 2023, Vol. 43
2023, Vol. 43文章信息
- 宋全健, 丰美萍, 边文华, 程晓鹏, 汪振华, 章守宇, 刘洪生
- SONG Quanjian, FENG Meiping, BIAN Wenhua, CHENG Xiaopeng, WANG Zhenhua, ZHANG Shouyu, LIU Hongsheng
- 大陈岛海域浮游动物群落季节变化及其影响因素
- Seasonal variation and environmental factors of zooplankton communities in sea areas around Dachen Island
- 生态学报. 2023, 43(17): 7128-7139
- Acta Ecologica Sinica. 2023, 43(17): 7128-7139
- http://dx.doi.org/10.20103/j.stxb.202207091960
-
文章历史
- 收稿日期: 2022-07-09
- 网络出版日期: 2023-04-28





2. 上海海洋大学海洋牧场工程技术研究中心, 上海 201306;
3. 上海海洋大学上海海洋大学环境DNA技术与水生态健康评估工程中心, 上海 201306
2. Engineering Technology Research Center of Marine Ranching, Shanghai Ocean University, Shanghai 201306, China;
3. Engineering Research Center of Environmental DNA and Ecological Water Health Assessment, Shanghai Ocean University, Shanghai 201306, China
浮游动物物种丰富、分布广泛、数量庞大, 是海洋生态系统中重要的生态类群, 不仅通过摄食行为制约着浮游植物和微生物的生长繁殖[1—4], 而且浮游动物作为许多鱼、虾类的饵料生物, 通过“上行效应”影响着鱼、虾等渔业资源的群落结构和总量[5—6]。同时, 浮游动物对环境变化的反应较为敏感[7—9], 因此, 水环境变化、浮游植物和渔业资源结构和数量的变动都影响浮游动物群落结构的改变[10—14]。研究浮游动物的种类组成、群落结构及季节变化特征对海洋环境和渔业资源的保护具有重要意义[15—18]。
大陈岛位于浙江省中部椒江口的东南部海域, 主要由上、下大陈岛两个岛屿组成。大陈岛海域的水文环境复杂, 不仅受到椒江水系淡水汇入的影响, 还受黑潮暖流的影响。黑潮暖流的次表层水爬升在此海域形成了明显的上升流[19—22], 上升流将底层的部分营养盐输送到表层, 椒江水系的汇入也为此海域带来了大量的陆地营养盐。因此, 大陈岛海域的营养盐丰富, 为浮游生物的大量生长和繁殖提供了良好的条件[23], 从而为许多经济鱼类提供了大量的饵料生物, 是许多经济鱼类的索饵场和产卵地[24]。大陈岛海域渔业资源丰富, 是鱼山渔场的重要部分, 鱼山渔场是仅次于舟山渔场的浙江省第二大渔场[25]。近年来关于大陈岛附近海域浮游动物群落结构的报道不多, 且多是对椒江口海域及浙江中部海域的研究[26—28]。虽然部分研究涉及到了对大陈岛海域的调查, 但是采样站点较少, 不能详细说明大陈岛海域的现状。本次研究对大陈岛海域做了多站点、多航次、多季节的调查, 对该海域浮游动物的物种组成、优势种、丰度、生物量、多样性及与环境因子的关系进行了深入分析, 为当地海洋牧场的规划和功能设计提供基础数据, 为更好的确定渔业资源目标种提供参考。
1 材料与方法 1.1 样品采集与分析于2020年9月(夏季)、11月(秋季)和2021年1月(冬季)、4月(春季)分别对浙江省大陈岛海域的浮游生物、环境因子等进行了4个航次的综合调查, 共设置了21个采样站点(图 1), 其中2020年9月和11月共20个站位(缺少站位DC21)。浮游动物用浅水Ⅰ型浮游生物网(网口面积0.2 m2, 筛绢孔径为505 μm)由底层(距海底2 m)至表层垂直拖网获得, 使用网口流量计测量通过浮游生物网的海水体积(m3), 样品均用5%的福尔马林溶液现场固定。带回实验室浓缩后, 用体视显微镜鉴定种类和计数, 湿重法称量重量。调查海域水温、盐度和叶绿素a浓度(Chl a)等环境因子均使用SeaBird CTD进行现场测定。

|
| 图 1 大陈岛海域浮游动物采样站位 Fig. 1 The location of sampling sites of zooplankton in sea areas around Dachen Island DC1, DC2, …, DC20为采样站位 |
采用优势度指数(Y)[29]来确定大陈岛海域浮游动物的优势种。
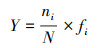
|
式中, N为样品中所有种类的总丰度;ni为第i种的丰度;fi为样品中第i种的出现频率, Y>0.02时为优势种。
物种多样性指标采用Shannon-Wiener多样性指数(H′)[30]和Pielou均匀度指数(J′)[31]。

|
式中, Pi为第i个物种的丰度占总丰度的百分比;S为物种数。
使用ARCGIS 10.8绘制采样站位图, 丰度、生物量分布图;为了减少偶见种对结果的干扰, 使用PRIMER 6对各航次出现频率在20%以上的物种进行群落聚类分析和多维度尺度分析(MDS)[32];使用SPSS 24.0对浮游动物的种类数、丰度、生物量等与海水温度、盐度等环境因子进行Pearson相关性分析;使用CONOCO 5对各航次的优势种与环境因子进行冗余分析(RDA)。
2 结果与分析 2.1 环境因子调查海域表层水温有明显的季节变化, 夏季水温最高, 冬季最低, 秋季、春季次之。夏、春季温度高值区出现在调查海域的中部;秋、冬季温度高值区出现在调查海域的东部。由于受到西侧椒江大量淡水汇入的影响, 盐度全年都呈现出从调查海域的西侧到东侧逐渐升高的趋势, 夏季椒江处在丰水期, 水流量大, 夏季盐度明显低于其他季节。Chl a呈现出春夏季较高, 秋冬季较低的特点, 夏季高值区出现在调查海域的东南部;秋、冬季高值区主要出现在西北部;春季分布较为均匀, 高值区出现在西南部(图 2)。

|
| 图 2 大陈岛海域海水温度、盐度和Chl a的平面分布 Fig. 2 Horizontal distribution of temperature, salinity and Chl a in sea areas around Dachen Island |
共鉴定浮游动物90种, 隶属于15大类群, 其中桡足类种类数最多, 有34种, 占总种类数的37.8%;水螅水母类8种、管水母类5种, 分别占总种类数的8.9%和5.6%;端足类和软体动物各4种, 分别占4.4%;介型类、毛颚类、十足类和尾索动物各3种, 多毛类、糠虾类和栉水母各2种, 涟虫类、磷虾类各1种, 共占总种类数的22.2%;浮游幼体共发现15类, 占16.7%(图 3)。

|
| 图 3 大陈岛海域浮游动物种类数、丰度和生物量的季节变化 Fig. 3 Seasonal variation of zooplankton species, abundance, and biomass in sea areas around Dachen Island |
浮游动物群落的种类组成有明显的季节变化, 夏季浮游动物种类最丰富, 共68种;其次是秋季, 为58种;冬、春季较少, 春季(26种)略高于冬季(20种)(图 3)。夏季种类数最高值出现在站点DC15(28种), 最低值出现在站点DC8(11种);秋季种类数的高值区出现在调查海域的东部和西北部, 最高值出现在站点DC19、DC23(23种), 最低值出现在站点DC7、DC14(2种);冬、春季种类数分布较为均匀(图 4)。

|
| 图 4 大陈岛海域浮游动物种类数分布 Fig. 4 Horizontal distribution of zooplankton species richness in sea areas around Dachen Island |
调查海域4个季节共计算出12种优势种(Y>0.02), 其中桡足类最多(5种)。浮游动物优势种也有着明显的季节变化, 秋季优势种最多(9种), 其中太平洋纺锤水蚤(Acartia pacifica)优势度最高;夏季次之(6种), 双生水母(Diphyes chamissonis)的优势度最高;春季和冬季各出现2种优势种, 其中中华哲水蚤(Calanus sinicus)是春季和冬季的绝对优势种, 其丰度分别占春季和冬季总丰度的91%和83%(图 3, 表 1)。
| 序号 Code |
种类 Species |
夏季 Summer |
秋季 Autumn |
冬季 Winter |
春季 Spring |
|||||||
| 优势度 | 平均丰度/(个/m3) | 优势度 | 平均丰度/(个/m3) | 优势度 | 平均丰度/(个/m3) | 优势度 | 平均丰度/(个/m3) | |||||
| SP1 | 百陶箭虫Sagitta bedoti | 0.03 | 3.86±3.38 | 0.02 | 1.94±4.52 | — | — | 0.02 | 9.08±4.14 | |||
| SP2 | 大西洋五角水母Muggiaea atlantica | — | — | 0.02 | 3.18±11.01 | / | / | — | — | |||
| SP3 | 磷虾带叉幼虫Furcilia larva | — | — | 0.03 | 3.04±4.53 | / | / | — | — | |||
| SP4 | 肥胖箭虫Sagitta enflata | 0.12 | 16.06±13.9 | — | — | / | / | / | / | |||
| SP5 | 磷虾节胸幼虫 Calyptopis larva | — | — | 0.06 | 5.37±7.76 | / | / | — | — | |||
| SP6 | 精致真刺水蚤Euchaeta concinna | 0.18 | 25.01±45.07 | 0.04 | 6.09±14.23 | — | — | — | — | |||
| SP7 | 双生水母Diphyes chamissonis | 0.33 | 44.25±46.05 | 0.04 | 3.23±2.43 | — | — | / | / | |||
| SP8 | 太平洋纺锤水蚤Acartia pacifica | — | — | 0.33 | 30.24±97.75 | / | / | / | / | |||
| SP9 | 微刺哲水蚤Canthocalanus pauper | 0.02 | 3.39±3.01 | 0.02 | 2.25±3.75 | / | / | / | / | |||
| SP10 | 针刺拟哲水蚤Paracalanus aculeatus | — | — | — | — | 0.07 | 2.07±2.4 | / | / | |||
| SP11 | 中华假磷虾Pseudeuphausia sinica | 0.16 | 22.92±41.6 | — | — | — | — | — | — | |||
| SP12 | 中华哲水蚤Calanus sinicus | — | — | 0.04 | 4.07±10.43 | 0.83 | 21.09±17.14 | 0.91 | 344.25±297.1 | |||
| SP: 种类Species;“—”表示此物种不是优势种(Y < 0.02);“/”表示未出现该物种 | ||||||||||||
调查海域浮游动物全年丰度范围为1.3—1190.9个/m3, 均值为153.4±214.7个/m3。有明显的季节变化, 春季丰度最高(380.2±296.2)个/m3, 冬季最低(25.3±19.1)个/m3, 夏季(135.3±112.6)个/m3高于秋季(67.9±90.5)个/m3。春季丰度最高值出现在站点DC20, 最低值出现在DC7站点;夏季高值区主要出现在调查海域东北部, 最高值出现在站点DC4, 最低值出现在站点DC9;秋季最高值出现在站点DC18, 最低值出现在站点DC14;冬季丰度分布较为均匀, 最高值出现在站点DC4, 最低值出现在站点DC15(图 3, 图 5)。

|
| 图 5 大陈岛海域浮游动物丰度分布 Fig. 5 Horizontal distribution of zooplankton abundance in sea areas around Dachen Island |
大陈岛海域浮游动物生物量的变化范围为2.2—3570.8 mg/m3, 年平均值为(411.9±561.7) mg/m3。同样有明显的季节变化, 夏季最高(895.0±802.5)mg/m3, 春季次之(623.4±358.7)mg/m3, 秋季(91.1±82.4)mg/m3高于冬季(46.0±85.0)mg/m3。夏季生物量高值区主要出现在调查海域东南部, 最高值出现在站点DC15, 最低值出现在DC19站位;春季最高值出现在站点DC2, 最低值出现在站点DC13;秋季最高值出现在站点DC2, 最低值出现在站点DC14;冬季生物量分布较为均匀, 最高值出现在站点DC4, 最低值出现在站点DC15(图 3, 图 6)。

|
| 图 6 大陈岛海域浮游动物生物量分布 Fig. 6 Horizontal distribution of zooplankton biomass in sea areas around Dachen Island |
大陈岛海域浮游动物群落多样性Shannon-Wiener指数(H′)全年的变化范围为0.36—3.64, 均值为1.71±0.96。多样性指数有明显的季节变化, 夏季最高, 冬季最低, 秋季高于春季。浮游动物Pielou均匀度指数(J′)的变化范围为0.15—0.93, 平均值为0.53±0.21, 秋季最高, 春季最低, 夏季高于冬季(表 2)。
| 季节 Seasons |
多样性指数(H′) Shannon-Weaver divklersity index (H′) |
均匀度指数(J′) Pielou′s evenness index (J′) |
|||
| 范围 | 均值 | 范围 | 均值 | ||
| 全年Annual | 0.36—3.64 | 1.71±0.96 | 0.15—0.93 | 0.53±0.21 | |
| 夏季Summer | 1.57—3.10 | 2.54±0.44 | 0.41—0.78 | 0.61±0.11 | |
| 秋季Autumn | 0.55—3.64 | 2.36±0.98 | 0.16—0.93 | 0.71±0.19 | |
| 冬季Winter | 0.36—1.41 | 0.84±0.35 | 0.23—0.82 | 0.46±0.16 | |
| 春季Spring | 0.36—2.41 | 1.24±0.64 | 0.15—0.80 | 0.37±0.20 | |
聚类分析结果表明(图 7), 除去秋季航次的DC7和DC14两个站点单独聚成一类外, 以33%的相似度, 可以划分为4个类群, 分别为夏季类群、秋季类群、冬季类群和春季类群, 其中, 夏季航次的DC10站点被划入了秋季类群。单因素方差分析(One-way ANOSIM)的检验结果表明, 4个类群中的任意两个类群之间都存在显著的差异性(全局检验: R=0.923, P=0.001), 4个类群内部都表现出了较好的聚类效果, 具有明显的季节变化特征。夏季类群与冬季类群、春季类群之间的差异性最大(R=1, P=0.001), 夏季类群和秋季类群之间的差异性最小(R=0.783, P=0.001)。

|
| 图 7 大陈岛海域调查站位浮游动物群落的聚类分析和多维度尺度分析(MDS)标序 Fig. 7 Cluster and MDS ordination of zooplankton communities at different survey stations in sea areas around Dachen Island |
浮游动物的种类数、丰度、生物量、多样性指数、均匀度指数与水温、盐度、Chl a、浮游植物丰度进行Pearson相关性分析, 结果表明(表 3), 浮游动物丰度与Chl a浓度呈显著正相关(P < 0.05), 与其他环境因子无显著相关性;浮游动物生物量与水温和Chl a呈极显著正相关(P < 0.01), 与盐度呈极显著负相关(P < 0.01);浮游动物种类数与水温和Chl a浓度呈极显著的正相关(P < 0.01), 与浮游植物丰度呈显著正相关(P < 0.05), 与盐度呈极显著负相关(P < 0.01);浮游动物的多样性指数(H′)与水温呈极显著的正相关(P < 0.01), 与盐度呈极显著负相关(P < 0.01);浮游动物均匀度指数(J′)与水温呈极显著正相关(P < 0.01)。
| 相关系数 Correlation coefficient |
温度/(℃) Temperature |
盐度 Salinity |
叶绿素a/(μg/L) Chl a |
浮游植物丰度/(个/L) Phytoplankton abundance |
| 丰度Abundance/(个/m3) | 0.032 | -0.021 | 0.251* | -0.051 |
| 生物量Biomass/(mg/m3) | 0.466** | -0.367** | 0.411** | 0.124 |
| 种类数Species | 0.800** | -0.533** | 0.293** | 0.243* |
| 多样性指数(H′) Diversity index | 0.685** | -0.401** | 0.166 | 0.16 |
| 均匀度指数(J′) Evenness index | 0.351** | -0.134 | -0.075 | 0.062 |
| *表示显著相关(P < 0.05), **表示极显著相关(P < 0.01) | ||||
为了进一步探究大陈岛海域的环境因子对浮游动物群落的影响, 本文对各航次的优势种(共12种)与环境因子进行RDA冗余分析, 结果显示第一主轴和第二主轴的特征值分别为10.94%和5.55%, 前两轴累计解释了浮游动物群落的81.52%, Monte Carlo检验显示, 水温(F=9.2, P=0.002)、盐度(F=4.3, P=0.002)和Chl a(F=3.2, P=0.002)对浮游动物群落有极显著的影响, 解释贡献率分别为50.9%, 23.0%和16.8%。如图所示(图 8), 肥胖箭虫、双生水母、精致针刺水蚤和中华假磷虾等与海水温度呈现出明显的正相关关系, 因此它们在夏季丰度较高;中华哲水蚤与水温有明显的负相关关系, 因此其在春季和冬季丰度较高。百陶箭虫与Chl a有较高的正相关性;反之, 磷虾带叉幼虫、大西洋五角水母、太平洋纺锤水蚤等与Chl a有明显的负相关性。针刺拟哲水蚤偏好在盐度较高的海水中生活。此外, RDA分析图可以体现出浮游动物不同种类的生态习性, 不同物种在分析图中的位置越接近, 说明它们的生态习性越相似。

|
| 图 8 大陈岛海域浮游动物与环境因子的冗余分析 Fig. 8 Redundancy analysis (RDA) of zooplankton constrained by environmental factors WT:水温Water temperature;S:盐度Salinity;Chl a:叶绿素a;PA: 浮游植物丰度Phytoplankton abundance;SP1:百陶箭虫Sagitta bedoti;SP2:大西洋五角水母Muggiaea atlantica;SP3:磷虾带叉幼虫Furcilia larva;SP4:肥胖箭虫Sagitta enflata;SP5:磷虾节胸幼虫Calyptopis larva;SP6:精致真刺水蚤Euchaeta concinna;SP7:双生水母Diphyes chamissonis;SP8:太平洋纺锤水蚤Acartia pacifica;SP9:微刺哲水蚤Canthocalanus pauper;SP10:针刺拟哲水蚤Paracalanus aculeatus;SP11:中华假磷虾Pseudeuphausia sinica;SP12:中华哲水蚤Calanus sinicus |
调查发现大陈岛海域浮游动物群落有明显的季节变化特征, 丰度春季最高, 种类数和生物量夏季最高, 种类数、丰度和生物量都在冬季达到了一年中的最低值。大陈岛海域浮游动物的Shannon-Wiener指数(H′)和Pielou均匀度指数(J′)都表现出夏、秋季较高, 冬、春较低的特点, 春季群落和冬季群落都是单一物种占绝对优势, 生物多样性低, 夏季群落和秋季群落物种丰富, 生物多样性较高, 与孙鲁峰等[26]在大陈岛附近海域(椒江口海域)的研究相似。本次调查共发现浮游动物90种(包括浮游幼体15类), 桡足类最多(34种), 其全年平均丰度占浮游动物总丰度的74.88%, 是调查海域优势度最高的类群, 这一结果与我国大多数海域类似[6, 11, 33—35]。此外, 管水母类、毛颚类和浮游幼体的丰度高、种类多, 也是组成该海域浮游动物群落的重要类群。
有研究表明, 在全球部分海域, 出现了水母类在浮游动物群落中占比逐渐上升的现象, 海洋生态系统有胶质化的趋势, 这一现象与全球气候变暖显著相关[17]。杜明敏[11]等的研究表明, 中国近海水母类的丰度和多样性明显增加, 在东海和南海海域, 桡足类仍然是浮游动物群落中最具优势的类群, 但是随着水母类浮游动物多样性的升高, 桡足类的优势度有所下降;在渤海和黄海的夏季, 虽然桡足类的丰度在浮游动物总丰度中最占优势的地位仍未改变, 但是水母类的种类数已经超过了桡足类。本次调查中, 共发现水母类浮游动物15种, 夏季水母类浮游动物丰度占总丰度的比例为33.84%, 超过了桡足类(占总丰度的比例为24.55%)成为夏季的第一优势类群(图 3), 水母类浮游动物会捕食鱼卵、饵料浮游动物等, 与经济物种竞争, 进而影响渔业资源的结构和总量[17], 水母类作为海洋生态系统食物链中的重要一环, 其数量和种类的爆发常会引起区域性的海洋灾害[36], 因此, 本次调查发现的大陈岛海域水母类浮游动物在夏季爆发的现象值得关注。
3.2 环境因子对浮游动物群落季节变化的影响浮游动物的群落结构会受到不同环境因子的共同影响, 大陈岛海域浮游动物和环境因子的相关性分析和RDA分析表明, 海水温度、盐度和Chl a是影响浮游动物群落的物种组成、分布及多样性水平的重要环境因子, 在浮游动物的生长繁殖过程中起到重要作用[16, 32, 37]。水温会影响浮游动物的代谢速度, 水温过高或过低均会对浮游动物的生长发育和繁殖产生不利的影响[38—39], 大陈岛海域冬季和春季水温较低, 对多数浮游动物的生长繁殖有抑制作用, 所以冬、春季浮游动物的种类数和多样性指数较低。而有研究表明, 中华哲水蚤对温度的耐受范围为5—23℃, 最适温度在10—18℃[16, 40], 水温过高, 中华哲水蚤的死亡率显著升高[40]。因此中华哲水蚤是大陈岛海域冬季和春季的绝对优势种, 尤其是在春季大量出现, 致使春季浮游动物的平均丰度明显高于其他季节。随着温度的升高, 夏季中华哲水蚤的丰度大幅降低, 而浮动动物种类数迅速增多, 物种多样性和生物量达到了一年中的最高值。夏季主要以大型浮游动物(水母类、毛颚动物门、糠虾目等)为主(图 3), 而中华哲水蚤体型较小, 因此, 夏季浮游动物的生物量高于春季。盐度与浮游动物的生长发育和繁殖过程有着密切关系, 不同物种具有不同的最适盐度范围与生态幅[37], 浮游动物可以通过自身的调节机制来适应一定范围的盐度变化[41—42], 许多研究表明, 盐度也是影响浮游动物的群落结构和分布的关键因素[43—44], 大陈岛海域浮游动物与环境因子的相关性分析表明, 浮游动物的生物量、种类数和多样性指数均与盐度呈极显著的负相关(P < 0.01), 这表明盐度越高可能越不利于大陈岛海域浮游动物群落的发展。
充足的饵料来源是保持浮游动物群落稳定发展的关键因素, 冬季水温偏低, 浮游植物和浮游动物丰度都处在一年中的最低水平, 随着水温的逐渐上升, 浮游植物开始大量增殖, 浮游植物为浮游动物的生长繁殖提供丰富的饵料, 各种浮游幼体大量出现, 多种浮游幼体成为了优势种, 表明了大陈岛海域的大多数动物偏好在水温较高、饵料丰富的季节繁殖。综上所述, 浮游动物的群落结构是多种因素共同影响的结果, 多种生物与分生物因素共同驱动着大陈岛海域浮游动物群落的发展变化。
| [1] |
徐兆礼. 中国海洋浮游动物研究的新进展. 厦门大学学报: 自然科学版, 2006, 45(S2): 16-23. |
| [2] |
Richardson A J. In hot water: zooplankton and climate change. ICES Journal of Marine Science, 2008, 65(3): 279-295. DOI:10.1093/icesjms/fsn028 |
| [3] |
Yuliana, Mutmainnah. Assessment of coastal water productivity of Fitu Village, Ternate Island based on its zooplankton community structure. IOP Conference Series: Earth and Environmental Science, 2021, 890(1): 012011. DOI:10.1088/1755-1315/890/1/012011 |
| [4] |
张亮, 宿凯, 张绍萍, 孙滨, 蒲思潮, 王俊健, 纪莹璐. 日照岚山港北作业区邻近海域春、秋季浮游动物群落结构研究. 海洋学报, 2022, 44(1): 79-88. |
| [5] |
Baars M, Fransz H. Grazing pressure of copepods on the phytoplankton stock of the central North Sea. Netherlands Journal of Sea Research, 1984, 18(1/2): 120-142. |
| [6] |
庞碧剑, 蓝文陆, 黎明民, 李天深. 北部湾近岸海域浮游动物群落结构特征及季节变化. 生态学报, 2019, 39(19): 7014-7024. |
| [7] |
Fullgrabe L, Grosjean P, Gobert S, Lejeune P, Leduc M, Engels G, Dauby P, Boissery P, Richir J. Zooplankton dynamics in a changing environment: a 13-year survey in the northwestern Mediterranean Sea. Marine Environmental Research, 2020, 159: 104962. DOI:10.1016/j.marenvres.2020.104962 |
| [8] |
Bellier E, Engen S, Jensen T C. Seasonal diversity dynamics of a boreal zooplankton community under climate impact. Oecologia, 2022, 199(1): 139-152. DOI:10.1007/s00442-022-05165-0 |
| [9] |
Kmiha-Megdiche S, Drira Z, Ayadi H. Response of zooplankton population to environmental forcing-case of southern and northern coasts of sfax, tunisia. International Journal of Environmental Sciences & Natural Resources, 2020, 23(2): 556110. |
| [10] |
张才学, 龚玉艳, 孙省利, 施玉珍, 杨国欢, 柯盛. 2010年夏季雷州半岛沿岸海域浮游动物群落结构特征. 海洋学报, 2014, 36(4): 91-99. DOI:10.3969/j.issn.0253-4193.2014.04.008 |
| [11] |
杜明敏, 刘镇盛, 王春生, 张东声, 章菁. 中国近海浮游动物群落结构及季节变化. 生态学报, 2013, 33(17): 5407-5418. |
| [12] |
徐兆礼, 晁敏, 陈亚瞿. 东海浮游动物生物量分布特征. 海洋学报, 2004, 26(3): 93-101. |
| [13] |
毕洪生, 孙松, 高尚武, 张光涛, 张芳. 渤海浮游动物群落生态特点Ⅲ.部分浮游动物数量分布和季节变动. 生态学报, 2001, 21(4): 513-521. |
| [14] |
陈洪举. 黄、东海浮游动物群落结构和多样性研究[D]. 青岛: 中国海洋大学, 2010.
|
| [15] |
叶文建, 杜萍, 寿鹿, 蔡小霞, 陈悦, 汤雁滨, 廖一波, 黄伟, 刘小涯. 舟山海域大中型浮游动物群落时空变化及受控要素. 生态学报, 2021, 41(1): 254-267. |
| [16] |
周孔霖. 黄海中华哲水蚤度夏机制的新探索: 温度和饵料的作用[D]. 青岛: 中国科学院大学(中国科学院海洋研究所), 2016.
|
| [17] |
邵倩文, 刘镇盛, 章菁, 孙栋, 林施泉. 长江口及邻近海域浮游动物群落结构及季节变化. 生态学报, 2017, 37(2): 683-691. |
| [18] |
Li K Z, Ma J, Huang L M, Tan Y H, Song X Y. Environmental drivers of temporal and spatial fluctuations of mesozooplankton community in Daya bay, northern South China Sea. Journal of Ocean University of China, 2021, 20(4): 1013-1026. DOI:10.1007/s11802-021-4602-x |
| [19] |
曹欣中. 浙江近海上升流季过程的初步研究. 水产学报, 1986, 10(1): 51-69. |
| [20] |
何德华, 杨关铭, 方绍锦, 沈伟林, 刘红斌, 高爱根, 黄淑莘. 浙江沿岸上升流区浮游动物生态研究——Ⅰ.生物量及主要类群丰度. 海洋学报, 1987, 9(1): 79-92. |
| [21] |
胡敦欣, 吕良洪, 熊庆成, 丁宗信, 孙寿昌. 关于浙江沿岸上升流的研究. 科学通报, 1980, 25(3): 131-133. |
| [22] |
许建平, 曹欣中, 潘玉球. 浙江近海存在沿岸上升流的证据. 海洋湖沼通报, 1983(4): 17-25. |
| [23] |
孙鲁峰. 浙江中部近海上升流对浮游生物分布的影响[D]. 上海: 上海海洋大学, 2012.
|
| [24] |
周永东, 金海卫, 蒋日进, 张洪亮, 张亚洲, 潘国良. 浙江中北部沿岸春、夏季鱼卵和仔稚鱼种类组成与数量分布. 水产学报, 2011, 35(6): 880-889. |
| [25] |
陈剑, 徐兆礼. 闽江口和椒江口浮游动物饵料特征和渔场属性的比较. 中国水产科学, 2015, 22(4): 770-779. |
| [26] |
孙鲁峰, 孙岳, 徐兆礼. 椒江口海域浮游动物群落季节变化特征. 上海海洋大学学报, 2014, 23(1): 131-138. |
| [27] |
樊紫薇. 浙江中部近岸海域鱼卵、仔稚鱼群落结构特征研究[D]. 舟山: 浙江海洋大学, 2020.
|
| [28] |
王婕妤, 唐静亮, 胡颢琰, 黄备, 魏娜. 浙江近岸海域浮游动物的生态分布特征. 浙江海洋学院学报: 自然科学版, 2008, 27(4): 384-389. |
| [29] |
徐兆礼, 陈亚瞿. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系. 生态学杂志, 1989, 8(4): 13-15. |
| [30] |
Burks A W, Shannon C E, Weaver W. The mathematical theory of communication. The Philosophical Review, 1951, 60(3): 398. |
| [31] |
Pielou E C. An Introduction to Mathematical Ecology. New York: Wiley-Interscience, 1969.
|
| [32] |
黄彬彬, 吴风霞, 郑淑娴, 田丰歌, 何静. 雷州半岛以东近岸海域浮游动物群落结构与环境因子关系. 广东海洋大学学报, 2022, 42(2): 53-61. |
| [33] |
骆鑫, 曾江宁, 徐晓群, 杜萍, 廖一波, 刘晶晶. 舟山海域夏、秋季浮游动物的分布特征及其与环境因子的关系. 生态学报, 2016, 36(24): 8194-8204. |
| [34] |
龚玉艳, 肖雅元, 徐姗楠, 刘永, 杨玉滔, 李纯厚. 陵水湾浮游动物群落结构特征及其季节变化. 中国水产科学, 2019, 26(1): 71-81. |
| [35] |
张光星, 陈石泉, 陈丹丹, 吴钟解, 李元超, 蔡泽富, 王道儒. 海南岛近岸海域夏初浮游动物与环境因子的关系. 海洋学报, 2014, 36(12): 125-132. |
| [36] |
Dumont H J. The distribution and ecology of the fresh- and brackish-water medusae of the world. Hydrobiologia, 1994, 272(1): 1-12. |
| [37] |
薛泽, 朱丽岩, 王博渊, 刘正宇. 温度和盐度对两种海洋桡足类动物摄食和代谢的影响. 海洋与湖沼, 2020, 51(1): 95-102. |
| [38] |
Lenz P H, Hower A E, Hartline D K. Temperature compensation in the escape response of a marine copepod, Calanus finmarchicus(Crustacea). The Biological Bulletin, 2005, 209(1): 75-85. |
| [39] |
Devreker D, Souissi S, Seuront L. Effects of chlorophyll concentration and temperature variation on the reproduction and survival of Temora longicornis (Copepoda, Calanoida) in the Eastern English Channel. Journal of Experimental Marine Biology and Ecology, 2005, 318(2): 145-162. |
| [40] |
Pu X M, Sun S, Yang B, Zhang G T, Zhang F. Life history strategies of Calanus sinicus in the southern Yellow Sea in summer. Journal of Plankton Research, 2004, 26(9): 1059-1068. |
| [41] |
Mouny P, Dauvin J C. Environmental control of mesozooplankton community structure in the Seine Estuary (English Channel). Oceanologica Acta, 2002, 25(1): 13-22. |
| [42] |
刘耀谦, 孙省利, 张才学. 2014—2015年硇洲岛大型海藻场浮游动物的群落结构和季节变化. 海洋学报, 2018, 40(12): 94-111. |
| [43] |
Telesh I V, Khlebovich V V. Principal processes within the estuarine salinity gradient: a review. Marine Pollution Bulletin, 2010, 61(4/5/6): 149-155. |
| [44] |
Zakaria H Y, Radwan A A, Said M A. Influence of salinity variations on zooplankton community in el-mex bay, alexandria, Egypt. National institute of oceanography and fisheries, Egypt, 2007, 33(3): 52-67. |