 2023, Vol. 43
2023, Vol. 43文章信息
- 饶洁, 段丁琪, 唐强, 马明国, 韦杰, 贺秀斌
- RAO Jie, DUAN Dingqi, TANG Qiang, MA Mingguo, WEI Jie, HE Xiubin
- 三峡水库消落带植被高程梯度分异及其对生境胁迫的响应
- Vegetation differentiation along elevation gradient in the water level fluctuation zone of the Three Gorges Reservoir and its response to habitat stressing
- 生态学报. 2023, 43(16): 6649-6660
- Acta Ecologica Sinica. 2023, 43(16): 6649-6660
- http://dx.doi.org/10.5846/stxb202205061258
-
文章历史
- 收稿日期: 2022-05-06
- 网络出版日期: 2023-02-21

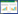




2. 重庆师范大学地理与旅游学院, 三峡库区地表过程与环境遥感重庆市重点实验室, 重庆 401331;
3. 中国科学院、水利部成都山地灾害与环境研究所, 山地表生过程与生态调控重点实验室, 成都 610299
2. Chongqing Key Laboratory of Earth Surface Process and Environment Remote Sensing in the Three Gorge Reservoir Area, School of Geography and Tourism, Chongqing Normal University, Chongqing 401331, China;
3. Key Laboratory of Mountain Surface Processes and Ecological Regulation, Institute of Mountain Hazards and Environment, Chinese Academy of Sciences, Chengdu 610299, China
三峡水库蓄水运行后水位周期性地在145m和175m间波动, 形成了垂直落差达30m、长661km、总面积349km2的水库消落带[1—2]。消落带是水库水位调节在建库河段形成的水位变动区, 是水陆交错带, 坡面水文过程和水库水位节律复合作用形成独特的生态系统格局和过程。消落带是三峡水库重要的结构和功能单元, 承担河岸缓冲带和河流生态廊道的角色, 提供固土护岸、拦沙截污、维持生物多样性等多种生态功能[3—4]。然而, 三峡水库蓄水运行形成的极端生境胁迫导致消落带植被退化严重, 生态系统结构和功能严重受损。一方面, 高压极端淹水胁迫导致生境破碎化加剧, 原有陆生植被快速消亡, 适生性植被发生渐进演替, 消落带植被格局显著改变, 生物多样性降低[5—6];另一方面, 夏季干旱、土壤侵蚀、泥沙掩埋、土壤贫瘠化等因素导致消落带生境空间异质性增加, 生态系统承载力降低, 成为植被演替主要障碍性因素[7—8]。当前在库区或样地尺度, 开展了消落带植被群落结构时空分布[5]、岸坡土壤种子库[9]等调查;从生长繁殖适应策略[10]、C\N\P生态化学计量特征[11]、植株形态和功能性状[12]、光合生理特征[13]等视角剖析了适生物种对极端淹水、泥沙掩埋等生境胁迫的适应机理或策略。然而, 缺乏消落带植被演替进程对多因子复合生境胁迫响应的综合理解。
水库消落带植被退化与生态功能受损是长江流域水电开发面临的特色问题, 开展消落带生态修复, 提升河流受损廊道的综合生态功能, 为服务长江流域生态保护国家战略和保障长江流域水资源安全具有重要意义。植被作为水库消落带生态系统重要的结构和功能单元, 恢复受损植被是消落带生态功能重建的重要途径[14—15]。然而, 当前三峡水库消落带植被恢复缺少极端生境胁迫下植被演替过程机制理论体系支撑。生物量能够衡量消落带不同植被群落生产力水平高低, 定量反映植被生长差异, 对表征消落带生态系统演替具有极强的指示作用。本研究选取三峡水库典型自然恢复消落带为研究对象, 聚焦高程梯度上植被的空间再分配格局, 拟结合野外调查和室内分析, 从极端水淹胁迫、土壤侵蚀—泥沙沉积、土壤基质环境状况三个层次探讨生境因子对植被高程梯度分异的影响, 揭示消落带植被演替与特殊生境条件的关联特性, 对深入认识极端生境胁迫下植被演替进程和消落带土壤—植被互馈效应, 开展消落带植被恢复和生态功能重建具有重要意义。
1 研究区概况三峡工程在坝址宜昌至库尾江津间形成了长661km、水域面积1080km2水库型河段(图 1), 其中涪陵以上为变动回水区, 典型特征是冬季为雍水河段, 夏季为自然流河段, 涪陵以下为永久水库区。三峡库区奉节以东为喀斯特岩溶区, 地貌以高山峡谷为主, 消落带坡度陡, 基岩出露;奉节以西为低山丘陵区, 地势相对平缓, 河谷开阔, 消落带分布面积广, 土地资源优良, 是开展消落带生态修复的重点区域。本研究选择三峡水库中游重庆市忠县石宝镇长江干流左岸土质消落带为研究区。该区属亚热带湿润性季风气候, 年均气温18.2℃, 年均降水量1172.1mm, 降水集中在6—8月, 占全年的80%以上, 相对湿度81%[16];土壤为紫色土, pH值为7.21左右, 土质结构松散, 抗蚀性能低下。蓄水前消落带土地利用主要为水田和旱地, 蓄水后以自然演替的草本植物为主, 代表性优势物种如狗牙根、苍耳等。

|
| 图 1 三峡水库范围和研究区位置 Fig. 1 Map of the Three Gorges Reservoir and location of the study area |
三峡水库独特的“蓄清排浊”运行模式形成了特定的水库水位节律。在单个运行周期内水位在145m和175m间波动:即每年9月中旬起, 水库开启蓄水周期以满足发电需求, 至10月底水位逐渐上涨至175m最高水位;11月至次年1月, 水位维持在175m左右运行;2月至6月, 水库开始腾空库容, 水位从175m逐渐下降至145m;6月至9月, 水位维持在145m左右运行以满足长江防洪排沙需求, 受雨季长江上游洪峰影响, 呈现不同规模的自然洪峰脉冲(图 2)。三峡水库消落带成陆期与库区夏季光热水资源同期, 且土地资源优良, 为植被生长提供有利生境条件。消落带植被利用夏季出露空窗期捕获光热水资源, 快速完成自身生活史。1月至3月, 植被降低对水淹环境的敏感性, 依靠萌发过程占据土壤辍块[17];4月至7月, 最优化资源适配为植被生长提供物质保障, 加快生产代谢活动, 促进物质积累[18];8月至9月, 植被水平空间资源利用率饱和, 群落构成趋于成熟。然而, 在消落带高程梯度上, 水位节律形成的淹水时长、出露时令差异导致植被物候出现迟滞效应, 对群落结构和生产力产生重要影响。

|
| 图 2 三峡水库水位变动与消落带植被生命周期 Fig. 2 Changes of water level of the Three Gorges Reservoir and life cycle of vegetation in the water level fluctuation zone |
野外调查和样品采集于2021年8月完成。此时植被处于成熟期, 植被群落结构和生物量趋于稳定。经实地踏勘, 结合坡度、高程以及地表覆盖土壤质地类型和颜色差异, 沿高程自低向高划分为(Ⅰ)侵蚀区、(Ⅱ)沉积区、(Ⅲ)沉积区和(Ⅳ)侵蚀区。沿145—175m高程间隔一定距离设置三条代表性样带(T1、T2、T3), 并依据高程梯度、植被优势群落分布、侵蚀和沉积情况, 沿每条样带布设1m×1m的植被调查样方, 共设置调查样方16个。现场记录各样方内植物种名、盖度、高度及生活型。齐地刈割1/4(0.25m2)样方采集植被地上生物量, 称定鲜重后装入牛皮袋并编号。土壤样品采集在清除表层枯落物和腐殖质层后, 根据“梅花五点法”使用土钻钻取0—10cm, 10—20cm表土混合样, 装入聚乙烯自封袋并编号, 共采集32份土样;使用环刀在样方中央采集原状土样, 现场称定鲜重后装入自封袋编号。
2.2 样品测试植物样品带回实验室, 经85℃烘干48h至恒重后称干重代表植被生物量。土壤样品置于阴凉处自然风干, 剔除石块、植物根系等杂物后用玛瑙钵手工研磨, 过2mm尼龙筛后部分样品用于土壤pH、速效氮、速效磷的测定, 剩余样品过0.149mm尼龙筛后用于土壤有机质、全氮、全磷的测定。原状土经105℃烘干24h后称干重, 计算鲜重与干重的差值占鲜重的重量百分比作为土壤含水率[19];pH值采用pH电位法测定[19];有机碳和全氮采用元素分析仪测定[20];速效氮采用碱解—扩散法测定[19];全磷采用硫酸—高氯酸—钼锑抗比色法测定[19];速效磷采用碳酸氢钠提取—钼锑抗比色法测定[19]。
2.3 数据处理 2.3.1 水文变量三峡水库逐日水位数据收集自长江水文网。利用三峡水库逐日水位数据计算单个运行周期内消落带不同高程的平均淹水深度(Mean Inundation Depth, MID)和累积淹水时长(Cumulative Inundation Duration, CID), 计算方法如下。
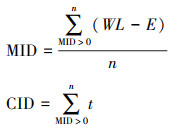
|
式中, WL为三峡水库逐日水位;E为消落带145—175m高程;t为MID>0的天数;n为总天数。
2.3.2 物种多样性指数选取重要值(Important value, IV)评价群落中植被物种相对重要性;物种多样性指数采用Simpson优势度指数、Shannon-Weiner指数、Pielou均匀度指数和Patrick丰富度指数作为评价指标, 用以表征群落内植物种类优势地位、复杂程度、物种分布均匀程度以及物种密度的特征[21], 分别采用以下公式计算:
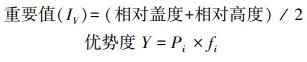
|
Shannon-Weiner多样性指数 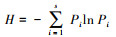
Pielou均匀度指数 E=H/lnS
Patrick丰富度指数 R=S
式中, Pi=Ni/N;Ni为某物种重要值;N为样方内所有物种重要值之和;S为样方内的总物种数。
3 结果与分析 3.1 植被群落组成和物种多样性的高程梯度分异特征对研究消落带野外调查共发现草本植物15科25种23属, 其中禾本科(Gramineae)6种6属, 菊科(Compositae)3种3属, 蓼科(Polygonaceae)3种2属, 分别占物种总数量的24%、12%和12%, 是优势科;其它科植物单种、单属现象明显。整体而言, 一年生草本占物种总数量的52%, 多年生草本占48%, 表明消落带优势生活型存在空间支配主导性。在消落带高程梯度上, 多年生草本在低高程段呈现优势性, 而随高程升高, 多年生草本植物占比降低, 一年生草本植物占比增加。通过对物种多样性指数沿高程进行线性回归, 发现消落带植被物种Shannon-Weiner多样性指数、Pielou均匀度指数以及Patrick丰富度指数随海拔升高均呈增加趋势(图 3), 拟合系数r分别为0.804、0.791和0.586, 且Shannon-Weiner多样性指数高程之间差异显著(P < 0.05), 表明低高程段植被结构趋于单一化。

|
| 图 3 消落带高程梯度植物多样性指数和生活型变化 Fig. 3 Variability of species diversity index and vegetation lifestyle with elevations |
消落带植被优势物种沿高程梯度呈现空间差异性。随海拔升高, 形成以“狗牙根(Cynodon dactylon)-酸模叶蓼(Polygonum lapathifolium)+狗牙根-狗牙根-苍耳(Xanthium sibiricum)”为优势群落的总体分布格局(表 1)。消落带上部主要聚集苍耳等高大直立一年生草本(优势度为0.13±0.02);消落带中部为过渡带, 优势物种多样化分布, 易形成共优群落来维持群落稳定性, 其中狗牙根(0.28±0.09)和酸模叶蓼优势度较高(0.13±0.04);消落带下部主要聚集耐淹性强的狗牙根等多年生草本(优势度为0.44±0.09)。同一优势物种在不同高程优势性具有差异, 狗牙根作为研究消落带断面优势种, 形成消落带下部(0.44±0.09)>消落带中部(0.29±0.09)>消落带上部(0.12±0.11)的分布格局, 越靠近消落带下缘狗牙根聚集度越高。
| 种 Species |
科 Families |
属 Genera |
优势度Dominant value | |||||
| 145—150 | 150—155 | 155—160 | 160—165 | 165—170 | 170—175 | |||
| 狗牙根 Cynodon dactylon |
禾本科 Gramineae |
狗牙根属 Cynodon |
0.376 | 0.502 | 0.196 | 0.376 | 0.233 | 0.116 |
| 苍耳 Xanthium sibiricum |
菊科 Compositae |
苍耳属 Xanthium |
— | — | 0.014 | 0.01 | 0.031 | 0.128 |
| 酸模叶蓼 Polygonum lapathifolium |
蓼科 Polygonaceae |
蓼属 Polygonum |
— | — | 0.176 | 0.073 | — | 0.043 |
| 葎草 Humulus scandens |
桑科 Moraceae |
葎草属 Humulus |
— | — | — | — | — | 0.032 |
| 香附子 Cyperus rotundus |
莎草科 Cyperaceae |
莎草属 Cyperus |
0.017 | 0.01 | — | — | — | 0.011 |
| 长鬃蓼 Polygonum longisetum |
蓼科 Polygonaceae |
蓼属 Polygonum |
— | — | — | — | — | 0.002 |
| 牛鞭草 Hemarthria sibirica |
禾本科 Gramineae |
牛鞭草属 Hemarthria |
— | 0.003 | 0.015 | 0.033 | 0.081 | 0.044 |
| 石荠苎 Mosla scabra |
唇形科 Lamiaceae |
石荠苎属 Mosla |
— | — | — | 0.023 | 0.017 | 0.01 |
| 狗尾草 Setaria viridis |
禾本科 Gramineae |
狗尾草属 Setaria |
— | — | — | 0.008 | — | 0.004 |
| 合萌 Aeschynomene indica |
豆科 Leguminosae |
合萌属 Aeschynomene |
— | — | — | — | 0.004 | — |
| 莲子草 Alternanthera sessilis |
苋科 Amaranthaceae |
莲子草属 Alternanthera |
— | — | — | 0.009 | — | — |
| 喜旱莲子草 Alternanthera philoxeroides |
苋科 Amaranthaceae |
莲子草属 Alternanthera |
— | 0.018 | 0.03 | 0.017 | — | — |
| 鳢肠 Eclipta prostrata |
菊科 Compositae |
鳢肠属 Eclipta |
— | — | 0.006 | 0.005 | — | — |
| 稗 Echinochloa crus-galli |
禾本科 Gramineae |
稗属 Echinochloa |
— | — | 0.004 | 0.003 | — | — |
| 鬼针草 Bidens pilosa |
菊科 Compositae |
鬼针草属 Bidens |
— | — | 0.007 | — | — | — |
| 马唐 Digitaria sanguinalis |
禾本科 Gramineae |
马唐属 Digitaria |
— | — | 0.007 | — | — | — |
| 大青 Clerodendrum cyrtophyllum |
马鞭草科 Verbenaceae |
大青属 Clerodendrum |
— | — | 0.003 | — | — | — |
| 短叶水蜈蚣 Kyllinga brevifolia |
莎草科 Cyperaceae |
水蜈蚣属 Kyllinga |
0.006 | 0.006 | — | — | — | — |
| 水苦荬 Veronica undulata |
玄参科 Scrophulariaceae |
婆婆纳属 Veronica |
— | 0.002 | — | — | — | — |
| 灯芯草 Juncus effusus |
灯芯草科 Juncaceae |
灯芯草属 Juncus |
— | 0.003 | — | — | — | — |
| 酸模 Rumex acetosa |
蓼科 Polygonaceae |
酸模属 Rumex |
— | 0.005 | — | — | — | — |
| 蔊菜 Rorippa indica |
十字花科 Brassicaceae |
蔊菜属 Rorippa |
— | 0.002 | — | — | — | — |
| 水蕨 Ceratopteris thalictroides |
水蕨科 Parkeriaceae |
水蕨属 Ceratopteris |
— | 0.002 | — | — | — | — |
| 扯根菜 Penthorum chinense |
虎耳草科 Saxifragaceae |
扯根菜属 Penthorum |
0.006 | — | — | — | — | — |
研究消落带植被总生物量变化范围为199.68—1211.2g/m2。通过对植被生物量沿高程进行线性回归(图 4), 发现植被生物量沿高程梯度自低向高呈显著增加趋势(P < 0.05)。但植被生物量随高程变化存在波动点, 总体呈现“W”型的分布格局。对侵蚀和沉积区植被生物量的分析发现, Ⅳ侵蚀区(736.13g/m2)>Ⅲ沉积区(684.5g/m2)>Ⅰ侵蚀区(405.68g/m2)>Ⅱ沉积区(302.16g/m2), 且差异显著(P < 0.05), 如图 5。

|
| 图 4 消落带植被生物量和水文变量随高程变化 Fig. 4 Changes of vegetation biomass and hydrological variables along with elevation gradient |

|
| 图 5 植被生物量在侵蚀—沉积区域的分异 Fig. 5 Vegetation biomass divergence in different erosion-deposition zones |
总体来说, 土壤理化性质差异较大, 空间异质性较强(表 2)。除全磷外, 其余土壤因子变异系数均大于15%, 其中速效磷变异系数高达49.22%, 其次为速效氮和土壤含水率。0—10cm土壤含水率、有机质、全氮、速效氮、全磷、速效磷均值分别为43.38%、17.99g/kg、1.07g/kg、112.35mg/kg、0.41g/kg、15.69mg/kg;而10—20cm土壤含水率、有机质、全氮、速效氮、全磷、速效磷均值分别为42%、17.54g/kg、1.04g/kg、112.19mg/kg、0.41g/kg、14.99mg/kg, 表明在土壤剖面上, 土壤养分随土壤深度增加而减少, 但差异性不显著(P>0.05)。
| 因子 Factors |
土层 Soil layer |
最小值 Minimum |
最大值 Maximum |
平均数 Mean |
标准误差 Standard deviation |
变异系数 Coefficient of variation |
| 含水率 | 0—10cm | 0.25 | 0.52 | 0.43 | 0.07 | 16.62 |
| Soil water content | 10—20cm | 0.17 | 0.55 | 0.42 | 0.11 | 27.61 |
| 有机质 | 0—10cm | 14.85 | 20.97 | 17.99 | 1.47 | 8.18 |
| Organic matter | 10—20cm | 10.95 | 24.01 | 17.54 | 4.04 | 23.05 |
| 全氮 | 0—10cm | 0.92 | 1.25 | 1.07 | 0.08 | 7.89 |
| Total nitrogen | 10—20cm | 0.63 | 1.39 | 1.04 | 0.21 | 20.08 |
| 速效氮 | 0—10cm | 59.85 | 168.35 | 112.35 | 35.01 | 31.16 |
| Available nitrogen | 10—20cm | 65.1 | 208.6 | 112.19 | 47.85 | 42.65 |
| 全磷 | 0—10cm | 0.36 | 0.47 | 0.41 | 0.03 | 0.08 |
| Total phosphorus | 10—20cm | 0.36 | 0.46 | 0.41 | 0.03 | 0.07 |
| 速效磷 | 0—10cm | 6.75 | 27.85 | 15.69 | 5.97 | 0.38 |
| Available phosphorus | 10—20cm | 7.65 | 23.85 | 14.99 | 6.45 | 0.43 |
相关性分析表明, 消落带植被生物量与累积淹水时长(R=-0.677, P < 0.01)、平均淹水深度(R=-0.691, P < 0.01)呈极显著负相关, 与土壤含水率(R=0.52, P < 0.05)、土壤全氮(R=0.52, P < 0.05)呈显著正相关(图 6)。主成分分析表明, KMO检验系数超过0.5, Bartlett检验值小于指定显著水平0.05;前2个PCA轴累积解释方差比例达69.15%, 且特征根都大于1, 表明土壤理化性质在前两个排序轴载荷值较高(图 7)。主成分1贡献率为51.29%, 累积淹水深度、平均淹水时长和土壤含水率权重系数较大, 绝对值均超过了0.8, 主要反映水淹胁迫的影响;主成分2的贡献率为17.86%, 全氮和速效氮权重系数较大, 反映土壤养分的影响。

|
| 图 6 植被生物量与生境因子相关性 Fig. 6 The correlationship between riparian vegetation biomass and environmental factors **代表在0.01水平上差异显著;*代表在0.05水平上差异显著 |

|
| 图 7 前2个主成分载荷图 Fig. 7 The payloads of first two principal components |
三峡水库水位周期性波动改变河流自然洪枯规律并形成特定的水库水位节律, 导致淹水累计时长、出露时令、淹水强度等指标在消落带高程梯度上存在明显差异性。受制于高强度淹水胁迫, 消落带植被群落结构处于剧烈退化阶段, 原生植被迁移、变异或消亡, 适生植被出现渐进演替, 植被物种多样性锐减并显著低于自然河岸带[5, 22—24]。植被多样性指数与高程呈正相关性, 可能与资源条件分化和植被生态适应差异相关[6, 25]。消落带下部水淹胁迫强, 植被定殖受阻, 不耐淹植被由于缺乏适应极端环境的机体结构和功能而消亡, 群落构成单一化;而消落带上部微生境条件复杂, 资源组合最优, 有利于生态位宽度较大的植被物种定殖生长, 群落物种多样性丰富[18, 26]。依托种子繁殖和杂草策略生长的一年生草本具有生活周期短、适应性强的特点, 能建立异质生境耐受性以规避水淹干扰, 在植被筛选驯化过程中逐步取代多年生草本成为消落带优势生活型[6, 22, 27]。然而在高强度、长时间水淹的消落带下部, 一年生草本种子活性丧失成苗率低, 难以完成自身生活史;而多年生草本具有极强的再生能力和发达的通气组织, 依靠根系克隆整合快速恢复与扩张[28—29], 在消落带下部易形成斑块化分布格局。因此不同生活型物种沿消落带高程梯度具有相反演化分布格局[30], 即随高程降低, 一年生占比降低, 多年生占比升高。
4.2 多因子复合生境胁迫对消落带植被生物量的影响三峡水库消落带季节性淹没—出露形成特定的宏观生境, 然而土壤侵蚀、泥沙沉积、土壤基质环境变化导致消落带生境破碎化增强, 生态系统脆弱性和敏感性增加。水位周期性涨落过程中, 水体掏蚀和泥沙沉积破坏库岸土壤结构稳定, 引起土壤养分流失, 造成植被生长基质失稳[14]。水位周期性涨落形成的累积淹水时长、平均淹水深度等是决定植被生物量高程梯度分异的首要胁迫因子, 与植被生物量呈极显著负相关性[5, 31], 即随高程增加, 植被承受的累积淹水时长、平均淹水深度越小(图 4), 而生物量积累越大, 这可能与植被塑型效应和生理特征调节差异有关[32—33]。消落带下部高强度、长时间水淹和出露时令晚等特点, 植被光合作用和物质代谢过程受阻, 通过减少种群密度并采取高生殖分配对策降低能量消耗, 减缓植株生长[34];而消落带上部植被利用消落带成陆期充足的光热水资源激励种子萌发, 增加叶生物量分配进行营养生长, 并积累碳水化合物促进地上部分再生产过程[10, 35], 形成植被生物量随高程增加而升高的趋势。
然而, 植被生物量沿高程变化存在波动值[36—37](图 4), 反映侵蚀—沉积过程以及土壤基质环境的再分配作用。侵蚀—沉积过程通过改变土层厚度、表层土壤肥力和基质环境等间接影响上覆植被生长发育过程。土壤侵蚀导致原始剖面表层土壤颗粒流失, 养分淋溶损耗等贫瘠化效应;并引发土壤团粒结构、总孔隙度和毛管孔隙度等物理特性恶化, 削弱了土壤持水、抗蚀和抗旱能力, 对植被种子萌发、建植和根叶生长产生消极影响[38]。水库水体悬移质泥沙在消落带沉积, 对植被生长产生掩埋效应。适度泥埋引起植被“补偿生长”现象, 促进根叶协同发展, 有利于土壤肥力和水分吸收;而过度泥沙掩埋影响土壤通透性, 抑制植被出苗、地上分株形成, 降低光合作用效力, 造成植被生长差异[39—41]。研究发现, Ⅱ沉积区植被生物量呈现低值, 可能是净泥沙沉积量超过植被特有忍耐限度, 植被出芽和生长发育过程受到抑制作用。因此, 侵蚀和沉积过程是影响植被生长的次要胁迫因子。
土壤作为消落带物质能量循环重要一环, 为消落带植被生长协调和供应营养物质, 决定植被群落生产力状况。水位周期性涨落导致大粒径团聚体崩解为微团聚体, 加速土壤养分的释放、运动和扩散, 造成消落带土壤贫瘠状态[42—43]。植被生物量与土壤含水率、全氮呈显著正相关[44—45], 表明其是决定植被生物量高程梯度分异的主要土壤限制性因子。高浓度全氮能刺激种子萌发[46];促进根系吸收以维持植株N化学计量平衡;促进植被叶绿素合成以增加生态系统生产力, 因此土壤全氮与植被生物量变化步调一致。而土壤含水率作为能量循环的重要载体通过影响养分在土壤中的转化和运移, 决定植被对土壤水分和养分利用效率, 有利于生物量积累。
综上, 沿消落带高程梯度的水淹、侵蚀—沉积、土壤基质环境三类生境胁迫要素对消落带植被萌芽、生长和成熟三阶段产生不同作用效应, 造成物种多样性、植被生物量呈现显著高程梯度分异特征(图 8)。但本研究仅考虑单时段内, 植被对三类生境胁迫要素的响应, 缺乏动态性和综合性。后续研究可从多时段、多因素角度定量描述消落带植被筛选驯化过程和时空分异特征, 探讨其对多因子极端生境胁迫的响应。

|
| 图 8 环境驱动力对植被演替的影响 Fig. 8 The impacts of habitat drivers on vegetation succession |
三峡水库消落带植被退化和生态功能下降是以强烈的人为干扰为外因, 以多因素复合生境胁迫驱动, 以植被群落构成、物种多样性和生物量减少为本质, 以高程梯度植被格局分异为标志的生态系统适应性演替过程。植被作为消落带综合生态功能重建的主体, 优势生境适宜性物种筛选是开展水库消落带植被生态恢复的关键。三峡水库消落带植被群落演替过程尚未稳定, 亟需开拓具有耐淹、耐旱适应性, 根系发达、固岸能力强和高经济价值的先锋物种构建强自稳定性的植被群落[47]。消落带植被恢复需重视土壤基质保育, 采取必要的调水保土措施保持土壤、调节土壤水分、维持土壤肥力, 达到稳固岸坡和维持土地生产潜力目的。结合植被生态型特征和立地条件高程差异, 因地制宜, 分区优化物种配置[48]。在保障消落带自然恢复过程中, 适当辅以人工措施, 采取林草混交模式进行合理的物种组配, 使物种组成分布渐趋均匀, 满足生态系统稳定、不同区位生态服务功能提供、景观美学的需求。
5 结论本研究选取三峡水库典型自然恢复消落带, 联合野外调查和室内分析, 揭示了消落带植被的高程梯度分异特征, 系统分析了极端水淹、侵蚀—沉积、土壤理化性质等生境胁迫类型对消落带植被高程梯度分异的影响, 提出了消落带植被恢复的重要方向, 取得如下主要结论:
(1) 消落带在植被物种多样性、群落组成和生物量三方面呈现明显的高程梯度分异规律。研究消落带植被以适生草本为主, 单种单属、单优群落现象明显, 物种多样性指数与高程呈正相关关系;一年生草本与多年生草本成为消落带优势生活型, 植被群落结构趋于单一化, 随海拔增加一年生草本占比增加, 多年生占比减少;植被生物量沿高程梯度自低向高呈显著递增趋势, 但存在局部波动。
(2) 水淹、侵蚀—沉积、土壤理化性质三类生境胁迫类型是影响消落带植被生物量高程梯度分异的主要因素, 具有层次性效应。三峡水库水位季节性波动形成的淹没时长、出露时令、淹水强度等是决定植被生物量高程梯度分异的首要胁迫因子, 对植被生长产生极显著负效应;侵蚀—沉积过程作为次要胁迫因子, 改变土层厚度、土壤持水性能和肥力状况, 对植被生长产生土壤贫瘠化效应或泥沙掩埋效应;土壤含水率、土壤全氮作为植被生长主要限制性因子, 提供植被生理过程必需的水分和养分, 形成显著正相关效应。
(3) 三峡水库消落带植被恢复的重点任务是针对极端淹水、夏季间歇性干旱、土壤贫瘠化和泥沙掩埋等生境胁迫的优质抗逆物种的选育, 其次是土壤基质保育, 而植被格局优化配置有助于提升植被的综合生态效益, 规避过度恢复带来的负面环境效应。
| [1] |
Wu J G, Huang J H, Han X G, Xie Z Q, Gao X M. Three-gorges Dam—experiment in habitat fragmentation?. Science, 2003, 300(5623): 1239-1240. DOI:10.1126/science.1083312 |
| [2] |
Fu B J, Wu B F, Lü Y H, Xu Z H, Cao J H, Niu D, Yang G S, Zhou Y M. Three Gorges Project: efforts and challenges for the environment. Progress in Physical Geography: Earth and Environment, 2010, 34(6): 741-754. DOI:10.1177/0309133310370286 |
| [3] |
New T, Xie Z Q. Impacts of large dams on riparian vegetation: applying global experience to the case of China′s Three Gorges Dam. Biodiversity and Conservation, 2008, 17(13): 3149-3163. DOI:10.1007/s10531-008-9416-2 |
| [4] |
Yang F, Wang Y, Chan Z L. Review of environmental conditions in the water level fluctuation zone: perspectives on riparian vegetation engineering in the Three Gorges Reservoir. Aquatic Ecosystem Health & Management, 2015, 18(2): 240-249. |
| [5] |
朱妮妮, 郭泉水, 秦爱丽, 裴顺祥, 马凡强, 朱莉, 简尊吉. 三峡水库奉节以东秭归和巫山段消落带植物群落动态特征. 生态学报, 2015, 35(23): 7852-7867. |
| [6] |
张爱英, 熊高明, 樊大勇, 谢宗强. 三峡水库运行对淹没区及消落带植物多样性的影响. 生态学杂志, 2016, 35(9): 2505-2518. |
| [7] |
李强. 泥沙掩埋和干旱对三峡库区消落带狗牙根生长恢复的影响. 生态学报, 2016, 36(1): 200-208. |
| [8] |
童笑笑, 陈春娣, 吴胜军, 贾振毅, 易雪梅, 马茂华. 三峡库区澎溪河消落带植物群落分布格局及生境影响. 生态学报, 2018, 38(2): 571-580. |
| [9] |
Zhang A Y, Xie Z Q. C4 herbs dominate the reservoir flood area of the Three Gorges Reservoir. Science of the Total Environment, 2021, 755: 142479. DOI:10.1016/j.scitotenv.2020.142479 |
| [10] |
姚洁, 曾波, 杜珲, 潘晓娇, 苏晓磊. 三峡水库长期水淹条件下耐淹植物甜根子草的资源分配特征. 生态学报, 2015, 35(22): 7347-7354. |
| [11] |
孔维苇, 王晓锋, 卢虹宇, 刘婷婷, 龚小杰, 刘欢, 袁兴中. 三峡库区消落带4种典型草本植物的生态化学计量特征. 生态学报, 2020, 40(13): 4493-4506. |
| [12] |
Su X L, Bejarano M D, Yi X M, Lin F, Ayi Q L, Zeng B. Unnatural flooding alters the functional diversity of riparian vegetation of the Three Gorges Reservoir. Freshwater Biology, 2020, 65(9): 1585-1595. DOI:10.1111/fwb.13523 |
| [13] |
冯大兰, 刘芸, 钟章成, 杨娟, 谢君. 三峡库区消落带芦苇(Phragmites communis(reed))的光合生理响应和叶绿素荧光特性. 生态学报, 2008, 28(5): 2013-2021. |
| [14] |
程瑞梅, 王晓荣, 肖文发, 郭泉水. 消落带研究进展. 林业科学, 2010, 46(4): 111-119. |
| [15] |
Gurnell A. Plants as river system engineers. Earth Surface Processes and Landforms, 2014, 39(1): 4-25. DOI:10.1002/esp.3397 |
| [16] |
储立民, 常超, 谢宗强, 熊高明. 三峡水库蓄水对消落带土壤重金属的影响. 土壤学报, 2011, 48(1): 192-196. |
| [17] |
刘瑞雪, 詹娟, 史志华, 陈龙清. 丹江口水库消落带土壤种子库与地上植被和环境的关系. 应用生态学报, 2013, 24(3): 801-808. |
| [18] |
简尊吉, 马凡强, 郭泉水, 秦爱丽, 肖文发. 三峡水库峡谷地貌区消落带优势植物种群生态位. 生态学杂志, 2017, 36(2): 328-334. |
| [19] |
曹恒生, 唐荣华, 朱泉娣, 中国土壤学会农业化学专业委员会. 土壤农业化学常规分析方法. 北京: 科学出版社, 1983: 457.
|
| [20] |
王巧环, 任玉芬, 孟龄, 李虹, 傅慧敏, 王华锋. 元素分析仪同时测定土壤中全氮和有机碳. 分析试验室, 2013, 32(10): 41-45. |
| [21] |
郭燕, 杨邵, 沈雅飞, 肖文发, 程瑞梅. 三峡水库消落带现存植物自然分布特征与群落物种多样性研究. 生态学报, 2019, 39(12): 4255-4265. |
| [22] |
Grime J P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. The American Naturalist, 1977, 111(982): 1169-1194. |
| [23] |
Nilsson C, Jansson R. Floristic differences between riparian corridors of regulated and free-flowing boreal rivers. Regulated Rivers: Research & Management, 1995, 11(1): 55-66. |
| [24] |
王强, 袁兴中, 刘红, 张跃伟, 陈忠礼, 李波. 三峡水库初期蓄水对消落带植被及物种多样性的影响. 自然资源学报, 2011, 26(10): 1680-1693. |
| [25] |
Nilsson C, Jansson R, Zinko U. Long-term responses of river-margin vegetation to water-level regulation. Science, 1997, 276(5313): 798-800. |
| [26] |
郭燕, 杨邵, 沈雅飞, 肖文发, 程瑞梅. 三峡库区消落带现存草本植物组成与生态位. 应用生态学报, 2018, 29(11): 3559-3568. |
| [27] |
王业春, 雷波, 张晟. 三峡库区消落带不同水位高程植被和土壤特征差异. 湖泊科学, 2012, 24(2): 206-212. |
| [28] |
张想英, 樊大勇, 谢宗强, 熊高明, 李兆佳. 克隆整合有助于狗牙根抵御水淹. 植物生态学报, 2010, 34(9): 1075-1083. |
| [29] |
洪明, 郭泉水, 聂必红, 康义, 裴顺祥, 金江群, 王祥福. 三峡库区消落带狗牙根种群对水陆生境变化的响应. 应用生态学报, 2011, 22(11): 2829-2835. |
| [30] |
苏琴琴, 俞幸池, 覃红玲, 谢姣姣, 荆书芳, 林锋, 苏晓磊, 阿依巧丽, 曾波, 张小萍. 三峡水库消落区不同生活史类型植物群落的空间分布格局. 生态学报, 2020, 40(13): 4507-4515. |
| [31] |
Wang Q, Yuan X Z, Willison J H M, Zhang Y W, Liu H. Diversity and above-ground biomass patterns of vascular flora induced by flooding in the drawdown area of China′s Three Gorges Reservoir. PLoS One, 2014, 9(6): e100889. |
| [32] |
Zhu Z H, Chen Z L, Li L, Shao Y. Response of dominant plant species to periodic flooding in the riparian zone of the Three Gorges Reservoir (TGR), China. Science of the Total Environment, 2020, 747: 141101. |
| [33] |
Wei G W, Chen Y H, Sun X S, Matsubara S, Luo F L, Yu F H. Elevation-dependent selection for plasticity in leaf and root traits of Polygonum hydropiper in response to flooding. Environmental and Experimental Botany, 2021, 182: 104331. |
| [34] |
李彦杰, 刘仁华, 周大祥, 杨俊年, 蒋梦芸, 吴巍, 郭世浩, 高鑫. 三峡库区消落带野生狗牙根对水淹适生性的转录组分析. 生态学报, 2018, 38(23): 8434-8441. |
| [35] |
李强, 丁武泉, 王书敏, 朱启红, 杨俊, 柯胜钱, 秦露, 杨林静, 郑捷月, 孟燚雯. 三峡库区多年高水位运行对消落带狗牙根生长恢复的影响. 生态学报, 2020, 40(3): 985-992. |
| [36] |
Zhang Z Y, Wan C Y, Zheng Z W, Hu L, Feng K, Chang J B, Xie P. Plant community characteristics and their responses to environmental factors in the water level fluctuation zone of the Three Gorges Reservoir in China. Environmental Science and Pollution Research, 2013, 20(10): 7080-7091. |
| [37] |
Chen Z L, Yuan X Z, Roß-Nickoll M, Hollert H, Schaeffer A. Moderate inundation stimulates plant community assembly in the drawdown zone of China′s Three Gorges Reservoir. Environmental Sciences Europe, 2020, 32(1): 79. |
| [38] |
焦菊英, 王宁, 杜华栋, 王东丽. 土壤侵蚀对植被发育演替的干扰与植物的抗侵蚀特性研究进展. 草业学报, 2012, 21(5): 311-318. |
| [39] |
潘瑛, 谢永宏, 陈心胜, 李峰. 湿地植物对泥沙淤积的适应. 生态学杂志, 2011, 30(1): 155-161. |
| [40] |
Tang Q, Bao Y H, He X B, Zhou H D, Cao Z J, Gao P, Zhong R H, Hu Y H, Zhang X B. Sedimentation and associated trace metal enrichment in the riparian zone of the Three Gorges Reservoir, China. Science of the Total Environment, 2014, 479/480: 258-266. |
| [41] |
李强, 丁武泉, 朱启红, 宋力. 模拟沙埋对三峡库区低位消落带狗牙根恢复生长的影响. 生态学杂志, 2015, 34(4): 919-924. |
| [42] |
程瑞梅, 刘泽彬, 肖文发, 王娜, 王晓荣, 沈雅飞. 三峡库区典型消落带土壤化学性质变化. 林业科学, 2017, 53(2): 19-25. |
| [43] |
张淑娟, 贺秀斌, 鲍玉海, 唐强. 三峡水库消落带不同水位高程土壤团聚体变化特征. 水土保持研究, 2021, 28(1): 25-30. |
| [44] |
张志永, 万成炎, 郑志伟, 邹曦, 潘晓洁, 胡莲, 冯坤, 陈小娟. 三峡水库小江消落区生境异质性对植物群落影响. 长江流域资源与环境, 2013, 22(11): 1506-1513. |
| [45] |
徐建霞, 王建柱. 三峡库区香溪河消落带植被群落特征与土壤环境相关性. 生态学杂志, 2018, 37(12): 3661-3669. |
| [46] |
Li T T, Zhu Z H, Shao Y, Chen Z L, Roß-Nickoll M. Soil seedbank: importance for revegetation in the water level fluctuation zone of the reservoir area. Science of the Total Environment, 2022, 829: 154686. |
| [47] |
樊大勇, 熊高明, 张爱英, 刘曦, 谢宗强, 李兆佳. 三峡库区水位调度对消落带生态修复中物种筛选实践的影响. 植物生态学报, 2015, 39(4): 416-432. |
| [48] |
贺秀斌, 鲍玉海. 三峡水库消落带土壤侵蚀与生态重建研究进展. 中国水土保持科学, 2019, 17(4): 160-168. |