 2023, Vol. 43
2023, Vol. 43文章信息
- 倪秀雅, 冯永霞, 李如华, 尚鹤, 陈展
- NI Xiuya, FENG Yongxia, LI Ruhua, SHANG He, CHEN Zhan
- 重庆酸雨区不同林型对土壤酸化和真菌群落的影响
- Effects of different plantation types on soil acidification and soil fungal community in acid rain area of Chongqing
- 生态学报. 2023, 43(17): 7203-7215
- Acta Ecologica Sinica. 2023, 43(17): 7203-7215
- http://dx.doi.org/10.20103/j.stxb.202207071942
-
文章历史
- 收稿日期: 2022-07-07
- 网络出版日期: 2023-04-28

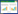




酸沉降是全球面临的主要环境问题之一, 由于酸沉降对生态系统的负面影响, 已引起了学界和政府的广泛关注[1—2]。我国由于人口众多, 工业快速发展, 酸雨已成为一个比较严重的环境问题。我国政府采取了一系列措施控制SO2和NOX等酸性气体的排放[3—4], SO2排放量在2000年代显著下降, NOx的排放量在2012—2015年间也降低了约20%[5]。同时, 近年来我中国政府尤其关注控制农业源NH3的排放[5], 但不容忽视的是大气中的NH3在中和酸性降水中起着重要的作用, 能有助于减轻酸雨[6]。而大气中NH3水平降低可能会加剧酸性降水使得陆地生态系统面临酸雨污染, 尤其是我国南方和西南地区的土壤缓冲能力较低对酸雨尤其敏感[7]。Liu等[5]研究发现, 如果在SO2和NOX减排15%的同时, NH3减排50%的话, 则会显著加剧降水的酸化, 雨水pH值在北方降低1个单位之多, 而南方也将降低0.1—0.5个单位, 且重酸雨面积会相应地大幅增加。鉴于我国南部和四川盆地的降水酸度要大得多, 即使pH值小幅下降也会使本已严峻的局势恶化[5], 因此, 酸沉降在我国南方仍然是一个严重的环境问题[2, 8—9]。
酸沉降对森林生态系统造成严重的影响, 尤其是土壤酸化对森林的影响是长期的和持久的。酸化导致土壤中Ca等阳离子的流失则可以从根本上改变土壤过程, 损害敏感树种的营养状况, 影响其生长[10]。在美国西北部的阿迪朗达克地区, 由于酸沉降引起土壤酸化而造成土壤中Ca缺乏、盐基离子含量低[11—12], 从而影响了糖枫树(Acer saccharum Marsh)的更新, 使其冠层状况变差, 并可能降低基茎的生长[13]。土壤酸化会造成土壤中铝的活化, 对植物产生铝毒, 由于酸沉降造成土壤中铝浓度升高, 限制了植物对水和营养物质的吸收, 从而降低云杉(Picea rubens Sarg.)对其他环境胁迫的抵抗力, 致使其发生衰退[10]。酸沉降会导致土壤中氮含量的积累, 影响微生物过程, 如矿化、硝化和反硝化等, 从而影响森林中NO、N2O等温室气体的排放[14]。有研究表明, 尽管酸沉降输入降低, 由于土壤中吸附的SO42-的历史存量使得酸化土壤的恢复减慢[1, 15—16]。笔者前期研究发现, 在重庆铁山坪地区即使降雨pH值偏中性(6.58—7.33)[17], 但森林土壤pH值仍然偏酸性, 在3.80—4.84之间[18], 说明即使从20世纪八十年代开始采取措施降低酸性气体排放, 但酸化土壤的恢复存在滞后效应。
重庆铁山坪是我国酸雨研究的热点地区之一, 自2000年以来, 铁山坪就已成为中挪合作的中国酸沉降综合影响观测研究项目的五个观测点之一[19]。铁山坪的森林主要为起源于20世纪60年代天然林皆伐后自然更新形成的马尾松次生林, 因受酸沉降危害造成森林衰退, 因此, 20世纪80和90年代以来, 陆续在马尾松林下及其采伐迹地、火烧迹地补植了阔叶树种, 或结合森林防火栽植或补植了防火树种木荷, 现已形成了马尾松纯林(Pinus massoniana Lamb)、香樟林[Cinnamomum camphora (Linn) Presl]、木荷林(Schima superba Gardn. et Champ)以及针阔混交林等多种森林类型镶嵌分布的空间格局。土壤pH值是土壤微生物的一个重要驱动因子, 土壤酸化必然引起土壤微生物的响应。土壤微生物在土壤碳氮磷等元素循环中起着至关重要的作用, 与土壤健康密切相关。不同森林类型的土壤细菌群落差异可能影响土壤酸化缓冲能力。研究发现, 在重庆铁山坪受损马尾松林改造成香樟纯林或马尾松-香樟混交林后, 其土壤中一些有利于土壤团聚体形成的细菌以及增强环境胁迫抗性细菌的相对丰度较高, 而产酸和嗜酸菌丰度较低;相比之下, 马尾松纯林和种植木荷的土壤pH值较低, 且产酸菌和嗜酸菌丰度较高[20]。土壤真菌是另一类重要的土壤微生物, 相比于细菌, 真菌更直接地依赖于凋落物和树木的营养相互作用, 许多真菌是专一性根共生体或病原体[21—22];且真菌更倾向于酸性环境[23], 土壤酸化必将对土壤真菌产生影响。因此, 了解土壤酸化背景下不同林型的土壤真菌群落结构有助于我们更好地了解不同营林措施对酸化土壤及其森林质量的影响, 为酸雨区森林生态系统健康可持续发展提供科学依据。
本研究以重庆铁山坪林场的马尾松纯林(Pi)、香樟纯林(Ci)、木荷纯林(Sc)、马尾松-香樟混交林(Pi_Ci)以及马尾松-木荷(Pi_Sc)混交林为研究对象, 通过对土壤理化性质、土壤真菌群落结构的分析, 探讨在酸雨区将马尾松纯林改造成针阔混交林或阔叶林后对土壤酸化和土壤真菌群落结构的影响, 以及对森林生态系统功能可能存在的潜在影响。
1 材料与方法 1.1 研究区概况研究地点为重庆市铁山坪林场(N29°38′, E106°41′), 位于重庆市中心东北方向约25 km处, 属四川盆地低山丘陵区, 海拔变化在242—579 m, 坡度变化在5—30°。该地为亚热带湿润气候, 年降水量1100 mm, 年均气温18 ℃。土壤以砂岩上发育的山地黄壤为主, 厚度50—80 cm, 土壤密度1.25—1.64 g/cm3, 土壤酸化比较严重(pH值3.80—4.84)[18]。铁山坪林场现有森林1200 hm2, 以20世纪60年代破坏后天然更新形成的马尾松次生林为主, 马尾松纯林林龄平均为62年。香樟林为20世纪80年代初在马尾松林采伐迹地营造的人工林, 林龄约为37年。木荷林是20世纪90年代初在马尾松林火烧迹地上营造的人工林, 林龄约为29年。马尾松-香樟混交林是20世纪80年代初在马尾松林下通过星状补植香樟形成的, 平均林龄50年。马尾松-木荷混交林是20世纪90年代初建设生物防火隔离带时, 在马尾松林下带状栽植木荷形成的, 平均林龄45年。
1.2 研究方法 1.2.1 样地设置2018年6月, 选择土壤、地形条件基本一致的代表性地点, 每种林型设置规格为20 m × 20 m的典型样地各4个, 共20个。
1.2.2 土壤取样及分析2018年8月, 在各个样地内按“S”型布设10个取样点, 每个取样点挖取一个土壤剖面, 分别采集腐殖质层(O层)和淋溶层(A层)的土壤样品。每个样地10个点的土壤分层混合, 每层一个混合土壤样品, 分别表示为Pi_O(马尾松纯林O层土壤), Pi_A(马尾松纯林A层土壤), Ci_O(香樟纯林O层土壤), Ci_A(香樟纯林A层土壤), Sc_O(木荷纯林O层土壤), Sc_A(木荷纯林A层土壤), Pi_Ci_O(马尾松-香樟混交林O层土壤), Pi_Ci_A(马尾松-香樟混交林A层土壤), Pi_Sc_O(马尾松-木荷混交林O层土壤), Pi_Sc_A(马尾松-木荷混交林A层土壤)。采集的土壤分成两部分, 一部分运输至实验室, 用于土壤性质的分析;另一部分-80℃保存, 用于土壤真菌群落结构的分析。
土壤性质测定:土壤pH测定采用电位法;土壤阳离子交换量(CEC)采用草酸铵-氯化铵浸提后用半微量凯氏定氮法测定;土壤有机碳(SOC)用重铬酸钾容量-外加热发测定;土壤全氮(TN)利用H2SO4-H2O2消煮后半微量定氮法测定;土壤全磷(TP)采用NaOH熔融-钼锑抗比色法分析;土壤全钾(TK)利用NaOH熔融-火焰光度法测定;氨氮和硝氮则采用0.01mol/L氯化钙浸提后利用AA3流动注射分析仪进行分析测定。
DNA提取和聚合酶链式反应(PCR)扩增:根据试剂盒E.Z.N.A.® soil DNA kit (Omega Bio-tek, 诺克罗斯,乔治亚州,美国)说明书进行微生物群落总DNA抽提, 使用1%的琼脂糖凝胶电泳检测DNA的提取质量, 使用NanoDrop2000测定DNA浓度和纯度;使用ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS2R(5′-GCTGCGTTCTTCATCG ATGC-3′)对ITS1基因进行PCR扩增, 扩增程序如下:95℃预变性3 min, 37个循环(95℃变性30 s, 55℃退火30 s, 72℃延伸45 s), 然后72℃稳定延伸10 min, 最后在4℃进行保存(PCR仪:ABI GeneAmp®9700型)。PCR反应体系为:5×TransStart FastPfu缓冲液4 μL, 2.5 mmol/L dNTPs 2 μL, 上游引物(5 μmol/L)0.8 μL, 下游引物(5uM)0.8 μL, TransStart FastPfu DNA聚合酶0.4 μL, 模板DNA 10 ng, ddH2O补足至20 μL。每个样本3个重复。
Illumina Miseq测序:将同一样本的PCR产物混合后使用2%琼脂糖凝胶回收PCR产物, 利用试剂盒AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, 联合市,加利福尼亚州,美国) 进行回收产物纯化, 经2%琼脂糖凝胶电泳检测, 并用QuantusTM Fluorometer (Promega, 美国) 对回收产物进行检测定量。使用NEXTflexTM Rapid DNA-Seq Kit(Bioo Scientific, 美国)进行建库:(1)接头链接;(2)使用磁珠筛选去除接头自连片段;(3)利用PCR扩增进行文库模板的富集;(4)磁珠回收PCR产物得到最终的文库。利用Illumina公司的Miseq PE300平台进行测序(上海美吉生物医药科技有限公司)。
1.2.3 数据处理及统计分析(1) 土壤性质数据
本文目的之一是探讨不同林型与土壤酸化间的关系, 为了更直观地表现不同林型之间土壤性质的差异, 将各林型不同土层的土壤合并后运用SPSS18对土壤性质进行单因素方差分析(ANOVA)和Tukey′s显著性检验(HSD), 检测不同林型间土壤酸化指标和土壤养分之间的显著性水平。
(2) 土壤真菌群落数据
使用fastp[24](https://github.com/OpenGene/fastp, version 0.20.0)软件对原始测序序列进行质控, 使用FLASH[25](http://www.cbcb.umd.edu/software/flash, version 1.2.7)软件进行拼接:(1)过滤测序仪单次测序所得到的碱基序列(简称“序列reads”)尾部质量值20以下的碱基, 设置50bp的窗口, 如果窗口内的平均质量值低于20, 从窗口开始截去后端碱基, 过滤质控后50bp以下的reads, 去除含N碱基的reads;(2)根据两条具有物理关系的一对序列(PE reads)之间的重叠(overlap)关系, 将成对reads拼接(merge)成一条序列, 最小overlap长度为10 bp;(3)拼接序列的overlap区允许的最大错配比率为0.2, 筛选不符合序列;(4)根据序列首尾两端的条码(barcode)和引物区分样品, 并调整序列方向, barcode允许的错配数为0, 最大引物错配数为2。
使用UPARSE[26]软件(http://drive5.com/uparse/, version 7.1), 根据97%[26—27]的相似度对序列进行操作分类单元(OTU)聚类并剔除嵌合体。利用RDP分类器[28](http://rdp.cme.msu.edu/, version 2.2) 对每条序列进行物种分类注释, 比对unite8.0/its_fungi, 设置比对阈值为70%。
本研究利用美吉生物云平台(https://cloud.majorbio.com/)完成高通量测序数据分析。利用Mothur软件计算Alpha多样性指数(Shannon, Sobs, Chao1, Ace), 为了更直观地表现不同林型之间土壤真菌群落多样性的差异, 将各林型不同土层的土壤合并后运用SPSS18对多样性指数进行单因素方差分析。利用R软件(version3.3.1)基于Bray-cutis距离算法进行主坐标分析(PCoA分析)区分不同林型土壤真菌群落结构特征, 结合PERMONAVA分析确定林型、土层对土壤真菌群落结构的影响。用97%相似性的样本OTU表进行消除趋势对应分析(DCA), 其结果显示土壤真菌数据适合用冗余分析(RDA分析)探讨土壤性质与土壤真菌群落结构的相互关系。为找出造成不同林型土壤真菌群落差异的主要菌群, 利用SPSS18对合并土层的真菌属水平分类单位丰度进行单因素方差分析。
2 结果与分析 2.1 土壤酸化和土壤养分状况将Pi改造成Ci以及Pi_Ci后, 土壤pH值显著提高0.43个单位, 而NH4 ∶ NO3则显著降低三倍;而改造成Sc和Pi_Sc后pH值没有显著变化, 但Sc的NH4 ∶ NO3与Pi相比显著提高了约35%。Pi的改造对土壤SOC和TN含量都没有显著影响, 但改造成Ci显著提高了TP含量, 而改造成Sc和Pi_Sc后TP、TK含量均显著低于Pi(表 1)。
| 林型Forest Types | pH | CEC/(cmol/kg) | NH4∶NO3 | SOC/(g/kg) | TN/(g/kg) | TP/(g/kg) | TK/(g/kg) |
| Pi | 4.07±0.04b | 8.42±1.11 | 1.70±0.21b | 47.31±12.23 | 2.58±0.62 | 0.39±0.03b | 7.79±0.11a |
| Ci | 4.50±0.04a | 8.45±0.84 | 0.55±0.05c | 39.35±7.43 | 2.82c±0.49 | 0.55±0.02a | 7.92±0.12a |
| Sc | 4.09±0.04b | 6.73±1.19 | 2.62±0.31a | 40.31±9.24 | 2.19±0.55 | 0.20±0.02c | 2.53±0.07c |
| Pi_Ci | 4.50±0.01a | 8.15±1.19 | 0.60±0.06c | 41.71±10.96 | 2.74±0.65 | 0.42±0.03b | 7.91±0.10a |
| Pi_Sc | 3.99±0.07b | 8.86±1.56 | 2.16±0.03ab | 52.37±12.81 | 2.80±0.75 | 0.25±0.03c | 3.02±0.04b |
| Pi:马尾松纯林;Ci:香樟纯林;Sc:木荷纯林;Pi_Ci:马尾松香樟混交林;Pi_Sc:马尾松木荷混交林;不同字母表示不同林型之间差异显著(P < 0.05); pH:酸碱度Potential of hydrogen;CEC:土壤阳离子交换量Soil cation exchange capacity;NH4 ∶ NO3:硝态氮与铵态氮的比值;SOC:土壤有机碳Soil organic carbon;TN:土壤全氮Total nitrogen;TP:土壤全磷Total phosphorus;TK:土壤全钾Total potassium | |||||||
通过对不同林型不同土层的土壤样本进行高通量测序, 按最小样本序列上进行样本序列抽平处理后, 共得到优化序列2105668条, 经分析属于7门、29纲、88目、216科、451属、727种、3713OTUs。
2.2.1 真菌群落多样性除Shannon指数外, Chao1、Sobs及Ace指数均以Ci林型最高, 且Chao1指数显著高于Pi和Pi_Sc样本, Sobs指数显著高于Pi_Ci和Pi_Sc, Ace指数显著高于Pi。虽然Sc林型的Shannon指数最高, 但Pi、Ci以及Pi_Ci与其没有显著差异(表 2)。所以, 总体来讲, 五种林型中Ci林型土壤真菌多样性更丰富。相关性分析表明(表 3), 土壤pH值与Chao1和Sobs指数显著正相关;CEC与Shannon、Chao1、Sobs和Ace均呈正相关;而NH4 ∶ NO3则与Chao1和Sobs呈显著负相关;TK与Chao1指数显著正相关。
| 林型Forest types | Chao1指数Chao1 index | Ace指数Ace index | 生态优势度指数Sobs index | 香农指数Shannon index |
| Pi | 760±38bc | 723±50b | 521.25±8.63ab | 3.91±0.20ab |
| Ci | 1140±99a | 944±65a | 621.25±25.73a | 3.94±0.14ab |
| Sc | 908±75ab | 783±80ab | 543.00±29.68ab | 4.27±0.22a |
| Pi_Ci | 1065±145a | 779±52ab | 474.25±25.15b | 3.74±0.32ab |
| Pi_Sc | 582±59c | 799±55ab | 499.25±19.63b | 3.46±0.09b |
| 不同字母表示不同林型之间差异显著(P<0.05) | ||||
| 土壤性质Soil properties | Chao指数Chao index | Ace指数Ace index | 生态优势度指数Sobs index | 香农指数Shannon index |
| pH | 0.446** | 0.416** | ||
| CEC | 0.337* | 0.762** | 0.358* | 0.522** |
| NH4∶NO3 | -0.495** | -0.446** | ||
| TK | 0.361* | |||
| *在0.05水平上显著相关;**在0.01水平上显著相关 | ||||
从图 1可以看出, 在Ci和Pi_Ci两林型O层和A层土壤样品中的优势物种表现一致, 子囊菌门(Ascomycota)相对丰度最高(平均为43.44%), 其次为被孢霉门(Mortierellomycota)(18.24%), 担子菌门(Basidiomycota)和罗兹菌门(Rozellomycota)相对低一些(平均相对丰度分别为11.11%和6.06%)。而Pi、Sc及Pi_Sc的O层和A层土壤样品中, 相对丰度排在前两位的优势物种比较一致, 分别是子囊菌门(37.72%)和担子菌门(22.63%)。而相对丰度排在第三位的物种在O层和A层中表现不同, O层中为罗兹菌门(平均22.83%), 而A层中相对丰度排在第三位的则是被孢霉门(平均为8.39%)。子囊菌门在各样本中分布比例比较均衡(25.20—63.16%);担子菌门分布比例差别较大, Pi_A中分布比例达48.62%, 而在Pi_Sc_O中分布比例只有(6.13%);被孢霉门在Ci_O中分布比例最大(32.33%), 同样在Pi_Sc_O中分布比例最少(2.99%);而Pi_Sc_O中罗兹菌门分布比例高达35.12%, 而在Pi_A中分布比例只有1.61%。由此可见, 不同林型的不同土层样本之间土壤真菌群落组成存在差异。

|
| 图 1 不同林型不同土层土壤真菌门水平群落组成 Fig. 1 Soil fungal communities at the phylum level under different five forest types Pi_O, 马尾松纯林O层土壤; Pi_A, 马尾松纯林A层土壤; Ci_O, 香樟纯林O层土壤; Ci_A, 香樟纯林A层土壤; Sc_O, 木荷纯林O层土壤; Sc_A, 木荷纯林A层土壤; Pi_Ci_O, 马尾松-香樟混交林O层土壤; Pi_Ci_A, 马尾松-香樟混交林A层土壤; Pi_Sc_O, 马尾松-木荷混交林O层土壤; Pi_Sc_A, 马尾松-木荷混交林A层土壤; O层:腐殖质层土壤;A层:淋溶层土壤 |
主坐标分析(PCoA)结果表明(图 2), PC1轴解释了土壤真菌群落结构变异量的21.97%, 且该轴将不同林型的土壤真菌群落区分为两个大的类群, 由图 2中可以看出, Pi、Sc、Pi_Sc的土壤真菌群落分布在PC1轴的右侧, 而Ci和Pi_Ci则分布在左侧。而PC2轴解释了土壤真菌群落结构变异量的19.68%, 且不同土层在PC2轴上区分开来, O层基本分布在PC2轴的下半部分, 而A层则聚集在PC2轴的上半部分。置换多元(因素)方差分析(PERMANOVA)分析结果表明, 森林类型、土层及两者交互作用均对土壤真菌群落结构产生显著影响(P=0.001);林型和土层交互作用的R2值为0.84041, 林型的R2值为0.4112, 土层的R2值为0.17492, 说明林型对土壤真菌群落结构的影响大于土层。

|
| 图 2 不同林型不同土层土壤真菌群落PCoA分析 Fig. 2 Principal coordinates analysis of soil fungal communities PCoA分析:主坐标分析Principal co-ordinates analysis; PC: 主成分Principal component |
RDA分析结果(图 3)表明, 土壤pH值对真菌群落影响最大(P=0.001), 其次为CEC和SOC(P值分别为0.002和0.003), TN含量也显著影响真菌群落结构(P=0.004), 而TP和TK对真菌群落结构的影响不显著。

|
| 图 3 不同林型不同土层土壤真菌群落与土壤性质的RDA分析 Fig. 3 Redundancy analyses to reveal soil fungal communities in different forest types and correlations with edaphic properties RDA分析:冗余分析Redundancy analysis; 环境因子箭头的长短可以代表环境因子对于物种数据的影响程度的大小; NH4_NO3; 硝态氮与铵态氮的比值, 即NH4 ∶ NO3; pH:酸碱度Potential of hydrogen;TP:土壤全磷Total phosphorus;TN:土壤全氮Total nitrogen;CEC:土壤阳离子交换量Soil cation exchange capacity;SOC:土壤有机碳Soil organic carbon |
通过分析发现, 除Pi_Ci外, 其他林型都有一些特征菌(表 4)。Russula、Geminibasidium、Tomentella、Sebacina属及Russulaceae和Herpotrichiellaceae科在Pi土壤中的相对丰度显著高于其他四个林型, 且其他四个林型中这些真菌的丰度很低或未检出。Ci土壤中的Mortierella属相对丰度是Pi、Sc及Pi_Sc三个林型的三倍之多, 且Hygrocybe属只出现在Ci林型中。而Sc土壤中Cladophialophora、Sarcodon、Paecilomyces、Chloridium及Chamaeleomyces的相对丰度显著高于其他林型。而Venturiales目和Penicillium属在Pi、Sc及Pi_Sc中丰度都相对较高, 且Pi_Sc中这两类真菌含量显著高于Ci和Pi_Ci, 且Ci和Pi_Ci中未检出Venturiales。
| 土壤真菌Soil fungus | Pi | Ci | Sc | Pi_Ci | Pi_Sc |
| Russula | 10.82±3.99a | 0.00±0.00b | 0.01±0.01b | 0.00±0.00b | 0.26±0.10b |
| Russulaceae | 7.73±3.14a | 0.00±0.00b | 0.00±0.00b | 0.69±0.28b | 0.00±0.00b |
| Geminibasidium | 3.90±1.51a | 0.12±0.07b | 0.14±0.06b | 0.03±0.01b | 0.07±0.02b |
| Tomentella | 2.35±0.83a | 0.00±0.00b | 0.00±0.00b | 0.01±0.00b | 0.00±0.00b |
| Sebacina | 1.13±0.27a | 0.00±0.00b | 0.00±0.00b | 0.00±0.00b | 0.00±0.00b |
| Herpotrichiellaceae | 0.61±0.29a | 0.00±0.00b | 0.00±0.00b | 0.00±0.00b | 0.00±0.00b |
| Mortierella | 7.86±1.47b | 22.93±3.96a | 7.92±0.87b | 13.84±2.53ab | 6.18±1.53b |
| Hygrocybe | 0.00±0.00b | 1.53±0.64a | 0.00±0.00b | 0.00±0.00b | 0.00±0.00b |
| Cladophialophora | 0.52±0.20b | 0.53±0.17b | 2.44±0.93a | 0.63±0.23b | 0.10±0.04b |
| Sarcodon | 0.00±0.00b | 0.00±0.00b | 4.12±1.96a | 0.00±0.00b | 0.00±0.00b |
| Paecilomyces | 0.18±0.06b | 0.76±0.28ab | 1.76±0.69a | 0.70±0.26ab | 0.04±0.01b |
| Chloridium | 0.21±0.04b | 0.38±0.09b | 1.44±0.39a | 0.16±0.05b | 0.39±0.10b |
| Chamaeleomyces | 0.03±0.01b | 0.10±0.04b | 0.68±0.24a | 0.20±0.09b | 0.03±0.01b |
| Venturiales | 0.45±0.18ab | 0.00±0.00b | 1.66±0.64ab | 0.00±0.00b | 2.17±0.84a |
| Penicillium | 6.54±1.05ab | 0.84±0.34b | 4.28±0.87ab | 0.94±0.33b | 11.05±4.38a |
| 不同字母表示不同林型之间差异显著(P<0.05) | |||||
表 5相关性分析表明, Ci中含量丰富的Mortierella和Hygrocybe与土壤pH值、总磷含量及硝态氮含量呈显著正相关, 且Mortierella与土壤NH4 ∶ NO3呈显著负相关。Pi中含量高的Russulaceae与土壤TN、SOC及NO3-N呈负相关, 而Sebacina与pH呈显著负相关。Sc中相对丰度最高的Sarcodon、Chloridium及Chamaeleomyces均与TK呈显著负相关, 后两者与NH4-N呈显著正相关;且Sarcodon与CEC、TN、TP及TK呈显著负相关, 与NH4 ∶ NO3呈正相关。而Venturiales除了与pH无相关性、与NH4 ∶ NO3呈正相关外, 与其他所有土壤指标呈显著负相关关系;且Penicillium也与大多数土壤性质呈显著负相关, 与pH、SOC、及NH4-N无相关性。
| 土壤真菌Soil fungus | pH | CEC | NH4∶NO3 | TN | TP | TK | SOC | NH4-N | NO3-N |
| Mortierella | 0.515 (0.001) |
-0.477 (0.002) |
0.426 (0.006) |
0.430 (0.006) |
0.392 (0.012) |
||||
| Hygrocybe | 0.461 (0.003) |
0.431 (0.006) |
0.671 (<0.001) |
||||||
| Russulaceae | -0.331 (0.037) |
-0.329 (0.038) |
-0.312 (0.050) |
||||||
| Sebacina | -0.314 (0.049) |
0.320 (0.044) |
|||||||
| Cladophialophora | 0.430 (0.006) |
||||||||
| Sarcodon | -0.402 (0.010) |
0.592 (<0.001) |
-0.328 (0.039) |
-0.426 (0.006) |
-0.362 (0.022) |
||||
| Chloridium | -0.527 (<0.001) |
0.491 (0.001) |
|||||||
| Chamael eomyces | -0.348 (0.028) |
0.358 (0.023) |
|||||||
| Venturiales | -0.612 (<0.001) |
0.587 (<0.001) |
-0.597 (<0.001) |
-0.710 (<0.001) |
-0.521 (0.001) |
-0.508 (0.001) |
-0.384 (0.015) |
-0.532 (<0.001) |
|
| Penicillium | -0.357 (0.024) |
-0.389 (0.013) |
-0.520 (0.001) |
-0.330 (0.038) |
-0.381 (0.015) |
||||
| NH4-N:铵氮;NO3-N:硝氮;括号内数字为P值 | |||||||||
土壤酸化及其恢复是一个复杂的过程, 很难用一个单一指标来进行定量描述[29], 而土壤pH是土壤性质最重要的因素之一[30], 并且直接受酸沉降的影响。重庆铁山坪酸雨区将马尾松纯林改造成香樟纯林或马尾松-香樟混交林后显著提高土壤pH, 说明香樟能有效缓解土壤的酸化。土壤中硝态氮和铵态氮的比例也会对土壤酸化产生影响。研究表明当NH4 ∶ NO3低于1时, 说明土壤中以NO3-N为主, 则植物吸收的NO3-N比NH4-N多;而当土壤中NH4-N比例高时, 则植物根系吸收NH4+释放H+, 则根际酸化会加剧, 从而加重铝毒影响盐基离子的吸收[31]。在本研究中, 香樟纯林和马尾松-香樟混交林的NH4 ∶ NO3均显著低于马尾松纯林且均低于1, 说明这两个林型中植物以吸收NO3-N为主, 不会加剧土壤酸化;而马尾松纯林的NH4 ∶ NO3大于1, 且改造成木荷纯林和马尾松-木荷混交林后比值均显著增加, 说明会加剧根际酸化, 影响盐基离子吸收, 且土壤TP、TK也显著降低(表 1), 均不利于植物生长。
土壤微生物是生态系统中重要的分解者, 其多样性和组成在土壤健康调控中起关键作用[32]。而真菌是土壤微生物的重要组成部分, 在营养物质循环、能量流动过程中起着重要作用, 土壤真菌是评价土壤质量的关键指标[33]。在酸沉降区域, 对受损马尾松纯林进行阔叶林替代或林下补值阔叶树种形成针阔混交林是常用的营林措施[18]。研究发现种植香樟, 如香樟纯林和马尾松-香樟混交林, 其土壤细菌群落结构和组成优于马尾松纯林、木荷纯林以及马尾松-木荷混交林, 有利于缓解土壤酸化[18, 20]。
研究发现, 香樟纯林的土壤真菌多样性高于其他林型, 这应该与香樟纯林土壤酸化没有其他林型严重有关;且多样性指数与土壤酸化指标(pH值、CEC、NH4 ∶ NO3)显著相关, 说明土壤酸化缓解后有利于土壤真菌多样化, 这与对细菌多样性的影响一致[20]。张义杰等[34]发现, 在酸化土壤中施用生石灰后能提高土壤pH的同时, 也显著增加了真菌多样性指数, 这与本文研究结果一致。土壤真菌群落多样性发生变化的同时, 不同林型的真菌群落结构也发生显著变化, 而且从PCoA图可以看出土壤O层和A层的真菌群落结构分化明显, Stone等[35]同样发现土壤深度是影响微生物群落结构的主要因子, 这主要是由于不同深度土壤理化性质的差异造成的[36], 而这种理化性质之间的差异主要是由于地上凋落物和细根周转对表层土壤的影响要大于深层土壤[37]。从PCoA图上还可以看出, 不同林型的真菌聚成两大类, 香樟纯林和马尾松-香樟混交林的真菌群落结构更相似, 而马尾松纯林、木荷纯林与马尾松-木荷混交林的真菌群落结构更接近, 这与五个林型下土壤细菌群落结构表现一致[20]。同样, RDA分析结果表明, 反应土壤酸化特征的pH、CEC对真菌群落结构的影响最显著, 真菌群落多样性与这些指标呈正相关, 表明土壤真菌群落多样性和结构的差异与不同林型下土壤酸化程度存在密切相关性。其次, SOC、TN两项土壤性质也同样显著影响真菌群落结构, 这与刘立玲等[38]的研究结果是一致的, 均认为SOC、TN与真菌群落结构的变化密切相关[39]。这是因为碳和氮是真菌的营养元素, 为真菌提供养分来源。同时土壤的碳氮水平影响着真菌分解有机物的酶的合成, 从而对真菌群落结构产生影响[40]。又有研究表明, 无论是松树林、橡树林还是草地, 土壤的SOC和TN含量随着土层变深而递减, 其垂直分布呈“倒三角”趋势[41], 这也很好地解释了PCoA图中显示的土壤O层和A层的真菌群落结构分化明显的现象。
通过对不同林型土壤真菌多样性差异的分析, 发现在香樟纯林中Mortierella和Hygrocybe这两属真菌明显高于其他林型。Mortierella为溶磷菌, 是参与土壤磷循环的重要微生物类群, 能增加土壤中有效磷含量, 对植物生长有促进作用[42], 而且也有利于其他微生物对磷的吸收利用[43]。在本研究中, Mortierella与土壤有效磷呈显著正相关(表 4), 表明该菌的丰度越高土壤中的磷含量越多, 这与土壤总磷含量结果也一致, 从而有利于植物生长。Zhang等[44]研究证实土壤酸化显著降低了Mortierella的相对丰度, 这与本文研究结果一致, 相关性分析表明该菌丰度与土壤pH呈显著正相关, 土壤pH越低的土壤中该菌丰度相对较低。另外, 有研究表明, Mortierella属的很多种类参与降解植物残体以及土壤有机污染物的降解, 能有效促进土壤碳循环, 保持土壤的健康状况[45—46];且该属真菌对土传病菌镰孢菌(Fusarium spp.)有一定的抑制能力[47—48]。因此, 在受损马尾松林地种植香樟后, Mortierella丰度大大提高, 有利于土壤健康和植物生长。只在香樟林中出现的Hygrocybe的营养模式和功能还不甚清楚[49]。
马尾松是典型的外生菌根树种。本文在马尾松纯林土壤中检测出Russulaceae、Russula、Tomentella以及Sebacina的相对丰度较高, 而在其他林型中相对丰度则较低或未检出。Russula不仅是外生菌根真菌, 也是嗜酸菌[50], 偏好酸性环境, 同时也产生丁酸[51]。作为外生菌根菌Sebacina属真菌能在逆境中提高宿主植物对土壤中营养元素的吸收, 有利于宿主在逆境条件下存活[52]。Sebacina属真菌也是一类嗜酸菌, 出现在低pH值和高腐殖质环境中[53], 这与本研究结果一致, 即Sebacina相对丰度与土壤pH值呈负相关, 与SOC呈正相关。Geminibasidium为腐生菌, 在马尾松纯林中丰度较高, 该林型土壤pH值也是五个林型中较低的。王楠等[54]在对毛竹阔叶林土壤真菌结构的研究中发现, 模拟酸雨胁迫下Geminibasidium相对含量发生显著变化, 在酸度最高的酸雨处理下其相对丰度最高, 由此认为该属真菌可以作为酸雨胁迫下土壤真菌群落结构变化的指示物种之一。
木荷纯林中相对丰度较高的几种真菌多为病原真菌。Cladophialophora是一种植物病原菌, 可引起宿主叶片产生斑点[55];Paecilomyces除作为昆虫病原真菌能侵染昆虫外, 还可以寄生在植物病原菌和植物病原线虫上, 对生物防治有重要作用[56];同样也是植物病原菌[57], 以及产酸菌[58], 可产生赤霉酸[59]和醋酸[60]。Chloridium是一类异养硝化菌, 能在酸性土壤中调控土壤异养硝化作用, 其丰度与土壤异养硝化速率呈显著正相关, 随土壤酸度增加而增加[44]。这与本文结果一致。Chloridium丰度与土壤NH4-N呈正相关, 而硝化作用的底物是NH4+, 更多的底物存在才能促进异样消化速率。
本文研究发现Venturiales和Penicillium真菌在马尾松纯林、木荷纯林以及马尾松-木荷混交林土壤中相对丰度比较高。Venturiales目中许多真菌都是重要的植物病原菌[61]。而Penicillium在自然界中分布极其广泛, 促进碳氮磷等多种元素循环, 是最有力的N2O产生菌之一[62], 因此, 在马尾松、木荷及其混交林土壤中该菌含量丰富可能会促进N2O的排放。Penicillium也是嗜酸菌和产酸菌, 能产生大量有机酸[58]。在木荷纯林及其与马尾松混交林土壤中发生大量产酸菌, 可进一步加速土壤酸化, 丰富的植物病原菌则会对森林健康产生威胁。
4 结论(1) 马尾松纯林改造成香樟林后可缓解土壤酸化, 提高土壤养分;而改造成木荷林后可能进一步加剧土壤酸化, 降低土壤养分含量。
(2) 不同林型土壤真菌群落多样性以香樟林最为丰富, 且土壤pH值、CEC和NH4 ∶ NO3显著影响真菌多样性。
(3) 林型和土壤厚度均对真菌群落结构具有显著影响, 且林型影响土壤酸度进而对真菌群落结构产生显著影响。香樟纯林中有益菌较多, 而马尾松纯林以外生菌根真菌占优势, 木荷及其与马尾松的混交林则含有更多的植物病原真菌和产酸菌。
| [1] |
Duan L, Yu Q, Zhang Q, Wang Z F, Pan Y P, Larssen T, Tang J, Mulder J. Acid deposition in Asia: Emissions, deposition, and ecosystem effects. Atmospheric Environment, 2016, 146: 55-69. DOI:10.1016/j.atmosenv.2016.07.018 |
| [2] |
Yu H L, He N P, Wang Q F, Zhu J X, Gao Y, Zhang Y H, Jia Y L, Yu G R. Development of atmospheric acid deposition in China from the 1990s to the 2010s. Environmental Pollution, 2017, 231: 182-190. DOI:10.1016/j.envpol.2017.08.014 |
| [3] |
Liu F, Zhang Q, van der A R J, Zheng B, Tong D, Yan L, Zheng Y X, He K B. Recent reduction in NOx emissions over China: synthesis of satellite observations and emission inventories. Environmental Research Letters, 2016, 11(11): 114002. DOI:10.1088/1748-9326/11/11/114002 |
| [4] |
Li C, Zhang Q, Krotkov N A, Streets D G, He K B, Tsay S C, Gleason J F. Recent large reduction in sulfur dioxide emissions from Chinese power plants observed by the Ozone Monitoring Instrument. Geophysical Research Letters, 2010, 37(8): L08807. |
| [5] |
Liu M X, Huang X, Song Y, Tang J, Cao J J, Zhang X Y, Zhang Q, Wang S X, Xu T T, Kang L, Cai X H, Zhang H S, Yang F M, Wang H B, Yu J Z, Lau A K H, He L Y, Huang X F, Duan L, Ding A J, Xue L K, Gao J, Liu B, Zhu T. Ammonia emission control in China would mitigate haze pollution and nitrogen deposition, but worsen acid rain. Proceedings of the National Academy of Sciences of the United States of America, 2019, 116(16): 7760-7765. |
| [6] |
Lajtha K, Jones J. Trends in cation, nitrogen, sulfate and hydrogen ion concentrations in precipitation in the United States and Europe from 1978 to 2010:a new look at an old problem. Biogeochemistry, 2013, 116(1): 303-334. |
| [7] |
Larssen T, Carmichael G R. Acid rain and acidification in China: the importance of base cation deposition. Environmental Pollution, 2000, 110(1): 89-102. DOI:10.1016/S0269-7491(99)00279-1 |
| [8] |
Kang Y, Liu M, Song Y, Huang X, Yao H, Cai X, Zhang H, Kang L, Liu X, Yan X. High-resolution ammonia emissions inventories in China from 1980 to 2012. Atmospheric Chemistry and Physics, 2016, 16(4): 2043-2058. DOI:10.5194/acp-16-2043-2016 |
| [9] |
Wang S X, Zhao B, Cai S Y, Klimont Z, Nielsen C P, Morikawa T, Woo J H, Kim Y, Fu X, Xu J Y, Hao J M, He K B. Emission trends and mitigation options for air pollutants in East Asia. Atmospheric Chemistry and Physics, 2014, 14(13): 6571-6603. DOI:10.5194/acp-14-6571-2014 |
| [10] |
Driscoll C T, Wang Z. Ecosystem effects of acidic deposition. Encyclopedia of Water: Science, Technology, and Society, 2019, 1-12. |
| [11] |
Johnson A H, Moyer A, Bedison J E, Richter S L, Willig S A. Seven decades of calcium depletion in organic horizons of Adirondack forest soils. Soil Science Society of America Journal, 2008, 72(6): 1824-1830. DOI:10.2136/sssaj2006.0407 |
| [12] |
Sullivan T J, Fernandez I J, Herlihy A T, Driscoll C T, McDonnell T C, Nowicki N A, Snyder K U, Sutherland J W. Acid-base characteristics of soils in the adirondack mountains, New York. Soil Science Society of America Journal, 2006, 70(1): 141-152. DOI:10.2136/sssaj2005.0073 |
| [13] |
Sullivan T J, Lawrence G B, Bailey S W, McDonnell T C, Beier C M, Weathers K C, McPherson G T, Bishop D A. Effects of acidic deposition and soil acidification on sugar maple trees in the Adirondack Mountains, New York. Environmental Science & Technology, 2013, 47(22): 12687-12694. |
| [14] |
Sutton M, Howard C M, Erisman J, Billen G, Bleeker A, Grennfelt P, Grinsven H, Grizzetti B. The European nitrogen assessment: sources, effects and policy perspectives. Cambridge University Press, 2011.
|
| [15] |
Pannatier E G, Thimonier A, Schmitt M, Walthert L, Waldner P. A decade of monitoring at Swiss Long-Term Forest Ecosystem Research (LWF) sites: can we observe trends in atmospheric acid deposition and in soil solution acidity?. Environmental Monitoring and Assessment, 2011, 174(1): 3-30. |
| [16] |
Marx A, Hintze S, Sanda M, Jankovec J, Oulehle F, Dusek J, Vitvar T, Vogel T, van Geldern R, Barth J A C. Acid rain footprint three decades after peak deposition: Long-term recovery from pollutant sulphate in the Uhlirska catchment (Czech Republic). Science of the Total Environment, 2017, 598: 1037-1049. DOI:10.1016/j.scitotenv.2017.04.109 |
| [17] |
Chen Z, Wang Y H, Chen R S, Ni X Y, Cao J X. Effects of forest type on nutrient fluxes in throughfall, stemflow, and litter leachate within acid-polluted locations in southwest China. International Journal of Environmental Research and Public Health, 2022, 19(5): 2810. DOI:10.3390/ijerph19052810 |
| [18] |
王轶浩, 陈展, 周建岗, 张媛媛. 重庆酸雨区马尾松纯林改造对土壤酸化特征及团聚体稳定性的影响. 生态学报, 2021, 41(13): 5184-5194. |
| [19] |
向仁军. 中国南方典型酸雨区酸沉降特性及其环境效应研究[D]. 长沙: 中南大学, 2012.
|
| [20] |
Chen Z, Maltz M R, Zhang Y G, O'Brien B J, Neff M, Wang Y H, Cao J X. Plantations of Cinnamomum camphora (linn) presl with distinct soil bacterial communities mitigate soil acidity within polluted locations in southwest China. Forests, 2021, 12(6): 657. DOI:10.3390/f12060657 |
| [21] |
Gao C, Shi N N, Liu Y X, Peay K G, Zheng Y, Ding Q, Mi X C, Ma K P, Wubet T, Buscot F, Guo L D. Host plant genus-level diversity is the best predictor of ectomycorrhizal fungal diversity in a Chinese subtropical forest. Molecular Ecology, 2013, 22(12): 3403-3414. DOI:10.1111/mec.12297 |
| [22] |
Wardle D A. The influence of biotic interactions on soil biodiversity. Ecology Letters, 2006, 9(7): 870-886. DOI:10.1111/j.1461-0248.2006.00931.x |
| [23] |
Grantina L, Seile E, Kenigsvalde K, Kasparinskis R, Tabors G, Nikolajeva V, Jungerius P, Muiznieks I. The influence of the land use on abundance and diversity of soil fungi: comparison of conventional and molecular methods of analysis. Environmental and Experimental Biology, 2011, 9: 9-21. |
| [24] |
Chen S F, Zhou Y Q, Chen Y R, Gu J. Fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics, 2018, 34(17): i884-i890. DOI:10.1093/bioinformatics/bty560 |
| [25] |
Magoč T, Salzberg S L. FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics, 2011, 27(21): 2957-2963. DOI:10.1093/bioinformatics/btr507 |
| [26] |
Edgar R C. UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nature Methods, 2013, 10(10): 996-998. DOI:10.1038/nmeth.2604 |
| [27] |
Stackebrandt E, Goebel B M. Taxonomic note: a place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. International Journal of Systematic and Evolutionary Microbiology, 1994, 44(4): 846-849. DOI:10.1099/00207713-44-4-846 |
| [28] |
Wang Q, Garrity G M, Tiedje J M, Cole J R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Applied and Environmental Microbiology, 2007, 73(16): 5261-5267. DOI:10.1128/AEM.00062-07 |
| [29] |
Berger T W, Türtscher S, Berger P, Lindebner L. A slight recovery of soils from Acid Rain over the last three decades is not reflected in the macro nutrition of beech (Fagus sylvatica) at 97 forest stands of the Vienna Woods. Environmental Pollution: Barking, Essex: 1987, 2016, 216: 624-635. |
| [30] |
吴淑娟, 任运涛, 吴彩霞, 傅华. 草地土壤pH值测定方法比较研究. 土壤通报, 2018, 49(2): 343-348. |
| [31] |
Heij G, De Vries W, Posthumus A, Mohren G. Studies in Environmental Science, 1991, pp. 97-137, Elsevier.
|
| [32] |
朱永官, 彭静静, 韦中, 沈其荣, 张福锁. 土壤微生物组与土壤健康. 中国科学: 生命科学, 2021, 51(1): 1-11. |
| [33] |
Alguacil M M, Torrecillas E, Caravaca F, Fernández D A, Azcón R, Roldán A. The application of an organic amendment modifies the arbuscular mycorrhizal fungal communities colonizing native seedlings grown in a heavy-metal-polluted soil. Soil Biology and Biochemistry, 2011, 43(7): 1498-1508. DOI:10.1016/j.soilbio.2011.03.026 |
| [34] |
张义杰, 徐杰, 陆仁窗, 叶辰, 黄惠川, 杨敏, 何霞红, 朱书生. 生石灰对林下酸化土壤的调控作用及三七生长的影响. 应用生态学报, 2022, 33(4): 972-980. |
| [35] |
Stone M M, DeForest J L, Plante A F. Changes in extracellular enzyme activity and microbial community structure with soil depth at the Luquillo Critical Zone Observatory. Soil Biology and Biochemistry, 2014, 75: 237-247. DOI:10.1016/j.soilbio.2014.04.017 |
| [36] |
Ding X X, Liu G L, Fu S L, Chen H Y H. Tree species composition and nutrient availability affect soil microbial diversity and composition across forest types in subtropical China. CATENA, 2021, 201: 105224. DOI:10.1016/j.catena.2021.105224 |
| [37] |
Feng C, Wang Z, Ma Y, Fu S L, Chen H Y H. Increased litterfall contributes to carbon and nitrogen accumulation following cessation of anthropogenic disturbances in degraded forests. Forest Ecology and Management, 2019, 432: 832-839. DOI:10.1016/j.foreco.2018.10.025 |
| [38] |
刘立玲, 周光益, 党鹏, 陈洁, 尚海, 邱丽琼, 朱宁华. 湘西石漠化区3种造林模式土壤真菌群落结构差异. 生态学报, 2022, 42(10): 4150-4159. |
| [39] |
Li J R, Chen L, Wang H, Ouyang S, Liu X H, Lu J. Pattern and drivers of soil fungal community along elevation gradient in the Abies georgei forests of Segila mountains, Southeast Tibe. Global Ecology and Conservation, 2022, 39: e02291. |
| [40] |
高文慧, 郭宗昊, 高科, 薛晨, 昌梦园, 刘远, 王光利. 生物炭与炭基肥对大豆根际土壤细菌和真菌群落的影响. 生态环境学报, 2021, 30(1): 205-212. |
| [41] |
Jiang W T, Gong L, Yang L H, He S P, Liu X H. Dynamics in C, N, and P stoichiometry and microbial biomass following soil depth and vegetation types in low mountain and hill region of China. Scientific Reports, 2021, 11(1): 19631. |
| [42] |
Borrell A N, Shi Y, Gan Y, Bainard L D, Germida J J, Hamel C. Fungal diversity associated with pulses and its influence on the subsequent wheat crop in the Canadian prairies. Plant and Soil, 2017, 414(1): 13-31. |
| [43] |
李芳. 长期不同施肥条件下黄淮海平原旱作土壤微生物群落结构特征的演变[D]. 郑州: 河南农业大学, 2018.
|
| [44] |
Zhang Y, Dai S Y, Huang X Q, Zhao Y, Zhao J, Cheng Y, Cai Z C, Zhang J B. pH-induced changes in fungal abundance and composition affects soil heterotrophic nitrification after 30 days of artificial pH manipulation. Geoderma, 2020, 366: 114255. |
| [45] |
Osono T. Colonization and succession of fungi during decomposition of Swida controversa leaf litter. Mycologia, 2005, 97(3): 589-597. |
| [46] |
Ellegaard-Jensen L, Aamand J, Kragelund B B, Johnsen A H, Rosendahl S. Strains of the soil fungus Mortierella show different degradation potentials for the phenylurea herbicide diuron. Biodegradation, 2013, 24(6): 765-774. |
| [47] |
Xiong W, Li R, Ren Y, Liu C, Zhao Q Y, Wu H S, Jousset A, Shen Q R. Distinct roles for soil fungal and bacterial communities associated with the suppression of vanilla Fusarium wilt disease. Soil Biology and Biochemistry, 2017, 107: 198-207. |
| [48] |
Shen Z Z, Penton C R, Lv N N, Xue C, Yuan X F, Ruan Y Z, Li R, Shen Q R. Banana Fusarium wilt disease incidence is influenced by shifts of soil microbial communities under different monoculture spans. Microbial Ecology, 2018, 75(3): 739-750. |
| [49] |
Halbwachs H, Karasch P, Griffith G W. The diverse habitats of Hygrocybe-peeking into an enigmatic lifestyle. Mycosphere, 2013, 4(4): 773-792. |
| [50] |
Rineau F, Garbaye J. Effects of liming on ectomycorrhizal community structure in relation to soil horizons and tree hosts. Fungal Ecology, 2009, 2(3): 103-109. |
| [51] |
Zhang N, Li C, Niu Z H, Kang H Y, Wang M S, Zhang B, Tian H T. Colonization and immunoregulation of Lactobacillus plantarum BF_15, a novel probiotic strain from the feces of breast-fed infants. Food & Function, 2020, 11(4): 3156-3166. |
| [52] |
Ghimire S R, Craven K D. Enhancement of switchgrass (Panicum virgatum L.) biomass production under drought conditions by the ectomycorrhizal fungus Sebacina vermifera. Applied and Environmental Microbiology, 2011, 77(19): 7063-7067. |
| [53] |
Sigisfredo G, Kai R, Robert B, Franz O, Michael W. Phylogenetic diversity and structure of sebacinoid fungi associated with plant communities along an altitudinal gradient. FEMS Microbiology Ecology, 2013, 83(2): 265-278. |
| [54] |
王楠, 潘小承, 王传宽, 白尚斌. 模拟酸雨对毛竹阔叶林过渡带土壤真菌结构及其多样性的影响. 环境科学, 2020, 41(5): 2476-2484. |
| [55] |
Badali H, Gueidan C, Najafzadeh M J, Bonifaz A, van den Ende A H G G, de Hoog G S. Biodiversity of the genus Cladophialophora. Studies in Mycology, 2008, 61: 175-191. |
| [56] |
罗科, 张青文, 杨奇华. 拟青霉属真菌在植物保护中的研究利用. 世界农业, 1990(11): 32-33. |
| [57] |
Senthilkumar M, Anandham R, Krishnamoorthy R. Paecilomyces. Beneficial Microbes in Agro-Ecology. Amsterdam: Elsevier, 2020: 793-808.
|
| [58] |
Khan R, Gupta K. Screening and characterization of acid producing fungi from different mine areas of Chhattisgarh region. KAVAKA, 2018. |
| [59] |
El-Sheikh M A, Rajaselvam J, Abdel-Salam E M, Vijayaraghavan P, Alatar A A, Devadhasan Biji G. Paecilomyces sp. ZB is a cell factory for the production of gibberellic acid using a cheap substrate in solid state fermentation. Saudi Journal of Biological Sciences, 2020, 27(9): 2431-2438. |
| [60] |
Senthilkumar M, Anandham R, Krishnamoorthy R. Paecilomyces. Beneficial Microbes in Agro-Ecology. Amsterdam: Elsevier, 2020: 793-808.
|
| [61] |
Zhang Y, Crous P W, Schoch C L, Bahkali A H, Guo L D, Hyde K D. A molecular, morphological and ecological re-appraisal of Venturiales-a new order of Dothideomycetes. Fungal Diversity, 2011, 51(1): 249-277. |
| [62] |
Jirout J, Šimek M, Elhottová D. Fungal contribution to nitrous oxide emissions from cattle impacted soils. Chemosphere, 2013, 90(2): 565-572. |