 2023, Vol. 43
2023, Vol. 43文章信息
- 赵小雨, 蔡立哲, 饶义勇, 杨德援, 林靖翔, 陈昕韡, 傅素晶, 周细平
- ZHAO Xiaoyu, CAI Lizhe, RAO Yiyong, YANG Deyuan, LIN Jingxiang, CHEN Xinwei, FU Sujing, ZHOU Xiping
- 考洲洋人工种植红树林湿地大型底栖动物群落环境响应
- Environmental response of macrofaunal community in planted mangrove wetlands in Kaozhou Bay, China
- 生态学报. 2023, 43(13): 5505-5516
- Acta Ecologica Sinica. 2023, 43(13): 5505-5516
- http://dx.doi.org/10.5846/stxb202206141693
-
文章历史
- 收稿日期: 2022-06-14
- 网络出版日期: 2023-03-07





2. 厦门大学嘉庚学院, 漳州 363105;
3. 中国水产科学研究院南海水产研究所, 广州 510300;
4. 自然资源部第三海洋研究所, 厦门 361000
2. Xiamen University Tan Kah Kee College, Zhangzhou 363105, China;
3. South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou 510300, China;
4. The Third Institute of Oceanography, Ministry of Natural Resources, Xiamen 361000, China
红树林湿地是热带和亚热带地区沿海潮间带重要的生态系统, 可为人类生产生活提供多种生态服务功能[1]。随着人口的快速增长和城市化的发展, 在过去几十年里, 大约三分之一的红树林已经退化[2]。为此, 全球范围内在广泛开展红树林造林计划, 以期恢复退化的红树林并扩大红树林面积[3]。大型底栖动物是红树林生态系统的关键组成部分, 在连接初级生产者和顶级捕食者方面发挥着重要作用[4], 可以增强沉积物孔隙度, 驱动沉积物再造、养分循环和有机质分解等重要过程[5—7]。红树林植被的变化会带来生境的异质性进而影响大型底栖动物的分布和生存[8—9], 大型底栖动物群落结构的变化又会影响当地生物多样性和生态系统功能[10—11], 因此, 研究红树林湿地大型底栖动物群落结构特征具有重要的生态学意义[12—13]。
考洲洋位于惠东县南部, 具有极为丰富的滩涂资源以及优越的水域条件。近年来由于考洲洋海域的水产养殖密度逐渐上升, 加之其周边是重要的皮革产区, 大量污染物质被排入湾内, 导致海水环境质量不断降低[14], 对考洲洋生物群落组成带来了负面影响[15]。2017年开始实施的考洲洋—罂公洲至赤岸区域海岸带整治及生态修复工程项目, 其主要内容就是种植红树, 计划完成红树种植133hm2, 种植红树800万株[16]。截止到2018年4月, 完成红树育苗750万株, 种植红树500万株[17]。红树林生态修复工程有效地改善了考洲洋的海水质量[18], 为鱼类和底栖动物提供了重要庇护和摄食场所。目前关于考洲洋的研究主要集中在水质条件[19]、沉积物[20]、重金属污染物[21]、有机污染物[22]等环境质量监测, 以及红树林植被调查[23]和渔业资源调查[24], 但是对考洲洋红树林湿地大型底栖动物时空变化了解较少, 因此有必要对该地区红树林湿地大型底栖动物群落结构进行分析。本文于2018年和2019年在考洲洋盐洲大桥附近两处人工种植红树林湿地开展大型底栖动物调查研究, 探讨考洲洋人工种植红树林湿地大型底栖动物对环境的响应, 以期为红树林生态系统功能恢复和生物多样性保护提供理论依据。
1 材料与方法 1.1 研究区域与样品处理方法盐洲大桥北起惠州市惠东黄埠镇望京洲村, 南连盐洲联新村李甲海堤, 是惠东县2008年十大重点建设项目之一, 盐洲大桥及引道全长3.899 km。本研究于2018年1月(冬季)、2018年7月(夏季)、2019年4月(春季)和2019年11月(秋季), 对新吉盐公路与盐洲大桥交叉处附近的人工种植红树林湿地(X断面和Y断面)林内、林缘和光滩共6个取样点进行了4个季节的大型底栖动物生态定量调查(图 1)。

|
| 图 1 考洲洋盐洲大桥附近人工种植红树林湿地大型底栖动物调查断面布设示意图 Fig. 1 Sampling sites of macrofauna in planted mangrove wetlands near Yanzhou Bridge in Kaozhou Bay |
大型底栖动物样品定量采集选择在大潮期间, 在盐洲大桥两侧红树林湿地设置2条断面, 每条断面各3个取样点, 分别位于红树林林内(XH和YH取样点, 潮高180 cm左右, 参考大亚湾潮汐表, 下同)、林缘(XM和YM取样点, 潮高130 cm左右)和光滩(XL和YL取样点, 潮高80 cm左右);每个取样点设置5个平行样框, 样框面积为25 cm×25 cm, 深度30 cm。采样时, X断面XH取样点红树林种植1—2年, Y断面YH取样点为红树林种植5—6年。每个样框采集到的沉积物于塑料桶中进行淘洗, 经孔径为0.5 mm的筛网过筛分选, 将筛网截留的生物和残渣一起装在标本瓶中, 加入5%中性甲醛溶液进行固定和保存, 贴好标签。样本带回实验室后, 再次经0.5 mm筛网进行分选, 并在解剖镜下镜检分离生物和杂质, 然后对大型底栖动物进行鉴定分类、计数和称重(感重0.0001 g)。样本采集、保存和分选鉴定均按照《海洋调查规范》第6部分:海洋生物调查指南(GB/T 12763.6—2007)和《海洋监测规范》第7部分:近海污染生态调查和生物监测(GB/T 17378.7—2007)进行。
1.2 数据处理与分析本研究中大型底栖动物物种数采用两种计算方式, 即统计值和平均值, 如某断面的物种数统计值是该断面各取样点获得的所有物种数, 而某断面的物种数平均值是该断面各取样点物种数的平均值[25]。本研究采用Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)和Marglef丰富度指数(d)衡量大型底栖动物群落多样性水平。计算公式为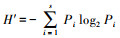
2018—2019年的四个季节(冬季、夏季、春季和秋季)在X和Y断面共获得大型底栖动物48种, 其中X断面共获得30种, Y断面共获得39种。在X断面, 光滩(XL)大型底栖动物物种数最多, 为23种, 其次是林内(XH), 19种, 林缘(XM)最少, 18种;在Y断面, 也是光滩(YL)大型底栖动物物种数(30种)最多, 其次是林内(YH)26种, 林缘(YM)最少, 25种。Y断面红树林三种生境的大型底栖动物物种数均分别高于X断面三种生境的物种数。X断面和Y断面大型底栖动物物种数均是夏季最多, 分别为18种和28种, 秋季最少, 分别为10种和15种(图 2)。

|
| 图 2 考洲洋人工种植红树林湿地大型底栖动物物种数、栖息密度和生物量的时空变化 Fig. 2 Temporal and spatial variations of macrofaunal species numbers, density and biomass in planted mangrove wetlands in Kaozhou Bay H: 林内 Inside mangrove;M: 林缘 Mangrove margin;L: 光滩 Bare flat |
大型底栖动物物种数平均值的时空分布与统计值的时空分布不尽一致。Y断面统计值空间分布是光滩(30种)>林内(26种)>林缘(25种), 而平均值是光滩(8种)>林缘(6种)>林内(5种)。X断面统计值季节分布是夏>春>冬>秋, 而平均值是夏>秋>冬>春;Y断面统计值季节分布是夏>冬>春>秋, 而平均值是冬>夏>春>秋(图 2)。
在X断面的大型底栖动物平均栖息密度, 从不同生境看, 光滩最高, 林内第二, 林缘最低, 分别是1278个/m2、1221个/m2和721个/m2;从季节上看, 冬季最高, 夏季次之, 春季第三, 秋季最低, 分别是2803个/m2、635个/m2、507个/m2和349个/m2。在Y断面的大型底栖动物平均栖息密度, 从不同生境看, 光滩最高, 林缘第二, 林内最低, 分别是2008个/m2、720个/m2和478个/m2;从季节上看, 冬季最高, 夏季次之, 春季第三, 秋季最低, 分别是1739个/m2、1639个/m2、759个/m2和139个/m2。可见, X和Y断面大型底栖动物平均栖息密度的空间变化略有不同, X断面是光滩>林内>林缘, Y断面是光滩>林缘>林内, 但两条断面的季节变化是一致的, 均是冬>夏>春>秋(图 2)。
在X断面的大型底栖动物平均生物量, 从不同生境看, 林内最高, 林缘次之, 光滩最低, 分别是46.925 g/m2、33.285 g/m2和25.614 g/m2;从季节上看, 春季最高, 夏季次之, 秋季第三, 冬季最低, 分别是54.994 g/m2、37.669 g/m2、29.378 g/m2和19.058 g/m2。在Y断面的大型底栖动物平均生物量, 从不同生境看, 林缘最高, 光滩第二, 林内最低, 分别是52.058 g/m2、49.481 g/m2和35.090 g/m2;从季节上看, 秋季最高, 冬季次之, 春季第三, 夏季最低, 分别是85.251 g/m2、36.866 g/m2、32.925 g/m2和27.130 g/m2。可见, X和Y断面大型底栖动物平均生物量的空间变化明显不同, X断面是林内>林缘>光滩, Y断面则是林缘>光滩>林内, 两条断面的季节变化也明显不同, X断面是春>夏>秋>冬, Y断面是秋>冬>春>夏(图 2)。
2.2 考洲洋人工种植红树林湿地大型底栖动物多样性指数、均匀度指数和丰富度指数在X断面的大型底栖动物群落平均多样性指数(H′), 从不同生境看, 林内最高, 林缘第二, 光滩最低, 分别是1.535、1.369和1.307;从季节上看, 夏季最高, 秋季次之, 冬季第三, 春季最低, 分别是1.757、1.658、1.128和1.072。在Y断面的大型底栖动物群落平均多样性指数(H′), 从不同生境看, 林内最高, 林缘第二, 光滩最低, 分别是1.772、1.771和1.690;从季节上看, 冬季最高, 秋季次之, 夏季第三, 春季最低, 分别是1.980、1.805、1.660和1.407(图 3)。

|
| 图 3 考洲洋人工种植红树林湿地大型底栖动物多样性指数(H′)、均匀度指数(J) 和丰富度指数(d) 的时空变化 Fig. 3 Temporal and spatial variations of macrofaunal diversity index (H′), evenness index (J) and species richness index (d) in planted mangrove wetlands in Kaozhou Bay |
在X断面的大型底栖动物群落平均均匀度指数(J), 从不同生境看, 林缘最高, 林内第二, 光滩最低, 分别是0.732、0.729和0.600;从季节上看, 秋季最高, 夏季次之, 春季第三, 冬季最低, 分别是0.814、0.708、0.643和0.585。在Y断面的大型底栖动物群落平均丰富度指数(J), 从不同生境看, 林内最高, 林缘第二, 光滩最低, 分别是0.762、0.680和0.607;从季节上看, 秋季最高, 冬季次之, 春季第三, 夏季最低, 分别是0.887、0.654、0.610和0.573(图 3)。
在X断面的大型底栖动物群落平均丰富度指数(d), 从不同生境看, 林内最高, 光滩第二, 林缘最低, 分别是0.584、0.579和0.456;从季节上看, 夏季最高, 秋季次之, 春季第三, 冬季最低, 分别是0.769、0.569、0.421和0.398。在Y断面的大型底栖动物群落平均丰富度指数(d), 从不同生境看, 光滩最高, 林缘第二, 林内最低, 分别是0.990、0.818和0.779;从季节上看, 冬季最高, 夏季次之, 春季第三, 秋季最低, 分别是1.122、0.948、0.669和0.663(图 3)。
综合上述结果可见, X和Y断面大型底栖动物平均多样性指数(H′)、均匀度指数(J)和丰富度指数(d)的空间变化略有不同, 两条断面三种指数的季节变化明显不同。
2.3 考洲洋人工种植红树林湿地大型底栖动物群落的方差分析双因素方差分析表明, 大型底栖动物物种数的6组对比数据, 即两断面(X和Y断面)、X和Y断面林内(XH和YH取样点)、林缘(XM和YM取样点)、光滩(XL和YL取样点)、X断面之间的取样点(XH、XM和XL)和Y断面之间的取样点(YH、YM和YL)的大型底栖动物物种数均有显著季节差异(P < 0.05), 且均有季节×断面/取样点显著差异(P < 0.05);大型底栖动物物种数的空间差异除了在两林内之间不显著外, 其余5组对比数据存在显著空间差异(P < 0.05)(表 1)。
| 参数 Parameters |
对比 Comparison |
季节 Season |
断面/取样点 Transects/Sites |
季节×断面/取样点 Season×Transects/Sites |
||||||||
| df | F | P | df | F | P | df | F | P | ||||
| 物种数 | 两断面 | 3 | 15.727 | < 0.001* | 1 | 35.509 | < 0.001* | 3 | 8.994 | < 0.001* | ||
| Species number | 两林内 | 3 | 10.827 | < 0.001* | 1 | 4.134 | 0.051 | 3 | 11.027 | < 0.001* | ||
| 两林缘 | 3 | 13.926 | < 0.001* | 1 | 27.348 | < 0.001* | 3 | 6.812 | 0.001* | |||
| 两光滩 | 3 | 21.199 | < 0.001* | 1 | 70.281 | < 0.001* | 3 | 31.498 | < 0.001* | |||
| X断面 | 3 | 12.697 | < 0.001* | 2 | 4.885 | 0.012* | 6 | 8.235 | < 0.001* | |||
| Y断面 | 3 | 41.466 | < 0.001* | 2 | 20.163 | < 0.001* | 6 | 9.845 | < 0.001* | |||
| 栖息密度 | 两断面 | 3 | 24.818 | < 0.001* | 1 | 0.001 | 0.980 | 3 | 6.054 | 0.001* | ||
| Density | 两林内 | 3 | 23.017 | < 0.001* | 1 | 23.748 | < 0.001* | 3 | 31.424 | < 0.001* | ||
| 两林缘 | 3 | 66.420 | < 0.001* | 1 | 0.000 | 0.989 | 3 | 15.913 | < 0.001* | |||
| 两光滩 | 3 | 19.470 | < 0.001* | 1 | 5.422 | 0.027* | 3 | 4.321 | 0.012* | |||
| X断面 | 3 | 38.435 | < 0.001* | 2 | 3.587 | 0.035* | 6 | 1.732 | 0.134 | |||
| Y断面 | 3 | 29.966 | < 0.001* | 2 | 44.476 | < 0.001* | 6 | 14.794 | < 0.001* | |||
| 生物量 | 两断面 | 3 | 4.398 | 0.006* | 1 | 2.614 | 0.109 | 3 | 7.448 | < 0.001 | ||
| Biomass | 两林内 | 3 | 5.824 | 0.003* | 1 | 1.360 | 0.252 | 3 | 4.044 | 0.015* | ||
| 两林缘 | 3 | 1.327 | 0.283 | 1 | 2.747 | 0.108 | 3 | 5.143 | 0.005* | |||
| 两光滩 | 3 | 2.160 | 0.113 | 1 | 7.983 | 0.008* | 3 | 5.431 | 0.004* | |||
| X断面 | 3 | 5.803 | 0.002* | 2 | 3.906 | 0.027* | 6 | 4.051 | 0.002* | |||
| Y断面 | 3 | 7.640 | < 0.001* | 2 | 1.136 | 0.330 | 6 | 1.655 | 0.154 | |||
| H′ | 两断面 | 3 | 7.370 | < 0.001* | 1 | 14.381 | < 0.001* | 3 | 6.277 | 0.001* | ||
| 两林内 | 3 | 3.798 | 0.020* | 1 | 3.941 | 0.056 | 3 | 5.371 | 0.004* | |||
| 两林缘 | 3 | 3.350 | 0.031* | 1 | 4.628 | 0.039* | 3 | 1.487 | 0.237 | |||
| 两光滩 | 3 | 3.858 | 0.019* | 1 | 8.959 | 0.005* | 3 | 9.345 | < 0.001* | |||
| X断面 | 3 | 11.542 | < 0.001* | 2 | 1.705 | 0.193 | 6 | 2.368 | 0.044* | |||
| Y断面 | 3 | 4.679 | 0.006* | 2 | 0.367 | 0.695 | 6 | 2.810 | 0.021* | |||
| J | 两断面 | 3 | 14.910 | <0.001* | 1 | 0.043 | 0.835 | 3 | 2.950 | 0.036* | ||
| 两林内 | 3 | 2.314 | 0.095* | 1 | 0.320 | 0.576 | 3 | 1.566 | 0.217 | |||
| 两林缘 | 3 | 5.069 | 0.006* | 1 | 1.067 | 0.310 | 3 | 0.905 | 0.450 | |||
| 两光滩 | 3 | 16.672 | < 0.001* | 1 | 0.000 | 0.994 | 3 | 3.390 | 0.030* | |||
| X断面 | 3 | 5.219 | 0.003* | 2 | 4.065 | 0.023* | 6 | 0.865 | 0.527 | |||
| Y断面 | 3 | 20.877 | < 0.001* | 2 | 8.496 | 0.001* | 6 | 1.291 | 0.281 | |||
| d | 两断面 | 3 | 9.822 | < 0.001* | 1 | 48.291 | < 0.001* | 3 | 10.251 | < 0.001* | ||
| 两林内 | 3 | 9.568 | < 0.001* | 1 | 14.272 | 0.001* | 3 | 9.421 | < 0.001* | |||
| 两林缘 | 3 | 6.902 | 0.001* | 1 | 28.039 | < 0.001* | 3 | 6.565 | 0.001* | |||
| 两光滩 | 3 | 7.804 | 0.001* | 1 | 50.564 | < 0.001* | 3 | 26.010 | < 0.001* | |||
| X断面 | 3 | 14.992 | < 0.001* | 2 | 3.608 | 0.035* | 6 | 7.140 | < 0.001* | |||
| Y断面 | 3 | 20.912 | < 0.001* | 2 | 7.412 | 0.002* | 6 | 6.916 | < 0.001* | |||
| H′:多样性指数 Diversity index;J:均匀度指数 Evenness index;d:丰富度指数 Richness index; *表示因子间具有显著差异(P < 0.05) | ||||||||||||
大型底栖动物栖息密度的6组对比数据, 即X和Y断面、两林内、两林缘、两光滩、X断面之间的取样点和Y断面之间的取样点的大型底栖动物栖息密度均有显著季节差异(P < 0.05);4组对比数据, 即两林内、两光滩、X和Y断面之间的取样点的大型底栖动物栖息密度存在显著空间差异(P < 0.05);5组对比数据, 即两断面、两林内、两林缘、两光滩和Y断面之间的取样点的大型底栖动物栖息密度存在季节×断面/取样点显著差异(P < 0.05)(表 1)。
大型底栖动物生物量的4组对比数据, 即X和Y断面、两林内、X断面之间的取样点和Y断面之间的取样点的大型底栖动物生物量有显著季节差异(P < 0.05);2组对比数据, 即两光滩和X断面之间的取样点的大型底栖动物生物量存在显著空间差异(P < 0.05);5组对比数据, 即X和Y断面、林内、林缘、光滩和X断面之间的取样点的大型底栖动物生物量存在季节×断面/取样点显著差异(P < 0.05)(表 1)。
多样性指数(H′)的6组对比数据, 即X和Y断面、两林内、两林缘、两光滩、X断面之间的取样点和Y断面之间的取样点的H′均有显著季节差异(P < 0.05);3组对比数据, 即两断面、两林缘和两光滩的大型底栖动物H′存在显著空间差异(P < 0.05);5组对比数据, 即两断面、两林内、两光滩、X和Y断面之间的取样点的大型底栖动物H′存在季节×断面/取样点显著差异(P < 0.05)(表 1)。
大型底栖动物均匀度指数(J)的6组对比数据, 即X和Y断面、两林内、两林缘、两光滩、X断面之间的取样点和Y断面之间的取样点的J均有显著季节差异(P < 0.05);2组对比数据, 即X断面之间的取样点和Y断面之间的取样点的大型底栖动物J存在显著空间差异(P < 0.05);2组对比数据, 即两断面和两光滩的大型底栖动物J存在季节×断面/取样点显著差异(P < 0.05)(表 1)。
大型底栖动物丰富度指数(d)的6组对比数据, 即两断面、两林内、两林缘、两光滩、X断面之间的取样点和Y断面之间的取样点的d均有显著季节差异(P < 0.05)、存在显著空间差异(P < 0.05)和存在季节×断面/取样点显著差异(P < 0.05)(表 1)。
综合上述6种参数的双因素方差分析结果得出, 两断面、两林缘和两光滩的大型底栖动物物种数、多样性指数(H′)和丰富度指数(d)均有显著空间差异。
2.4 考洲洋人工种植红树林湿地大型底栖动物群落的聚类和nMDS分析聚类分析表明, 冬季在相似度40%水平上大型底栖动物群落可以分为2组群落, 除了Y断面林内为一个群落外, 其余Y断面林缘和光滩, 以及X断面的林内、林缘和光滩为另一个群落;SIMPER分析结果显示, YH群落主要贡献种为腺带刺沙蚕(Neanthes glandicincta)和蜾蠃蜚(Corophium sp.), 贡献率分别为41.31%和15.16%, 后者群落主要贡献种为腺带刺沙蚕(77.89%)。夏季在相似度45%水平上大型底栖动物群落可以分为2组群落, 群落1为X断面的林内、林缘和光滩, 主要贡献种为腺带刺沙蚕和持真节虫(Euclymene annandalei), 贡献率分别为54.67%和25.58%;群落2为Y断面的林内、林缘和光滩, 主要贡献种为腺带刺沙蚕(66.04%)。春季在相似度25%水平上大型底栖动物群落可以分为2组群落, 除了X断面林内为一个群落外, 其余X断面林缘和光滩, 以及Y断面的林内、林缘和光滩为另一个群落;XH群落主要贡献种为羽须鳃沙蚕(Dendronereis pinnaticirris)和日本拟背尾水虱(Paranthura japonica), 贡献率分别为54.89%和23.72%;后者群落主要贡献种为腺带刺沙蚕(64.23%)。秋季在相似度35%水平上大型底栖动物群落可以分为2组群落, 除了Y断面林内为一个群落外, 其余Y断面林缘和光滩, 以及X断面的林内、林缘和光滩为另一个群落;YH群落主要贡献种为持真节虫(55.44%)和腺带刺沙蚕(36.73%), 后者群落主要贡献种为小头虫(Capitella sp.), 贡献率为71.58%(图 4)。nMDS结果与Cluster聚类分析结果一致(图 5)。

|
| 图 4 考洲洋人工种植红树林湿地大型底栖动物群落的Cluster分析 Fig. 4 Cluster analysis of macrofaunal community in planted mangrove wetlands in Kaozhou Bay |

|
| 图 5 考洲洋人工种植红树林湿地大型底栖动物群落的非参数多变量排序图 Fig. 5 Nonmetric multidimensional scaling (nMDS) analysis of macrofaunal community in planted mangrove wetlands in Kaozhou Bay |
冬季考洲洋盐洲大桥附近红树林湿地Y断面的大型底栖动物有明显的空间分布趋势。Y断面的潮高是YH>YM>YL(大约180 cm>130 cm>80 cm), 冬季大型底栖动物物种数、栖息密度、生物量、H′和d均是随着潮高降低而增加, 即YH < YM < YL。Y断面夏季大型底栖动物栖息密度随着潮高降低而增加, 但多样性指数和丰富度指数随着潮高降低而降低。2019年大亚湾冬季和夏季潮差较大, 冬季潮差在187—236 cm之间, 夏季潮差在186—216 cm之间, 春季潮差在121—158 cm之间, 秋季潮差在144—154 cm之间[27], 2020年大亚湾也是冬季和夏季潮差较大, 春季和秋季潮差较小[28](表 2)。可见, 红树林湿地大型底栖动物群落在冬季和夏季有明显空间分布趋势, 而在春季和秋季没有明显空间分布趋势, 与冬季和夏季潮差较大, 春季和秋季潮差较小有关。考洲洋邻近大亚湾, 潮汐相似, 但考洲洋仅靠一条狭长的水道与外海相通, 潮差可能更小。潮差大, 潮间带淹没范围宽, 反之, 潮间带淹没范围小。较高的潮区, 海水淹没时间较短, 必然影响大型底栖动物的摄食和栖息, 特别是夏季和秋季, 太阳高照, 底质表面温度升高, 导致底质表面日温差增大, 不适宜一些耐温范围较窄的大型底栖动物栖息。X断面4个季节的大型底栖动物群落空间分布差异比Y断面小, 是因为X断面是新种植红树林的湿地, 底表凹凸不平, 因此潮汐的影响较小。
| 季节 Season |
时间 Time |
最大潮高/cm Maximum tidal height |
最小潮高/cm Minimum tidal height |
潮差/cm Tidal range |
| 冬季 Winter | 2019年1月7日 | 219 | 32 | 187 |
| 2019年1月22日 | 242 | 6 | 236 | |
| 2020年1月12日 | 231 | 15 | 216 | |
| 2020年1月25日 | 217 | 25 | 192 | |
| 平均 | 227 | 20 | 208 | |
| 春季 Spring | 2019年4月3日 | 165 | 44 | 121 |
| 2019年4月18日 | 187 | 29 | 158 | |
| 2020年4月7日 | 199 | 25 | 174 | |
| 2020年4月26日 | 182 | 34 | 146 | |
| 平均 | 183 | 33 | 150 | |
| 夏季 Summer | 2019年7月4日 | 209 | -7 | 216 |
| 2019年7月18日 | 198 | 12 | 186 | |
| 2020年7月6日 | 209 | -1 | 210 | |
| 2020年7月22日 | 204 | 2 | 202 | |
| 平均 | 205 | 2 | 204 | |
| 秋季 Autumn | 2019年10月3日 | 206 | 52 | 154 |
| 2019年10月18日 | 201 | 57 | 144 | |
| 2020年10月6日 | 184 | 63 | 121 | |
| 2020年10月20日 | 170 | 34 | 136 | |
| 平均 | 190 | 52 | 139 |
冬季和夏季潮差较春季和秋季大, 冬季和夏季大型底栖动物物种数、栖息密度、多样性指数和丰富度指数较春季和秋季的高, 表明潮汐是影响考洲洋红树林湿地大型底栖动物季节分布的重要环境因子。
潮汐影响潮间带大型底栖动物分布在许多文献中有报道。红树林湿地大型底栖动物群落的垂直分带现象主要由潮位线、食物适应性及底质的组成结构等因素所决定[29]。深圳湾潮间带红树林近岸端栖息的大型底栖动物种类主要是无齿相手蟹(Orisarma dehaani)和褶痕相手蟹(Parasesarma plicatum)等, 林带中间部分分布的大型底栖动物主要有拟黑螺(Melanoides sp.)和紫游螺(Neripteron violaceum)等, 而在红树林近海端主要分布的大型底栖动物则是弧边招潮蟹(Tubuca arcuata)、锯脚泥蟹(Ilyoplax dentimerosa)、印尼拟蟹守螺(Pirenella alata)和多毛类的沙蚕等[30—31]。深圳湾潮间带没有红树林光滩腺带刺沙蚕(Neanthes glandicincta)栖息密度高, 但在红树林内很少采集到[32]。泉州湾蟳埔潮间带大型底栖动物功能群物种数和栖息密度明显受潮汐影响。由于沙滩靠近海堤, 潮流经海堤后回流, 导致潮水对沙滩的冲刷作用增强, 一些大型和小型底栖动物容易被冲刷到沉积物表面并被潮水打晕, 因而肉食性物种数和栖息密度在沙滩所占比例均是5种功能群中最高的, 而在互花米草(Spartina alterniflora)生境, 杂食者物种数和碎屑食者栖息密度分别是5种功能群中最高的, 这是因为互花米草的阻碍作用, 减弱了潮水对互花米草生境沉积物的冲刷, 使更多的杂食者和碎屑食者能栖息于互花米草生境[33]。杭州湾南岸低潮带受潮汐扰动影响较大, 杂食者和植食者的种类较少, 故各功能群的多样性指数较低;中潮带营养源及食物较丰富, 功能群类型趋于多样化[34]。潮汐导致潮间带的空间异质性(沉积物粒径的差异), 空间异质性导致泉州湾蟳埔潮间带大型底栖动物功能群组成的差异[33]。在长江口横沙岛、长兴岛潮间带不同潮位间大型底栖动物的总平均栖息密度具有显者差异, 而总平均生物量差异不显著[35]。泉州湾蟳埔潮间带不同潮层除大型底栖动物生物量外, 大型底栖动物物种数、栖息密度、多样性指数、均匀度指数和丰富度指数均有极显著差异, 可见潮汐是影响泉州湾蟳埔潮间带大型底栖动物群落的主要环境因子[36]。
3.2 红树种植年限对考洲洋人工种植红树林湿地大型底栖动物群落的影响考洲洋盐洲大桥附近红树林湿地X断面林内大型底栖动物的总物种数、平均栖息密度和生物量低于Y断面林内, 但X断面林内大型底栖动物的多样性指数、均匀度指数和丰富度指数均低于Y断面。方差分析表明两条断面林内的大型底栖动物栖息密度和丰富度指数有显著空间差异, 但两条断面林内大型底栖动物的物种数、生物量、多样性指数和均匀度指数无显著空间差异。这说明红树植物种植年限对大型底栖动物栖息密度和丰富度指数有显著影响, 而对大型底栖动物的物种数、生物量、多样性指数和均匀度指数无显著影响。这是因为盐洲大桥两侧红树林种植年限相差不到5年, 影响还比较小。如果种植年限长, 则大型底栖动物群落结构将发生明显变化[13]。
红树种植年限与红树的覆盖度有关。随着红树植物的生长, 其高度和树冠的覆盖度也随之增加。植被类型和覆盖度可以通过影响影响营养源的分布和食物结构, 进一步影响底栖动物功能群的结构[37]。红树林内常年凋落的红树枝叶, 在沉积物表面堆积、腐烂, 为一些适合高有机质环境的物种, 如小头虫(Capitella sp.)、可口革囊星虫(Phascolosoma arcuatum)等, 提供了稳定的栖息环境和食物来源, 而光滩由于没有植被的覆盖而大大增加了光滩上大型底栖动物的暴露机率[38]。红树种植2年的湿地大型底栖动物栖息密度比光滩的高[39]。浙江西门岛密秋茄树(Kandelia obovata)生境大型底栖动物的物种数、生物量、栖息密度、均匀度指数、多样性指数、丰度指数和M-AMBI指数平均值均高于疏秋茄树生境[40]。大型底栖动物随着红树种植年限的增加群落结构发生变化[13]。
3.3 红树林湿地大型底栖动物群落的环境响应不同植被之间大型底栖动物群落结构存在差异。广州南沙十四涌潮间带光滩、海桑和茳芏3种生境的大型底栖动物物种数、栖息密度均呈显著差异[41]。新形成的互花米草生境大型底栖动物的物种数和丰富度较高, 但随着时间的推移, 互花米草生境中的大型底栖动物物种数和多样性都会下降[42]。
磷酸盐、无机氮、石油类是影响考洲洋海域环境质量的主要因子, 除磷酸盐存在较普遍超标外, 无机氮、石油类、锌超标因子主要集中在考洲洋与吉隆河交汇处, 工业与城镇用海区, 麒麟河入海口附近陆源入海排污集中的区域[18]。2018年春季和冬季的主要超标因子为DIP和DIN, 夏季的超标因子为DIP, 秋季水质良好[19]。从水质环境看, 考洲洋内部海域均处于富营养化状态, 总体水质状况为良好以下, 潮水的涌入能改善考洲洋的水质状况[18]。受生活污水和工业污水影响, 深圳湾潮间带曾经大量出现小头虫和刚鳃虫(Chaetozone setosa)等耐污大型底栖动物[25], 但2018年和2019年盐洲大桥附近红树林湿地未发现大量的耐污大型底栖动物。
4 结论Y断面冬季的大型底栖动物群落的物种数、栖息密度、生物量、多样性指数(H′)和丰富度指数(d), 以及夏季的栖息密度均随潮区降低而增加;而Y断面冬季的均匀度指数(J)、夏季的H′和J则是随潮区降低而减少。聚类(Cluster)和非度量多维尺度(nMDS)分析表明冬季和夏季X断面和Y断面大型底栖动物群落相似性较低, 而春季和秋季X断面和Y断面大型底栖动物群落相似性较高。冬季和夏季最大潮高、潮差、大型底栖动物物种数、栖息密度、多样性指数和丰富度指数较春季和秋季的高。上述结果可以证实潮汐不仅影响考洲洋红树林湿地大型底栖动物的空间分布, 也影响大型底栖动物的季节分布。红树种植1—2年红树林湿地(X断面)大型底栖动物物种数、H′和J低于种植5—6年的红树林湿地(Y断面), 表明红树种植年限对大型底栖动物群落有影响。
| [1] |
Walters B B, Rönnbäck P, Kovacs J M, Crona B, Hussain S A, Badola R, Primavera J H, Barbier E, Dahdouh-Guebas F. Ethnobiology, socio-economics and management of mangrove forests: a review. Aquatic Botany, 2008, 89(2): 220-236. DOI:10.1016/j.aquabot.2008.02.009 |
| [2] |
Alongi D M. Present state and future of the world's mangrove forests. Environmental Conservation, 2002, 29(3): 331-349. DOI:10.1017/S0376892902000231 |
| [3] |
Leung J Y S, Cheung N K M. Can mangrove plantation enhance the functional diversity of macrobenthic community in polluted mangroves?. Marine Pollution Bulletin, 2017, 116(1/2): 454-461. |
| [4] |
Kon K, Kurokura H, Tongnunui P. Influence of a microhabitat on the structuring of the benthic macrofaunal community in a mangrove forest. Hydrobiologia, 2011, 671(1): 205-216. DOI:10.1007/s10750-011-0718-0 |
| [5] |
Bonaglia S, Brüchert V, Callac N, Vicenzi A, Chi Fru E, Nascimento F J A. Methane fluxes from coastal sediments are enhanced by macrofauna. Scientific Reports, 2017, 7(1): 13145. DOI:10.1038/s41598-017-13263-w |
| [6] |
Kon K, Kurokura H, Tongnunui P. Effects of the physical structure of mangrove vegetation on a benthic faunal community. Journal of Experimental Marine Biology and Ecology, 2010, 383(2): 171-180. DOI:10.1016/j.jembe.2009.11.015 |
| [7] |
Giri C, Ochieng E, Tieszen L L, Zhu Z, Singh A, Loveland T, Masek J, Duke N. Status and distribution of mangrove forests of the world using earth observation satellite data. Global Ecology and Biogeography, 2011, 20(1): 154-159. DOI:10.1111/j.1466-8238.2010.00584.x |
| [8] |
陈光程, 余丹, 叶勇, 陈彬. 红树林植被对大型底栖动物群落的影响. 生态学报, 2013, 33(2): 327-336. |
| [9] |
肖泓, 尹煮石, 张敏, 陈桂珠, 彭逸生. 广东电白人工红树林大型底栖动物群落的功能特征及其影响因子. 生态学杂志, 2020, 39(6): 1813-1822. |
| [10] |
Kristensen E, Bouillon S, Dittmar T, Marchand C. Organic carbon dynamics in mangrove ecosystems: a review. Aquatic Botany, 2008, 89(2): 201-219. DOI:10.1016/j.aquabot.2007.12.005 |
| [11] |
Nagelkerken I, Blaber S J M, Bouillon S, Green P, Haywood M, Kirton L G, Meynecke J O, Pawlik J, Penrose H M, Sasekumar A, Somerfield P J. The habitat function of mangroves for terrestrial and marine fauna: a review. Aquatic Botany, 2008, 89(2): 155-185. DOI:10.1016/j.aquabot.2007.12.007 |
| [12] |
陈国贵, 王文卿, 谷宣, 洪伟, 林宇枫, 王瑁. 漳江口红树植物群落周转对大型底栖动物群落结构的影响. 生态学报, 2021, 41(11): 4310-4317. |
| [13] |
Rao Y Y, Cai L Z, Zhou X P, Fu S J, Peng W Q, Chen X W, Zheng B. Changes of community structure and functional feeding groups of benthic macrofauna after mangrove afforestation in a subtropical intertidal zone, China. Wetlands, 2021, 41(8): 114. DOI:10.1007/s13157-021-01489-4 |
| [14] |
Luo H T, Wang Q, Nie X P, Ren H, Shen Z, Xie X F, Yang Y F. Heavy metal contamination in the cultivated oyster Crassostrea rivularis and associated health risks from a typical mariculture zone in the South China Sea. Bulletin of Environmental Contamination and Toxicology, 2018, 101(1): 33-41. DOI:10.1007/s00128-018-2360-2 |
| [15] |
左娅, 陈向明, 康德礼, 叶长鹏. 基于耗散结构理论的考洲洋生态修复研究. 生态科学, 2015, 34(4): 71-75. |
| [16] |
鄢春梅, 李文凤, 谢绍茂. 美丽海湾建设背景下考洲洋海岸带整治与生态修复实践. 广东园林, 2021, 43(3): 61-65. |
| [17] |
管丽娟, 廖静. 惠州考洲洋: 用红树林捍卫美丽海湾. 海洋与渔业, 2018(4): 41-42. |
| [18] |
马正波, 梁浩亮, 梁婷婷, 范开文, 谢文琦. 考洲洋水环境质量现状及受潮汐影响变化分析. 环境保护科学, 2017, 43(4): 72-78. |
| [19] |
梁婷婷, 王军星, 余清华, 杨小平, 钟锦明. 考洲洋海域水质季节变化及评价方法初步研究. 海洋开发与管理, 2021, 38(7): 96-102. |
| [20] |
Wang X N, Gu Y G, Wang Z H, Ke C L, Mo M S. Biological risk assessment of heavy metals in sediments and health risk assessment in bivalve mollusks from Kaozhouyang Bay, South China. Marine Pollution Bulletin, 2018, 133: 312-319. |
| [21] |
杨小平, 陈冰玲, 谢文琦, 梁婷婷, 肖思巧. 考洲洋入海口附近海域海水重金属含量分析. 中国资源综合利用, 2021, 39(5): 119-124. |
| [22] |
Yu Z L, Lin Q, Gu Y G, Ke C L, Sun R X. Spatial-temporal trend and health implications of polycyclic aromatic hydrocarbons (PAHs) in resident oysters, South China Sea: a case study of Eastern Guangdong coast. Marine Pollution Bulletin, 2016, 110(1): 203-211. |
| [23] |
梁曾飞, 黄哲, 彭泰来, 宿明, 舒服, 张蓓. 惠东县红树林资源现状及保护对策. 浙江林业科技, 2021, 41(1): 108-112. |
| [24] |
应锐, 蒋力, 尹芳敏, 黄建荣, 黎祖福, 王英永, 冯建祥. 基于刺网、地笼的考洲洋游泳动物群落结构和多样性分析. 南方水产科学, 2019, 15(4): 1-10. |
| [25] |
蔡立哲. 深圳湾底栖动物生态学. 厦门: 厦门大学出版社, 2015: 1-303.
|
| [26] |
周红, 张志南. 大型多元统计软件PRIMER的方法原理及其在底栖群落生态学中的应用. 青岛海洋大学学报: 自然科学版, 2003, 33(1): 58-64. |
| [27] |
国家海洋信息中心. 2019潮汐表. 第3册. 台湾海峡至北部湾. 北京: 海洋出版社, 2018: 1-587.
|
| [28] |
国家海洋信息中心. 2020潮汐表. 第3册. 台湾海峡至北部湾. 北京: 海洋出版社, 2019: 1-587.
|
| [29] |
Morton B, Morton J. The Sea Shore Ecology of Hong Kong. Hong Kong, China: Hong Kong University Press, 1983.
|
| [30] |
余日清, 陈桂珠, 章金鸿, 黄玉山. 排放生活污水对红树林底栖动物群落季节变化的影响. 中国环境科学, 1997, 17(6): 18-21. |
| [31] |
余日清, 陈桂珠, 黄玉山, 谭凤仪. 深圳福田红树林区底栖大型动物群落的空间分带及灌污的可能影响. 生态学报, 1996, 16(3): 283-288. |
| [32] |
黄昆, 蔡立哲, 徐华林, 傅素晶, 周细平, 林和山. 深圳湾福田红树林湿地腺带刺沙蚕分布的生态响应. 厦门大学学报: 自然科学版, 2009, 48(5): 756-762. |
| [33] |
李想, 蔡立哲, 卓异, 郭涛, 饶义勇, 颜露露, 傅素晶, 陈昕韡. 泉州湾蟳埔潮间带大型底栖动物功能群研究. 海洋通报, 2014, 33(5): 497-504. |
| [34] |
李欢欢, 鲍毅新, 胡知渊, 葛宝明. 杭州湾南岸大桥建设区域潮间带大型底栖动物功能群及营养等级的季节动态. 动物学报, 2007, 53(6): 1011-1023. |
| [35] |
陶世如, 姜丽芬, 吴纪华, 赵斌, 李博. 长江口横沙岛、长兴岛潮间带大型底栖动物群落特征及其季节变化. 生态学杂志, 2009, 28(7): 1345-1350. |
| [36] |
卓异, 蔡立哲, 郭涛, 傅素晶, 陈昕韡, 吴辰. 泉州湾蟳埔潮间带大型底栖动物群落的时空分布. 生态学报, 2014, 34(5): 1244-1252. |
| [37] |
袁兴中, 陆健健, 刘红. 长江口底栖动物功能群分布格局及其变化. 生态学报, 2002, 22(12): 2054-2062. |
| [38] |
辛俊宏, 任一平, 徐宾铎, 张崇良, 薛莹, 纪毓鹏. 胶州湾西北部潮滩湿地大型底栖动物功能群. 应用生态学报, 2011, 22(7): 1885-1892. |
| [39] |
颜露露. 同安湾下潭尾人工红树林湿地大型底栖动物群落恢复研究[D]. 厦门: 厦门大学, 2016.
|
| [40] |
李文君, 饶义勇, 林怡雯, 吕宝强, 王航俊, 陈昕韡, 蔡立哲. 西门岛潮间带秋茄与互花米草生境秋季大型底栖动物群落比较. 泉州师范学院学报, 2017, 35(2): 1-7. |
| [41] |
黄睿婧, 蔡立哲, 叶洁琼, 周细平, 刘莎, 许鹏, 彭欣, 曹婧. 广州南沙十四涌潮间带三种生境的大型底栖动物群落比较. 生态学杂志, 2010, 29(6): 1187-1192. |
| [42] |
仇乐, 刘金娥, 陈建琴, 王国祥, 常青. 互花米草扩张对江苏海滨湿地大型底栖动物的影响. 海洋科学, 2010, 34(8): 50-55. |