 2023, Vol. 43
2023, Vol. 43文章信息
- 袁洪艺, 杜灵通, 潘海珠, 乔成龙, 田静, 易志远, 吴宏玥, 张祎, 施光耀, Irumva Olivier
- YUAN Hongyi, DU Lingtong, PAN Haizhu, QIAO Chenglong, TIAN Jing, YI Zhiyuan, WU Hongyue, ZHANG Yi, SHI Guangyao, IRUMVA Olivier
- 基于参数优化的人工灌丛生态系统碳水通量模拟
- Simulation of carbon and water fluxes of planted shrub ecosystem in desert steppe biome zone based on parameter optimization
- 生态学报. 2023, 43(13): 5546-5557
- Acta Ecologica Sinica. 2023, 43(13): 5546-5557
- http://dx.doi.org/10.5846/stxb202207031892
-
文章历史
- 收稿日期: 2022-07-03
- 网络出版日期: 2023-03-07





2. 宁夏大学西北退化生态系统恢复与重建教育部重点实验室, 银川 750021;
3. 宁夏大学生态环境学院, 银川 750021
2. Key Laboratory for Restoration and Reconstruction of Degraded Ecosystem in Northwest China of Ministry of Education, Ningxia University, Yinchuan 750021, China;
3. School of Ecology and environment, Ningxia University, Yinchuan 750021, China
陆地生态系统碳水循环是地圈和生物圈物质与能量交换的重要过程, 当前, 气候变化和人类活动引起的陆地生态系统碳水循环过程及其内在机制的变化研究已经成为全球变化生态学研究的热点。其中, 占据着全球陆地面积40%的旱地(干旱和半干旱)系统对全球陆地碳汇的趋势和年际变化具有显著的影响[1]。此外, 有研究表明全球碳通量的年际变化可能由半干旱生态系统主导, 但这种变化的潜在机制在这些特定地区尚不清楚[2], 因此研究其生态系统的碳水循环, 可为干旱半干旱区应对全球气候变化提供理论依据。模型模拟是开展陆地生态系统碳水循环研究的重要手段, 自20世纪70年代以来已发展出一系列的模型, 主要分为经验模型(如Miami和Thornthwaite模型)、生态过程机理模型(如TEM、BATS和Biome-BGC模型)和遥感模型(如VPM和CASA模型)三大类[3]。其中, Biome-BGC模型能够详细描述植物生态生理过程及模拟不同空间尺度(站点—区域—全球)碳水通量变化的时间特征(日、月和年), 具有外延性好、机理性强和综合性高的特点, 是当前分析与预测大尺度生态系统过程的首要选择[4—5], 因而被广泛的应用于不同类型生态系统的碳水通量模拟研究中。Thornton等[6]以北美常绿针叶林为研究对象, 模拟了该生态系统碳水通量对气候变化和历史干扰的响应;Chiesi等[7]利用Biome-BGC模型, 研究了意大利地中海常绿硬叶灌木ET和GPP的变化特征;康满春等[8]利用Biome-BGC模型模拟了不同气候变化情景下杨树人工林生态系统的碳水通量及水分利用效率;张越等[9]利用Biome-BGC模型对秦岭太白红杉林生产力、碳源/汇的动态变化进行了模拟, 并对未来气候变化的强度及气候响应机理进行了研究。此外, 已有学者利用Biome-BGC模型研究了灌丛化前后荒漠草原生态系统的碳储量及蒸散变化[10—11], 表明该模型在研究荒漠草原区碳水通量变化时具有一定的适用性。
前人研究表明, Biome-BGC模型能较好的模拟陆地生态系统碳水通量变化, 但也发现模型能否准确地模拟真实的生态过程, 受到多方面因素的影响, 主要包括模型对生态系统的简化和近似、参数取值的合理性[12], 更重要的是模型模拟没有充分利用地面碳通量测量数据, 如来自涡度通量塔的测量数据[13], 这些因素都制约着模型精度的提高, 致使模拟结果存在不确定性。因此, 一些学者通过修改模型结构和内部模块的算法及数据同化方法来减小模拟误差[14—15], 另一些学者则将长时间的通量观测数据结合优化算法, 来求解模型参数的最优取值以减小模拟误差[5]。然而, 改进模型方法要求较强的专业知识和编程能力, 且在实际模拟工作中缺乏普适性[16], 数据同化方法又涉及大量的数据处理和计算过程[12]。因此, 相对改进模型和数据同化而言, 参数优化更易实现。目前, 参数优化方法逐渐替代了人工试错法和经验法等传统的模型调参方法, 避免了传统模型调参方法主观性强、费时费力等缺陷。随着计算机技术快速发展和模型模拟软件的推广, 目前已发展出可自动识别搜索的优化方法, 如HYDRUS-1D、SCE-UA算法和参数估计(Parameter ESTimation, PEST)等[17], 他们可在参数空间上自动搜索, 并按照一定的寻优规则来识别较优或最优的模型参数集[5]。其中, PEST在模型参数优化方面有着独特的优势, 已在水文、生态、农业等领域得到了广泛的应用, Nolan等[18]首次利用PEST模型优化了RzwQM2农业系统模拟模型;温永斌等[19]利用PEST优化参数后的Biome-BGC模型模拟了千烟洲森林总初级生产力(Gross primary productivity, GPP)和蒸散(Evapotranspiration, ET), 得出PEST对Biome-BGC模型进行参数优化能显著提高模拟精度;梁浩等[17]利用PEST对土壤水力学参数、氮转化参数和作物遗传参数同时进行优化, 并进行了综合敏感度分析。现有研究表明, 经PEST优化的Biome-BGC模型, 其模拟效果得到明显改善[19—20], 成为提升生态系统碳水循环模拟精度的有效手段。
为此, 本研究通过气象数据和植被生理生态参数, 驱动Biome-BGC模型, 利用涡度相关系统实测数据并结合PEST方法对Biome-BGC模型的生理生态参数进行优化, 用优化后的模型对宁夏盐池荒漠草原区人工灌丛生态系统的GPP和ET进行模拟, 探讨人工灌丛生态系统碳水通量特征及与环境因子之间的关系, 以期揭示荒漠草原人工干扰下的碳水循环变化规律。
1 研究区概况及数据与方法 1.1 研究区概况本研究以位于宁夏回族自治区东部的盐池县(37°04′—38°10′N, 106°30′—107°47′E)荒漠草原为例, 该县是宁夏面积最大的县, 总面积6769 km2, 海拔1295—1951 m, 北邻毛乌素沙地, 东南接黄土高原, 地势南高北低[11]。盐池干旱少雨, 属于典型的温带大陆性气候, 1958—2017年的日平均气温为8.3℃, 年平均降水量为297mm, 全年降水分布不均, 主要集中在7—9月, 潜在蒸发量高达2136 mm[21]。土壤以灰钙土为主, 其次是黑垆土和风沙土, 疏松的土壤结构, 极易受风力侵蚀, 导致土地退化严重。盐池县自然植被在区系上属于欧亚草原区的亚洲中部类型区, 是我国中部典型草原区向西部荒漠草原区的过渡地带, 主要植被类型有灌丛、草原、草甸、沙地植被和荒漠草原植被。原生灌木植物主要以北沙柳(Salix psammophila)和小叶锦鸡儿(Caragana microphylla)为主, 草本植物主要有大针茅(Stipa grandis)、长芒草(Stipa bungeana)和冰草(Agropyron cristatum)等[22]。其中南部黄土丘陵区为干草原, 而中北部风沙区为荒漠草原, 以旱生和中旱生类植物群落为主, 在退化荒漠草原区, 人为种植了大量中间锦鸡儿(Caragana liouana)灌木用于防风固沙, 中间锦鸡儿种植条带间距6—7 m, 株距平均不足1 m, 平均高度1.40 m, 现已形成约8.9×104 hm2的以中间锦鸡儿为优势种的灌丛化荒漠草原景观[10]。本研究依托位于盐池县花马池镇杨寨子自然村的宁夏大学荒漠草原生态系统定位观测研究站(37.83°N, 107.33°E)开展, 监测点从1986年开始种植中间锦鸡儿灌木, 这一举措使得原有的生态结构和碳水循环发生极大地变化。
1.2 数据与方法 1.2.1 Biome-BGC模型简介Biome-BGC是美国蒙塔那大学陆地动态数值模拟研究小组(Numerical Terra Dynamic Simulations Group, NTSG)开发的一款生物地球化学模型, 可用来模拟常绿针叶林、常绿阔叶林、落叶针叶林、落叶阔叶林、C3草本植物、C4草本植物和灌木林7种植被类型的物质循环过程, 能够对陆地生态系统的植被、凋落物和土壤中的碳、水、氮存储和通量流动进行模拟[23], 其蒸散过程依靠Penman-Monteith公式求得[24]:
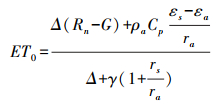
|
(1) |
式中, ET0为潜在蒸散量, Δ为饱和水汽压与温度关系曲线上的斜率, Rn为植被表面的净辐射, G为土壤热通量密度, ρa表示空气密度, Cp表示空气的比热, εs-εa表示空气蒸汽压差, γ是湿度常数, rs和ra是表面阻力和空气动力阻力。
Biome-BGC模型运行需要三个输入文件, 第一个为初始化文件, 它提供模型模拟的大部分基础信息, 包括对模拟对象的物理特性描述、模拟时间框架描述、其他所需输入文件和输出文件名称以及输出变量列表等;第二个为气象数据文件, 它包含模拟地点的温度(日最高温和日最低温)、降水量、水汽压亏缺、辐射和日照时长;第三个为植物生理生态参数文件, 包括冠层截留系数、最大气孔导度、比叶面积以及光合产物分配到植物各器官所占的比例等43个参数。
1.2.2 PEST参数优化方法参数估计(Parameter ESTimation, PEST)是一种非线性参数优化方法, 它不仅能够进行参数优化、敏感性分析, 还可用于模型的预测分析。PEST最初是针对参数多且复杂的水文模型而开发, 故应用于水文领域的研究较多, 但经不断发展现可应用于不同学科的模型优化。PEST设置灵活, 不受模型结构的限制, 可同时优化多个参数, 能够满足于不同学科对参数优化的具体需求[25—26]。PEST的目标函数是模型模拟值与实际观测值之间的差异函数, 是基于Gauss-Marquardt-Levenberg算法对目标函数求最小值来实现参数优化计算[19]。在PEST优化过程中, 其运行的基础是先对待优化参数进行识别, 核心是对参数进行优化调整。PEST方法优化Biome-BGC模型参数时, PEST与Biome-BGC两者的结构相互独立, 算法通过识别模型输入参数文件(·tpl)、观测数据指令文件(·ins)和控制文件(·pst), 以此建立PEST、Biome-BGC模型及通量观测数据之间的的联系, 最后实现彼此之间的数据交换。PEST算法优化原理及软件使用指南详见其官网(http://www.pesthomepage.org/)。
本文利用PEST对盐池荒漠草原区人工灌丛生态系统的生理生态参数进行优化, 首先从Biome-BGC的43个生理生态参数中选出要进行优化的参数, 其中用来描述植被类型及固定组合比例的参数不优化, 如被模拟的人工灌木是C3光合类型, 研究区未经历火烧等, 故火灾死亡率参数也不参与优化。对28个需要优化的参数设定取值范围, 根据White等[23]提出的模型参数平均值的置信区间为20%, 将模型各个参数的取值范围设定为(x-Δx, x+Δx), 其中x为参数基准值, Δx=0.2x, 且参数间相互独立, 最终将需要优化的参数和实测的GPP和ET值输入PEST进行参数优化, 最终生成基于GPP和基于ET的两套优化参数值。
1.2.3 数据来源Biome-BGC模型所需的日尺度气象数据来源于中国气象数据网(http://data.cma.cn/), 研究获取了盐池县1986—2018年逐日最高温度、最低温度及降雨量, 再利用山地小气候模型(MT-CLM)对这些数据进行模拟, 获得驱动Biome-BGC模型所需的七种气象指标。2016—2018年日尺度的GPP和ET来源于宁夏大学盐池荒漠草原生态定位研究站的开路涡度相关系统观测所得, 由红外气体分析仪(LI-7500A, LI-COR, 美国)和三维超声风速仪(WindMaster Pro, Gill, 英国)组成的观测系统架设在人工灌丛之上, 距地面高度3 m, 数据采样频率为10 Hz, 原始数据经异常剔除、坐标旋转和空气密度效应(WPL)订正等处理后, 用于PEST模型优化输入和Biome-BGC模型模拟结果验证。1986—2018年的大气CO2浓度数据来源于青海省瓦里关观测站, 人工灌丛的生理生态参数参照文献[27]模型缺省值。
1.2.4 数据处理与分析碳水通量数据采用EddyPro 7.0涡度协方差数据处理软件进行预处理, 数据统计分析及图形绘制在Excel 2019中完成, 相关性分析在SPSS 26.0中进行, 通过方差分析来检验模拟值与观测值的差异大小。数据投影采用地理坐标系, 大地基准面采用WGS-1984。
2 结果和分析 2.1 PEST参数优化结果利用PEST和2017年研究站涡度相关系统实测的人工灌丛逐日GPP、ET数据对28个驱动Biome-BGC模型所需的生理生态参数进行优化, 得到两套优化的参数值(表 1)。从优化结果来看, 当利用实测GPP优化参数时, PEST对整株植物死亡率等15个参数进行数值下调, 对转换生长占生长季比例等11个参数进行数值上调, 只有叶片和细根年周转率、死木质组织碳氮比2个参数未做优化调整;当利用实测的ET优化参数时, PEST对活立木年周转率等9个参数进行数值下调, 对整株植物死亡率等12个参数进行数值上调, 有7个参数未做优化调整。整体来看, PEST参数优化算法对Biome-BGC模型的大多数生理生态参数都进行了优化调整。
| 参数 Parameters |
单位 Units |
初始值[27] Initial value |
取值范围 Value range |
基于GPP的优化值 Optimization Value based on GPP |
基于ET优化值 Optimization Value based on ET |
| 转换生长占生长季比例Transfer growth period as fraction of growing season | — | 0.30 | [0.24, 0.36] | 0.36 | 0.30 |
| 落叶时段占生长季比例Litterfall as fraction of growing season | — | 0.30 | [0.24, 0.36] | 0.31 | 0.30 |
| 叶片和细根年周转率Annual leaf and fine root turnover fraction | 1/a | 1.0 | [0.8, 1.2] | 1.0 | 1.0 |
| 活立木年周转率Annual live wood turnover fraction | 1/a | 0.70 | [0.56, 0.84] | 0.84 | 0.56 |
| 整株植物死亡率Annual whole-plant mortality fraction | 1/a | 0.020 | [0.016, 0.024] | 0.016 | 0.024 |
| 细根与叶片碳分配比New fine root C: new leaf C | kgC/kgC | 0.980 | [0.784, 1.176] | 0.872 | 0.827 |
| 茎与叶片碳分配比New stem C: new leaf C | kgC/gC | 1.06 | [0.848, 1.272] | 0.848 | 1.272 |
| 活木与木质组织碳分配比New live wood C: new total wood C | kgC/kgC | 0.10 | [0.08, 0.12] | 0.08 | 0.08 |
| 粗根与茎分配比New croot C: new stem C | kgC/kgC | 0.940 | [0.752, 1.128] | 1.128 | 1.128 |
| 当前生长比例Current growth proportion | — | 0.50 | [0.40, 0.60] | 0.40 | 0.47 |
| 叶片碳氮比C: N of leaves | kgC/kgN | 10.63270 | [8.50616, 12.75924] | 9.03651 | 10.50428 |
| 凋落物碳氮比C: N of leaf litter, after retranslocation | kgC/kgN | 93.0 | [74.4, 111.6] | 93.7 | 93.0 |
| 细根碳氮比C: N of fine roots | kgC/kgN | 12.6497 | [10.1198, 15.1796] | 14.3650 | 10.1200 |
| 活木质组织碳氮比C: N of live wood | kgC/kgN | 31.5280 | [25.2224, 37.8336] | 34.6950 | 31.1550 |
| 死木质组织碳氮比C: N of dead wood | kgC/kgN | 729.0 | [583.2, 874.8] | 729.0 | 729.0 |
| 冠层截留系Canopy water interception coefficient | 1 LAI-1 d-1 | 0.0410 | [0.0328, 0.0492] | 0.0328 | 0.0480 |
| 冠层消光系数Canopy light extinction coefficient | — | 0.430 | [0.344, 0.516] | 0.340 | 0.520 |
| 叶面积与投影叶面积指数比All-sided to projected leaf arera ratio | — | 2.60 | [2.08, 3.12] | 3.10 | 2.65 |
| 冠层比叶面积Canopy average specific leaf area (projected area basis) | m2/kgC | 19.80 | [15.84, 23.76] | 20.97 | 23.42 |
| 阴生叶和阳生叶的比叶面积比例Ratio of shaded SLA: sunlit SLA | — | 2.0 | [1.6, 2.4] | 1.6 | 2.19 |
| 酮糖二磷酸羧化酶中氮含量与叶氮含量Fraction of leaf N in Rubisco | — | 0.330 | [0.264, 0.396] | 0.396 | 0.264 |
| 最大气孔导度Maximum stomatal conductance (projected area basis) | m/s | 0.0030 | [0.0024, 0.0036] | 0.0024 | 0.0036 |
| 表皮层导度Cuticular conductance (projected area basis) | m/s | 0.00010 | [0.00008, 0.00012] | 0.00008 | 0.00012 |
| 边界层导度Boundary layer conductance (projected area basis) | m/s | 0.080 | [0.064, 0.096] | 0.096 | 0.064 |
| 气孔开始缩小时的叶片水势Leaf water potential: start of conductance reduction | MPa | -0.420 | [-0.504, -0.336] | -0.441 | -0.356 |
| 气孔完全闭合时的叶片水势Leaf water potential: complete conductance reduction | MPa | -2.310 | [-2.772, -1.848] | -2.606 | -1.848 |
| 气孔开始缩小时的饱和水汽压差Vapor pressure deficit: start of conductance reduction | Pa | 930.0 | [744.0, 1116.0] | 929.8 | 930.0 |
| 气孔完全闭合时的饱和水汽压差Vapor pressure deficit: complete conductance reduction | Pa | 4100 | [3280, 4920] | 4098 | 4100 |
| GPP: 总初级生产力Gross primary productivity;ET: 蒸散evapotranspiration | |||||
由于Biome-BGC模型设置了生长季起止日期, 故对GPP只在生长季输出有效模拟值, 而ET则在整年都会输出有效模拟值, 因此本研究将参数优化前后Biome-BGC模型输出的2017年GPP、ET与实验站涡度相关系统实测的GPP、ET进行对比。从日尺度的结果来看(图 1), 参数优化后, Biome-BGC模型输出的GPP与实测值间的平均绝对误差(MAE)和均方根误差(RMSE)均有显著下降, 模型输出的GPP与站点实测GPP间的线性关系更强, 拟合优度(R2)更高, 且二者的线性斜率更接近于1, 可见参数优化显著提高了Biome-BGC模型对GPP的模拟效果。同样的优化效果也表现在对ET的模拟结果中, 优化后Biome-BGC模型输出的ET也与实测值更相近。从图 1的散点对比来看, Biome-BGC模型在GPP和ET的高值区往往会出现低估, 而经过参数优化后的Biome-BGC模型能一定程度的校正这种偏离现象, 使得模拟值与实测值的线性拟合斜率更接近于1∶1线。

|
| 图 1 2017年参数优化前后Biome-BGC模拟的日GPP、ET与站点实测值对比 Fig. 1 Daily GPP and ET simulated by Biome-BGC before and after parameter optimization compared to observed values in 2017 GPP: 总初级生产力Gross primary productivity;ET: 蒸散evapotranspiration |
从月尺度的结果来看(图 2), 参数优化能显著的提高GPP和ET的模拟效果, 极大地降低模拟值与实测值间的平均绝对误差(MAE)和均方根误差(RMSE)。同时, 参数优化后的月尺度GPP、ET与实测值间的线性拟合斜率系数更接近于1, 虽然GPP的拟合优度(R2)只从0.89提高到0.91, ET的拟合优度(R2)从0.85提高到0.93, 但优化后的散点更接近于1:1线, 可知参数优化对模型有极大的校准作用, 提升了模型的精度。从2017年总GPP和ET的结果对比来看, 未参数优化的Biome-BGC模型模拟的年GPP为306.64 gC/m2, 而参数优化后模拟的年GPP为497.62 gC/m2, 实验站涡度相关系统实测的年GPP为482.99 gC/m2, GPP模拟的相对误差由36.5%降低到3.0%。未参数优化的Biome-BGC模型模拟的年ET为263.26 mm, 而参数优化后模拟的ET为397.18 mm, 涡度相关系统实测的年ET为431.20 mm, 参数优化后ET模拟的相对误差由38.9%降低到7.9%。

|
| 图 2 2017年参数优化前后Biome-BGC模拟的月GPP、ET与站点实测值对比 Fig. 2 Monthly GPP and ET simulated by Biome-BGC before and after parameter optimization compared to observed values in 2017 |
为进一步验证参数优化后模型是否具有稳定的普适性, 本节使用实验站涡度相关系统实测的2016—2018年GPP、ET数据, 对使用2017年实测数据优化得出的Biome-BGC模型生理生态参数的普适性效果进行检验。从GPP的模拟结果来看(图 3), Biome-BGC模型能模拟出盐池荒漠草原区人工灌丛生态系统年内GPP的周期性变化, 参数优化前Biome-BGC模型对生长期内GPP的模拟整体偏低, 而经过参数优化后的Biome-BGC模型则显著提高了GPP的模拟值。2016—2018年人工灌丛GPP实测总值为1653.61 gC/m2, Biome-BGC模型原始模拟的GPP总值只有949.30 gC/m2, 但参数优化后, 模拟GPP总值增高到1344.52 gC/m2, 相对误差从42.6%降低到18.7%。从散点对比可以看出, PEST参数优化能显著提高Biome-BGC模拟的效果, 降低模拟值与实测值间的平均绝对误差(MAE)和均方根误差(RMSE)。

|
| 图 3 2016—2018年参数优化前后Biome-BGC模型模拟的日GPP与实测值对比 Fig. 3 Relationship between daily GPP simulated by Biome-BGC model before and after parameter optimization with observed GPP over 2016—2018 |
从ET的模拟结果来看(图 4), PEST参数优化也能改善ET的模拟效果。2016—2018年未经参数优化模拟出的人工灌丛ET总值只有785.99 mm, 但参数优化后, 模拟ET总值增高到1143.66 mm, 更接近1261.33 mm的实测总值, 相对误差从37.7%降低到9.3%。模型模拟的ET与实测值间的均方根误差(RMSE)由2.32 mm/d降低到了1.39 mm/d, 平均绝对误差(MAE)由0.81 mm/d降低到了0.67 mm/d, 模型参数优化后输出的ET与实测ET间的线性关系更强, 拟合优度(R2)更高, 且二者的线性斜率更接近于1。

|
| 图 4 2016—2018年参数优化前后Biome-BGC模型模拟的日ET与实测值对比 Fig. 4 Relationship between daily ET simulated by Biome-BGC model before and after parameter optimization with observed ET over 2016—2018 |
利用优化后的生理生态参数对1986—2018年盐池荒漠草区人工灌丛生态系统长时间序列的GPP和ET进行模拟(图 5)。在近30多年间, 盐池荒漠草原的GPP和ET存在较强的年际波动变化特征。GPP的多年平均值为410.32 g C m-2 a-1, 2018年出现最大值为455.47 gC/m2, 1988年出现最小值为367.45 gC/m2;ET多年平均值为293.28 mm/a, 1996年出现最大值为401.67 mm, 2000年出现最小值141.80 mm。从年际变化特征来看, GPP年际变率较小, 变异系数为0.05, 但在近30余年中表现出显著的上升趋势, 增幅为1.47 g C m-2 a-1;而ET的变异系数则高达0.22, 年际变化率较大, 且无显著变化趋势。从年内的GPP、ET变化特征来看, 盐池荒漠草原区人工灌丛生态系统的GPP和ET在年内呈“单峰”形态变化, 具有显著的季节性规律。GPP和ET均从生长季初期(3月)开始增加, 生长季中期达到年内最大, 生长季结束后(11月)降到低谷, 其中GPP在7月达到峰值79.53 gC/m2, ET在8月达到峰值60.20 mm。虽然人工种植灌丛增强了植物蒸腾量和生态系统的蒸散总量[10], 但对荒漠草原区人工灌丛生态系统而言, 其丛下土壤蒸发依然是占蒸散总量的68.6%[28], 故年内最大ET出现在多年平均降雨量最高的8月份。

|
| 图 5 1986—2018年盐池GPP和ET的年际和年内变化 Fig. 5 GPP and ET interannual and monthly trends in Yanchi County, 1986 to 2018 |
盐池荒漠草原区人工灌丛生态系统的1986—2018年GPP、ET与各个环境气候因子之间的关系如图 6所示。从中可以看出GPP与降水量存在显著正相关关系(r=0.40, P < 0.05), 与气温也存在正相关关系(r=0.49, P < 0.01), 即该区域的人工灌丛的初级生产力大小主要受控于降水和气温, 在降水充沛的条件下, 年平均气温的增高会促进中高纬度地区植被的生产活性, 导致生态系统的GPP升高[29]。GPP与太阳短波辐射存在着显著的负相关关系(r=-0.36, P < 0.05), 即太阳辐射越强, 则会抑制植物光合作用, 减弱总初级生产力, 这与月尺度的GPP与太阳短波辐射关系认识较为一致[30]。ET与降雨的相关性更为显著(r=0.51, P < 0.01), 即半干旱区的人工灌丛生态系统蒸散受降水影响明显, 自然降水的多少决定了年蒸散总量的变化。ET与太阳短波辐射(r=-0.53, P < 0.01)及饱和水汽压差(r=-0.54, P < 0.01)存在着显著的负相关关系, 这表明在光饱和点之后, 强烈的太阳短波辐和较高的饱和水汽压差会抑制干旱区人工灌丛植被的水分交换过程[30]。

|
| 图 6 盐池1986—2018年GPP、ET与环境气候因素的关系 Fig. 6 Relationship between GPP, ET and environmental climate factors in Yanchi County from 1986 to 2018 |
GPP是陆地生态系统碳循环中的重要指标, 它表示植被通过光合作用所固定下来的碳的总量, 能够反映植物群落在自然环境条件下的生产能力[31]。本文通过Biome-BGC模型对荒漠草原区人工灌丛生态系统的GPP进行了模拟, 发现未经参数优化的Biome-BGC模型模拟的GPP结果比实测值偏小, 误差相对较大。当经过PEST参数优化后, GPP的模拟结果逐渐接近实测值, 但依然存在2017年的模拟值略高于实测值、而2016—2018的模拟总值又低于实测值的偏差现象, 即PEST参数优化方法对Biome-BGC模拟GPP的精度有很好的提升作用, 但不同年份的提升效果存在差异, 导致这一现象的原因这可能是未对待优化的参数作敏感性分析所致。已有研究表明, Biome-BGC模型的部分生理生态参数对模型模拟结果影响非常敏感[32], 其取值会对碳水通量的模拟结果造成很大的影响。斯瑶[4]通过傅里叶幅度灵敏度检验扩展法(extended fourier amplitude sensitivity test, EFAST)分析发现叶片和细根的年周转率(LFRT)、最大气孔导度(Gsmax)以及细根和叶片碳的比值(FRC:LC)等参数对GPP的模拟结果具有很大的影响, 其中, LFRT的一阶敏感高达29.80%, 其次是FRC:LC的贡献率为20.50%。而本文仅对28个参数进行整体优化, 未对敏感参数进行甄选, 这将是未来进一步深化研究的方向。其次, 本研究根据盐池荒漠草原人工灌丛的物候期, 人为设置了生长季起始和结束日期, 故模型并未对非生长季节的GPP进行模拟, 而涡度相关系统观测的该生态系统非生长季依然有微弱的碳交换, 这也导致模拟值比实际观测值偏小的原因。因此今后可尝试从删选优化敏感性参数和改进Biome-BGC模拟结构来进一步提高GPP的模拟精度。由于受到最大气孔导度[33]、冠层截留系数和根深等参数设置的直接影响及单层土壤模式、Penman-Monteith公式的适用性[34]和土壤蒸发对模型计算结构的影响, 用模型默认参数对特定生态系统的ET模拟时会产生较大的误差[35]。本研究通过PEST参数优化提升了ET模拟精度, 极大的提高了模拟值, 使其更接近实测ET。但相对于GPP而言, 参数优化后模拟的ET与实测ET在日尺度上的误差依然较大, 其较低的R2可能与Biome-BGC模型简化了土壤水平衡过程、冠层水分截留等有关, 其次可能是对降雨量较低的荒漠草原区的生态系统水分循环细微信息捕捉不够有关。因此, 未来对干旱半干旱区ET模拟效果的提升, 仅仅通过优化Biome-BGC模型参数的方法依然效果有限, 更精确的模拟还需从模型碳水循环计算机理过程上进行优化改进。
3.2 影响荒漠草原区人工灌丛生态系统碳水通量的因素GPP和ET作为指示陆地生态系统碳水循环强弱的重要指标, 其受控因素多样且复杂。对于荒漠草原区人工灌丛生态系统而言, 人为的灌木种植活动是影响其碳水通量的首要原因, 本研究发现, 随着人工灌木演替入侵荒漠草原后, 生态系统的GPP出现显著增加趋势, 碳固持能力持续增强, 这与Du[21]得出的人工灌丛化将荒漠草原区由碳源变为碳汇的认识一致, 即人类活动改变了荒漠草原区生态系统的碳固持趋势。但控制GPP和ET波动的因素则更多的是环境气象因素, 温度在极端高温和极端低温时会抑制植物的光合作用能力, 只有在温度最适时, GPP达到最大值[36];天然降水是干旱半干旱区地表蒸散的主要水分供给来源, 它直接影响地表土壤含水量大小, 进而影响蒸散大小[37]。除了本文2.4节提到的降水量、气温、太阳短波辐射和饱和水汽压差等4种因素外, CO2浓度[38]、土壤含水量及土壤温度[39]等环境因素也会对GPP和ET产生影响。康满春[8]等利用Biome-BGC模型对CO2浓度增加情景下的杨树人工林GPP进行模拟, 发现GPP对大气CO2浓度上升的响应程度(27.7%—43.6%)远高于对气温上升(1.2%—5.8%)和降水变化(1.2%—3.5%)的响应, 原因可能是由于CO2浓度升高一方面对植物起到“施肥”作用, 另一方面通过影响植物对温度、水分及营养物质需求等方面来作用于植物生长, 从而提升植物光合效率。而土壤含水量成为引起亚热带森林GPPmax年际变化的主要控制因素, 其可能是通过调节温度变暖来影响植被生产力[40]。因此, 在今后的研究中要加强多环境因素对荒漠草原区人工灌丛生态系统碳水通量的影响分析, 以便为荒漠草原退化生态系统修复与治理提供更为可靠的理论支撑和决策依据。
3.3 模型-数据融合方法近年来, 通过数据融合模型来提高模型对碳水通量的模拟效果已经成为陆地生态系统研究碳水循环过程的热点话题之一, 其主要包括数据同化和参数估计两类[41], 本研究用到的PEST模型优化参数便属于后者。PEST仅生成Biome-BGC模型参数对应的一个最佳估计值, 并不改变模型内部结构, 但如若不改动Biome-BGC模型, 则很难优化模型结构中随生态过程而产生动态变化的状态变量。相对而言, 数据同化方法则需要编程改进模型, 将观测数据融合进模型中, 这样不仅修正了模拟的状态变量, 还优化了参数[41]。因此, 后续研究可通过改动模型结构, 将模型与观测数据融合, 或者将数据同化和参数估计这两种方法结合使用, 进一步提高模型估算碳水通量的准确性, 为研究陆地生态系统碳水循环过程提供新的方法与途径。
4 结论本文针对如何高效模拟荒漠草原区人工灌丛生态系统碳水循环这一个科学问题, 集合PEST模型与涡度相关观测数据来优化Biome-BGC模型的生理生态参数, 进而模拟宁夏盐池荒漠草原区人工灌丛生态系统的GPP和ET, 对参数优化后的模拟效果进行了评价, 并分析了研究区种植人工灌丛以来的碳水循环变化规律, 得出如下结论:(1)PEST参数优化后可提高Biome-BGC模型模拟GPP和ET的精度, 可改善模型对荒漠草原区人工灌丛生态系统碳水循环的模拟效果;(2)经PEST模型优化后的模型参数具有较强的普适性, 可以推广应用于荒漠草原区人工灌丛生态系统长时间序列的GPP和ET模拟中去;(3)宁夏盐池县杨寨子村自1986年种植人工灌木林以来, 生态系统的GPP出现显著增加趋势, 但ET年际变化率较大, 且无显著变化趋势。
| [1] |
Barnes M L, Farella M M, Scott R L, Moore D J P, Ponce-Campos G E, Biederman J A, MacBean N, Litvak M E, Breshears D D. Improved dryland carbon flux predictions with explicit consideration of water-carbon coupling. Communications Earth & Environment, 2021, 2(1): 248. |
| [2] |
Zhang Y, Xiao X M, Guanter L, Zhou S, Ciais P, Joiner J, Sitch S, Wu X C, Nabel J, Dong J W, Kato E, Jain A K, Wiltshire A, Stocker B D. Precipitation and carbon-water coupling jointly control the interannual variability of global land gross primary production. Scientific Reports, 2016, 6(1): 39748. DOI:10.1038/srep39748 |
| [3] |
刘晓文. 陆地生态系统过程模型参数估计及其在碳水通量模拟中的研究——以CEVSA模型为例[D]. 甘肃: 兰州大学, 2020: 93.
|
| [4] |
斯瑶, 张振振. 不同气象条件下Biome-BGC模型碳通量模拟精度评价. 三峡生态环境监测, 2019, 4(3): 59-67. |
| [5] |
张廷龙, 孙睿, 胡波, 冯丽超. 利用模拟退火算法优化Biome-BGC模型参数. 生态学杂志, 2011, 30(2): 408-414. |
| [6] |
Thornton P E, Law B E, Gholz H L, Clark K L, Falge E, Ellsworth D S, Golstein A H, Monson R K, Hollinger D, Falk M, Chen J, Sparks J P. Modeling and measuring the effects of disturbance history and climate on carbon and water budgets in evergreen needleleaf forests. Agricultural and Forest Meteorology, 2002, 113(1/4): 185-222. |
| [7] |
Chiesi M, Chirici G, Corona P, Duce P, Salvati R, Spano D, Vaccari F P, Maselli F. Use of BIOME-BGC to simulate water and carbon fluxes within Mediterranean macchia. iForest-Biogeosciences and Forestry, 2012, 5(2): 38-43. |
| [8] |
康满春, 朱丽平, 许行, 查同刚, 张志强. 基于Biome-BGC模型的北方杨树人工林碳水通量对气候变化的响应研究. 生态学报, 2019, 39(7): 2378-2390. |
| [9] |
张越, 刘康, 张红娟, 张丹丹, 陈慕亚. 基于BIOME-BGC模型的秦岭北坡太白红杉林碳源/汇动态和趋势研究. 热带亚热带植物学报, 2019, 27(3): 235-249. |
| [10] |
丹杨, 杜灵通, 王乐, 马龙龙, 乔成龙, 吴宏玥, 孟晨. 荒漠草原人工灌丛化对蒸散发及其组分的影响——以盐池县为例. 生态学报, 2020, 40(16): 5638-5648. |
| [11] |
王乐, 杜灵通, 马龙龙, 丹杨. 人工灌丛化对荒漠草原生态系统碳储量的影响. 生态学报, 2022, 42(1): 246-254. |
| [12] |
樊华烨, 李英, 张廷龙, 高焕霖, 呼帅. 陆地植被水碳通量模型模拟与数据同化研究进展. 应用生态学报, 2020, 31(6): 2098-2108. |
| [13] |
Ma X L, Huete A, Cleverly J, Eamus D, Chevallier F, Joiner J, Poulter B, Zhang Y G, Guanter L, Meyer W, Xie Z Y, Ponce-Campos G. Drought rapidly diminishes the large net CO2 uptake in 2011 over semi-arid Australia. Scientific Reports, 2016, 6(1): 37747. DOI:10.1038/srep37747 |
| [14] |
Mao F J, Li P H, Zhou G M, Du H Q, Xu X J, Shi Y J, Mo L F, Zhou Y F, Tu G Q. Development of the BIOME-BGC model for the simulation of managed Moso bamboo forest ecosystems. Journal of Environmental Management, 2016, 172: 29-39. |
| [15] |
郑磊, 宋世凯, 袁秀亮, 董嘉琪, 李龙辉. 基于Biome-BGC模型和集合卡尔曼滤波方法的阔叶红松林生态系统水碳通量模拟. 生态学杂志, 2017, 36(6): 1752-1760. |
| [16] |
刘丽慧, 孙皓, 李传华. 基于改进土壤冻融水循环的Biome-BGC模型估算青藏高原草地NPP. 地理研究, 2021, 40(5): 1253-1264. |
| [17] |
梁浩, 胡克林, 李保国. 基于PEST的土壤-作物系统模型参数优化及灵敏度分析. 农业工程学报, 2016, 32(3): 78-85. |
| [18] |
Nolan B T, Puckett L J, Ma L W, Green C T, Bayless E R, Malone R W. Predicting unsaturated zone nitrogen mass balances in agricultural settings of the United States. Journal of Environmental Quality, 2010, 39(3): 1051-1065. |
| [19] |
温永斌, 韩海荣, 程小琴, 李祖政. 基于Biome-BGC模型的千烟洲森林水分利用效率研究. 北京林业大学学报, 2019, 41(4): 69-77. |
| [20] |
温永斌, 韩海荣, 程小琴, 李祖政. 不同幅度景观格局与水分利用效率耦合研究——以千烟洲为例. 北京林业大学学报, 2019, 41(12): 88-95. |
| [21] |
Du L T, Zeng Y J, Ma L L, Qiao C L, Wu H Y, Su Z B, Bao G. Effects of anthropogenic revegetation on the water and carbon cycles of a desert steppe ecosystem. Agricultural and Forest Meteorology, 2021, 300: 108339. |
| [22] |
郑琪琪, 杜灵通, 宫菲, 丹杨, 王乐. 基于GF-1遥感影像的宁夏盐池柠条人工林景观特征研究. 西南林业大学学报, 2019, 39(1): 152-159. |
| [23] |
White M A, Thornton P E, Running S W, Nemani R R. Parameterization and sensitivity analysis of the BIOME-BGC terrestrial ecosystem model: net primary production controls. Earth Interactions, 2000, 4(3): 1-85. |
| [24] |
Yu X, Lamačová A, Duffy C, Krám P, Hruška J. Hydrological model uncertainty due to spatial evapotranspiration estimation methods. Computers & Geosciences, 2016, 90: 90-101. |
| [25] |
Iskra I, Droste R. Application of non-linear automatic optimization techniques for calibration of HSPF. Water Environment Research, 2007, 79(6): 647-659. |
| [26] |
Skahill B E, Baggett J S, Frankenstein S, Downer C W. More efficient PEST compatible model independent model calibration. Environmental Modelling & Software, 2009, 24(4): 517-529. |
| [27] |
王乐. 气候变化和人工灌丛对荒漠草原生态系统碳循环的影响[D]. 银川: 宁夏大学, 2020: 64.
|
| [28] |
马龙龙, 杜灵通, 丹杨, 王乐, 乔成龙, 吴宏玥. 基于茎流-蒸渗仪法的荒漠草原带人工灌丛群落蒸散特征. 植物生态学报, 2020, 44(8): 807-818. |
| [29] |
Wang Z Q, Chang J F, Peng S S, Piao S L, Ciais P, Betts R. Changes in productivity and carbon storage of grasslands in China under future global warming scenarios of 1.5℃ and 2℃. Journal of Plant Ecology, 2019, 12(5): 804-814. |
| [30] |
Du L T, Ma L L, Pan H Z, Qiao C L, Meng C, Wu H Y, Tian J, Yuan H Y. Carbon-water coupling and its relationship with environmental and biological factors in a planted Caragana liouana shrub community in desert steppe, northwest China. Journal of Plant Ecology, 2022, 15(5): 947-960. |
| [31] |
Beer C, Reichstein M, Tomelleri E, Ciais P, Jung M, Carvalhais N, Rödenbeck C, Arain M A, Baldocchi D, Bonan G B, Bondeau A, Cescatti A, Lasslop G, Lindroth A, Lomas M, Luyssaert S, Margolis H, Oleson K W, Roupsard O, Veenendaal E, Viovy N, Williams C, Woodward F I, Papale D. Terrestrial gross carbon dioxide uptake: global distribution and covariation with climate. Science, 2010, 329(5993): 834-838. |
| [32] |
刘秋雨, 张廷龙, 孙睿, 王博闻, 叶欣欣, 李一哲. Biome-BGC模型参数的敏感性和时间异质性. 生态学杂志, 2017, 36(3): 869-877. |
| [33] |
Sándor R, Barcza Z, Hidy D, Lellei-Kovács E, Ma S, Bellocchi G. Modelling of grassland fluxes in Europe: evaluation of two biogeochemical models. Agriculture Ecosystems & Environment, 2016, 215: 1-19. |
| [34] |
张廷龙, 孙睿, 胡波, 冯丽超, 张荣华. 改进Biome-BGC模型模拟哈佛森林地区水、碳通量. 生态学杂志, 2011, 30(9): 8. |
| [35] |
Ueyama M, Ichii K, Hirata R, Takagi K, Asanuma J, Machimura T, Nakai Y, Ohta T, Saigusa N, Takahashi Y, Hirano T. Simulating carbon and water cycles of larch forests in East Asia by the BIOME-BGC model with AsiaFlux data. Biogeosciences, 2010, 7(3): 959-977. |
| [36] |
Lin S R, Li J, Liu Q H, Gioli B, Paul-Limoges E, Buchmann N, Gharun M, Hörtnagl L, Foltýnová L, Dušek J, Li L H, Yuan W P. Improved global estimations of gross primary productivity of natural vegetation types by incorporating plant functional type. International Journal of Applied Earth Observation and Geoinformation, 2021, 100: 102328. |
| [37] |
杨秀芹, 王国杰, 潘欣, 张余庆. 基于GLEAM遥感模型的中国1980-2011年地表蒸散发时空变化. 农业工程学报, 2015, 31(21): 132-141. |
| [38] |
Qiu R N, Han G, Ma X, Sha Z Y, Shi T Q, Xu H, Zhang M. CO2 concentration, a critical factor influencing the relationship between solar-induced chlorophyll fluorescence and gross primary productivity. Remote Sensing, 2020, 12(9): 1377. |
| [39] |
姜鹏, 秦美欧, 李荣平, 孟莹, 杨霏云, 温日红, 孙沛, 方缘. 中国典型生态系统GPP的季节变异及其影响要素. 生态环境学报, 2022, 31(4): 643-651. |
| [40] |
石旭霞, 宋沼鹏, 侯继华, 张雷明, 牛书丽, 王安志, 项文化, 王辉民. 中国东部森林最大总初级生产力的时空分布特征及其影响因子. 生态学杂志, 2019, 38(7): 1949-1961. |
| [41] |
李新, 马瀚青, 冉有华, 王旭峰, 朱高峰, 刘丰, 何洪林, 张臻, 黄春林. 陆地碳循环模型-数据融合: 前沿与挑战. 中国科学: 地球科学, 2021, 51(10): 1650-1663. |