 2023, Vol. 43
2023, Vol. 43文章信息
- 方康, 徐国策, 李鹏, 王斌, 陈新, 马天文, 魏全, 马凌
- FANG Kang, XU Guoce, LI Peng, WANG Bin, CHEN Xin, MA Tianwen, WEI Quan, MA Ling
- 大理河沉积物微生物碳源利用特征及其影响因素
- Characteristics and influencing factors of microbial carbon source utilization in Dali River sediments
- 生态学报. 2023, 43(13): 5571-5580
- Acta Ecologica Sinica. 2023, 43(13): 5571-5580
- http://dx.doi.org/10.5846/stxb202108162257
-
文章历史
- 收稿日期: 2021-08-16
- 网络出版日期: 2023-03-09





2. 中国电建集团西北勘测设计研究院有限公司, 西安 710065
2. PowerChina Northwest Engineering Corporation Limited, Xi′an 710065, China
近年来, 随着人口增长和经济快速发展, 进入河流的污染物不断增加, 由于目前对这些污染物的处理工艺还不够成熟和彻底, 造成了河流水质出现了不同程度的恶化, 部分河流出现了水体富营养化、有机物含量超标、农药残留等严重环境问题, 致使河流原有的环境被破坏, 严重威胁河流水域生态环境安全, 同时也对人类自身的健康生存环境产生不利影响[1—2]。与河流水体相比, 沉积物具有良好的吸附性, 进入河流中的污染物经过长期的累积并不断汇集到河流沉积物环境中[3]。沉积物中的污染物受到外界的干扰后, 通过释放作用, 造成水体发生二次污染, 加剧河流水质恶化, 进而导致河流生态系统失衡[4]。微生物是世界上最多样化和最丰富的生物类群, 具有调节全球生物化学循环的功能, 几乎影响着生态系统内所有的生物[5—7]。由于微生物群落对外界环境变化高度敏感, 常常被作为监测和反映河流生态环境变化的指示指标[8]。同时, 微生物通过参与河流污染物的迁移、自净等过程, 从而影响河流生态环境[9—10]。因此, 分析受污染河流沉积物中微生物群落的功能多样性, 阐明主要污染物对沉积泥沙中微生物代谢反应及功能特性的作用规律, 这对河流生态保护和生态系统质量提升具有重要意义。
目前, 对于河流微生物的研究大多集中于群落组成和多样性上, 而关于微生物群落代谢活性的研究更多是通过基因序列, 对微生物群落的代谢功能进行推测。考虑到微生物群落表现出的功能可塑性和功能冗余现象, 推测代谢可塑性是细菌群落的内在属性, 而功能冗余的表达更依赖于环境因素[10]。Aßhauer等通过宏基因组以及16S rRNA数据预测功能谱表明与鸟枪法测序方法相比, 在对16S rRNA数据的分类学进行分析时, 微生物群落功能的描述不够全面[11]。Stilianos等研究得出环境条件通过塑造代谢生态位, 进而影响海洋微生物群落中功能种群的分布, 而对单个功能群的分类组成只有微弱的影响[12]。大理河是无定河最大的支流, 地处黄河中游地区[13], 在自然地理环境上, 具有其独特的生态学研究意义。由于气候变化和社会经济活动的影响, 大理河出现了水质恶化、水土流失等一系列生态环境问题, 成为了流域可持续发展不可忽视的制约因素。近年来, 大理河的生态环境安全与健康问题日益受到重视, 但目前对大理河流域沉积物中微生物群落功能的研究鲜有报道。因此, 本文采用了Biolog Eco微平板法, 分析了大理河流域沉积物中微生物的碳源利用特征, 讨论了污染对沉积物中微生物群落功能的影响。这对于更好地的了解大理河水生态健康状况, 更有效地保护水生态环境具有非常重要的现实意义。
1 研究区概况大理河是无定河流域最重要的支流之一, 河流全长159.9 km, 流域面积3904.24 km2, 流域范围在109°14′—110°13′ E, 37°30′—37°56′ N。河流自西向东流经陕西省榆林市的靖边县、横山区、子洲县, 在绥德县城附近注入无定河[13]。大理河是沿岸川道地区农业的灌溉水源, 例如大理渠涉及横山、子洲2县5个乡(镇)的34个村民委员会的用水, 其水质状况与周围居民的生产生活息息相关。近年来由于流域内工业的发展和人口数量的增长, 大量的工业废水和生活污水, 排放到大理河中, 致使大理河尤其是中下游河流遭到严重污染。
2 材料与方法 2.1 样品采集2020年10月, 根据大理河自然条件, 共布设了18个采样点(图 1), 其中干流14个、支流(小理河) 4个;每个样点进行3次重复采样, 使用抓斗式泥沙采样器共采集到沉积物样品54个, 采集样品的时候使用GPS进行定位, 准确记录每个采样点的位置。将每个样点采集的样品一分为二, 一份样品200 g装于无菌采样袋中, 及时压住封条;另一份样品500 g装入塑封袋中, 将样品置于低温箱中, 带回实验室用于后续测试分析。

|
| 图 1 研究区位置及采样点分布图 Fig. 1 The location of the study area and the distribution of sampling points |
将采集的沉积物样品带回实验室, 在实验室通风处自然风干, 去除样品中小石子、腐殖质等杂物后研磨过0.15 mm筛。利用全自动间断化学分析仪(Clever Chem2000德国)测定样品中全磷(TP)、全氮(TN)、氨氮(NH4+)和硝氮(NO3-)。使用有机碳分析仪(TOC) multi N/CⓇ 3100(德国耶拿公司)测定沉积物中有机碳(TOC)含量, 用1 mol/L的盐酸溶液浸泡24 h, 去除沉积物中的无机碳, 用TOC分析仪测定沉积物中的有机碳含量, 测定结果如表 1所示。
| 流域 River basin |
pH | 全氮 TN/(g/kg) |
全磷 TP/(g/kg) |
有机碳 TOC/(g/kg) |
硝氮 NO3--N/(mg/kg) |
氨氮 NH4+-N/(mg/kg) |
| 上游Upstream | 8.0a | 0.14a | 0.16a | 15.97a | 1.51a | 4.07a |
| 中游Midstream | 7.9b | 0.28b | 0.29b | 20.17b | 3.41b | 4.66a |
| 下游Downstream | 7.6b | 0.21b | 0.29b | 20.57b | 2.65b | 4.93a |
| 支流Tributary | 8.1a | 0.24ab | 0.21b | 16.43a | 2.44b | 4.61a |
| 同列不同字母表示显著差异(P < 0.05);TN: Total Nitrogen; TP: Total Phosphorus; TOC: Total Organic Carbon | ||||||
使用Biolog Eco微平板法测定微生物群落碳源利用强度, Biolog Eco微平板法是一种基于群落水平的生理特性分析方法, 具有灵敏度高、分辨能力强的优点, 可以最大限度地保留微生物群落原有的代谢特征[14]。吸光度值用SpectraMaxⓇ iD3多功能微孔板读板机测定。
在超净工作台上取事先活化(25 ℃培养24 h)相当于10 g干沉积物的湿样品置入150 mL三角瓶中, 加入90 mL灭菌的生理盐水(0.85% NaCl), 用无菌棉花封口, 充分震荡30 min后, 静置15 min, 用移液枪吸取10 mL上清液, 按逐步稀释法将上清液稀释为10-3 g/mL悬浮液, 向Biolog Eco板的各孔中加入150 μL悬浮液, 盖好盖子置于恒温(25 ℃)培养箱中培养, 每隔24 h用多功能微孔板读板机在590 nm和750 nm波长下测定各孔的吸光度值, 连续测定168 h[14]。
2.3 计算方法 2.3.1 沉积物微生物碳源利用强度微生物碳源利用强度采用平均颜色变化率(Average Well Color Development, AWCD)来描述, AWCD值越高表明微生物丰度越高、碳源利用强度越大[15]。
平均颜色变化率AWCD:
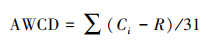
|
(1) |
式中, Ci为每个碳源孔测定的两波段吸光度值的差值;R为对照孔的吸光度值;31为碳源
种类数;Ci-R < 0的孔均在计算中记为零。
2.3.2 沉积物微生物群落功能多样性采用Simpson多样性指数、Shannon-Wiener多样性指数、McIntosh多样指数以及Pielou均匀度指数来表征微生物群落功能多样性[16]。
Shannon-Wiener多指数H:
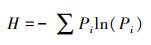
|
(2) |
Simpson指数D:
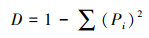
|
(3g11) |
McIntosh指数U:
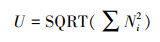
|
(4g11) |
Pielou均匀度指数J:
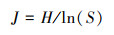
|
(5) |
式中, Pi为第i孔的相对吸光值与平板所有反应孔相对吸光值总和的比率, 即Pi=(Ci-R)/Σ(Ci-R);Ni为第i孔的相对吸光值;S为颜色变化孔的数目, 即Ci-R>0孔的数目。
2.4 碳源种类Biolog Eco板含有31种碳源, 可以分为6大类, 其中氨基酸(Amino acids) 6种、糖类(Carbohydrates)10种、羧酸(Carboxyl acids) 5种、聚合物(Polymers) 4种、胺类(Amines)2种和酚酸类(Phenolic acids)4种[16]。
3 结果与分析 3.1 微生物群落碳源利用强度分析大理河流域上、中、下游及支流沉积物中微生物碳源利用强度(AWCD)的时间变化如图 2所示, 随着培养时间的延长, 碳源利用能力在不断增强, 0—120 h内平均颜色变化率(AWCD)快速增加, 说明悬浊液中的微生物在不断适应Biolog Eco板各孔的环境, 处于快速生长期, Biolog Eco板内底物碳源被大量利用, 此时微生物代谢强度越来越高, 在第120 h后慢慢步入稳定期。流域上、中、下游及支流沉积物中微生物平均颜色变化率(AWCD)总体上呈现, 上游>支流>中游>下游。在进入稳定期后, 沉积物中微生物平均颜色变化率(AWCD)的大小顺序为:上游(0.82)、支流(0.71)、中游(0.57)、下游(0.55), 与上游平均颜色变化率(AWCD)相比, 支流、中游、下游分别降低了13.4%、30.5%、30.7%。

|
| 图 2 微生物群落AWCD值的变化特征 Fig. 2 Variation characteristics of AWCD values of microbial communities AWCE: 平均颜色变化率Average well color development |
对不同培养时间沉积物中微生物对各类碳源的利用进行计算, 结果如图 3所示, 随着培养时间的延长, 微生物对碳源的利用的强度逐渐增大。对比流域上游、中游、下游及支流沉积物中微生物对碳源利用的大小顺序, 沉积物中微生物对不同类型碳源的利用强度明显不同:上游沉积物中微生物对糖类、氨基酸类、酚酸类以及胺类利用强度较高;中游沉积物中微生物对糖类、酚酸类和胺类利用强度较高;下游沉积物中微生物则对糖类、氨基酸类碳源利用强度较高;支流沉积物中微生物对糖类、氨基酸、酚酸类利用强度较高。由此可见, 流域沉积物中微生物主要利用的碳源为糖类和氨基酸类, 其次是酚酸类、胺类, 对羧酸类和聚合物类利用较少。

|
| 图 3 不同培养时间下微生物群落6类碳源利用情况 Fig. 3 The utilization of six types of carbon sources in the microbial community under different incubation times A、B、C、D、E、F代表的是六种碳源, 分别对应的是糖类、氨基酸类、羧酸类、酚酸类、胺类、聚合物类 |
根据微生物群落碳源利用强度(AWCD)值, 选择培养第24 h、120 h、168 h数据, 分别代表微生物培养初期、稳定期、培养结束时进行微生物群落功能多样性分析, 结果如图 4所示。流域上、中、下游以及支流沉积物中微生物群落功能多样性指数呈现出了不同的变化规律。从图 4可知, 随着培养时间延长, 功能多样指数逐渐增大。到达稳定期后, Shannon-Wiener指数(H)、Pielou均匀度(J)指数变化不明显;上游及支流McIntosh指数(U)变化较小, 中游和下游McIntosh指数(U), 呈现下降趋势;而中游、下游及支流Simpson指数(D)变化不显著, 上游Simpson指数(D)呈现上升趋势。

|
| 图 4 不同培养时间微生物群落功能多样性指数变化图 Fig. 4 Changes of microbial community functional diversity index at different culture time |
培养结束时, 流域沉积物中微生物群落的Shannon-Wiener指数(H)从大到小为上游、下游、中游, 与上游相比, 中游、下游及支流分别降低了13.30%、12.45%、3.62%;从流域上游到下游, Pielou均匀度(J)指数逐渐增大, 从上游的0.774增加到0.922, 增幅达19.12%;Simpson指数(D)变化不明显, 中游较上游增加了4.42%、下游较上游增加了5.63%;McIntosh指数(U)变化最为明显, 上游(1.286)到下游(0.985), 降低了23.41%。
3.4 流域沉积物中微生物群落代谢特征分析对不同样点沉积物中微生物培养168 h的AWCD数据, 标准化后进行主成分分析, 从31种因子中提取出两个主成分因子, 以分析沉积物中微生物群落的代谢特征, 结果如图 5所示。沉积物中微生物群落的两个主成分因子解释度分别为60.9% 和21.4%, 累积达到82.3%。沉积物中微生物利用的碳源, 在坐标图上的主要分散在3个区域, 上游群落主要位于第一象限, 中游位于第二象限, 下游和支流位于第三象限。

|
| 图 5 流域沉积物中微生物碳源利用的主成分变量 Fig. 5 Principal component variables of the utilization of microbial carbon sources in sediments of the river basin |
微生物代谢活动相关的碳源种类主成分分析结果见表 2, 根据碳源在前两个主成分上的得分系数, 选择相关性较高的碳源进行归类。与PC 1相关性较高的碳源有25种, 其中糖类7种, 胺类、酚酸类各2种, 聚合物类4种, 氨基酸类、羧酸类各6种;与PC 2相关性较高的碳源有11种, 其中糖类4种, 胺类、聚合物类、羧酸类各1种, 氨基酸类、酚酸类各2种。由此可知, 对沉积物中微生物代谢活动起主要贡献作用的碳源是糖类和氨基酸类, 这与3.2节碳源利用特征的分析结果一致。
| 碳源种类 Types of carbon sources |
碳源名称 Carbon source name |
主成分相关系数 | 碳源种类 Types of carbon sources |
碳源名称 Carbon source name |
主成分相关系数 | |||
| PC1 | PC2 | PC1 | PC2 | |||||
| 糖类Carbohydrates | β-甲基-D-葡萄糖苷 | 0.697 | 0.780 | 氨基酸类 Amino acids |
L-精氨酸 | 0.787 | -0.398 | |
| D-木糖/戊醛糖 | 0.799 | -0.332 | L-天门冬酰胺 | 0.802 | -0.527 | |||
| i-赤藓糖醇 | 0.811 | -0.187 | L-苯丙氨酸 | 0.833 | -0.095 | |||
| D-甘露醇 | 0.750 | -0.496 | L-丝氨酸 | 0.779 | -0.588 | |||
| N-乙酰-D葡萄糖氨 | 0.825 | -0.226 | L-苏氨酸 | 0.828 | -0.076 | |||
| D-纤维二糖 | 0.338 | 2.119 | 甘氨酰-L-谷氨酸 | 0.831 | -0.031 | |||
| α-D-乳糖 | 0.834 | 0.116 | 羧酸类 Carboxyl acids |
γ-羟丁酸 | 0.822 | 0.397 | ||
| D-半乳糖酸γ-内酯 | -0.141 | -0.070 | α-丁酮酸 | 0.762 | 0.150 | |||
| D-L-α-磷酸甘油 | -0.064 | 0.663 | D-葡糖胺酸 | 0.825 | -0.226 | |||
| 1-磷酸葡萄糖 | 0.809 | 0.211 | D-苹果酸 | 0.786 | -0.499 | |||
| 胺类Amines | 苯乙胺 | 0.504 | 1.545 | D-半乳糖醛酸 | -0.141 | -0.070 | ||
| 腐胺 | 0.819 | -0.336 | 丙酮酸甲酯 | 0.834 | -0.031 | |||
| 聚合物类Polymers | 吐温40 | 0.793 | -0.287 | 衣康酸 | 0.834 | -0.031 | ||
| 吐温80 | 0.807 | 0.380 | 酚酸类 Phenolic acids |
2-羟基苯甲酸 | -0.050 | 2.145 | ||
| α-环式糊精 | 0.812 | 0.528 | 4-羟基苯甲酸 | -0.111 | -0.955 | |||
| 肝糖 | 0.832 | 0.032 | ||||||
| 主成分相关系数:Principal component correlation coefficient;PC 1: 主成分1 Principal component 1; PC 2: 主成分2 Principal component 2 | ||||||||
采用冗余分析方法(RDA)对培养168 h的AWCD值以及功能多样性指数与沉积物理化因子间关系进一步分析, 结果如图 6所示。上游RDA结果表明, 氨氮、硝氮、有机碳、全氮、全磷含量、pH是解释上游微生物群落碳源利用及功能多样性差异的主要因子;中游RDA结果表明, 硝氮、全氮、氨氮、全磷、有机碳含量是解释中游微生物群落碳源利用及功能多样性差异的主要因子;下游RDA结果表明, 氨氮、全氮、硝氮、有机碳含量、pH是解释下游微生物群落碳源利用及功能多样性差异的主要因子;支流RDA结果表明, 硝氮、氨氮、全氮、有机碳含量是解释支流微生物群落碳源利用及功能多样性差异的主要因子。综上所述, 影响流域沉积物中微生物群落碳源利用特征及功能多样性差异的因子有所差异, 全氮、氨氮、硝氮、有机碳含量是主要影响因子;全磷含量对上游及中游沉积物中微生物群落碳源利用及功能多样性影响较大, 而pH则是对上游及下游沉积物中微生物群落碳源利用及功能多样性影响较大。

|
| 图 6 微生物群落碳源利用特征和功能多样性指数与环境因子冗余分析 Fig. 6 Redundancy analysis of carbon source utilization characteristics and functional diversity index of microbial communities and environmental factors AWCD: 平均颜色变化率;H: Shannon-Wiener多样性指数;U: McIntosh多样性指数;D: Simpson多样性指数;J: Pielou均匀度指数 |
微生物是生态系统的重要组成部分, 环境因素变化对其影响较为显著, 而这种影响主要是通过群落代谢特征和功能差异来实现[17]。Biolog Eco方法主要根据微生物对碳源利用能力的差异来描述微生物群落功能的动态变化[18]。碳源利用强度(AWCD)值的变化可以反映沉积物中微生物代谢活性的差异, AWCD值增加越快, 表明微生物代谢活性越高[17]。本次研究中, 随着培养时间的延长, 微生物碳源利用能力在不断增强, 0—120 h微生物群落AWCD值变化最为明显, 120 h后AWCD值趋于平缓, 表明微生物群落对碳源利用达到一个较高的水平;受到其栖息生境的影响, 上游和支流沉积物中为微生物碳源利用较高, 表明上游和支流沉积物环境有利于微生物生存, 其环境为微生物聚集和生长繁殖提供了更有利的场所。
不同种类微生物在环境中的占比不同, 构成了其独特的群落结构, 进而会导致其群落碳源利用特征和功能也不尽相同[19]。本次研究中, 沉积物中微生物主要利用的碳源为糖类和氨基酸类, 其次是酚酸类、胺类, 对羧酸类和聚合物类利用较低。有研究表明, 葡萄糖是蔗糖和糖原合成的中间体, 葡萄糖为微生物生命活动提供基本的能源物质[8], 而在本文研究中微生物对单一碳源葡萄糖的利用强度较高, 这可能跟微生物利用糖类化合物, 最终为其提供生命活动所需要的营养物质有关[5]。此外, 微生物对氨基酸类化合物利用程度较高, Du等研究发现氨基酸类化合物富集会提高嗜氨基酸细菌活性, 导致其对氨基酸类化合物的利用程度增强[20]。而酚酸类化合物可以影响微生物的丰度, 添加酚类物质可以促进降解酚酸化合物的微生物的生长, 其代谢产物可以提高其它物种相对丰度[21—22]。中游沉积物中微生物在培养前期对碳源的利用强度较低, 随着酚酸类碳源的利用逐渐增大, 其它碳源的利用强度不断增大, 尤其是对聚合物类碳源的利用强度。由于大理河不同河段受人类活动干扰程度不一, 沉积物环境差异显著, 进而导致微生物在环境因素的选择下, 其群落功能必然存在差异。
流域沉积物中微生物功能多样性指数分析发现, 随着培养时间延长, 微生物功能多样性指数不断增大;到达稳定期后, 上游到下游随着河流沉积物中污染物含量的增加, 沉积物中微生物Shannon-Wiener多样性指数(H)和McIntosh指数(U)逐渐减小, 而Simpson指数(D)和Pielou均匀度指数(J)逐渐增大。通过野外调查以及实验分析发现上游和支流采样点附近农业点源、面源污染较少, 相对中游、下游来说, 上游及支流人口密度较小, 河流受污染程度较低, 更有利于微生物生存。而河流中下游由于长期污染, 沉积物中微生物群落优势菌群得以继续生存, 而其余物种在环境因素的影响下, 群落丰度逐渐降低甚至衰亡, 群落结构变得单一, 导致微生物群落多样性降低。
有研究表明河流微生物群落功能除了受自然过程的影响, 还受污水排放、土地利用、引水灌溉等一些系列人为因素的影响, 而且这些因素主要产生负面影响[23—24]。另外, 造成沉积物中微生物群落动能多样性差异, 可能是由于Biolog Eco板中的底物改变了沉积物中的优势菌群, 促进某些优势菌群生长代谢, 抑制其它丰度较低种群生长代谢, 致使某些功能菌群消失, 从而降低微生物群落碳源利用强度和功能多样性[25]。培养到达稳定期后, 上游Simpson指数(D)继续上升, 由于Biolog Eco板中的底物碳源被大量利用, 导致次优势菌群碳源利用处于劣势, 相对丰度下降, 进而Simpson指数(D)继续增大;而中游及下游McIntosh指数(U)逐渐减小, 由于流域中游及下游污染较为严重, 到达稳定期后, 沉积物中的污染物成为限制微生物群落碳源利用的主要因素。
沉积物是河流生态系统中氮、磷等物质循环的重要场所, 而沉积物中微生物通过同化、异化等代谢过程参与氮污染物降解、转化[26—27]。由于微生物对环境变化非常敏感, 其群落的碳源利特征和功能会随着环境因素的变化而变化[8, 28]。本研究发现, 影响沉积物中微生物群落碳源利用及功能多样性差异的因子有所差异, 全氮、氨氮、硝氮含量是主要影响因子, 其次是全磷含量和pH。汪丽娜的研究结果表明全氮、铵态氮和硝态氮是AOA群落结构变化的关键影响因子, 铵态氮、全氮和pH是AOB群落结构变化的关键影响因子, 影响Anammox群落结构的关键因子是pH、全氮和铵态氮[29]。pH对水体中营养盐的比例分配和存在形态存在一定的影响, 不同微生物适应生长的pH值范围也不同, 从而影响微生物的生长繁殖及群落功能[30]。钟芳[31]等研究得出细菌群落丰度与有机质、全氮、速效钾呈显著的正相关关系。侯晓杰[32]等研究发现微生物对碳源利用受到pH、速效钾显著影响, 有机碳、速效氮含量和土壤碳氮比与微生物群落功能多样性密切相关。此外, 利用Biolog Eco微平板法研究微生物群落功能多样性存在一定缺陷, 不可培养菌群的功能难以发现, 在以后的研究中, 须结合其他群落结构分析方法开展沉积物中微生物功能型菌群研究[33]。随着近些年来的高通量测序技术以及其它分子生物学方法技术改进, 将分子生物学方法和Biolog Eco方法相结合, 有助于更进一步揭示沉积物中微生物碳源利用特征和功能多样性与污染之间的关系。
5 结论(1) 大理河流域沉积物中微生物群落碳源利用特征由高到低为:上游、支流、中游、下游, 与上游相比, 支流、中游、下游分别降低了13.4%、30.5%、30.7%。
(2) 大理河流域沉积物中微生物功能多样性McIntosh指数(U)、Shannon-Wiener指数(H)从上游到下游逐渐减小, Pielou均匀度指数(J)、Simpson指数(D)从上游到下游逐渐增大。
(3) 与微生物代谢活动相关性较高碳源为糖类, 其次是氨基酸类;聚合物类、羧酸类、胺类、酚酸类与微生物代谢活动相关性较低。
(4) 沉积物中全氮、氨氮、硝氮、有机碳含量是影响微生物群落碳源利用及功能多样性差异的主要因子, 全磷含量、pH的影响次之。
| [1] |
Preisner M, Neverova-Dziopak E, Kowalewski Z. Analysis of eutrophication potential of municipal wastewater. Water Science & Technology, 2020, 81(9): 1994-2003. |
| [2] |
Yang P, Xia J, Zhan C S, Mo X G, Chen X J, Hu S, Chen J. Estimation of water consumption for ecosystems based on Vegetation Interfaces Processes Model: A case study of the Aksu River Basin, Northwest China. Science of the Total Environment, 2018, 613-614: 186-195. DOI:10.1016/j.scitotenv.2017.09.045 |
| [3] |
Roberto A A, Van J B, Leff L G. Sediment bacteria in an urban stream: spatiotemporal patterns in community composition. Water Research, 2018, 134: 353-369. DOI:10.1016/j.watres.2018.01.045 |
| [4] |
Gerbersdorf S. U, Hollert H, Brinkmann M, Wieprecht, S, Holger Schüttrumpf H, Manz W. Anthropogenic pollutants affect ecosystem services of freshwater sediments: the need for a "triad plus x" approach. Journal of Soils and Sediments, 2011, 11(6): 1099-1114. DOI:10.1007/s11368-011-0373-0 |
| [5] |
Kuang J L, Huang L A, He Z. L, Chen L. C, Hua ZS, Jia P, Lli S J, Liu J, li T, Zhou JZ, Shu w S. Prelieting taxononic and funcionalstructure of microbial communities in acid mine drainage. The ISME Journal, 2016, 106: 1527-1539. |
| [6] |
Namita P A, Subash A, Liu X B, Shen L, Zheng Q G. Bacterial Diversity in Alpine Lakes: A Review from the Third Pole Region. Journal of Earth Science, 2019, 30(02): 387-396. DOI:10.1007/s12583-018-1206-5 |
| [7] |
Guo X P, Lu D P, Niu Z S, Feng J N, Chen Y R, Tou F Y, Liu M, Yang Yi. Bacterial community structure in response to environmental impacts in the intertidal sediments along the Yangtze Estuary, China. Marine Pollution Bulletin, 2018, 126(07.): 141-149. |
| [8] |
Guo X P, Yang Y, Niu Z S, Lu D P, Zhu C H, Feng J N, Wu J Y, Chen Y R, Tou F Y, Liu M, Hou L J. Characteristics of microbial community indicate anthropogenic impact on the sediments along the Yangtze Estuary and its coastal area, China. The Science of the total environment, 2019, 648: 306-314. DOI:10.1016/j.scitotenv.2018.08.162 |
| [9] |
Ducrotoy J P. The use of biotopes in assessing the environmental quality of tidal estuaries in Europe. Estuarine, Estuarine Coastal & Shelf Science, 2010, 86(3): 317-321. |
| [10] |
吴兰, 程家劲, 贺勇, 陈圆圆, 刘颖, 魏鑫, 刘亚军. 基于Biolog-Eco法对鄱阳湖不同湿地类型下土壤微生物功能多样性. 南昌大学学报(理科版), 2020, 44(6): 585-592. DOI:10.13764/j.cnki.ncdl.2020.06.013 |
| [11] |
Aßhauer K P, Wemheuer B, Daniel R, Meinicke P. Tax4Fun: predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics (Oxford, England), 2015, 31(17): 2882-2884. |
| [12] |
Louca S, Parfrey L W, Doebeli M. Decoupling function and taxonomy in the global ocean microbiome. Science, 2016, 353(6305): 1272-1277. DOI:10.1126/science.aaf4507 |
| [13] |
贾路, 任宗萍, 李占斌, 李鹏, 徐国策, 张铁钢, 杨媛媛. 基于耦合协调度的大理河流域径流和输沙关系分析. 农业工程学报, 2020, 36(11): 86-94, 328. |
| [14] |
Song B, Zeng Z T, Zeng G M, Gong J L, Xiao R, Chen M, Tang X, Ye S J, Shen M C. Effects of hydroxyl, carboxyl, and amino functionalized carbon nanotubes on the functional diversity of microbial community in riverine sediment. Chemosphere, 2021, 262: 128053. DOI:10.1016/j.chemosphere.2020.128053 |
| [15] |
高晓奇, 肖能文, 叶瑶, 付梦娣, 李俊生. 基于Biolog-ECO分析长庆油田土壤微生物群落功能多样性特征. 应用与环境生物学报, 2014, 20(05): 913-918. |
| [16] |
王俊力, 刘福兴, 乔红霞, 付子轼, 孟格蕾, 蔡敏. 滤解带构建对硬质驳岸水体微生物功能多样性的影响. 生态与农村环境学报, 2022, 38(03): 343-349. |
| [17] |
田雅楠, 王红旗. Biolog法在环境微生物功能多样性研究中的应用. 环境科学与技术, 2011, 34(03): 50-57. |
| [18] |
Gryta A, Frąc M, Oszust K. The application of the Biolog EcoPlate approach in ecotoxicological evaluation of dairy sewage sludge. Applied biochemistry and biotechnology, 2014, 174(4): 1434-1443. |
| [19] |
成水平, 王月圆, 吴娟. 人工湿地研究现状与展望. 湖泊科学, 2019, 31(06): 1489-1498. |
| [20] |
Du R, Bai J, Wang S Z, Wu Q H. Response of soil microbial community function to chemical-aided remediation of multi-metal contaminated soils using Jatropha curcas. Huanjing Kexue Xuebao/Acta Scientiae Circumstantiae, 2011, 31(3): 575-582. |
| [21] |
郭莹, 王一明, 巫攀, 彭双. 长期施用粪肥对水稻土中微生物群落功能多样性的影响. 应用与环境生物学报, 2019, 25(03): 593-602. |
| [22] |
李森森, 马大龙, 臧淑英, 王璐璐, 孙弘哲. 不同干扰方式下松江湿地土壤微生物群落结构和功能特征. 生态学报, 2018, 38(22): 7979-7989. |
| [23] |
Saxena G, Marzinelli E M, Naing N N, He Z L, Liang Y T, Tom L, Mitra S, Ping H, Joshi U M, Reuben S, Mynampati K C, Mishra S, Umashankar S, Zhou J Z, Andersen G L, Kijelleberg S, Swarup S. Ecogenomics reveals metals and land-usepressures on microbial communities in the waterways of a megacity. Environmental Science Technology, 2015, 49(3): 1462-1471. |
| [24] |
Tang V T, Rene E R, Fu D F, Singh R P, Behera S K, Pugazhendhi A. Effect of mixed microbial culture addition on enhanced river water quality: Pollutant removal and microbial community characteristics. Environmental Technology & Innovation, 2020, 18: 100707. |
| [25] |
王杰, 李刚, 修伟明, 赵建宁, 王慧, 杨殿林. 贝加尔针茅草原土壤微生物功能多样性对氮素和水分添加的响应. 草业学报, 2014, 23(04): 343-350. |
| [26] |
Wang W J, Yi Y J, Yang Y F, Zhou Y, Zhang S H, Wang X, Yang Z F. Impact of anthropogenic activities on the sediment microbial communities of Baiyangdian shallow lake. International Journal of Sediment Research, 2020, 35(2): 180-192. |
| [27] |
宋吉雪, 明红霞, 石婷婷, 苏洁, 陈泉睿, 金媛, 樊景凤. 辽河口沉积物中参与主要生物地化循环的微生物功能基因分析. 海洋与湖沼, 2021, 52(04): 904-916. |
| [28] |
李轶, 雷梦婷, 杨楠, 张文龙, 牛丽华, 王龙飞. 河流微生物生态学的研究进展. 水资源保护, 2022, 38(01): 190-197. |
| [29] |
汪丽娜. 赣江南昌段底泥脱氮功能微生物群落结构研究[D]. 南昌: 江西农业大学, 2021.
|
| [30] |
汤琼. 筑坝对长江上游微生物群落的影响研究[D]. 重庆: 重庆交通大学, 2021.
|
| [31] |
钟芳, 赵瑾, 孙荣高, 李正平, 王万鹏. 兰州南北两山五类乔灌木林草地土壤养分与土壤微生物空间分布研究. 草业学报, 2010, 19(03): 94-101. |
| [32] |
侯晓杰, 汪景宽, 李世朋. 不同施肥处理与地膜覆盖对土壤微生物群落功能多样性的影响. 生态学报, 2007(02): 655-661. |
| [33] |
闫法军, 田相利, 董双林, 阳钢, 刘瑞娟, 张凯. 刺参池塘底质微生物群落功能多样性的季节变化. 生态学报, 2014, 34(11): 2996-3006. |