枫香Liquidambar acalycina;C:薄叶润楠Machilus leptophylla;D:八角枫+缺萼枫香Alangium chinense+Liquidambar acalycina;E:八角枫+薄叶润楠Alangium chinense+Machilus leptophylla;F:缺萼枫香+薄叶润楠Liquidambar acalycina+Machilus leptophylla;G:八角枫+缺萼枫香+薄叶润楠Alangium chinense+Liquidambar acalycina+Machilus leptophylla; 不同字母表示差异显著; *:P<0.05;***:P<0.001 |
在混合凋落物DEFG组合中, 叶凋落物的质量损失随孔径的增加呈上升趋势:相比于小孔径, D(八角枫+缺萼枫香)、E(八角枫+薄叶润楠)、F(缺萼枫香+薄叶润楠)和G(八角枫+缺萼枫香+薄叶润楠)在大孔径中分别上升了21.33%, 12.98%, 15.53%和30.27%。
2.2.3 混合凋落物分解的混合效应4种凋落物组合在3种不同分解袋孔径下共计12种混合处理, 0.05mm孔径即仅微生物参与分解的处理的实际质量损失低于预期质量损失, 表现为负的非加和效应, 其中3个为显著负的非加和效应:八角枫+缺萼枫香(D, P<0.05)、八角枫+薄叶润楠(E, P<0.05)、八角枫+缺萼枫香+薄叶润楠(G, P<0.001)。底栖动物加入后(孔径为0.25mm和2mm)表现为正的非加和效应, 即实际质量损失显著高于预期质量损失, 其中2个处理为显著正的非加和效应, 缺萼枫香+薄叶润楠(F, P<0.05)、八角枫+缺萼枫香+薄叶润楠(G, P<0.0001, 图 3)。

|
| 图 3 不同凋落物处理分别在3种分解袋孔径中(0.05mm、0.25mm和2mm) 的质量损失 Fig. 3 Mass loss of litter in three mesh size (0.05mm, 0.25mm and 2mm) with treatment *: P < 0.05;**: P < 0.01;***: P < 0.001 |
如表 5所示, 在金佛山溪流的凋落物分解实验中共发现了32个大型底栖动物类群, 平均密度为2072个/m2。大部分底栖动物属于昆虫纲(97.4%), 极少部分为蛛形纲(2.1%)和腹足纲(0.4%)。其中, 绿襀科、扁蜉科、溪尼甲科以及摇蚊科(枝长跗摇蚊属)是撕食叶片的优势类群。根据鉴定出的大型底栖动物, 共划分出六个摄食群功能: 刮食者(49.1%), 其他包括撕食者(18.2%)、食碎屑者(17.5%)、捕食者(9.9%)、滤食者(5.2%)和钻食者(0.1%);共发现16个小型底栖动物类群, 平均密度为9529个/m2。其中, 小型摇蚊科幼虫占比最高, 达到59.2%, 其次是线虫(26.5%)和小型蜉蝣目幼虫(5.6%)。
| 类群 Taxa |
功能摄食群 Functional feeding group |
丰度/% Abundance |
|
| 大型底栖动物 | 昆虫纲Insecta | 97.43 | |
| Macroinvertebrates | 鞘翅目Coleoptera | 18.02 | |
| 溪尼甲科Elmidae | |||
| Limmius | SC/SH | 15.45 | |
| Macronychus | SC/SH | 0.24 | |
| Elmis | SC/SH | 0.14 | |
| Esolus | SC/SH | 0.78 | |
| 长角泥甲科Elmidae | SC/SH | 0.82 | |
| 扁角泥甲科Psephenidae | SC | 0.34 | |
| 龙虱科Dytiscidae | |||
| 粒龙虱属Laccophilus | SH/H | 0.24 | |
| 蜉蝣目Epbemeroptera | 39.72 | ||
| 蜉蝣科Ephemeridae | |||
| 蜉蛇属Ephemera | |||
| 东方蜉Ephemera orientalis | SH/F | 1.46 | |
| 四节蜉科Baetidae | |||
| 四节蜉属Baetis | SC/DF | 27.25 | |
| 扁蜉科Heptageniidae | |||
| 似动蜉属Cinygmina | SC | 3.55 | |
| 三叉扁蝴蜉属Ecdyonurus | |||
| 德拉扁蜉蝣Ecdyonurus dracon | SC/SH | 6.77 | |
| 新蜉科Neoephemeridae | |||
| 新蜉属Neoephemera | DF | 0.68 | |
| 襀翅目Plecoptera | 6.13 | ||
| 绿襀科Chloroperlidae | 3.90 | ||
| Siphonoperla | SH | ||
| 襀科Perlidae | |||
| Dinocras | SH/PR | 0.43 | |
| 黑襀科Capniidae | |||
| Capnia | SH | 0.58 | |
| Capnioneura | SH | 1.22 | |
| 双翅目Diptera | 33.41 | ||
| 大蚊科Tipulidae | SH/DF/PR | 0.63 | |
| 沼大蚊科Limoniidae | SH/SC/Ht | 2.04 | |
| 摇蚊科Chironmidae | |||
| 长足摇蚊亚科Tanypodinae | |||
| Paramerina | PR | 3.90 | |
| 长跗摇蚊族Tanytarsini | |||
| 枝长跗摇蚊属Cladotanytarsus | DF/SH/SC | 11.60 | |
| 长跗摇蚊属Tanytarsus | DF/SH/SC | 2.04 | |
| 摇蚊族Chironomini | |||
| 多足摇蚊属Polypedilum | DF/SH/F | 5.17 | |
| 直突摇蚊亚科Orthocladiinae | |||
| 真开氏摇蚊属Eukiefferiella | SC/DF/F | 2.44 | |
| 毛胸摇蚊属Heleniella | SC/DF/F | 1.60 | |
| 拟毛突摇蚊属Parachaetocladius | SC/DF/F | ||
| 趋流摇蚊属Rheocricotopus | SC/DF/F | 0.78 | |
| 蠓科Cetatqogoridae | PR/DF/SH | 2.29 | |
| 舞硭科Empididae | SH | 0.14 | |
| 毛翅目Trichoptera | 0.14 | ||
| 多距石蛾科Polycentropodidae | PR/F | 0.14 | |
| 蛛形纲Arachnida | 2.14 | ||
| 真螨目Acariformes | |||
| 水螨Hydrachnidia | PR | 2.14 | |
| 腹足纲Gastropoda | 0.43 | ||
| 基眼目Basommatophora | |||
| 觿螺科Hydrobiidae科Hydrobi idae | SC/SH/F | 0.29 | |
| 扁卷螺科Planorbidae | SC/SH | 0.14 | |
| 小型底栖动物 | 介型纲Ostracoda | F | 4.30 |
| Meioinvertebrates | 线虫Nematode | DF/SH/P | 26.46 |
| 桡足类Copepoda | 2.14 | ||
| 猛水蚤 Harpacticoida | DF | 1.95 | |
| 剑水蚤Cyclopoida | DF | 0.19 | |
| 寡毛纲Oligochaeta | DF/SC | 0.22 | |
| 昆虫纲Insecta | 65.24 | ||
| 鞘翅目Coleoptera | 0.22 | ||
| Elmidae | |||
| Limmius | SC/SH | 0.19 | |
| 龙虱科Dytiscidae | |||
| 粒龙虱属Laccophilus | SH/PI | 0.02 | |
| 蜉蝣目Ephemeroptera | 5.63 | ||
| 蜉蝣科Ephemeridae | |||
| 蜉蝣属Ephemera | |||
| 东方蜉Ephemera orientalis | SH/F/DF/PR | 0.05 | |
| 四节蜉科Baetidae | |||
| 四节蜉属Baetis | SC/DF | 0.90 | |
| 扁蜉科Heptageniidae | |||
| 似动蜉属Cinygmina | SC | 1.14 | |
| 假蜉属Iron | SC | 3.48 | |
| 自扁蜉蝣属Heptagenia | SC/SH | 0.05 | |
| 襀翅目Plecoptera | 0.18 | ||
| 绿襀科Chloroperlidae | |||
| Siphonoperla | SH | 0.18 | |
| 双翅目Diptera | 59.20 | ||
| 摇蚊科Chironmidae | SC/DF/PR | 59.05 | |
| 蠓科Cetatqogoridae | PR/DF/SH | 0.15 | |
| 蛛形纲Arachnida | 1.64 | ||
| 真螨目Acariformes | |||
| 水螨Hydrachnidia | PR | 1.64 | |
| DF:食碎屑者Deposit feeder;SH:撕食者Shredder;SC:刮食者Scrapers;F:滤食者Filterfeeder;PI:钻食者Piercer;PR:捕食者Predator | |||
小型和大型底栖动物加入后, 凋落物的质量损失均增加(图 4)。微生物对凋落叶分解的贡献最大, 各组合均大于50%(相对贡献率为53.78%—76.38%)。单种叶片基质品质越低, 微生物贡献度越低(图 5), 三种凋落物相对贡献率分别为72.64%(八角枫)、67.82%(缺萼枫香)和53.78%(薄叶润楠)。在混合凋落物组合中, 大型底栖动物在组合D(八角枫+缺萼枫香)的相对贡献率仅为8.85%, 随着难分解叶片薄叶润楠的加入, 其对混合凋落物组合的相对贡献率也上升, 在组合E(八角枫+薄叶润楠)、F(缺萼枫香+薄叶润楠)、G(八角枫+缺萼枫香+薄叶润楠)中的贡献率分别为18.69%、13.74%和27.54%。

|
| 图 4 不同分解者类群对质量损失率的影响 Fig. 4 Impact of different decomposers on mass loss rate |

|
| 图 5 不同分解者类群的贡献度 Fig. 5 Contribution of different decomposers on mass loss |
混合凋落物初始加权性状CWM1显著影响其质量损失(图 6)。随着加权性状CWM1中N、P含量的增加, 混合凋落物质量损失增加, 混合效应减弱, 互补效应减弱, 选择效应增强;随着C含量、纤维素含量、木质素含量、木质素/N、C/N、和C/P的增加, 混合凋落物质量损失减少, 混合效应增强, 互补效应增强, 选择效应减弱(图 6, 图 7)。混合凋落物初始加权性状(CWM2)中C、N、P、木质素、韧性、木质素/N、N/P增加, 互补效应减弱, 选择效应增强(图 7)。同时, 随着混合凋落物的功能差异的增大, 互补效应增强, 选择效应减弱(图 8)。

|
| 图 6 混合凋落物初始性状对质量损失率和混合效应的影响 Fig. 6 Effect of initial traits on mass loss rate and mixing effect |

|
| 图 7 混合凋落物初始性状对互补和选择效应的影响 Fig. 7 Impact of initial traits of litter mixture on complementary and selectivity effects |

|
| 图 8 混合凋落物功能差异对互补和选择效应的影响 Fig. 8 Impact of functional divergence of litter mixture on complementary and selectivity effects |
不同物种分解速率的差异与叶凋落物的初始基质质量有关, 其中C、N、P、木质素的含量和韧性以及各营养元素间的比例是影响分解速率的重要因素[25]。本研究中叶凋落物的分解速率为八角枫>缺萼枫香>薄叶润楠(图 2)。其中, 八角枫(具有较高的N、P含量, 较低的韧性、木质素含量、木质素/N与C-营养素比值)的分解速率最快[26];而薄叶润楠反之, 与前人研究相似, 如Yue等发现柳树相比于柏树、落叶松、杜鹃木质素含量最低, 分解速率最快, 落叶松反之[27]。Gessner等[28]的研究也指出木质素含量高抑制凋落叶分解。较高的木质素含量, 会降低微生物的分解作用[29]。
在金佛山溪流中, 微生物对凋落物分解的相对贡献均大于底栖动物的贡献, 占50%以上, 豆鹏鹏等[30]的研究结果与本研究一致, 同时其他热带和温带溪流中也发现了这种现象[31]。本研究中基质质量最高的八角枫分解时, 微生物的相对贡献度最高(72.63%)(图 5), 这是由于叶片可降解性较高的凋落物(高N、P含量和低C/N、C/P)更有利于微生物定殖, 故微生物贡献度较高[32—33]。相反的, 薄叶润楠的基质质量最低、木质素含量最高, 其分解过程中大型底栖动物的相对贡献度为29.04%, 显著高于其他两种叶片。Compson等[34]在亚热带地区的研究中也发现了大型底栖动物中的撕食者偏好取食木质素含量较高的叶片。
此外, 水流因素会对凋落物分解造成影响, 流速和流量的增加能加速淋溶过程, 促使可溶性物质的溶解, 一定程度上可加速凋落物的物理破碎过程[35]。本研究使用了三种孔径的分解袋。其中, 小孔径网袋的质量损失主要是由于浸出和微生物分解造成的, 而中孔径网袋的质量损失主要是由于物理破碎、微生物分解、小型动物直接取食叶子及其与微生物的相互作用。大孔径网袋包括整个分解者群落(即微生物、小型动物和大型动物), 其中物理破碎、三种分解者群落及其交互作用可能导致质量损失[36]。本实验主要在流速为0.08—0.1m/s的低流速河段中进行。因此, 在不同孔径分解袋中, 由于水流引起的物理破碎所造成的叶质量损失在本研究中相对较小[37]。
3.2 凋落叶混合对其分解的影响 3.2.1 凋落物混合的负的非加和效应本研究中, 0.05mm(仅微生物参与)分解袋中混合凋落叶片实际质量损失均低于预期的质量损失, 呈现负的非加和效应(图 3)。这可能是由于混合凋落物中某种凋落物的叶层提取物抑制了参与凋落物分解的酶的活性, 干扰了混合凋落叶的分解[38]。此外, 分解缓慢的凋落物种类可能会阻碍分解者对较快分解成分的获取, 从而减慢混合物的分解速度, 本研究的薄叶润楠分解较慢, 其可能释放酚类等物质[38], 阻碍与其混合的八角枫和缺萼枫香等物种的分解。如, Mudrick等[39]也发现黑栎(Quercus nigra)和北美枫香(Liquidambar styraciflua)凋落物的混合, 黑栎的渗透液抑制了相邻的北美枫香的分解, 进而表现出负效应, 与本研究结论一致。
此外, 分解者类群是影响凋落物分解的关键因素。注意到, 一些昆虫的早龄虫如(摇蚊和东方蜉)因体长小于0.25mm, 被划分为小型底栖动物[40]。底栖动物的身体大小在构建群落和确定其功能上起关键作用, 小型底栖动物和大型底栖动物在沉积物和落叶的养分循环过程中可能扮演不同角色[41]。摇蚊幼虫或者其他底栖动物(如东方蜉)在不同阶段其摄食行为可能发生变化, 具有更大生物量的物种的生态系统功能更大[17, 42], 体长不同可能对凋落物分解过程的影响也不同。其中, 大型底栖动物中的撕食者会消耗粗颗粒有机物, 从而提高了落叶的分解速度[43], 对凋落物分解的影响最大[34]。0.05mm孔径的分解袋中只有微生物参与分解, 缺乏其他底栖动物(如撕食者)的协作。凋落叶在被大型动物取食时进一步破碎, 增加了叶片基质的表面积, 更有利于微生物进入叶片有营养的内部组织[44]。
3.2.2 正的非加和效应本研究发现小型底栖动物加入分解后, 分解者群落复杂度增加, 混合凋落物的质量损失为正的非加和效应, 但不显著(图 3);大型底栖动物的加入更一步促进了混合凋落物的分解, 其中缺萼枫香+薄叶润楠(F)和八角枫+缺萼枫香+薄叶润楠(G)两个混合凋落物处理的正的非加和效应显著, 其速率分别是预期的1.5倍和1.6倍(图 3)。
金佛山溪流中小型底栖动物群主要是由小型摇蚊科幼虫和线虫组成(表 5), 前者约占小型底栖动物密度的60%。小型底栖动物可能通过直接取食或间接改变微生物群落结构来影响凋落物分解过程[45]。研究表明摇蚊幼虫可能直接取食高质量的叶片, 可利用被真菌分解一定程度的凋落物作为补充的食物来源[46]。碎屑上附着的微生物还可作为与叶凋落物分解相关的摇蚊幼虫的直接食物来源[47]。此外, 小型摇蚊科幼虫可以捕食微生物, 对微生物生物量有下行效应的控制作用, 进而促进微生物群落处于一个更活跃的生长阶段, 从而增加了对有机质的需求[48]。线虫可能不直接取食叶片, 而是以叶片上的生物膜(如真菌、硅藻、细菌及其胞外分泌物)为食物来源, 可能使微生物的丰度和群落结构发生改变, 导致微生物的活性的增加[49], 从而间接影响凋落物的分解过程。Wang等[40]研究发现小型底栖动物动物提高了缺萼枫香(Liquidambar acalycina)分解速率, 小型底栖动物的下行效应使其真菌生物量和多样性分别增加了100.08%和多40%。因此, 小型底栖动物取食凋落物并增加微生物群落活跃度, 最终促进了混合凋落物的分解。
金佛山溪流中大型底栖动物主要以蜉蝣目为优势类群, 主要摄食群功能类群是刮食者(49.1%)和撕食者(18.2%)和食碎屑者(17.5%)(表 5)。其中, 撕食者对叶片有一定的直接切碎作用[34]。大型底栖动物也可通过取食微生物和对微环境的生物扰动间接影响凋落物的分解[50]。此外, 由于大型底栖动物(如水螨类)对小型底栖动物存在捕食关系, 进而控制其数量和生物量[51]。大型底栖动物进一步促进了小型底栖动物分解叶片的能力(图 4), 发挥了正效应。大型底栖动物对叶凋落物的直接摄食促进叶片分解, 这种取食行为也进一步导致叶片破碎, 进而增加叶基质的表面积, 促进微生物进入叶内部营养组织, 从而使小型底栖动物和微生物可以更容易地利用这些碎片[52]。王芳[9]研究发现大型底栖动物存在时凋落叶破碎产生细颗粒有机物, 小型底栖动物进一步利用。同时, 小型底栖动物可以利用叶屑和较小的微生物(如细菌)[53], 也可作为其他较大的底栖动物的食物来源。因此, 分解者群落的复杂性的增加对凋落物分解具有关键的正向作用[54]。
本研究表明随着混合凋落物的功能差异的增大, 互补效应增强, 选择效应减弱(图 8)。这是由于凋落物多样性主要通过互补效应促进了凋落物分解[55]。Conn等[56]也发现了橡木和松树凋落物叶的混合可以优化氮和磷等营养物质的保留, 从而促进混合凋落物分解。分解者优先利用高质量凋落物, 随后使低质量的凋落物具有了相对较高的养分有效性, 继而引起低质量凋落物分解加快并最终使混合凋落物的整体分解速率加快[10]。
4 结论(1) 单种叶凋落物的基质质量越高(高N、P含量和低C/N、C/P), 分解速率越快, 微生物贡献度越高;基质质量越低, 分解速率越慢, 底栖动物贡献度越高。
(2) 金佛山溪流凋落物的混合分解与凋落物种类和分解者类群的复杂性密切相关。混合凋落物种类增加, 物种间互补效应增强, 凋落物分解越快, 正的非加和效应增强。仅微生物参与时, 混合凋落物的分解均呈现负的非加和效应, 其中含基质质量较高凋落叶种(八角枫)的组合呈现显著负的非加和效应;小型底栖动物加入后, 混合凋落物呈现正的非加和效应, 大型底栖动物加入后含有基质质量较低凋落叶种(缺萼枫香和薄叶润楠)的组合呈现显著正的非加和效应。
(3) 本研究探明了亚热带源头溪流微生物和底栖动物对凋落物混合分解的相对贡献, 为阐明凋落物的混合效应机制提供了科学依据。
| [1] |
Vannote R L, Minshall G W, Cummins K W, Sedell J R, Cushing C E. The River continuum concept. Canadian Journal of Fisheries and Aquatic Sciences, 1980, 37(1): 130-137. DOI:10.1139/f80-017 |
| [2] |
Riskin S H, Neill C, Jankowski K J. Solute and sediment export from Amazon forest and soybean headwater streams. Ecological Applications, 2017, 27(1): 193-207. DOI:10.1002/eap.1428 |
| [3] |
Gessner M O, Chauvet E. A case for using litter breakdown to assess functional stream integrity. Ecological Applications, 2002, 12(2): 498-510. DOI:10.1890/1051-0761(2002)012[0498:ACFULB]2.0.CO;2 |
| [4] |
Moore J C, Berlow E L, Coleman D C, Ruiter P C, Dong Q, Hastings A, Johnson N C, McCann K S, Melville K, Morin P J, Nadelhoffer K, Rosemond A D, Post D M, Sabo J L, Scow K M, Vanni M J, Wall D H. Detritus, trophic dynamics and biodiversity. Ecology Letters, 2004, 7(7): 584-600. DOI:10.1111/j.1461-0248.2004.00606.x |
| [5] |
王璐. 长白山溪流冬季凋落物分解与大型底栖动物定殖[D]. 长春: 东北师范大学, 2020.
|
| [6] |
Jabiol J, Chauvet E. Biodiversity and litter decomposition: a case study in a Mediterranean stream. Freshwater Science, 2015, 34(2): 423-430. DOI:10.1086/680094 |
| [7] |
官昭瑛, 何莹, 安玉蓉, 蔡吉花, 童晓立. 外来植物凋落物分解对底栖动物多样性及其摄食功能群的影响. 生态学报, 2010, 30(11): 2828-2835. |
| [8] |
李宜浓, 周晓梅, 张乃莉, 马克平. 陆地生态系统混合凋落物分解研究进展. 生态学报, 2016, 36(16): 4977-4987. |
| [9] |
王芳. 不同分解者类群对溪流凋落物分解的贡献研究[D]. 重庆: 重庆大学, 2020.
|
| [10] |
Liu J, Liu X Y, Song Q N, Compson Z G, LeRoy C J, Luan F G, Wang H, Hu Y L, Yang Q P. Synergistic effects: a common theme in mixed-species litter decomposition. New Phytologist, 2020, 227(3): 757-765. DOI:10.1111/nph.16556 |
| [11] |
Hättenschwiler S, Tiunov A V, Scheu S. Biodiversity and litter decomposition in terrestrial ecosystems. Annual Review of Ecology, Evolution, and Systematics, 2005, 36: 191-218. DOI:10.1146/annurev.ecolsys.36.112904.151932 |
| [12] |
Smith V C, Bradford M A. Do non-additive effects on decomposition in litter-mix experiments result from differences in resource quality between litters?. Oikos, 2003, 102(2): 235-242. DOI:10.1034/j.1600-0706.2003.12503.x |
| [13] |
张慧慧, 白云玉, 张英洁, 靳英华, 许嘉巍, 陶岩, 水新利, 赵琛, 刘丽杰. 长白山苔原带3种类型的凋落物化学组分与生态化学计量特征对模拟氮沉降的响应. 生态学报, 2022(21): 1-14. |
| [14] |
魏复盛. 国家环境保护总局, 水和废水监测分析方法编委会编 (4版). 北京: 中国环境科学出版社, 2002.
|
| [15] |
Graça M, Bärlocher F, Gessner M. Methods to study litter decomposition: a practical guide. 1st ed. New York: Springer, 2005: 3-5.
|
| [16] |
Wang F, Lin D M, Li W, Dou P P, Han L, Huang M F, Qian S H, Yao J M. Meiofauna promotes litter decomposition in stream ecosystems depending on leaf species. Ecology and Evolution, 2020, 10(17): 9257-9270. DOI:10.1002/ece3.6610 |
| [17] |
Tachet H, Richoux P, Bournaud M. Invertébrés d'eau douce Systematique, biologie, écologie. 1st ed. Paris: CNRS, 2000: 29-30.
|
| [18] |
Giere O. Meiobenthology·The Microscopic Motile Fauna of Aquatic Sediments. 2nd ed. German: Springer-Verlag Berlin Heidelberg, 2009: 1-2.
|
| [19] |
Lin G G, Zeng D H. Functional identity rather than functional diversity or species richness controls litter mixture decomposition in a subtropical forest. Plant and Soil, 2018, 428(1/2): 179-193. |
| [20] |
Seastedt T R, Todd T C, James S W. Experimental manipulations of the arthropod, nematode and earthworm communities in a North American tallgrass prairie. Pedobiologia, 1987, 30: 9-17. DOI:10.1016/S0031-4056(23)00351-7 |
| [21] |
Lavorel S, Grigulis K, McIntyre S, Williams N S G, Garden D, Dorrough J, Berman S, Quétier F, Thébault A, Bonis A. Assessing functional diversity in the field-methodology matters!. Functional Ecology, 2008, 22(1): 134-147. |
| [22] |
Laliberté E, Legendre P. A distance-based framework for measuring functional diversity from multiple traits. Ecology, 2010, 91(1): 299-305. DOI:10.1890/08-2244.1 |
| [23] |
Loreau M, Hector A. Partitioning selection and complementarity in biodiversity experiments. Nature, 2001, 412(6842): 72-76. DOI:10.1038/35083573 |
| [24] |
Bates D, Mächler M, Bolker B, Walker S. Fitting linear mixed-effects models Using lme4. Journal of Statistical Software, 2015, 67(1): 1-48. |
| [25] |
张雨鉴, 王克勤, 宋娅丽, 郑兴蕊, 潘禹, 苏孟白, 李晓龙. 滇中亚高山地带性植被凋落物分解对模拟氮沉降的响应. 生态学报, 2020, 40(22): 8274-8286. |
| [26] |
Santschi F, Gounand I, Harvey E, Altermatt F. Leaf litter diversity and structure of microbial decomposer communities modulate litter decomposition in aquatic systems. Functional Ecology, 2018, 32(2): 522-532. DOI:10.1111/1365-2435.12980 |
| [27] |
Yue K, Peng C H, Yang W Q, Peng Y, Zhang C, Huang C P, Wu F Z. Degradation of lignin and cellulose during foliar litter decomposition in an alpine forest river. Ecosphere, 2016, 7(10): e01523. |
| [28] |
Gessner M O, Chauvet É. Importance of stream microfungi in controlling breakdown rates of leaf litter. Ecology, 1994, 75(6): 1807-1817. DOI:10.2307/1939639 |
| [29] |
王小平, 杨雪, 杨楠, 辛晓静, 曲耀冰, 赵念席, 高玉葆. 凋落物多样性及组成对凋落物分解及土壤微生物群落的影响——二氧化碳倍增条件下. 生态学报, 2020, 40(17): 6171-6178. |
| [30] |
豆鹏鹏, 林敦梅, 王芳, 黄屿玥, 高杰, 杨巍, 韦冰, 李伟, 姚婧梅. 三峡库区城市富营养化河流凋落物的混合分解效应. 中国环境科学, 2021, 41(8): 3775-3783. DOI:10.19674/j.cnki.issn1000-6923.20210526.001 |
| [31] |
Boyero L, Barmuta L A, Ratnarajah L, Schmidt K, Pearson R G. Effects of exotic riparian vegetation on leaf breakdown by shredders: a tropical-temperate comparison. Freshwater Science, 2012, 31(2): 296-303. DOI:10.1899/11-103.1 |
| [32] |
Ab Hamid S, Md Rawi C S. Ephemeroptera, Plecoptera and Trichoptera (Insecta) abundance, diversity and role in leaf litter breakdown in tropical headwater river. Tropical Life Sciences Research, 2017, 28(2): 89-105. DOI:10.21315/tlsr2017.28.2.7 |
| [33] |
Roberts M, Strauch A M, Wiegner T, MacKenzie R A. Leaf litter breakdown of native and exotic tree species in two Hawaiian streams that differ in flow. Pacific Science, 2016, 70(2): 209-222. DOI:10.2984/70.2.7 |
| [34] |
Compson Z G, Hungate B A, Whitham T G, Koch G W, Dijkstra P, Siders A C, Wojtowicz T, Jacobs R, Rakestraw D N, Allred K E, Sayer C K, Marks J C. Linking tree genetics and stream consumers: isotopic tracers elucidate controls on carbon and nitrogen assimilation. Ecology, 2018, 99(8): 1759-1770. DOI:10.1002/ecy.2224 |
| [35] |
Martins R T, Melo A S, Jr Gonçalves J F, Hamada N. Leaf-litter breakdown in urban streams of Central Amazonia: direct and indirect effects of physical, chemical, and biological factors. Freshwater Science, 2015, 34(2): 716-726. DOI:10.1086/681086 |
| [36] |
Nanda A, Asaeda T, Fujino T, Siong K, Nakajima T. Aggregation of Lepidostomatidae in small mesh size litter-bags: implication to the leaf litter decomposition process. Wetlands Ecology and Management, 2009, 17(4): 417-421. DOI:10.1007/s11273-008-9114-6 |
| [37] |
Ferreira V, Chauvet E, Canhoto C. Effects of experimental warming, litter species, and presence of macroinvertebrates on litter decomposition and associated decomposers in a temperate mountain stream. Canadian Journal of Fisheries and Aquatic Sciences, 2014, 72(2): 206-216. |
| [38] |
Song L J, Di Y, Shi B. The significance and development trend in research of plant polyphenols. Progress in Chemistry, 2000, 12(2): 161-170. |
| [39] |
Mudrick D A, Hoosein M, Hicks R R Jr, Townsend E C. Decomposition of leaf litter in an Appalachian forest: effects of leaf species, aspect, slope position and time. Forest Ecology and Management, 1994, 68(2/3): 231-250. |
| [40] |
Wang F, Lin D M, Li W, Dou P P, Han L, Huang M F, Qian S H, Yao J M. Meiofauna promotes litter decomposition in stream ecosystems depending on leaf species. Ecology and Evolution, 2020, 10(17): 9257-9270. DOI:10.1002/ece3.6610 |
| [41] |
Baldrighi E, Giovannelli D, Errico G D, Lavaleye M, Manini E. Exploring the relationship between macrofaunal biodiversity and ecosystem functioning in the deep sea. Frontiers in Marine Science, 2017, 4: 198. DOI:10.3389/fmars.2017.00198 |
| [42] |
Pozo J, Casas J, Menéndez M, Mollá S, Arostegui I, Basaguren A, Casado C, Descals E, García-Avilés J, González J M, Larrañaga A, López E, Lusi M, Moya O, Pérez J, Riera T, Roblas N, Salinas M J. Leaf-litter decomposition in headwater streams: a comparison of the process among four climatic regions. Journal of the North American Benthological Society, 2011, 30(4): 935-950. DOI:10.1899/10-153.1 |
| [43] |
Jr Gonçalves J F, Graça M A S, Callisto M. Litter decomposition in a Cerrado savannah stream is retarded by leaf toughness, low dissolved nutrients and a low density of shredders. Freshwater Biology, 2007, 52(8): 1440-1451. DOI:10.1111/j.1365-2427.2007.01769.x |
| [44] |
王璐, 杨海军, 李玲, 南晓飞, 张振兴, 李昆. 长白山地区溪流冻结初期凋落叶分解与底栖动物定殖的关系. 应用生态学报, 2017, 28(11): 3775-3783. DOI:10.13287/j.1001-9332.201711.035 |
| [45] |
Cook A R, Hoellein T J. Environmental drivers of leaf breakdown in an urban watershed. Freshwater Science, 2016, 35(1): 311-323. DOI:10.1086/685086 |
| [46] |
Callisto M, Jr Gonçalves J F, Graça M A S. Leaf litter as a possible food source for chironomids (Diptera) in Brazilian and Portuguese headwater streams. Revista Brasileira de Zoologia, 2007, 24(2): 442-448. DOI:10.1590/S0101-81752007000200023 |
| [47] |
Sanseverino A M, Nessimian J L. The food of larval Chironomidae (Insecta, Diptera) in submerged litter in a forest stream of the Atlantic Forest. Acta Limnologica Brasiliensia, 2008, 20: 15-20. |
| [48] |
Chen S J, Wang D. Effects of micro-, meio- and macroinvertebrates associated with burial on the decomposition of an aquatic macrophyte (Vallisneria natans) in a eutrophic shallow lake in China. Marine and Freshwater Research, 2018, 70(4): 554-562. |
| [49] |
刘均玲, 袁超, 何永姑, 王永强. 东寨港红树林小型底栖动物丰度与Chla、有机质的相关性. 生态学报, 2019, 39(1): 185-191. |
| [50] |
Mora-Gómez J, Elosegi A, Duarte S, Cássio F, Pascoal C, Romaní A M. Differences in the sensitivity of fungi and bacteria to season and invertebrates affect leaf litter decomposition in a Mediterranean stream. FEMS Microbiology Ecology, 2016, 92(8): fiw121. DOI:10.1093/femsec/fiw121 |
| [51] |
Majdi N, Tackx M, Buffan-Dubau E. Trophic positioning and microphytobenthic carbon uptake of biofilm-dwelling meiofauna in a temperate river. Freshwater Biology, 2012, 57(6): 1180-1190. DOI:10.1111/j.1365-2427.2012.02784.x |
| [52] |
Cummins K W, Wilzbach M, Gates D M. Shredders Riparian Vegetation stream invertebrates. BioScience, 1989, 39: 24-30. DOI:10.2307/1310804 |
| [53] |
Hakenkamp C C, Morin A. The importance of meiofauna to lotic ecosystem functioning. Freshwater Biology, 2000, 44(1): 165-175. DOI:10.1046/j.1365-2427.2000.00589.x |
| [54] |
Santschi F, Gounand I, Harvey E, Altermatt F. Leaf litter diversity and structure of microbial decomposer communities modulate litter decomposition in aquatic systems. Functional Ecology, 2018, 32(2): 522-532. DOI:10.1111/1365-2435.12980 |
| [55] |
Tonin A M, Gonçalves J F, Bambi P, Couceiro S R M, Feitoza L A M, Fontana L E, Hamada N, Hepp L U, Lezan-Kowalczuk V G, Leite G F M, Lemes-Silva A L, Lisboa L K, Loureiro R C, Martins R T, Medeiros A O, Morais P B, Moretto Y, Oliveria P C A, Pereira E B, Ferreira L P, Pérez J, Petrucio M M, Reis D F, Rezende RS, Roque N, Santos L E P, Siegloch A E, Tonello G, Boyero L. Plant litter dynamics in the forest-stream interface: precipitation is a major control across tropical biomes. Scientific Reports, 2017, 7: 10799. DOI:10.1038/s41598-017-10576-8 |
| [56] |
Conn C, Dighton J. Litter quality influences on decomposition, ectomycorrhizal community structure and mycorrhizal root surface acid phosphatase activity. Soil Biology and Biochemistry, 2000, 32(4): 489-496. DOI:10.1016/S0038-0717(99)00178-9 |