 2023, Vol. 43
2023, Vol. 43文章信息
- 陈晓海, 李银康, 应紫薇, 郭清扬, 颉晓勇
- CHEN Xiaohai, LI Yinkang, YING Ziwei, GUO Qingyang, XIE Xiaoyong
- 人为干扰栖息地退化对潮间带中华鲎幼鲎种群数量的影响
- Effects of habitat degradation due to human activities on abundance of juvenile Tachypleus tridentatus in the intertidal zone
- 生态学报. 2023, 43(16): 6824-6832
- Acta Ecologica Sinica. 2023, 43(16): 6824-6832
- http://dx.doi.org/10.5846/stxb202205111319
-
文章历史
- 收稿日期: 2022-05-11
- 网络出版日期: 2023-04-20





2. 上海海洋大学水产与生命学院, 上海 201306;
3. 南方海洋科学与工程广东省实验室, 广州 511458
2. College of Fisheries and Life, Shanghai Ocean University, Shanghai 201306, China;
3. Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou), Guangzhou 511458, China
中华鲎(Tachypleus tridentatus)又称中国鲎、东方鲎、三棘鲎, 隶属于肢口纲(Merostomata)剑尾目(Xiphosura), 主要分布于太平洋西岸[1]。由中华鲎血液生产的鲎试剂是新冠疫苗等生物医药产品必需的质检试剂[2—3], 近些年鲎资源严重衰退, 2019年中华鲎在IUCN红色名录中被更新为濒危(EN)等级, 2021年被列入国家二级重点保护野生动物名录, 鲎资源保护逐渐成为热点研究课题[4]。
过度捕捞、广泛误捕[5—9]等人类活动被公认是鲎面临的威胁因素[10], 而鲎栖息地退化同样是造成中华鲎资源量骤减的重要原因[11—12]。因沿海地区工业化及自然海滩开发等涉海工程加大对潮间带的开发利用, 使滩涂面积减少, 生境退化, 导致鲎失去繁殖和育幼场所, 直接威胁鲎生存[13—15]。涉海工业污染物和生活污水排放, 严重污染鲎栖息地, 影响鲎产卵和幼体生长发育[16—17]。厘清其中主要威胁因素, 尤其是人为干扰下栖息地退化的影响, 是有效开展中华鲎资源保护的关键环节, 但截至目前尚未有量化幼鲎栖息地破坏方面的研究报道。
鲎栖息地破坏在地理空间上表征为潮间带景观格局的改变, 反映人类开发利用强度[18]。人为干扰度指数(human disturbance index, HDI)模型可用于评价特定区域内景观类型受到人为干扰活动的影响强度[19], 可据此量化潮间带幼鲎栖息地生境遭受不同人类干扰强度。本研究以北部湾潮间带区域为对象, 通过调查研究区域2014、2017和2020年夏季潮间带滩涂幼鲎种群数据, 利用遥感和GIS空间技术, 量化北部湾潮间带景观格局及人为干扰强度变化, 评估人类活动造成的栖息地退化对中华鲎幼鲎种群规模的影响, 为中华鲎栖息地保护和种群资源恢复提供科学依据。
1 研究方法 1.1 研究区概况北部湾(Beibu Gulf)为中国广东雷州半岛、海南岛和广西壮族自治区及越南之间的海域, 平均水深42m, 是当今中华鲎最主要栖息海域[1]。本研究以北部湾潮间带为范围(21°30′—21°28′N, 108°33′—108°45′E)。向陆方向, 以1990年滨海湿地陆上边界为基准;向海方向, 以近海海域6m等深线为边界。研究区域面积共计2407km2(图 1)。

|
| 图 1 研究区遥感影像及中华鲎调查点 Fig. 1 Remote sensing image of the study area and survey sites of T.tridentatus 1.中三墩;2.湿地公园;3.竹林盐场;4.坡尾底;5.沙田;6.榕根山 |
通过美国陆地资源卫星(Landsat)选取2014、2017和2020年landsat 8 OLI TRIS遥感影像(分辨率30m), 使用ENVI 5.3软件对遥感影像进行几何精矫正、大气矫正、图像增强、镶嵌等预处理, 并选取6、5、4波段合成彩色影像, 于美国国家海洋大气管理局国家地理数据中心(NOAA)获取全球地形模型(ETOPO1), 并通过Arcmap 9.3软件地理分析工具获取近海海域6m等深线。
结合Shin等人[20]、Morton和Lee[13]对幼鲎种群丰度调查的随机样方法。于2014、2017与2020年夏季在研究区域6个调查点(中三墩、湿地公园、竹林盐场、坡尾底、沙田、榕根山)潮间带随机设置1—4个样方。各样方间距离大于200m;样方内由4条长500m, 宽4m且垂直于海岸线的样线组成, 每条样线间距大于125m。以此获取中华鲎幼鲎种群丰度数据。
1.3 景观分类及人为干扰度指数结合《土地利用现状分类》(GB/ T 21010—2017)与湿地景观与干扰度分析[18—19, 21—22], 将研究区域滨海湿地土地利用性质分为三大类(无干扰型、半干扰型和全干扰型), 以此基础上细分成10种景观类型(表 1), 各类景观对应不同干扰强度系数。根据所构建研究区潮间带景观分类体系, 采用ENVI 5.3软件人机交互监督分类研究区景观类型, 并依据HDI公式[22]构建模型。HDI公式如下:
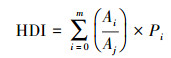
|
(1) |
| 一级分类 Level Ⅰ |
二级分类 Level Ⅱ |
含义 Definition |
人为干扰度指数 human disturbance index |
代码 Code |
| 无干扰(几乎无人为干扰) | 浅海水体 | 低潮不超过6m浅海水域 | 0.10 | A1 |
| Undisturbed (almost undisturbed by human) | 红树林 | 沿海生长红树植物的林地 | 0.15 | A2 |
| 滩涂 | 淤泥裸滩涂 | 0.17 | A3 | |
| 河口水域 | 河口区高低潮之间水域 | 0.20 | A4 | |
| 河流 | 潮流界以上淡水水域 | 0.23 | A5 | |
| 半干扰(人为、自然作用参半, 主要为农业、养殖业等生态系统) | 沙滩 | 滨海沙滩 | 0.30 | B1 |
| Partially disturbed (where human and nature impacts played equal roles, such as crop or fishery ecosystems) | 养殖 | 水产养殖池塘、鱼虾蟹养殖水面 | 0.63 | B2 |
| 耕地(或水田) | 水稻田、农用旱田 | 0.70 | B3 | |
| 全干扰(人造地物如公路等) | 居民点 | 农村居民地 | 0.95 | C1 |
| Completely disturbed (manmade entities like paved roads, etc) | 港口码头 | 渔业码头、商贸码头 | 0.98 | C2 |
式中, n为景观类型的数量, Ai为采样网格中第i种景观类型的面积(km2), Aj为采样网格总面积(km2), Pi为第i种景观类型所反映的干扰度强度系数。
采用Arcmap 9.3软件分析工具模块对研究区不同时期景观格局进行转移矩阵计算, 获得滨海湿地景观类型流转空间分布信息;通过渔网工具构建1km×1km的网格覆盖研究区域, 将HDI模型计算结果赋予每个网格中心点形成空间点集, 得到研究区不同时期人为干扰强度空间分布信息;对不同时期干扰度空间点集进行差值运算获得干扰度时空动态变化信息。
1.4 滨海湿地人为干扰指数与中国鲎幼鲎种群丰度的关系利用Arcmap 10.3软件按位置属性选取北部湾幼鲎调查点滨海湿地人为干扰度指数。使用SPSS 24.0软件Pearson法对各调查点人为干扰度指数与幼鲎种群丰度进行相关分析, 设置0.05为显著水平, 0.01为极显著水平。以HDI指数为预测变量, 丰度为响应变量进行曲线拟合建立对数、逆、二次项和幂回归方程, 并使用OriginPro 2021软件进行制图。于上述回归方程中选取最高决定系数(R2)作为最优回归方程。
对数回归方程:
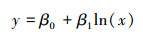
|
(2) |
逆回归方程:
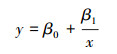
|
(3) |
幂回归方程:
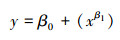
|
(4) |
二次项回归方程:
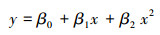
|
(5) |
式中, y为响应变量表示HDI指数, x为预测变量表示丰度, β0为常数项, β1为预测变量x对应的回归系数;(5)公式中x2为预测变量表示丰度的平方值, β2为其对应的回归系数。
2 结果 2.1 中华鲎幼鲎种群丰度情况北部湾研究区域各调查点中华鲎幼鲎数量随调查年份存在一定变化。各调查点中华鲎幼鲎丰度在每100m2 0.07到0.50只范围内, 普遍丰度较低(表 2)。
| 调查点 Survey sites |
2014年丰度 Abundance of 2014/(个/100m 2) |
2017年丰度 Abundance of 2017/(个/100m 2) |
2020年丰度 Abundance of 2022/(个/100m 2) |
| 中三墩 | 0.106 | 0.178 | 0.010 |
| 竹林盐场 | 0.096 | 0.158 | 0.020 |
| 湿地公园 | 0.078 | 0.069 | 0.085 |
| 坡尾底 | 0.088 | 0.783 | 0.080 |
| 沙田 | 0.256 | 0.806 | 0.440 |
| 榕根山 | 0.056 | 0.144 | 0.600 |
从2014—2020年, 滨海湿地无干扰类型景观面积减少17.03%(图 2), 其中滩涂和红树林景观共减少面积3.56%。无干扰转变为半干扰或全干扰类型景观中, 转化为养殖景观的面积比例较多, 浅海水体、红树林、滩涂、河口水域及河流分别向养殖景观转化面积91.35%、72.80%、76.06%、79.93%和42.38%(图 3)。半干扰与全干扰类型景观各增加面积48.87%和37.94%(图 2)。地理空间上, 不同类型景观转变为养殖区域主要集中于钦州湾茅尾海、北海市大风江水域及防城港市港口附近水域;转变为居民点和港口码头区域主要集中在钦州湾茅尾海沿岸(图 4)。

|
| 图 2 2014—2020北部湾潮间带不同景观类型斑块数量变化 Fig. 2 Changes in the number of patches of different landscape types in the intertidal zone of Beibu Gulf from 2014 to 2020 |

|
| 图 3 2014—2020北部湾潮间带无干扰类型景观面积流转 Fig. 3 Area dynamic transition of landsat in the intertidal zone of Beibu Gulf from 2014 to 2020 A1:浅海水体;A2:红树林;A3:滩涂;A4:河口水域;A5:河流;B1:沙滩;B2:养殖;B3:耕地(或水田);C1:居名点;C2:港口码头 |

|
| 图 4 2014—2020年北部湾潮间带景观变化空间分布 Fig. 4 Spatial distribution of landscape change in the intertidal zone of Beibu Gulf from 2014 to 2020 |
2014—2020年期间, 人类干扰度等级为无干扰的面积比例由72%减少至58%, 全干扰等级面积增加幅度不显著(图 5)。6年北部湾滨海湿地人为干扰空间分异特征显著(图 6)。2014年全干扰等级栅格零星分布, 主要集中于钦州湾、防城港市各大型港口附近, 半干扰等级栅格聚集面积较少, 分布较为分散。2020年半干扰等级栅格大面积聚集于钦州湾茅尾海和北海市大风江水域, 全干扰和半干扰等级栅格聚集于北部湾沿海, 形成明显的人为干扰带。6年来几乎北部湾滨海湿地全域呈现人为干扰逐渐增强趋势, 约64.72%滨海湿地的HDI指数不断增加, 其中6个中华鲎幼鲎调查点中, 竹林盐场、坡尾底、沙田和荣根山四个调查点的人类干扰强度增强(图 7)。

|
| 图 5 2014—2020北部湾潮间带人为干扰度等级构成变化 Fig. 5 Proportion of different HDI types in the intertidal zone of Beibu Gulf from 2014 to 2020 |

|
| 图 6 2014、2017和2020年北部湾潮间带人为干扰度空间分布 Fig. 6 Spatial distribution of HDI in the intertidal zone of Beibu Gulf at 2014, 2017 and 2020 year |

|
| 图 7 2014—2020年北部湾潮间带人为干扰度变化空间分布 Fig. 7 Spatial distribution of HDI′ s changes in the intertidal zone of Beibu Gulf from 2014 to 2020 |
经Preason相关性检验, 中华鲎幼鲎丰度与HDI指数间存在负显著相关性(Cor(丰度, HDI)=-0.535, P<0.05)。在对数、幂、二次项和逆回归方程中二次项回归模型决定系数(R2)最高(表 3), 得二次项回归方程y=1.6908-9.7905x+14.1944x2, 其中, 模型中的估计参数β1=-9.7905±3.7252;β2=14.1944±6.5943;β0=1.6908±0.4970(图 8)。回归方差分析结果显示中华鲎幼鲎丰度与HDI指数回归关系显著(F=6.2373, P<0.05), 决定系数为R2=0.4540。
| 回归方程 Regression models |
R2 | F | 显著性 Significance |
回归方程 Regression models |
R2 | F | 显著性 Significance |
|
| 对数Logarithmic | 0.3639 | 9.1521 | 0.0080 | 二次项Quadratic | 0.4540 | 6.2373 | 0.0107 | |
| 幂Power | 0.4244 | 11.7974 | 0.0034 | 逆回归Inverse | 0.3231 | 7.6368 | 0.0138 |

|
| 图 8 HDI与幼鲎丰度二次项回归方程曲线拟合 Fig. 8 Quadratic regression Curve estimation of HDI and abundantce of juvenile T.tridentatus |
中华鲎在靠近高潮线区域产卵, 幼鲎栖息于红树林附近泥滩[23—24], 无干扰类型景观和无干扰等级斑块适宜幼鲎生长。2014—2020年北部湾滨海湿地无干扰类型景观面积与无干扰等级斑块显著减少, 无干扰景观更多转化为人为干扰的养殖用地, 不断增强的人为干扰使幼鲎栖息地发生退化。即鲎栖息地由一个完整的、大面积的斑块逐渐分割成两个甚至多个不连续的、小面积的斑块, 对鲎种群产生不利影响。有研究表明, 岸上虾塘养殖模式阻碍了海陆交流造成近岸潮滩环境质量下降[5], 大面积滩涂养殖蛤蜊对红树林和海草植物群落造成危害, 使沿海生态环境“荒漠化”[25], 进而影响栖息此地的中华鲎产卵及幼鲎生存和觅食。Kwan等发现牡蛎养殖场大量的人工构筑物可能引起物理干扰[26], 改变幼鲎利用的潮间带生境, 阻碍幼鲎生长。
沿岸城市化加快了鲎栖息地消失的速度[27], 并使其生态环境质量进一步下降。无干扰类型景观中, 滩涂和红树林两类景观与幼鲎栖息地关系紧密[28—30], 因耕地、养殖、住宅、海岸带码头及工厂建设等需求, 北部湾滨海湿地于6年共损失3.56%滩涂和红树林面积;居民点和港口码头面积6年增加37.94%。居民点建设是影响景观生态风险格局的土地利用方式[31], 亦是公认对鲎造成重要影响的人类活动类型[10]。海岸带建设及港口码头造成的水环境污染对中华鲎卵和胚胎发育造成直接影响, 并间接减少潮间带底栖生物量[32—33]。因中华鲎在自然生态系统食物网中处于较弱势地位[34], 潮间带底栖生物量减少造成其觅食困难, 对幼鲎生存同样形成威胁。
人为因素(诸如城镇化建设、道路修建等)干扰导致的栖息地退化, 减少物种适宜生境面积, 使斑块间距与边缘地增多[35], 并增加环境污染物暴露风险, 使栖息地质量降低[36], 造成物种向外部非适宜生境扩散, 在改变适宜生境内群落结构同时增加生境内物种灭绝速率[37]。人为干扰下栖息地退化可能是幼鲎种群丰度下降的重要原因。北部湾及粤西一带海域容纳的中华鲎数量占中国整体蕴藏量的95%以上[2], 但近年来北部湾中华鲎数量正在急剧减少[38], 与20世纪90年代成鲎个体数量(60×104—70×104对)比较, 2013年中华鲎成体数量已锐减至不到30×104对[2]。本研究调查记录的中华鲎幼鲎平均丰度(0.23个/100m2)亦远低于Hu等在2008年的调查(1.17个/100m2)[39]。
3.2 人类干扰度指数与幼鲎种群丰度的相关性人为干扰度指数是一种基于土地利用状态变化的定量化人类活动强度评价模型, 揭示因人类活动及干扰产生的不同景观生态效应[40]。利用人类干扰度指数优化景观分布格局, 为生态环境风险防范提供科学依据。解若璠等探讨了湿地景观格局和人为干扰度的动态变化及响应[41], 潘晨等利用人为干扰度模型研究发现随人类干扰活动增强升金湖自然保护区景观破碎化加剧、相同区域间连通性减弱[42]。评价保护物种适宜栖息地方面, 任璘婧等通过人为干扰度模型分析长江口滩涂湿地景观变化对水鸟生境的影响, 发现鸻鹬类对人类干扰更加敏感, 中等强度的人类干扰在一定程度上扩大了雁鸭类的适宜生境面积[43]。
本研究中, 北部湾潮间带人类干扰度指数与幼鲎种群丰度相关性显著, 随HDI指数增加幼鲎种群丰度呈下降趋势, 潮间带HDI指数较高斑块不适合幼鲎生存。对两者回归建立模型决定系数均较低, 最大R2为0.4540, HDI指数仅可预测幼鲎种群丰度45.40%的变异。影响幼鲎种群丰度的因素除栖息地退化外, 还包括因食用、医药利用引起的过度捕捞或广泛的误捕行为等其他类型人为因素干扰。幼鲎对于栖息地的选择与环境因素中地理地貌(高程、坡度等)条件有关[44], 潮间带沉积物中叶绿素a及总有机碳含量也是决定幼鲎丰度与生长的重要因素[45]。因此后续需就幼鲎种群丰度与其他不同类型人为因素开展更为深入的研究。
3.3 北部湾潮间带中华鲎幼鲎栖息环境管理建议当前北部湾潮间带人类干扰不断增强, 幼鲎栖息地逐渐退化, 需引起相关管理部门重视。针对潮滩环境中养殖活动需优化沿岸海水养殖空间布局, 禁止养殖区域需依法严禁养殖, 有鲎生存的养殖区域应加强养殖尾水污染防控。针对全干扰类型景观, 建议合理控制沿岸工程建设规模。沿海农村居民点传统的渔业生产方式频繁与鲎直接接触, 容易对鲎造成误捕和伤害。可通过面向沿岸村民开展自然教育或公众科普, 提升其鲎保护意识, 减少鲎误捕和私宰等情况, 形成低成本高效益的中华鲎保护措施。
在人类干扰评估与潮间带开发利用规划基础上, 结合国家公园与HDI模型寻找适宜鲎生存地区, 合理规划建设保护区。在进行中华鲎人工放流增殖选址中可依据HDI指数高值斑块, 规避潮间带栖息地退化严重及人为干扰强烈的地区, 从放流地点选择角度提升增殖效果。对于已有鲎生存且HDI指数逐渐增强的潮滩, 可通过提高分辨率进行重点识别, 建议对6年内HDI指数增强的竹林盐场、坡尾底、沙田和荣根山4个调查点的潮滩尽早开展重点监控, 依科学数据指导潮间带幼鲎生境保护区建设和管理。
4 结论近6年来, 北部湾潮间带中华鲎幼鲎栖息地退化严重, 适宜幼鲎生存的无干扰景观(如红树林、滩涂等)不断减少, 且转化为养殖类型的景观较多;居民点及港口码头的面积增加。养殖、居民点及港口码头等非适宜鲎生存景观面积增加造成适宜栖息地减少, 同时降低生态环境质量, 不利于幼鲎生存。2014—2020年, 北部湾潮间带人为干扰逐渐加强, 无干扰等级HDI指数面积不断减少, 全干扰等级面积增加幅度不显著, 潮间带大部分面积HDI指数呈逐渐增强趋势。研究发现人为干扰度指数与潮间带幼鲎丰度间存在显著负相关, 可依据HDI指数优化幼鲎种群栖息地景观格局指导人工放流增值选址等鲎保护行动。
| [1] |
朱俊华, 吴宙, 冯炳斌, 邓帅帅, 甄文全, 廖永岩, 颉晓勇, Kwan K Y. 全球中华鲎资源保护现状及对策建议. 生物多样性, 2020, 28(5): 621-629. |
| [2] |
洪水根. 中国鲎生物学研究. 厦门: 厦门大学出版社, 2011: 154-155.
|
| [3] |
颉晓勇, 钟金香, 关杰耀, 贾晓平. 基于鲎试剂需求的中国鲎资源保护的迫切性研究. 中国渔业经济, 2021, 39(2): 109-116. DOI:10.3969/j.issn.1009-590X.2021.02.013 |
| [4] |
Yang H, Thompson J R, Flower R J. Save horseshoe crabs and coastal ecosystems. Science, 2019, 366(6467): 813-814. |
| [5] |
Laurie K, Chen C P, Cheung S G, Do V, Hsieh H, John A, Mohamad F, Seino S, Nishida S, Shin P, Yang M. Tachypleus Tridentatus (Errata Version Published in 2019). Gland: IUCN, 2019. |
| [6] |
Steele P, Bert T M, Johnston K H, Levett S. Efficiency of bycatch reduction devices in small otter trawls used in the Florida shrimp fishery. Fishery Bulletin, 2002, 100(2): 338-350. |
| [7] |
Supadminingsih F N, Wahju R I, Riyanto M. Composition of blue swimming crab Portunus pelagicus and horseshoe crab Limulidae on the gillnet fishery in Mayangan waters, Subang, West Java. AACL Bioflux, 2019, 12(1): 14-24. |
| [8] |
Smith D R, Brockmann H J, Beekey M A, King T L, Millard M J, Zaldívar-Rae J. Conservation status of the American horseshoe crab, (Limulus polyphemus): a regional assessment. Reviews in Fish Biology and Fisheries, 2017, 27(1): 135-175. DOI:10.1007/s11160-016-9461-y |
| [9] |
Meilana L, Fang Q H. Local knowledge-based study on the status of horseshoe crabs along the Indonesian coast. Regional Studies in Marine Science, 2020, 36: 101252. DOI:10.1016/j.rsma.2020.101252 |
| [10] |
Wang C C, Kwan K Y, Shin P K S, Cheung S G, Itaya S, Iwasaki Y, Cai L Z, Mohamad F, Fozi N F, Mat Zauki N A, Raman N J A, Chatterji A, Tripathy B, Sajan S, Min W W, Tan L J X, Supadminingsih F N, Wardiatno Y, Hsieh H L. Future of Asian horseshoe crab conservation under explicit baseline gaps: a global perspective. Global Ecology and Conservation, 2020, 24: e01373. DOI:10.1016/j.gecco.2020.e01373 |
| [11] |
李裕红, 颉晓勇, 关杰耀. 活化石中国鲎濒危现状及抢救性保护宣言. 湿地科学, 2018, 16(5): 690-692. |
| [12] |
鲍虞园, 叶国玲, 钟金香, 陈海刚, 李银康, 关杰耀, 颉晓勇. Hg2+对中国鲎幼体急性毒性及MT的诱导效应. 生态毒理学报, 2020, 15(6): 300-307. |
| [13] |
Morton B, Lee C N. Spatial and temporal distributions of juvenile horseshoe crabs (Arthropoda: Chelicerata) approaching extirpation along the northwestern shoreline of the New Territories of Hong Kong SAR, China. Journal of Natural History, 2010, 45(3/4): 227-251. |
| [14] |
翁朝红, 谢仰杰, 肖志群, 黄良敏, 李军, 王淑红, 张雅芝. 福建及中国其他沿岸海域中国鲎资源分布现状调查. 动物学杂志, 2012, 47(3): 40-48. |
| [15] |
Hsieh H L, Chen C P. Current status of Tachypleus tridentatus in Taiwan for red list assessment//Carmichael R H, Botton M L, Shin P K S, Cheung S G, eds. Changing Global Perspectives on Horseshoe Crab Biology, Conservation and Management. Cham: Springer, 2015: 383-396.
|
| [16] |
Botton M L, Itow T. The effects of water quality on horseshoe crab embryos and larvae//Tanacredi J T, Botton M L, Smith D, eds. Biology and Conservation of Horseshoe Crabs. New York: Springer, 2009: 439-454.
|
| [17] |
陈秋明. 厦门海域中国鲎资源的研究与保护. 环境科学与管理, 2009, 34(6): 9-11. |
| [18] |
梁发超, 刘黎明. 景观格局的人类干扰强度定量分析与生态功能区优化初探——以福建省闽清县为例. 资源科学, 2011, 33(6): 1138-1144. |
| [19] |
陈爱莲, 朱博勤, 陈利顶, 吴艳华, 孙然好. 双台河口湿地景观及生态干扰度的动态变化. 应用生态学报, 2010, 21(5): 1120-1128. |
| [20] |
Shin P K S, Li H, Cheung S G. Horseshoe crabs in Hong Kong: current population status and human exploitation//Tanacredi J T, Botton M L, Smith D, eds. Biology and Conservation of Horseshoe Crabs. New York: Springer, 2009: 347-360.
|
| [21] |
孙永光, 赵冬至, 吴涛, 卫宝泉, 高树刚, 李媛, 曹芳芳. 河口湿地人为干扰度时空动态及景观响应——以大洋河口为例. 生态学报, 2012, 32(12): 3645-3655. |
| [22] |
陈鹏, 傅世锋, 文超祥, 吴海燕, 宋志晓. 1989-2010年间厦门湾滨海湿地人为干扰影响评价及景观响应. 应用海洋学学报, 2014, 33(2): 167-174. |
| [23] |
Kwan B K Y, Hsieh H L, Cheung S G, Shin P K S. Present population and habitat status of potentially threatened Asian horseshoe crabs Tachypleus tridentatus and Carcinoscorpius rotundicauda in Hong Kong: a proposal for marine protected areas. Biodiversity and Conservation, 2016, 25(4): 673-692. |
| [24] |
颜明艳, 李琼珍, 宋洁, 王振华, 王有基, 胡梦红. 基于MAXENT模型评估北部湾潮间带中国鲎和圆尾鲎稚鲎的潜在地理分布及种群保育对策. 生态学报, 2019, 39(9): 3100-3109. |
| [25] |
Vietnam NBSAP (National Biodiversity Strategies and Action Plans). Vietnam's Fifth International Report to the United Nations Convention on Biological Diversity (Reporting period: 2009-2013). Hanoi: Ministry of Natural Resources and Environment, 2014.
|
| [26] |
Kwan K Y, Wong W T, Lam P Y, Chan H K, Lo H S, Cheung S G. Effects of rubble zones from oyster cultivation on habitat utilization and foraging behaviour of the endangered tri-spine horseshoe crab: an implication for intertidal oyster cultivation practices. Journal of Environmental Management, 2020, 271: 110925. |
| [27] |
Loveland R E, Botton M L. Sea level rise in Delaware Bay, U.S.A. : adaptations of spawning horseshoe crabs (Limulus polyphemus) to the glacial past, and the rapidly changing shoreline of the bay//Carmichael R H, Botton M L, Shin P K S, Cheung S G, eds. Changing Global Perspectives on Horseshoe Crab Biology, Conservation and Management. Cham: Springer, 2015: 41-63.
|
| [28] |
Jawahir A R N, Samsur M, Shabdin M L, Adha A R K. Distribution of two species of Asian horseshoe crabs at west coast of Sarawak's waters, east Malaysia. The Egyptian Journal of Aquatic Research, 2017, 43(2): 135-140. |
| [29] |
Chen C P, Yang M C, Fan L F, Qiu G L, Liao Y Y, Hsieh H L. Co-occurrence of juvenile horseshoe crabs Tachypleus tridentatus and Carcinoscorpius rotundicauda in an estuarine bay, Southwestern China. Aquatic Biology, 2015, 24(2): 117-126. |
| [30] |
Li H Y. The conservation of Horseshoe Crabs in Hong Kong[D]. Hong Kong, China: City University of Hong Kong, 2008.
|
| [31] |
刘春艳, 张科, 刘吉平. 1976-2013年三江平原景观生态风险变化及驱动力. 生态学报, 2018, 38(11): 3729-3740. |
| [32] |
Itow T. Crisis in the Seto Inland Sea: the decimation of the horseshoe crab. EMECS Newsletter, 1993, 3: 10-11. |
| [33] |
Botton M L. The conservation of horseshoe crabs: what can we learn from the Japanese experience?//Tanacredi J T, ed. Limulus in the Limelight: A Species 350 Million Years in the Making and in Peril? New York: Springer, 2001: 41-51.
|
| [34] |
郭清扬, 谷阳光, 鲍虞园, 李银康, 周传江, 颉晓勇. 中华鲎的食物组成及营养位置分析. 南方水产科学, 2021, 17(4): 35-40. |
| [35] |
尚伟平. 草原栖息地退化和破碎化背景下繁殖期栗斑腹鹀肠道微生物群落结构及其影响因素研究[D]. 长春: 吉林农业大学, 2021.
|
| [36] |
Andrén H. Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: a review. Oikos, 1994, 71(3): 355-366. |
| [37] |
张明海, 马建章. 野生动物生境破碎化理论探讨. 野生动物学报, 2014, 35(1): 6-14. |
| [38] |
Fu Y J, Huang S Y, Wu Z, Wang C C, Su M D, Wang X P, Xu P, Huang X, Wu H P, Wang Y J, Wang J Z, Xiao X B, Zhou H C, Xie X Y, Chen M, Huang S L, Liao Y Y, Kwan K Y. Socio-demographic drivers and public perceptions of consumption and conservation of Asian horseshoe crabs in northern Beibu Gulf, China. Aquatic Conservation: Marine and Freshwater Ecosystems, 2019, 29(8): 1268-1277. |
| [39] |
Hu M H, Wang Y J, Chen Y, Cheung S G, Shin P K S, Li Q Z. Summer distribution and abundance of juvenile Chinese horseshoe crabs Tachypleus tridentatus along an intertidal zone in Southern China. Aquatic Biology, 2009, 7(1/2): 107-112. |
| [40] |
刘世梁, 刘芦萌, 武雪, 侯笑云, 赵爽, 刘国华. 区域生态效应研究中人类活动强度定量化评价. 生态学报, 2018, 38(19): 6797-6809. |
| [41] |
解若璠, 沈永明, 劳桦. 基于人为干扰度的盐城滨海湿地景观格局动态变化及响应. 生态学杂志, 2022, 41(2): 351-360. |
| [42] |
潘晨, 周立志, 王晓辉, 徐文彬, 宋昀微. 人类活动对升金湖国家级自然保护区景观格局的影响. 生态科学, 2021, 40(2): 116-124. |
| [43] |
任璘婧, 李秀珍, 李希之, 闫中正, 孙永光. 长江口滩涂湿地景观变化对典型水鸟生境适宜性的影响. 长江流域资源与环境, 2014, 23(10): 1367-1374. |
| [44] |
Koyama A, Hirata T, Kawahara Y, Iyooka H, Kubozono H, Onikura N, Itaya S, Minagawa T. Habitat suitability maps for juvenile tri-spine horseshoe crabs in Japanese intertidal zones: a model approach using unmanned aerial vehicles and the structure from motion technique. PLoS One, 2020, 15(12): e0244494. |
| [45] |
Xie X Y, Wu Z, Wang C C, Fu Y J, Wang X P, Xu P, Huang X, Liao Y Y, Huang S L, Kwan K Y. Nursery habitat for Asian horseshoe crabs along the northern Beibu Gulf, China: implications for conservation management under baseline gaps. Aquatic Conservation: Marine and Freshwater Ecosystems, 2020, 30(2): 260-272. |