 2023, Vol. 43
2023, Vol. 43文章信息
- 刘朝红, 周晓兵, 陆永兴, 张世航, 刘学军, 张元明, 张玲卫
- LIU Chaohong, ZHOU Xiaobing, LU Yongxing, ZHANG Shihang, LIU Xuejun, ZHANG Yuanming, ZHANG Lingwei
- 古尔班通古特沙漠生物土壤结皮对土壤氮库垂直分布特征的影响
- Effects of biological soil crusts on vertical distribution of soil nitrogen pools in the Gurbantunggut Desert
- 生态学报. 2023, 43(12): 5005-5016
- Acta Ecologica Sinica. 2023, 43(12): 5005-5016
- http://dx.doi.org/10.5846/stxb202205121334
-
文章历史
- 收稿日期: 2022-05-12
- 网络出版日期: 2023-02-10






2. 中国科学院新疆生态与地理研究所 荒漠与绿洲生态国家重点实验室, 乌鲁木齐 830011;
3. 中国科学院大学, 北京 100049;
4. 中国农业大学资源与环境学院, 北京 100193
2. State Key Laboratory of Desert and Oasis Ecology, Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Urumqi 830011, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China;
4. College of Resources and Environmental Sciences, China Agricultural University, Beijing 100193, China
生物土壤结皮(Biological soil crusts, BSCs)是由藻类、地衣、苔藓等土壤微生物与表层土壤颗粒胶结而形成的复合体[1], 是全球干旱和半干旱区重要的地表覆盖类型。生物土壤结皮约占全球陆地面积的12%, 其在干旱区具有广阔的分布面积, 覆盖度可达70%以上[2]。生物土壤结皮能够将大气中的氮气固定, 转变为被植物吸收利用的铵态氮。因此, 生物土壤结皮是荒漠等干旱生态系统中氮素的重要来源, 缓解了荒漠生态系统的氮限制。已有研究发现, 生物土壤结皮的固氮量约占全球生物固氮总量的40%[3], 对荒漠生态系统氮循环产生了重要的影响。同时, 生物土壤结皮也能分解和矿化有机化合物, 改善土壤中的养分, 从而影响其他生物的生长, 具有极其重要的生态功能[4—5]。目前, 虽然国内外有关生物土壤结皮固定氮素的研究已有许多报道, 但研究结果多集中于生物土壤结皮层和0—20cm表层土壤不同形态氮素的影响[6—8], 对于生物土壤结皮覆盖下深层土壤的研究鲜见报道。因此, 进一步厘清生物土壤结皮影响下荒漠生态系统中土壤剖面氮库的分布特征, 对深入认识荒漠氮循环具有重要意义。
陆地生态系统中氮库形态较多, 包括铵态氮、硝态氮、微生物生物量氮等各种类型。在荒漠生态系统中, 广泛分布不同类型的生物土壤结皮不同类型生物土壤结皮对土壤各形态氮素含量的影响不同[9]。对土壤氮素的固定和转化速率作用也不同[10]。研究表明, 地衣结皮中各形态氮素含量比藻类结皮中更高[9]。生物土壤结皮对氮素含量影响限定在一定的土壤深度, 例如, 樊瑾等[11]在毛乌素沙地的研究表明生物土壤结皮增加土壤氮素的累积, 对结皮层氮素的提升显著高于下层土壤, 其影响可至其下0—20 cm土层。生物土壤结皮也能够从多个方面改变土壤理化性质。例如降低pH、增加土壤肥力、调节水文等作用, 进而间接影响土壤各形态氮素的分布转化[12]。此外, 同氮循环相关的土壤酶活性(如脲酶、硝酸盐还原酶和亮氨酸氨基肽酶)是表征土壤氮素状况的重要指标, 影响着土壤中氮素的循环。土壤酶活性的高低可以反应土壤微生物数量、有机质含量以及氮素动态。生物土壤结皮的发育改善了土壤微生物群落结构, 进而影响土壤酶活性[13]。为了对氮循环相关指标有一个系统认识, 综合性指标显得尤为重要, 如氮循环多功能特征指数。各氮库与酶活性等可整合形成氮循环多功能特征指数, 它的建立可指示植物群落的生产力[14], 了解荒漠生态系统的氮限制, 进而提高对荒漠生态系统功能的复杂性的认识。因此, 氮循环多功能特征指数可能影响荒漠生态多功能的关键因素之一, 其变化动态如何, 值得关注。
古尔班通古特沙漠作为典型温带荒漠生态系统, 其生物土壤结皮覆盖面积在30%以上, 对沙漠气候调节、土壤氮素固定等方面发挥着重要的生态功能[15]。本研究旨在解析不同类型生物土壤结皮及下层土壤的氮库分布特征及其影响因素, 确定生物土壤结皮影响程度和范围, 为生物土壤结皮多功能性的评估提供数据支持。
1 研究区概况研究区位于古尔班通古特沙漠腹地(87°15′ N, 45°30′ E), 该沙漠是中国最大的固定半固定沙漠, 海拔300至600 m, 面积约在4.88×104 km2。年平均降水量和年平均温度分别为129 mm和8.3℃, 年潜在蒸发量超过2000 mm[16—17]。古尔班通古特沙漠地表植被分属短命植物、1年生长营养期植物、多年生草本、小半灌木、灌木和小半乔木6种生活型。古尔班通古特沙漠地表发育有良好的生物土壤结皮, 是沙漠主要的地表景观之一。生物土壤结皮主要有藻结皮、地衣结皮和藓类结皮等类型及混生型, 其中藻类结皮主要分布在沙垄上部, 地衣结皮主要在沙垄的中下部, 藓类结皮以齿胁赤藓(Syntrichia canivervis)为主, 主要分布在丘间底部[18]。
2 研究方法 2.1 样品采集采样于2021年6月上旬在古尔班通古特沙漠腹地进行。为避免地形对结果的影响, 选取三块地势相对平坦、生物土壤结皮分布均匀的丘间低地设置3个30 m×30 m的大样方(相距1000 m), 相当于3个重复。样方内植被类型、生物土壤结皮盖度、厚度、粗糙度、抗压强度和抗剪强度等信息见表 1。在每个大样方内选择3个1 m×1 m的小样方, 其类型分别为裸沙、藻-地衣混生结皮和藓类结皮。通过裸沙、藻-地衣混生结皮和藓类结皮土壤剖面进行土壤样品采集, 取样层次分为结皮层和结皮层下0—2、2—5、5—10、10—20、20—30、30—50、50—70和70—100 cm 8个层次(共9层), 取样时避开植物和样方边缘。于实验室清除样品中可见植物根系、石子等异物, 每份土壤样品分2份, 一份样品置于4 ℃冰箱, 用于测定硝态氮和铵态氮含量, 一份样品自然风干后用于测定其他土壤理化性质。
| 类型 Type |
盖度 Cover/% |
厚度 Ts/mm |
粗糙度 Rs/% |
抗压强度 Cs/(N/cm2) |
抗剪强度 Ss/(kg/cm2) |
主要植被类型 Main vegetation types |
深度 Depth/cm |
有机碳 TOC/(g/kg) |
全磷 TP/(g/kg) |
酸碱度 pH |
电导率 EC/(μS/cm) |
含水率 SWC/% |
| 裸沙Bare soil | — | — | — | — | — | 以梭梭(Haloxylon ammodendron)为优势种的灌木和尖喙牻牛儿苗(Erodiumoxyrhinchum)为优势种的草本 | 0—2 | 1.00±0.26 | 0.03±0.01 | 7.88±0.10 | 122.67±13.12 | 0.06±0.04 |
| 2—5 | 0.99±0.14 | 0.03±0.00 | 7.90±0.09 | 108.17±10.69 | 0.07±0.02 | |||||||
| 5—10 | 1.55±0.50 | 0.04±0.02 | 8.07±0.05 | 98.17±4.63 | 0.11±0.00 | |||||||
| 10—20 | 0.92±0.04 | 0.03±0.01 | 8.03±0.02 | 93±2.03 | 1.3±1.16 | |||||||
| 20—30 | 1.01±0.27 | 0.02±0.00 | 8.20±0.07 | 84.8±7.78 | 0.05±0.03 | |||||||
| 藓类结皮Moss crust | 23.7±0.13 | 18.08±1.74 | 10.76±2.83 | 22.33±6.04 | 5.22±0.14 | BSC | 3.89±0.92 | 0.05±0.02 | 7.42±.03 | 138.23±46.45 | 0.09±0.03 | |
| 0—2 | 2.01±0.51 | 0.05±0.02 | 7.68±0.09 | 161.03±73.49 | 0.05±0.02 | |||||||
| 2—5 | 1.97±0.50 | 0.05±0.01 | 7.94±0.04 | 158.90±38.61 | 0.04±0.02 | |||||||
| 5—10 | 2.11±0.5 | 0.05±0.02 | 8.00±0.01 | 219.23±114.03 | 0.11±0.03 | |||||||
| 10—20 | 1.99±0.63 | 0.04±0.00 | 8.06±0.04 | 167.07±53.47 | 0.07±0.03 | |||||||
| 20—30 | 1.24±0.28 | 0.04±0.00 | 7.74±0.26 | 153.03±32.68 | 0.11±0.02 | |||||||
| 藻地衣混生结皮 | 25.31±0.07 | 10.74±1.20 | 11.45±1.58 | 8.94±1.89 | 2.36±0.29 | BSC | 11.49±0.27 | 0.07±0.01 | 7.39±0.02 | 389.33±92.02 | 0.04±0.02 | |
| Algal-lichen crust | 0—2 | 3.40±0.27 | 0.06±0.01 | 7.8±0.03 | 160.47±29.29 | 0.02±0.01 | ||||||
| 2—5 | 1.73±0.14 | 0.07±0.01 | 8.06±0.01 | 145.27±21.08 | 0.08±0.04 | |||||||
| 5—10 | 1.53±0.24 | 0.06±0.01 | 8.23±0.07 | 132.17±15.86 | 1.24±1.15 | |||||||
| 10—20 | 1.20±0.40 | 0.05±0.01 | 8.28±0.02 | 14.67±1.24 | 0.11±0.02 | |||||||
| 20—30 | 0.81±0.10 | 0.03±0.00 | 8.13±0.06 | 106.10±7.00 | 0.09±0.02 | |||||||
| Ts: 厚度Thickness;Rs: 粗糙度Roughness;Cs: 抗压强度Compressive strength;Ss: 抗剪强度Shear strength | ||||||||||||
土壤全氮(TN)和总磷(TP)含量用高氯酸-硫酸消化法测定(SEA1 AutoAnalyzer3, 德国);土壤铵态氮(NH4+-N)和硝态氮(NO3--N)用氯化钙(土水比1 ∶ 10)浸提, 通过连续流动分析仪测定含量(SEA1 AutoAnalyzer3, 德国), 两者之和为无机氮(IN);土壤微生物生物量氮(MBN)经氯仿熏蒸, 用硫酸钾(土水比1 ∶ 4)浸提, 碳氮分析仪(Analytik Jena Multi N/C3100, 德国)测定;土壤可溶性有机氮(DON)用过硫酸钾氧化法测定;土壤游离态氨基酸态氮(FAA)用可见分光光度法测定;土壤含水量(SWC)采用烘干法测定;土壤pH(1 ∶ 2.5)和电导率(EC)(1 ∶ 5)用电位法测定;土壤有机碳(TOC)含量采用干烧法测定(Analytik Jena Multi N/C3100, 德国);土壤脲酶(SUE)、硝酸盐还原酶(S-NR)和亮氨酸氨基肽酶(S-LAP)活性采用相关土壤酶试剂盒测定。
2.3 数据分析氮循环多功能性指数(N-Multifunctionality, N-MF)是指通过土壤氮循环反映土壤生态系统的多功能性[19], 即不同形态氮素为土壤提供的功能和服务的能力, 已经是研究氮循环整体功能的重要指标, 在土地利用和生态系统多功能性等方面[20—21]得到应用。本研究中, 用Z得分法标准化样方内每个氮素指标, 即用测定值减去平均值除以标准差[19]具体计算公式为:
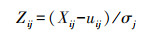
|
式中, Zij为第i类土壤类型第j土层土壤氮素指标的Z得分, Xij为第第i类土壤类型第j土层土壤氮素指标的测定值;ui为第j土层土壤氮素指标的平均值, σj为第j土层土壤氮素指标的标准差。
单个功能指数[21]计算公式为:
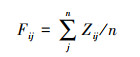
|
式中, Fij为为第i个样方第j土层土壤氮素指标的功能指数; n为该功能包含的生态系统功能指标数量。
对同一类型不同土层以及同一土层不同类型的土壤各氮素形态、土壤酶活性和土壤氮素多功能指数等所有指标采用单因素方差分析和LSD多重比较, 显著性水平设定为α=0.05。用Pearson法对不同形态氮素和土壤酶活性以及土壤理化性质进行相关分析, 土壤理化性质见表 1。采用SPSS 25统计分析软件进行数据处理及统计分析, 采用Origin 2021软件进行作图。
3 结果与分析 3.1 不同类型生物土壤结皮覆盖下土壤氮素和酶活性分布特征 3.1.1 不同类型生物土壤结皮土壤氮素含量及垂直分布特征不同类型生物土壤结皮全氮含量变化趋势明显(图 1), 即从结皮层到0—20 cm土层中全氮含量随土层加深而逐渐降低, 在20—100 cm土层趋于稳定。各土层全氮含量总体上表现为藓类结皮>藻-地衣混生结皮>裸沙, 结皮层全氮含量在132.72—170.62 mg/kg, 显著高于其下层土壤, 但裸沙的全氮含量在2—5 cm土层大于结皮覆盖土壤, 而结皮覆盖土壤在20 cm以下土层基本保持稳定。

|
| 图 1 不同类型生物土壤结皮土壤全氮、无机氮、硝态氮和铵态氮含量垂直分布特征 Fig. 1 Vertical distribution of total, inorganic, nitrate and ammonium nitrogen contents in different types of biological soil crusts S: 裸沙;AL: 藻-地衣混生结皮;M: 藓类结皮; 不同大写字母表不同类型同一土层区间差异显著(P < 0.05), 不通小写字母表示同一类型不同土层区间差异显著(P < 0.05) |
各土层无机氮含量总体上表现为藓类结皮>藻-地衣混生结皮>裸沙(图 1), 结皮层无机氮含量在0.18—12.85 mg/kg之间, 显著高于其下层土壤。裸沙的无机氮含量除2—5 cm土层含量显著高于其余土层, 其余土层差异不显著。藻-地衣混生结皮无机氮含量在结皮层到0—5 cm土层趋于稳定, 在5—10 cm土层下降, 在5—50 cm和50—100 cm土层分别趋于稳定。藓类结皮层无机氮含量显著高于其下层土壤, 0—20 cm土层无机氮含量基本稳定, 在20—30 cm土层上升, 30 cm以下土壤基本趋于稳定。
由图 1可知, 裸沙的硝态氮含量表现为先上升(0—5 cm)后下降(5—10 cm)最后趋于稳定(20 cm以下)。藻-地衣混生结皮硝态氮含量表现先上升(结皮层到0—10 cm)后下降(10—30 cm)最后趋于稳定(30 cm以下)。藓类结皮层硝态氮含量显著高于下层土壤, 在0—20 cm土层基本稳定, 20—30 cm土层硝态氮含量上升, 在50—70 cm土层硝态氮含量最低为3.43 mg/kg。
铵态氮含量变化趋势与无机氮含量变化趋势相似(图 1), 各土层铵态氮含量总体上表现为藓类结皮>藻-地衣混生结皮>裸沙, 结皮层显著高于其下层土壤。裸沙的铵态氮含量表现为先上升(0—5 cm)后下降(5—30 cm)最后趋于稳定(30 cm以下)。藻-地衣混生结皮铵态氮含量表现为先下降(结皮层到0—20 cm)后上升(20—30 cm)最后趋于稳定(30 cm以下)。藓类结皮在0—20 cm土层趋于稳定, 20—30 cm土层上升, 30 cm以下土壤基本趋于稳定。
不同类型生物土壤结皮游离态氨基酸氮含量变化趋势明显(图 2), 即从结皮层到0—20 cm土层游离态氨基酸氮含量随土层加深而逐渐降低, 但在20 cm土层以下逐渐趋于稳定。结皮层游离态氨基酸氮含量5.23—7.02 mg/kg之间, 显著大于其下层土壤;在0—20 cm土层游离态氨基酸氮含量总体上表现为藓类结皮>藻-地衣混生结皮>裸沙, 20 cm土层以下未有显著差异。

|
| 图 2 不同类型生物土壤结皮土壤游离态氨基酸氮、可溶性有机氮和微生物生物量氮含量垂直分布特征 Fig. 2 Vertical distribution characteristics of free amino acid nitrogen, soluble organic nitrogen and microbial nitrogen content in different types of biological soil crusts |
由图 2可知, 不同类型生物土壤结皮可溶性有机氮含量在各土层内总体上表现为藓类结皮和藻-地衣混生结皮>裸沙, 结皮层可溶性有机氮含量为6.01—1.58 mg/kg之间, 显著高于下层土壤。裸沙的可溶性有机氮含量整体表现为先下降(结皮层到0—20 cm)在上升(20—30 cm)最后趋于稳定(30 cm以下), 但2—5 cm土层可溶性有机氮含量最高为2.30 mg/kg;藻-地衣混生结皮和藓类结皮均表现为先下降(结皮层到0—10 cm)在趋于稳定(10 cm以下)。
微生物生物量氮含量在结皮层为35.15—51.52 mg/kg之间, 显著大于下层土壤(图 2)。裸沙的微生物生物量氮含量整体上在0—30 cm土层下降, 30 cm以下趋于稳定, 但2—5 cm土层微生物生物量氮含量最高为2.30 mg/kg;藻-地衣混生结皮和藓类结皮均表现为先下降(结皮层到0—10 cm)在趋于稳定(10 cm以下)。
3.1.2 不同类型生物土壤结皮覆盖下土壤酶活性的分布特征土壤脲酶(SUE)酶活性在0—10 cm土层表现为藓类结皮和藻-地衣混生结皮>裸沙(图 3), 结皮层>下层土壤(P < 0.05)。不同类型生物土壤结皮SUE酶活性变化趋势明显, 在0—10 cm土层中随土层加深而逐渐降低, 而在5 cm以下土层中SUE酶活性基本稳定, 各类型之间SUE酶活性未有显著差异。

|
| 图 3 不同类型生物土壤结皮土壤脲酶垂直分布特征 Fig. 3 Vertical distribution characteristics of urease in different types of biological soil crusts 不同大写字母表不同类型同一土层区间差异显著(P < 0.05);不通小写字母表示同一类型不同土层区间差异显著(P < 0.05) |
土壤亮氨酸氨基肽酶(S-LAP)酶活性(图 4)总体上表现为结皮层大于下层土壤。不同类型生物土壤结皮S-LAP酶活性变化趋势明显, 在0—10 cm土层中随土层加深而逐渐降低, 且裸沙土壤的S-LAP酶活性高于结皮;而在20 cm以下土层中S-LAP酶活性基本稳定, 结皮覆盖土壤S-LAP酶活性高于裸沙。

|
| 图 4 不同类型生物土壤结皮土壤亮氨酸氨基肽酶垂直分布特征 Fig. 4 Vertical distribution characteristics of leucine aminopeptidase in different types of biological soil crusts |
土壤硝酸盐还原酶(S-NR)酶活性在结皮层含量为3.37—5.84 μg d-1 g-1, 显著高于下层土壤(图 5)。不同类型生物土壤结皮S-NR酶活性变化趋势明显, 在0—10 cm土层中随土层加深而逐渐降低, 而在10 cm以下S-NR酶活性基本稳定, 各类型之间S-NR酶活性未有显著差异。

|
| 图 5 不同类型生物土壤结皮土壤硝酸盐还原酶垂直分布特征 Fig. 5 Vertical distribution characteristics of nitrate reductase in different types of biological soil crusts |
图 6表示不同形态氮素与土壤理化性质、土壤酶活性之间的相关性。分析发现, 不同氮素形态之间也存在相关性, 例如全氮与铵态氮、可溶性有机氮、游离态氨基酸氮、微生物生物量氮均呈正相关(P < 0.05)。土壤全氮、铵态氮、可溶性有机氮、游离态氨基酸氮、微生物生物量氮和土壤脲酶(SUE)、亮氨酸氨基肽酶(S-LAP)、硝酸盐还原酶(S-NR)与土壤pH、土壤含水率(SWC)呈负相关(P < 0.05), 与总磷(TP)、电导率(EC)、有机碳(TOC)呈正相关(P < 0.05)。

|
| 图 6 土壤氮素与环境因素的相关性 Fig. 6 Correlation of soil nitrogen with environmental factors |

|
| 图 7 不同类生物土壤结皮土壤氮循环多功能性指数分布特征 Fig. 7 Vertical distribution characteristics of the multifunctionality index of the nitrogen cycle in different types of biological soil crusts |
不同类型生物土壤结皮土壤氮循环多功能性指数(M-MF)变化趋势明显, 整体上表现为在0—100 cm土层中随土层加深先降低(0—20 cm)后趋于稳定(20 cm以下)的趋势, 但仍有个别土层会出现波动变化, 藓类结皮会在20—30 cm土层会有所上升, 裸沙在2—5 cm土层升高。各土层M-MF总体上表现为藓类结皮>藻-地衣混生结皮>裸沙。从土层来看, 结皮层中的M-MF显著大于其下层土壤, 其中以藓类结皮的结皮层M-MF为峰值3.363;其次, 裸沙在2—5 cm土层M-MF显著大于结皮覆盖土壤(P < 0.05)。
4 讨论已有研究表明, 生物土壤结皮可以改善土壤氮素养分状况[1, 22]。在古尔班通古特沙漠, 随着生物土壤结皮的演替, 土壤全氮、速效氮和可溶性氮含量逐渐增加[9]。本研究结果表明, 生物土壤结皮对土壤中氮库的贡献主要集中在结皮层, 而对下层土壤的影响有限, 这与已有的结果具有一致性[23—24]。
已有的古尔班通古特沙漠的研究表明, 生物土壤结皮对氮库的影响最深可达20 cm(研究土层深度为20 cm), 受地形、结皮类型等因素影响[17, 25], 但对20 cm以下的土壤氮库影响还尚不清楚。本试验研究未涉及地形等其他影响, 生物土壤结皮下层土壤氮库的变化可能主要与生物土壤结皮类型相关。本实验证实, 生物土壤结皮的形成增加了结皮层的不同形态氮素(全氮、无机氮、可溶性有机氮、游离态氨基酸态氮)的含量, 即增加了土壤氮库的总量。在0—20 cm土层内, 土壤不同形态氮素随土层加深而逐渐下降, 这与刘鑫等人[23]在古尔班通古特沙漠的研究结果相一致;但是在20—30 cm土层, 藓类结皮下土壤无机氮和铵态氮含量相比10—20 cm土层显著上升, 这可能是由于土壤水分入渗导致的氮素积累有关[26]。从整体上看, 生物土壤结皮中土壤不同形态氮素含量显著高于结皮下层土壤, 这可能是微生物或藻类能够固定氮素, 以及结皮层的形成可吸收土壤中的速效养分造成。例如, 藓类结皮会将土壤速效养分聚集在根系(假根)周围, 供其吸收利用, 导致结皮层的土壤氮库高于下层土壤[27]。但在2—5 cm土层, 裸沙的土壤氮库含量整体上均大于结皮覆盖土壤, 这可能是由于裸沙的表层(0—2 cm)土壤经风蚀, 导致土壤氮素下移以及结皮吸收其下层土壤氮素养分, 亦有可能是结皮死亡后被风沙掩埋, 表现为裸沙地貌, 导致裸沙在2—5 cm土层的氮库相对较高(取样时, 表层有裸沙覆盖, 视为裸沙对照)。在20—100 cm土层, 结皮覆盖下土壤氮库的变化与裸沙没有显著差异, 这说明结皮覆盖显著影响表层土壤(0—20 cm)的氮库, 这与已有研究相符[23—25], 而对于深层土壤氮库的影响较小, 从侧面证明了生物土壤结皮对于土壤氮库的影响在土层深度上有限制。
生物土壤结皮不仅能够改变土壤理化性质, 还能够增加碳、氮、磷等营养物质含量, 为土壤微生物提供稳定的生存环境和养分[28], 影响土壤酶活性。本研究中土壤胞外酶活性(脲酶、亮氨酸氨基肽酶、硝酸盐还原酶)随土壤深度的增加而逐渐下降, 生物土壤结皮覆盖下的土壤比无结皮土壤显著提高了土壤酶的活性, 这与前人的研究结果趋势相基本一致[29], 但土壤酶活性的提升幅度比前人报道效果要更加强烈。例如, 在黄土高原地区, 藓类结皮覆盖下的土壤脲酶比是无结皮土壤的3.5倍[29];在科尔沁沙地, 藓类结皮覆盖下0—20 cm土层的脲酶是无结皮土壤的3.3倍[30]。尽管以上研究所在的研究区域、气候条件、土壤深度、结皮类型等对土壤酶的影响不尽相同, 但这些研究均表明, 生物土壤结皮对土壤酶活性有明显的促进作用, 这也与吴楠和王彦峰等[29—31]的研究的结论类似。土壤酶直接参与土壤不同形态氮素的转化过程, 在一定程度上反应土壤不同形态氮素的转化动态。例如, 宁远英[30]发现土壤酶活性与土壤养分含量呈现正相关关系。土壤酶活性的提高促进土壤养分的积累, 土壤酶活性能反映土壤养分的动态, 可将土壤酶活性作为评价土壤肥力的指标[32—33]。在本研究中, 土壤SUE酶活性、S-LAP含量均与不同氮素形态呈现正相关关系。因此, 生物土壤结皮覆盖增加了土壤酶活性是促进土壤氮库含量增加的主要原因之一。
生物土壤结皮的发育增加了土壤氮库的积累, 但同时也增加了土壤氮库流失的风险。生物土壤结皮的形成改变了土壤表面孔隙状况, 土壤团聚体等特性, 从而影响土壤水分的入渗过程[34—35]。张静等[36]在自然状况下, 通过对有、无生物土壤结皮对凝结水的影响发现, 结皮会吸收大气中的水汽来促进其生长发育, 其具有强大的持水能力。虎瑞[37]在腾格里沙漠对生物土壤结皮氮转化的季节变化发现, 土壤速效氮主要受到土壤水分的影响。在本研究中, 除裸沙的无机氮、铵态氮以及藻-地衣混生结皮的硝态氮外, 土壤在20—30 cm土层的其他速效氮(无机氮、铵态氮和硝态氮)含量均呈现一定的上升, 这可能是由于生物土壤结皮的发育, 对水分的固持减缓了高温对温带沙漠氨化过程的抑制[38], 从而导致速效氮含量的增加。通过对环境因素与土壤不同形态氮素含量的相关性分析发现, 不同形态氮素与脲酶、亮氨酸氨基酶、电导率、总磷和有机碳呈正相关, 这可能是由于土壤碳、磷含量的提高, 增加了土壤微生物的代谢和活动和土壤氮循环过程, 促进了土壤酶活性和氮的固定效率, 进而影响土壤氮库的累积[39]。不同形态氮素与pH、土壤含水率呈负相关, 这可能是由于荒漠生态系统土壤pH普遍偏碱性, 生物土壤结皮的发育降低了土壤pH, 提高了土壤养分的有效性, 缓解荒漠土壤的氮限制[40], 同时结皮的发育增加了土壤水分的固持, 减少了土壤氮库的淋溶[41]。在干旱区生态环境中, 生物土壤结皮改变了土壤理化性质(包括pH等), 这很可能为结皮覆盖下土壤氮库增加提供了有利保障。
生物土壤结皮的发育促进了地下氮循环多功能指数增加, 有助于荒漠生态系统功能稳定。除水分外, 土壤氮库是影响荒漠生态系统的最重要因素。生物土壤结皮通过固定大气中的氮气, 形成植物和微生物可利用的氮素形态, 影响地下氮循环多功能。Elbert[1]在全球尺度上发现, 生物土壤结皮每年的氮吸收约为49 Tg, 占陆地生物固氮的近一半, 证实了生物土壤结皮的覆盖改变了土壤养分, 影响土壤氮库, 促进地下氮循环多功能的过程变化。本研究中, 土壤氮循环多功能指数随土层加深而逐渐降低, 各土层总体上表现为藓类结皮>藻-地衣混生结皮>裸沙, 其结果与土壤氮库分布特征相保持一致。研究结果进一步验证了生物土壤结皮的发育促进了土壤氮库的积累, 增加了地下氮循环过程强度, 进而促进了荒漠系统生态功能的发挥。
因此, 生物土壤结皮可显著提土壤表层(0—20 cm)氮库含量, 对土壤的氮库累积有着重要作用, 提升土壤肥力, 对荒漠植被维持和恢复有着重要作用。在本研究结果中, 生物土壤结皮对土壤深层(30—100 cm)氮库含量的影响并不显著, 土壤各氮素形态含量随各层的变化趋势有差异, 验证了生物土壤结皮对土壤氮库积累具有土层依赖性。
5 结论生物土壤结皮增加了土壤表层的氮库含量, 结皮层和0—100 cm土层的土壤氮库含量和土壤酶活性随土层加深而表现出在0—20 cm土层逐渐降低。除裸沙的无机氮、铵态氮以及藻-地衣混生结皮的硝态氮外, 土壤在20—30 cm土层的其他速效氮(无机氮、铵态氮和硝态氮)含量有所增加, 在30—100 cm土层趋于稳定, 其他各形态氮素含量和土壤酶活性在20—100 cm土层趋于稳定。结皮层和结皮下各层土壤氮库(除硝态氮以外)和土壤酶活性在整体上表现为藓类结皮>藻-地衣混生结皮>裸沙, 而硝态氮含量随土层加深呈现先升高后降低然后趋于稳定的趋势。结皮层土壤氮库含量和土壤酶活性显著高于其下层土壤, 但裸沙在2—5 cm土层土壤氮库含量大于藓类结皮和藻-地衣混生结皮相应土层。土壤氮库与脲酶、亮氨酸氨基酶、电导率、总磷和有机碳呈正相关, 与pH、土壤含水率呈负相关, 同时不同氮素形态之间也存在显著相关性。氮循环多功能随土层加深呈先降低后趋于稳定的趋势, 同样表现为藓类结皮>藻-地衣混生结皮>裸沙, 进一步验证了生物土壤结皮促进土壤氮库的累积, 但这种累积作用主要集中在表层土壤。
| [1] |
Elbert W, Weber B, Burrows S, Steinkamp J, Büdel B, Andreae M O, Pöschl U. Contribution of cryptogamic covers to the global cycles of carbon and nitrogen. Nature Geoscience, 2012, 5(7): 459-462. DOI:10.1038/ngeo1486 |
| [2] |
Rodriguez-Caballero E, Belnap J, Büdel B, Crutzen P J, Andreae M O, Pöschl U, Weber B. Dryland photoautotrophic soil surface communities endangered by global change. Nature Geoscience, 2018, 11(3): 185-189. DOI:10.1038/s41561-018-0072-1 |
| [3] |
Weber B, Wu D M, Tamm A, Ruckteschler N, Rodriguez-Caballero E, Steinkamp J, Meusel H, Elbert W, Behrendt T, Sörgel M, Cheng Y F, Crutzen P J, Su H, Pöschl U. Biological soil crusts accelerate the nitrogen cycle through large NO and HONO emissions in drylands. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(50): 15384-15389. DOI:10.1073/pnas.1515818112 |
| [4] |
姚小萌, 肖波, 王国鹏, 李胜龙, 田乐乐, 孙福海. 外源输入氮素在藓类生物土壤结皮中各氮组分的分配特征与归趋途径. 应用生态学报, 2020, 31(8): 2653-2662. |
| [5] |
Baumann K, Eckhardt K U, Acksel A, Gros P, Glaser K, Gillespie A W, Karsten U, Leinweber P. Contribution of biological soil crusts to soil organic matter composition and stability in temperate forests. Soil Biology and Biochemistry, 2021, 160: 108315. DOI:10.1016/j.soilbio.2021.108315 |
| [6] |
明姣, 赵允格, 许明祥, 杨丽娜, 王爱国, 姚春竹. 黄土丘陵区不同降水量带生物结皮对土壤氮素的影响. 植物营养与肥料学报, 2013, 19(1): 102-110. |
| [7] |
Chamizo S, Cantón Y, Miralles I, Domingo F. Biological soil crust development affects physicochemical characteristics of soil surface in semiarid ecosystems. Soil Biology and Biochemistry, 2012, 49: 96-105. DOI:10.1016/j.soilbio.2012.02.017 |
| [8] |
Delgado-Baquerizo M, Covelo F, Maestre F T, Gallardo A. Biological soil crusts affect small-scale spatial patterns of inorganic N in a semiarid Mediterranean grassland. Journal of Arid Environments, 2013, 91: 147-150. DOI:10.1016/j.jaridenv.2013.01.005 |
| [9] |
Zhou X B, Tao Y, Yin B F, Tucker C, Zhang Y M. Nitrogen pools in soil covered by biological soil crusts of different successional stages in a temperate desert in Central Asia. Geoderma, 2020, 366: 114166. DOI:10.1016/j.geoderma.2019.114166 |
| [10] |
Zhou X B, Smith H, Silva A G, Belnap J, Garcia-Pichel F. Differential responses of dinitrogen fixation, diazotrophic cyanobacteria and ammonia oxidation reveal a potential warming-induced imbalance of the N-cycle in biological soil crusts. PLoS One, 2016, 11(10): e0164932. DOI:10.1371/journal.pone.0164932 |
| [11] |
樊瑾, 李诗瑶, 余海龙, 黄菊莹. 毛乌素沙地不同类型生物结皮与下层土壤酶活性及土壤碳氮磷化学计量特征. 中国沙漠, 2021, 41(4): 109-120. |
| [12] |
周晓兵, 张丙昌, 张元明. 生物土壤结皮固沙理论与实践. 中国沙漠, 2021, 41(1): 164-173. |
| [13] |
周虹, 吴波, 高莹, 成龙, 贾晓红, 庞营军, 赵河聚. 毛乌素沙地臭柏(Sabina vulgaris)群落生物土壤结皮细菌群落组成及其影响因素. 中国沙漠, 2020, 40(5): 130-141. |
| [14] |
王毅, 孙建, 叶冲冲, 曾涛. 气候因子通过土壤微生物生物量氮促进青藏高原高寒草地地上生态系统功能. 植物生态学报, 2021, 45(5): 434-443. |
| [15] |
周晓兵, 尹本丰, 张元明. 模拟氮沉降对不同类型生物土壤结皮生长和光合生理的影响. 生态学报, 2016, 36(11): 3197-3205. |
| [16] |
李新荣, 贾玉奎, 龙利群, 王新平, 张景光. 干旱半干旱地区土壤微生物结皮的生态学意义及若干研究进展. 中国沙漠, 2001, 21(1): 4-11. |
| [17] |
吴文超, 岳平, 崔晓庆, 李凯辉, 刘学军. 古尔班通古特沙漠土壤微生物碳氮对环境因子的响应. 干旱区研究, 2018, 35(3): 515-523. |
| [18] |
张元明, 潘惠霞, 潘伯荣. 古尔班通古特沙漠不同地貌部位生物结皮的选择性分布. 水土保持学报, 2004, 18(4): 61-64. |
| [19] |
Wang H F, Lv G H, Cai Y, Zhang X N, Jiang L M, Yang X D. Determining the effects of biotic and abiotic factors on the ecosystem multifunctionality in a desert-oasis ecotone. Ecological Indicators, 2021, 128: 107830. DOI:10.1016/j.ecolind.2021.107830 |
| [20] |
张君红, 王健宇, 孟泽昕, 何佳, 董政宏, 刘凯茜, 陈文青. 土壤微生物多样性通过共现网络复杂性表征高寒草甸生态系统多功能性. 生态学报, 2022, 42(7): 2542-2558. |
| [21] |
郑佳华, 赵萌莉, 王琪, 张峰, 张彬, 张军. 不同利用方式对大针茅草原生态系统多功能性的影响. 草地学报, 2022, 30(5): 1054-1062. |
| [22] |
Barger N N, Weber B, Garcia-Pichel F, Zaady E, Belnap J. Patterns and controls on nitrogen cycling of biological soil crusts//Weber B, Büdel B, Belnap J, eds. Biological Soil Crusts: An Organizing Principle in Drylands. Cham: Springer, 2016: 257-285.
|
| [23] |
刘鑫, 荣晓莹, 张元明. 古尔班通古特沙漠生物土壤结皮对氨氧化微生物生态位的影响. 生物多样性, 2021, 29(1): 43-52. |
| [24] |
王闪闪, 赵允格, 石亚芳, 高丽倩, 杨巧云. 短期放牧干扰对黄土丘陵区生物结皮土壤氮素累积的影响. 应用生态学报, 2017, 28(12): 3848-3854. |
| [25] |
周晓兵, 张元明, 陶冶, 张丙昌. 古尔班通古特沙漠土壤酶活性和微生物量氮对模拟氮沉降的响应. 生态学报, 2011, 31(12): 3340-3349. |
| [26] |
肖波, 赵允格, 许明祥, 邵明安. 陕北黄土区生物结皮条件下土壤养分的积累及流失风险. 应用生态学报, 2008, 19(5): 1019-1026. |
| [27] |
姚春竹, 赵允格, 王媛, 张培培. 黄土丘陵区生物结皮条件下土壤有机质及氮素的累积特征. 西北农林科技大学学报: 自然科学版, 2014, 42(6): 197-204, 212-212. |
| [28] |
张国秀, 赵允格, 许明祥, 高丽倩. 黄土丘陵区生物结皮对土壤磷素有效性及碱性磷酸酶活性的影响. 植物营养与肥料学报, 2012, 18(3): 621-628. |
| [29] |
王彦峰, 王兵, 肖波, 温奋翔. 陕北黄土高原藓结皮的7种土壤酶活性及其剖面分布特征. 西北农林科技大学学报: 自然科学版, 2017, 45(3): 161-169. |
| [30] |
徐杰, 宁远英. 科尔沁沙地放牧恢复过程中生物结皮对土壤酶活性的影响. 内蒙古师范大学学报: 自然科学汉文版, 2010, 39(3): 302-307. |
| [31] |
吴楠, 张元明. 古尔班通古特沙漠生物土壤结皮影响下的土壤酶分布特征. 中国沙漠, 2010, 30(5): 1128-1136. |
| [32] |
Liu Y M, Yang H Y, Li X R, Xing Z S. Effects of biological soil crusts on soil enzyme activities in revegetated areas of the Tengger Desert, China. Applied Soil Ecology, 2014, 80: 6-14. |
| [33] |
Lebrun J D, Trinsoutrot-Gattin I, Vinceslas-Akpa M, Bailleul C, Brault A, Mougin C, Laval K. Assessing impacts of copper on soil enzyme activities in regard to their natural spatiotemporal variation under long-term different land uses. Soil Biology and Biochemistry, 2012, 49: 150-156. |
| [34] |
刘旦旦, 王健, 尹武君. 天然降雨对黄土坡地土壤侵蚀和养分流失的影响. 节水灌溉, 2011(8): 17-20, 24-24. |
| [35] |
陈磊, 李占斌, 李鹏, 穆军, 贾莲莲. 黄土高原人为加速侵蚀下水土与养分流失耦合研究. 水土保持学报, 2011, 25(3): 7-11. |
| [36] |
张静, 张元明, 周晓兵, 张丙昌, 魏美丽. 生物结皮影响下沙漠土壤表面凝结水的形成与变化特征. 生态学报, 2009, 29(12): 6600-6608. |
| [37] |
Hu R, Wang X P, Pan Y X, Zhang Y F, Zhang H, Chen N. Seasonal variation of net N mineralization under different biological soil crusts in Tengger Desert, North China. CATENA, 2015, 127: 9-16. |
| [38] |
Hu R, Wang X P, Xu J S, Zhang Y F, Pan Y X, Su X. The mechanism of soil nitrogen transformation under different biocrusts to warming and reduced precipitation: from microbial functional genes to enzyme activity. Science of the Total Environment, 2020, 722: 137849. |
| [39] |
周晓兵, 张元明. 干旱半干旱区氮沉降生态效应研究进展. 生态学报, 2009, 29(7): 3835-3845. |
| [40] |
肖波, 赵允格, 邵明安. 陕北水蚀风蚀交错区两种生物结皮对土壤理化性质的影响. 生态学报, 2007, 27(11): 4662-4670. |
| [41] |
王闪闪, 赵允格, 明姣, 张子辉, 郭雅丽. 黄土丘陵区生物结皮群落组成与水分入渗的关系. 应用生态学报, 2022, 33(7): 1819-1826. |