 2023, Vol. 43
2023, Vol. 43文章信息
- 李琪琪, 黄小娟, 李岚, 常生华, 侯扶江
- LI Qiqi, HUANG Xiaojuan, LI Lan, CHANG Shenghua, HOU Fujiang
- 黄土高原典型草原群落结构和土壤水分对划破的响应
- Responses of plant community and soil moisture to ripping intensity in the typical steppe of the Loess Plateau of China
- 生态学报. 2023, 43(15): 6131-6142
- Acta Ecologica Sinica. 2023, 43(15): 6131-6142
- http://dx.doi.org/10.5846/stxb202301040026
-
文章历史
- 收稿日期: 2023-01-04
- 网络出版日期: 2023-05-26






2. 草种创新与草地农业生态系统全国重点实验室,兰州 730020;
3. 农业农村部草牧业创新重点实验室,兰州 730020
2. State Key Laboratory of Herbage Improvement and Grassland Agro-ecosystems, Lanzhou 730020, China;
3. Key Laboratory of Grassland Livestock Industry Innovation Ministry of Agriculture, Lanzhou 730020, China
草原是全球和我国面积最大的陆地生态系统[1], 是人类重要的生态安全、食物安全和社会安全屏障[2]。然而, 由于人为活动和自然因素的共同影响, 世界草原约70%退化, 我国黄土高原是世界上受侵蚀最严重的地区之一, 主要表现为植被稀疏, 黄土裸露, 生态系统脆弱[3], 威胁社会经济可持续发展[4], 因此, 退化草原修复是黄土高原生态恢复的关键。划破作为草原生态修复的基础性措施, 为补播、施肥和有害生物防控等其它草原改良措施创造条件[5]。然而, 划破强度对草原植被和土壤的影响缺乏深入研究, 这却是草原生态修复和可持续管理的基础之一。
划破改善土壤的通透性和透水性, 促进植物生长发育, 提高地上生物量[6—7];也可能改变草原植物的群落组成, 影响草原土壤理化性质[8]。德国中部沼泽区, 划破提高草原生态系统的稳定性, 6—10年后草原生态修复较为理想[9]。巴西喜拉多热带草原, 划破通过改变土壤微生物和养分, 促进植被恢复[10]。青海省高寒草甸, 划破提升退化草原地上生物量30%—50%, 维持土壤稳定性[11]。甘肃省甘南州亚高山草甸, 轻度和中度划破增加草原物种丰富度, 重度划破则相反[12]。甘肃省玛曲县高寒草甸, 划破增加草原生产力, 降低物种丰富度, 轻度划破提高禾草丰富度和优质牧草占比[13]。综上, 划破强度是影响草原生产力和物种多样性的重要因素之一, 并对生产力—物种多样性关系的作用呈“驼峰”型曲线变化, 存在明显阈值, 在阈值的左侧, 随着划破强度增加, 植物群落物种多样性和生物量逐渐增大, 当划破强度超过阈值, 植物群落物种多样性和生物量显著降低, 因此, 明确划破强度的阈值是通过划破改良草原的关键(图 1)。

|
| 图 1 划破强度对生态系统的作用概念图 Fig. 1 Conceptual diagram of the effect of ripped intensity on ecosystem |
黄土高原位于我国东部季风区向西北干旱区的过渡地带, 草原面积20.15万km2, 天然草原面积占该区草原总面积的92.2%[14], 是全球受人类活动影响历史最悠久、强度最大的草原之一[15], 对划破敏感[16]。划破作为一种有效的草原修复方式, 可以显著提高草原物种多样性和生产力。因此, 为了恢复退化草原, 保持草原稳定健康发展, 本研究在黄土高原典型草原设置了3个划破强度, 通过比较3年后划破带和未划破带的植物群落物种多样性、生物量和土壤水分, 试图明确划破强度对草原生态修复是否存在作用和阈值, 为草原生态修复提供参考。
1 材料与方法 1.1 研究区概况研究区位于兰州大学环县草地农业试验站, 地处甘肃省环县甜水镇大良洼村(37.12°N, 106.82°E, 海拔1650 m)。温带大陆性季风气候, 年平均气温7.1℃, 年平均降水量359.3 mm(图 2), 年平均蒸发量1993.3 mm[17]。土壤类型为砂质黄绵土, 草原类型为微温微干典型草原[18], 植物群落由10科植物组成, 其中豆科植物7种, 禾本科植物7种, 菊科植物5种, 3科植物共占植物群落总数的67.9%(表 1)。生活型以多年生植物为主, 主要优势物种有达乌里胡枝子(Lespedeza davurica)、长芒草(Stipa bungeana Trin)、茵陈蒿(Artemisia capillaris Thunb)[19]等。植物返青期为3月下旬至4月上旬, 枯黄期为9月下旬至10月上旬。

|
| 图 2 研究区1998—2000年月均降水量和月均温 Fig. 2 Monthly average precipitation and temperature in the study area from 1998 to 2000 |
| 植物种类 Species |
科 Department |
植物生活型 Plant life form |
| 达乌里胡枝子Lespedeza davurica | 豆科 | 多年生 |
| 沙打旺Leguminosae | 豆科 | 多年生 |
| 苜蓿Medicago Sativa Linn | 豆科 | 多年生 |
| 米口袋Gueldenstaedtia verna(Georgi)Boriss | 豆科 | 多年生 |
| 黄花棘豆Oxytropis ochrocephala Bunge | 豆科 | 多年生 |
| 乳白花黄芪Astragalus galactites Pall | 豆科 | 多年生 |
| 砂珍棘豆Oxytropis psamocharis | 豆科 | 多年生 |
| 早熟禾Poa annua L | 禾本科 | 一年生 |
| 赖草Leymus secalinus (Georgi) Tzvel | 禾本科 | 多年生 |
| 白草Pennisetum centrasiaticum Tzvel | 禾本科 | 多年生 |
| 狗尾草Setaria viridis (L.) Beauv | 禾本科 | 一年生 |
| 长芒草Stipa bungeana Trin | 禾本科 | 多年生 |
| 糙隐子草Cleistogenes squarrosa (Trin.) Keng | 禾本科 | 多年生 |
| 野黍Eriochloa villosa (Thunb.) Kunth | 禾本科 | 一年生 |
| 蒲公英Taraxacum mongolicum Hand.-Mazz | 菊科 | 多年生 |
| 冷蒿Artemisia frigida Willd | 菊科 | 多年生 |
| 阿尔泰狗娃花Heteropappus altaicus (Willd.) Novopokr | 菊科 | 多年生 |
| 茵陈蒿Artemisia capillaris Thunb | 菊科 | 多年生 |
| 山苦荬Ixeris denticulata | 菊科 | 多年生 |
| 田旋花Convolvulus arvensis L | 旋花科 | 多年生 |
| 打碗花Calystegia hederacea Wall | 旋花科 | 一年生 |
| 扁蓄Polygonumaviculare L | 藜科 | 一年生 |
| 沙蓬Agriophyllum squarrosum (L.) Moq | 藜科 | 一年生 |
| 二裂委陵菜Potentilla bifurca Linn | 蔷薇科 | 多年生 |
| 地梢瓜Cynanchum thesioides (Freyn) K. Schum. | 萝藦科 | 多年生 |
| 远志Polygala tenuifolia Willd | 远志科 | 多年生 |
| 酢浆草Oxalis corniculata L | 酢浆草科 | 多年生 |
| 蚓果芥Torularia humilis (C. A. Mey.) O. E. Schulz | 十字花科 | 多年生 |
于1998年选择地形平坦、植物群落较为一致的区域, 随机设置了3个100 m×100 m的样地。采用耙宽3.6 m的圆盘耙划破草原, 划破深度为10 cm, 耙片划破间距为25 cm(图 3)。划破强度(Ripped intensity, RI)为划破带(Ripped belt, RB)占整个区域(Whole area, WA)的面积比例, 共设27.4% (RB1), 46.3% (RB2)和61.9% (RB3)3个划破强度, 与这三个划破强度相对应的是3个未划破带(Unripped belt, URB), 面积分别为72.6% (URB1)、53.7% (URB2)和38.1% (URB3), 每个区域4次重复。

|
| 图 3 划破强度示意图 Fig. 3 Schematic diagram of ripping intensity 灰色的线条代表划破区(R), 空白区域为未划破区(UR); WA:整区;RB:划破带;URB:未划破带 |
划破带和未划破带的每个区域随机设置4个1 m×1 m的样方。划破3年后, 在植物生长旺盛期8月进行植被调查并取样, 分种统计植物物种数量, 随后齐地面分种刈割植物地上部分, 65℃烘至恒重, 获取植物地上生物量指标。然后在每个样方中, 用直径100 mm的根钻分层(10 cm一层)取2钻0—40 cm的土样。土样过2 mm筛获得根样, 蒸馏水洗净烘干至恒重, 获取植物地下生物量指标。少部分土壤样品放置铝盒中, 105℃烘至恒重, 获取土壤水分。
1.4 数据分析群落生物量包括地上生物量(Aboveground biomass, AGB)、地下生物量(Belowground biomass, BGB)和总生物量(Total biomass, TB)。
每个划破强度WA的生物量为划破带和未划破带生物量与其面积比例的加权平均值。
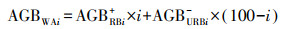
|
(1) |
式中, AGBWAi、AGBRBi+和AGBURBi-为划破强度i的整区、划破带和未划破带的地上生物量。
同理计算UGBWAi、TBWAi和土壤水分。
根冠比(Root-shoot ratio, RSR)为每个区域UGB与AGB的比值。
总物种数是一个区域内所有物种数, 3个整区仅计算总物种数。
物种丰富度(SR)是一个区域内单位面积的物种数。
Shannon Weiner指数
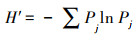
|
(2) |
Simpson指数
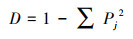
|
(3) |
均匀度指数
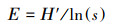
|
(4) |
式中, Pj是样方内第j个物种数量与群落物种数的比例[20]。
群落Sørensen相似性指数
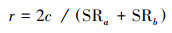
|
(5) |
式中, SRa+SRb分别为a和b两个样地的物种数, c为两个样地共有的物种数[21]。
1.5 统计分析使用SPSS 23.0(IBM, 美国)进行数据统计分析。采用单因素方差(one-way ANOVA)分析划破强度对植物群落物种多样性、生物量、根冠比和土壤水分的影响, Duncan法比较3个划破强度下各指标之间的差异(α=0.05), 划破强度对相关指标的影响采用二次项回归, 植物群落物种多样性、生物量及土壤水分之间的相关性采用Origin Pro 2022b插件correlation plot分析, 利用Excel 2018和Origin Pro 2022b软件作图。图表中数据为平均值±标准误。
2 结果与分析 2.1 植物群落物种多样性、生物量和土壤水分对划破强度和划破区域的响应划破强度和划破区域显著影响植物群落物种多样性、生物量和土壤水分。在3个划破强度下, 划破带和整区的地上生物量、地下生物量和总生物量差异显著(P<0.05)。划破带、未划破带和整区的地上生物量、地下生物量和总生物量在27.4%和46.3%划破强度下差异显著(P<0.05), 物种丰富度、土壤水分和根冠比在61.9%划破强度下差异显著(P < 0.05)(图 4)。

|
| 图 4 植物群落物种多样性、生物量和土壤水分对划破强度和划破区域的响应 Fig. 4 Response of plant community species diversity, biomass and soil moisture to ripped intensity and ripped area 大写字母代表三个划破强度下划破带、未划破带和整区各指标之间的差异, 小写字母代表同一划破强度下划破带、未划破带和整区各指标之间的差异 |
划破带、未划破带和整区的植物群落物种丰富度随划破强度的增加呈“驼峰”型曲线变化, 划破强度为46.3%时最高, 分别为14、16和22种/m2。3个划破强度下未划破带植物群落物种丰富度分别高于划破带9.1%—27.3%、6.3%—60.8%和7.1%—9.1%。划破带和未划破带总物种数随划破强度的增加呈“驼峰”型曲线变化, 在46.3%划破强度下最高达到18和20种/m2, 整区总物种数随划破强度的上升呈“马鞍”型曲线变化, 总物种数依次为未划破带>划破带>整区。根据拟合方程, 划破强度为43.7%、50.1%和55.3%时, 划破带、未划破带和整区植物群落物种丰富度分别达到峰值, 划破强度为43.7%—55.3%时物种丰富度最高(图 5)。

|
| 图 5 划破强度对植物群落物种丰富度的影响 Fig. 5 Effect of ripped intensity on species richness **, P < 0.01 |
3个划破强度中, 划破带和未划破带的Simpson指数、Shannon Weiner指数和均匀度指数差异显著(P < 0.01)。划破带Simpson指数、Shannon Weiner指数和均匀度指数较未划破带高。划破强度为46.3%时, Simpson指数、Shannon Weiner指数和均匀度指数均最低(表 2)。
| 划破强度/% Ripping intensity |
取样区域 Sampling area |
Simpson指数 Simpson index |
Shannon Weiner指数 Shannon Weiner index |
均匀度指数 Evenness index |
| 27.4 | 未划破带 | 0.46±0.00B | 1.14±0.01B | 0.45±0.01B |
| 划破带 | 0.82±0.01A | 1.95±0.05A | 0.81±0.01A | |
| 46.3 | 未划破带 | 0.24±0.04B | 0.62±0.07B | 0.22±0.02B |
| 划破带 | 0.76±0.03A | 1.67±0.07A | 0.63±0.03A | |
| 61.9 | 未划破带 | 0.71±0.01A | 1.51±0.03A | 0.58±0.02A |
| 划破带 | 0.76±0.02AB | 1.97±0.08AB | 0.71±0.03B | |
| 误差来源Source of error | ||||
| 划破强度Ripping intensity | *** | *** | *** | |
| 取样区域Sampling area | *** | *** | *** | |
| 划破强度×取样区域Ripping intensity×Sampling area | *** | *** | *** | |
| A和B表示对应划破强度下划破带与未划破带的Simpson指数、Shannon Weiner指数及均匀度指数的显著性 | ||||
划破带和整区的植物群落相似性随划破强度的增加呈先降低后升高的“马鞍”型曲线变化, 整区的植物群落相似性显著高于划破带23.85%—119.23%和未划破带44.43%—84.55%, 划破带和未划破带的群落相似性较低。3个划破强度下, 未划破带和划破带植物群落相似性负相关, 整区植物群落相似性的拟合优度最高(图 6)。

|
| 图 6 划破强度对植物群落相似性的影响 Fig. 6 Effect of ripped intensity on similarity of plant communities |
划破带、未划破带和整区的地上生物量随划破强度的增加呈“驼峰”型曲线变化, 划破强度为46.3%时最高, 分别为33.90 g/m2、95.38 g/m2、69.32 g/m2。3个划破强度下未划破带地上生物量分别是划破带的5.63%—7.53、1.21—1.62、2.63—3.1倍。划破带和未划破带地下生物量随划破强度的上升分别呈“马鞍”型和“驼峰”型曲线变化, 未划破带地下生物量最高, 3个划破强度下分别高于划破带88.2%—134.6%、52.4%—67.8%、2.5%—16.6%。总生物量随划破强度的增强呈“驼峰”型曲线变化, 划破带总生物量增速较未划破带和整区快, 未划破带总生物量高于划破带103.9%—152.9%、59.3%—75.8%、9.1%—22.6%。根冠比随划破强度的增大呈“马鞍”型曲线变化, 3个划破强度下划破带的根冠比显著高于未划破带0.21—3.15、0.46—0.63、2.12—3.00倍(图 7)。

|
| 图 7 划破强度对植物群落生物量的影响 Fig. 7 Effect of ripped intensity on plant community biomass ***, P < 0.001 |
划破强度为45.7%、45.2%和43.8%时, 划破带、未划破带和整区的地上生物量分别达到峰值39.4 g/m2、96.5 g/m2和70.4 g/m2, 分别可解释群落地上生物量99.4%、96.5%和98.2%的变异。综上所述, 当划破强度为43.8%—45.7%时群落植被地上生物量最高。
2.4 划破强度对草原土壤水分的影响划破带、未划破带和整区的土壤水分随划破强度的增加呈“马鞍”型曲线变化, 在46.3%划破强度下最低, 分别为11.26%、11.02%和11.13%。未划破带的土壤水分在27.4%划破强度下高于划破带2.9%—9.8%。划破强度为46.3%和61.9%时, 划破带土壤水分分别高于未划破带0.9%—3.2%和3.5%—4.6%。根据拟合方程, 划破强度为56.2%、50.1%和56.0%时, 划破带、未划破带和整区的土壤水分最低, 分别为9.9%、11.9%和9.7%(图 8)。

|
| 图 8 划破强度对土壤水分的影响 Fig. 8 Effect of ripped intensity on soil moisture content ***, P<0.001 |
划破带地上生物量与物种丰富度显著正相关(P < 0.05), 与Shannon Weiner指数和均匀度指数显著负相关(P < 0.05)。未划破带地上生物量与物种丰富度显著正相关(P < 0.05), 与Shannon Weiner指数、Simpson指数和均匀度指数显著负相关(P < 0.05), 根冠比与Shannon Weiner指数、Simpson指数和均匀度指数显著正相关(P < 0.05)。整区地上生物量与物种丰富度显著正相关(P < 0.05), 与Shannon Weiner指数、Simpson指数和均匀度指数显著负相关(P < 0.05), 总生物量与Simpson指数和均匀度指数显著负相关(P < 0.05)(图 9)。

|
| 图 9 植物群落物种多样性-生产力关系对划破强度的响应 Fig. 9 Response of plant community species diversity-productivity relationship to ripped intensity 图依次代表划破带、未划破带和整区植物群落地上生物量、地下生物量、总生物量和根冠比与生物多样性的关系, *, P < 0.05 |
划破强度为27.4%和61.9%时, 划破带和未划破带的地上生物量与土壤水分显著负相关(P < 0.05), 划破带地上生物量在46.3%划破强度下与土壤水分显著负相关(P < 0.05)。划破带、未划破带和整区的地下生物量、总生物量与土壤水分显著负相关(P < 0.05)(图 10)。

|
| 图 10 植物群落生物量与土壤水分的关系 Fig. 10 Relationship between plant biomass and soil moisture constant *, P < 0.05 |
土壤水分是植物生产力的重要限制因素, 划破调节土壤水分, 不断变化的水分状况影响养分的吸收和运输[22], 进而改变群落物种组成[23]。划破带物种丰富度低于未划破带, 可能由于划破引起较高的土壤透气性诱导了土壤保水率降低, 抑制了植物生长所需的水资源分配, 无法满足植物生长所需的基本条件, 物种多样性降低[24]。另一方面, 划破较大程度上抑制了矮小的物种以及实生苗的生长并降低了它们对其它物种的竞争能力, 物种丰富度随之降低[25]。
物种多样性随划破强度增加呈“驼峰”型曲线变化。划破强度为46.3%时植物群落物种丰富度最高, 可能是因为划破改善土壤中枯死的根系絮结, 提高生境质量[26], 有利于植物生长和其它物种的入侵, 从而增加物种丰富度[27]。在此范围内, 生物量随物种丰富度的上升而增加, 且与地下生物量不同的是划破带地上生物量随物种丰富度和Simpson指数的增加呈“马鞍”型曲线变化。一般而言, 物种丰富度和Simpson指数较高时, 群落地上生物量也较高[20], 在山地草甸和荒漠草甸的研究中也得到了同样的结论[28]。但本研究结果与之不同的是物种丰富度或Simpson指数较高时具有较低的地上生物量, 其原因可能是群落中物种较多(表 1), 植物根系优先生长, 地上部分发育缓慢, 生物量低。不同的划破强度改变了植物群落的组成, 一定程度上减少了竞争的相互作用, 促进了优势种和从属种的共存[29]。划破降低凋落物盖度, 提高一年生物种的光照利用率[30]。相反, 27.4%划破强度下物种丰富度最低, 可能是划破的干扰大于土壤水分的限制, 在本研究区域中土壤水分对植物生长的影响效应降低, 划破后物种间对光照和营养元素等资源的竞争加剧, 导致群落物种多样性降低[31]。在61.9%划破强度下, 凋落物的分解速率降低, 形成物理屏障, 影响种子的萌发和幼苗的存活[32], 植物生长发育所需的养分传输受到抑制, 导致植物长势弱且多样性降低。
3.2 划破对草原植物群落生物量的影响黄土高原典型草原划破3年后, 未划破带地上、地下生物量显著高于划破带, 可能因为:一划破去除多年生植物群落地上部分, 影响水分迁移, 进而抑制了植物群落地上部分的再生, 导致群落发生低补偿生长[33—34], 在祁连山高寒矮嵩草草甸的研究中也得到同样的结论[6];二划破过程中机械损伤以及人为干扰等因素破坏地表结构减少植被覆盖, 土壤水分蒸发加剧, 不利于地下部分的生长发育[35—36]。另外, 划破也可以通过降低土壤紧实度, 削弱了土壤保水保肥能力, 抑制植物根部生长[11], 使划破带植物群落地下生物量低于未划破带, 这与在祁连山中度退化高寒草甸的研究结果相反, 可能因为高寒草甸的年均降水量大于黄土高原典型草原, 砂质土壤的保水能力较差, 土壤含水量也随之降低[37—38];三划破可以减少凋落物的覆盖, 导致土壤水分流失加剧, 影响土壤温度, 土壤受侵蚀程度大[39], 不利于植物的生长和恢复[40];四划破降低了土壤微生物活性, 抑制土壤有机碳矿化[41], 减少植物群落生物量[42]。
划破强度为46.3%时植物群落地上部生长发育较好, 此时植物对资源的分配和利用效率最高, 并能削弱划破对土壤水分的负作用[43]。27.4%划破强度下群落生物量偏低, 可能因为植物根系呼吸受到抑制, 影响养分吸收效率。61.9%划破强度较大程度地破坏了植被和土壤原有结构, 增加了植物种子的掩埋深度, 抑制种子萌发、植被恢复和地上部分的生长[44]。
划破对植物群落地上、地下生物量之间分配的影响是植物对资源的权衡和环境适应能力的体现。划破带根冠比显著高于未划破带, 说明划破作为一种对草原的干扰方式和管理手段, 会导致大量氮从草原生态系统中去除, 从而减少土壤中氮的可用性, 迫使植物将更多的光合产物分配给地下器官[45], 进而导致群落内植物出现矮小化和向下生长的趋势[46]。划破强度为46.3%时根冠比最低, 表明该划破强度下分配模式倾向于地上部分的生长, 以达到植物对光资源竞争的最大利用率[47]。61.9%划破强度下土壤水分增加, 植物生长竞争力强, 更容易吸收深层土壤水分, 诱导更大的地下生物量分配[48]。
4 结论本试验以黄土高原典型草原为研究对象, 旨在探讨不同划破强度对草原植物群落物种多样性、生物量和土壤水分的影响。通过对比三个划破强度下退化草原的恢复状况, 发现:
划破是草地培育和草原修复的重要技术措施, 在生态环境较为脆弱的黄土高原采用划破可以改善退化草地, 维持生态系统的稳定性。划破强度显著影响退化草地恢复状况, 黄土高原典型草原植物群落对划破强度存在响应阈值, 划破强度为43.7%—55.3%时, 植物群落物种丰富度最高, 划破强度为43.8%—45.7%时植物群落地上生物量最高, 表明划破强度对典型草原的作用阈值为43.7%—55.3%。
| [1] |
Hou F J, Jia Q M, Lou S N, Yang C T, Ning J, Li L, Fan Q S. Grassland agriculture in China: a review. Frontiers of Agricultural Science and Engineering, 2021, 8(1): 35. DOI:10.15302/J-FASE-2020378 |
| [2] |
Bardgett R D, Bullock J M, Lavorel S, Manning P, Schaffner U, Ostle N, Chomel M, Durigan G, Fry E L, Johnson D, Lavallee J M, Provost G L, Luo S, Png K, Sankaran M, Hou X Y, Zhou H K, Ma L, Ren W B, Li X L, Ding Y, Li Y H, Shi H X. Combatting global grassland degradation. Nature Reviews Earth & Environment, 2021, 2(10): 720-735. |
| [3] |
Dong L B, Li J W, Liu Y L, Hai X Y, Li M Y, Wu J Z, Wang X Z, Shangguan Z P, Zhou Z C, Deng L. Forestation delivers significantly more effective results in soil C and N sequestrations than natural succession on badly degraded areas: Evidence from the Central Loess Plateau case. Catena, 2022, 208: 105734. DOI:10.1016/j.catena.2021.105734 |
| [4] |
Gang C C, Zhou W, Chen Y Z, Wang Z Q, Sun Z G, Li J L, Qi J G, Odeh I. Quantitative assessment of the contributions of climate change and human activities on global grassland degradation. Environmental Earth Sciences, 2014, 72(11): 4273-4282. DOI:10.1007/s12665-014-3322-6 |
| [5] |
Wang S L, Hu A, Zhang J, Hou F J. Effects of grazing season and stocking rate on seed bank in sheep dung on the semiarid Loess Plateau. The Rangeland Journal, 2019, 41(5): 405. DOI:10.1071/RJ19036 |
| [6] |
李小龙, 曹文侠, 张晓燕, 李文, 徐长林, 师尚礼. 划破草皮对祁连山不同地形高寒矮嵩草草甸土壤呼吸特征的影响. 草原与草坪, 2020, 40(2): 30-38. |
| [7] |
Mayel S, Jarrah M, Kuka K. How does grassland management affect physical and biochemical properties of temperate grassland soils? A review study. Grass and Forage Science, 2021, 76(2): 215-244. DOI:10.1111/gfs.12512 |
| [8] |
刘晶晶, 尹亚丽, 李世雄, 赵文, 董怡玲, 苏世锋. 不同调控措施对中度退化高寒草甸植被及土壤理化性质的影响. 草地学报, 2021, 29(9): 2074-2080. |
| [9] |
Streitberger M, Holtmann L, Bültmann H, Schmidt C, Fartmann T. Effects of montane heathland rejuvenation on phytodiversity depend on management intensity and succession. Ecological Engineering, 2021, 168: 106292. DOI:10.1016/j.ecoleng.2021.106292 |
| [10] |
Assis G B, Pilon N A L, Siqueira M F, Durigan G. Effectiveness and costs of invasive species control using different techniques to restore cerrado grasslands. Restoration Ecology, 2021, 29: e13219. |
| [11] |
李小龙, 曹文侠, 李文, 张晓燕, 徐长林, 韦应莉, 师尚礼. 划破草皮对不同地形高寒草甸草原土壤特征及地下生物量的影响. 草业学报, 2016, 25(6): 26-33. |
| [12] |
Wang Y X, Sun Y, Chang S H, Wang Z F, Fu H, Zhang W G, Hou F J. Restoration practices affect alpine meadow ecosystem coupling and functions. Rangeland Ecology & Management, 2020, 73(3): 441-451. |
| [13] |
Wang Y F, Cai Y F, Hou F J, Jia Z J, Bowatte S. Sheep grazing impacts on soil methanotrophs and their activity in typical steppe in the Loess Plateau China. Applied Soil Ecology, 2022, 175: 104440. DOI:10.1016/j.apsoil.2022.104440 |
| [14] |
袁梓裕, 张路, 廖李容, 王杰, 雷石龙, 刘国彬, 方怒放, 张超. 黄土高原草地植物多样性与群落稳定性的关系及其驱动因素. 生态学报, 2023, 43(1): 60-69. |
| [15] |
Naeem S, Zhang Y Q, Zhang X Z, Tian J, Abbas S, Luo L L, Meresa H K. Both climate and socioeconomic drivers contribute to vegetation greening of the Loess Plateau. Science Bulletin, 2021, 66(12): 1160-1163. DOI:10.1016/j.scib.2021.03.007 |
| [16] |
Hu A, Chang S H, Chen X J, Hou F J, Nan Z B. Temporal heterogeneity has no effect on the direction of succession in abandoned croplands in a semiarid area of northwest China. Land Degradation & Development, 2021, 32(1): 91-100. |
| [17] |
Li L, Zhang J, He X Z, Hou F J. Different effects of sheep excrement type and supply level on plant and soil C: N: P stoichiometry in a typical steppe on the loess plateau. Plant and Soil, 2021, 462(1): 45-58. |
| [18] |
Ren J Z, Hu Z Z, Zhao J, Zhang D G, Hou F J, Lin H L, Mu X D. A grassland classification system and its application in China. The Rangeland Journal, 2008, 30(2): 199. DOI:10.1071/RJ08002 |
| [19] |
Li L, He X Z, Zhang X M, Hu J Q, Wang M Y, Wang Z, Hou F J. Different effects of grazing and nitrogen addition on ecosystem multifunctionality are driven by changes in plant resource stoichiometry in a typical steppe. Plant and Soil, 2022, 481(1): 179-194. |
| [20] |
周洁艳, 安玉峰, 李多才, 张育文, 李岚. 祁连山高寒草原α多样性与生产力关系的研究. 中国草地学报, 2022, 44(3): 9-16. |
| [21] |
Buss J, Pinno B D, Quideau S. A comparison between reclamation stockpile and boreal forest seed banks and plant communities. Restoration Ecology, 2020, 28(5): 1247-1256. |
| [22] |
Fischer C, Leimer S, Roscher C, Ravenek J, Kroon H D, Kreutziger Y, Baade J, Beler H, Eisenhauer N, Weigelt A, Mommer L, Lange M, Gleixner G, Wilcke W, Schrder B, Hildebrandt A. Plant species richness and functional groups have different effects on soil water content in a decade-long grassland experiment. Journal of Ecology, 2019, 107(1): 127-141. |
| [23] |
Liu M, Liu G H, Zheng X X. Spatial pattern changes of biomass, litterfall and coverage with environmental factors across temperate grassland subjected to various management practices. Landscape Ecology, 2015, 30(3): 477-486. |
| [24] |
Vogel A, Scherer-Lorenzen M, Weigelt A. Grassland resistance and resilience after drought depends on management intensity and species richness. PloS one, 2012, 7(5): e36992. |
| [25] |
徐鑫磊, 宋彦涛, 赵京东, 乌云娜. 施肥和刈割对呼伦贝尔草甸草原牧草品质的影响及其与植物多样性的关系. 草业学报, 2021, 30(7): 1-10. |
| [26] |
李小龙, 曹文侠, 徐长林, 李文, 刘浩栋, 张晓燕, 师尚礼. 划破草皮对不同地形高寒草甸草原植被特征的影响. 草地学报, 2016, 24(02): 309-316. |
| [27] |
王悦骅, 靳宇曦, 王忠武, 韩国栋. 8年围封对内蒙古荒漠草原植物和土壤的影响. 草地学报, 2021, 29(10): 2339-2345. |
| [28] |
Chen L L, Baoyin T, Minggagud H. Effects of mowing regimes on above- and belowground biota in semi-arid grassland of Northern China. Journal of Environmental Management, 2021, 277: 111441. |
| [29] |
Li D, Nie Y Y, Xu L J, Ye L M. Enclosure in combination with mowing simultaneously promoted grassland biodiversity and biomass productivity. Plants, 2022, 11(15): 2037. |
| [30] |
Miao R H, Ma J, Liu Y Z, Liu Y C, Yang Z L, Guo M X. Variability of aboveground litter inputs alters soil carbon and nitrogen in a coniferous-broadleaf mixed forest of Central China. Forests, 2019, 10(2): 188. |
| [31] |
王萌, 徐冰, 张大勇, 朱璧如. 内蒙古草原锡林河流域植物群落生物量及多样性沿土壤水分含量梯度的变化. 北京师范大学学报: 自然科学版, 2016, 52(4): 445-449. |
| [32] |
Chen B M, D'Antonio C M, Molinari N, Peng S L. Mechanisms of influence of invasive grass litter on germination and growth of coexisting species in California. Biological Invasions, 2018, 20(7): 1881-1897. |
| [33] |
包乌云, 赵萌莉, 红梅, 高新磊. 刈割对人工草地产量和补偿性生长的影响. 中国草地学报, 2015, 37(5): 46-51. |
| [34] |
Wang B, Li P P, Huang C H, Liu G B, Yang Y F. Effects of root morphological traits on soil detachment for ten herbaceous species in the Loess Plateau. Science of The Total Environment, 2021, 754: 142304. |
| [35] |
Yang Z P, Minggagud H, Baoyin T, Li Y H. Plant production decreases whereas nutrients concentration increases in response to the decrease of mowing stubble height. Journal of environmental management, 2020, 253: 109745. |
| [36] |
李凤霞, 李晓东, 周秉荣, 祁栋林, 王力, 傅华. 放牧强度对三江源典型高寒草甸生物量和土壤理化特征的影响. 草业科学, 2015, 32(1): 11-18. |
| [37] |
孙天雨, 王雪, 李丹洋, 刘欣蕊, 王瑞丽, 张硕新. 黄土高原草地根系生物量沿环境梯度变化规律. 生态学报, 2021, 41(15): 6234-6244. |
| [38] |
刘晶晶, 尹亚丽, 李世雄, 赵文, 苏世锋, 董怡玲. 调控措施对祁连山中度退化高寒草甸土壤的影响. 应用生态学报, 2022, 33(4): 988-994. |
| [39] |
Lu X, Song X Y, Fu N, Cui S Y, Li L J, Li H Y, Li Y L. Effects of forest litter cover on hydrological response of hillslopes in the Loess Plateau of China. Catena, 2019, 181: 104076. |
| [40] |
Kotas P, Choma M, šantrůčková H, LepŠ J, Tříska J, KaŠtovská E. Linking above-and belowground responses to 16 years of fertilization, mowing, and removal of the dominant species in a temperate grassland. Ecosystems, 2017, 20(2): 354-367. |
| [41] |
Zhang S B, Fang Y Y, Luo Y, Li Y C, Ge T D, Wang Y X, Wang H L, Yu B, Song X Z, Chen J H, Zhou J S, Li Y F, Chang S X. Linking soil carbon availability, microbial community composition and enzyme activities to organic carbon mineralization of a bamboo forest soil amended with pyrogenic and fresh organic matter. Science of The Total Environment, 2021, 801: 149717. |
| [42] |
蒋翔, 马建霞. 我国草地生态恢复对不同因素响应的Meta分析. 草业学报, 2021, 30(2): 14-31. |
| [43] |
张存厚, 杨丽萍, 越昆, 刘朋涛, 张德龙. 锡林郭勒典型草原土壤水分对降水过程的响应. 干旱区资源与环境, 2022, 36(8): 133-139. |
| [44] |
Sun J, Ma B B, Lu X Y. Grazing enhances soil nutrient effects: Trade-offs between aboveground and belowground biomass in alpine grasslands of the Tibetan Plateau. Land Degradation & Development, 2018, 29(2): 337-348. |
| [45] |
Luo Y, Wang X Y, Cui M, Wang J, Gao Y Z. Mowing increases fine root production and root turnover in an artificially restored Songnen grassland. Plant and Soil, 2021, 465(1): 549-561. |
| [46] |
张晴晴, 梁庆伟, 娜日苏, 杨秀芳. 刈割对天然草地影响的研究进展. 畜牧与饲料科学, 2018, 39(1): 33-42. |
| [47] |
Wen J, Qin R M, Zhang S X, Yang X Y, Xu M H. Effects of long-term warming on the aboveground biomass and species diversity in an alpine meadow on the Qinghai-Tibetan Plateau of China. Journal of Arid Land, 2020, 12(2): 252-266. |
| [48] |
Zhou L Y, Zhou X H, He Y H, Fu Y L, Du Z G, Lu M, Sun X Y, Li C H, Lu C Y, Liu R Q, Zhou G Y, Bai S H, Thakur M P. Global systematic review with meta-analysis shows that warming effects on terrestrial plant biomass allocation are influenced by precipitation and mycorrhizal association. Nature Communications, 2022, 13(1): 4914. |