 2023, Vol. 43
2023, Vol. 43文章信息
- 徐铭泽, 刘鹏, 田赟, 赵洪贤, 靳川, 李满乐, 毛军, 魏晓帅, 贾昕, 查天山
- XU Mingze, LIU Peng, TIAN Yun, ZHAO Hongxian, JIN Chuan, LI Manle, MAO Jun, WEI Xiaoshuai, JIA Xin, ZHA Tianshan
- 毛乌素沙地油蒿叶性状对光能利用效率动态的影响
- Seasonal response of light use efficiency of Artemisia ordosica to leaf traits in Mu Us sandy land
- 生态学报. 2023, 43(12): 5122-5136
- Acta Ecologica Sinica. 2023, 43(12): 5122-5136
- http://dx.doi.org/10.5846/stxb202203240732
-
文章历史
- 收稿日期: 2022-03-24
- 网络出版日期: 2023-02-10






2. 宁夏盐池毛乌素沙地生态系统国家定位观测研究站, 北京 100089;
3. 北京林业大学水土保持国家林业局重点实验室, 北京 100089
2. Yanchi Ecology Research Station of Mu Us Desert, Beijing 100089, China;
3. Key Laboratory of State Forestry Administration on Soil and Water Conservation, Beijing Forestry University, Beijing 100089, China
光能利用效率(light use efficiency, LUE)是表征植物将光能吸收并转化为干物质的生理指标, 该指标从生态经济学角度反映了植物碳增益与光能吸收量之间的相互关系。在光能利用效率模型(如CASA模型和GLO-PEM模型等)中, LUE作为估算生态系统总初级生产力(GPP)或净初级生产力(NPP)的核心参数[1], 广泛运用于各类陆地生态系统的碳循环研究中[2—4]。
近年来LUE在时间尺度所表现出的变异性逐渐成为一个热点问题。在早期相关研究, LUE通常被设定为常数, 或根据植物功能群类型和生态系统类型将LUE划分为不同的常数区间进行计算[5—6]。相关研究表明LUE会随着日、月、季节和年际发生变化。如日尺度上呼伦贝尔贝加尔针茅草原LUE呈先降低后增加的单峰趋势[7];季节尺度杨树人工林LUE在生长季旺季达到峰值后迅速降低[8]。当前主流观点认为光能利用效率模型估算生态系统生产力的误差很大程度上来自于LUE取值的不确定性[9—12]。可见, 理解LUE的时间动态及其变异机制对于提高生产力模型的预测性具有重要意义。
目前针对LUE时间动态的研究主要集中于冠层或生态系统尺度, 发现环境因子对LUE具有明显的调节作用[13—15]。而叶尺度作为解释植物光合生理机理性变化和实现参数尺度扩展的基础单元, 当下关于LUE生物调节途径的认识仍不清晰[9]。一般认为, 植物叶片构型和性状配置模式反映了资源利用效率的最大潜力[16]。如形态结构类性状(比叶面积、叶片相对含水量等)反映了叶片对自然资源的捕获力和抵抗性, 叶氮和光合色素(叶绿素、类胡萝卜素)等生物化学性状则反映了叶片内部对资源的同化、运输效率。叶片的构建成本、功能属性与实际的投资收益比联系密切[17]。
近期研究表明, 叶性状在叶片发育完全后并非一直保持机械稳定性[18—19], 环境的季节波动所导致的性状塑性变化, 被认为是植物适应外部压力变化的重要方面[20]。如植物在适应季节性干旱时提高非光合系统的氮分配比例, 强化叶片的抗逆性[21—23]。旱生植物在应对严重水分亏缺时降低了整体的碳同化速率, 这与叶片的形态结构的调整有关[19]。这些研究共同强调, 生长季中, 植物不断调整其性状组合以高度适应环境, 叶片性状的高可塑性变化直接或间接地影响了植物对资源的整合。另外, 在胁迫环境中, 叶级资源利用效率的下调并不单单是对环境承压的应激反应, 还与叶片本身资源捕捉和同化能力的降低有关[24]。光能资源与水肥等限制性资源相比, 具有瞬时性、不可存储性等特点[12]。基于叶经济谱理论所指出的, 空间上叶片特征与资源获取策略间高度耦合[17, 25], 那么叶性状—资源利用效率间的静态关系可以延伸至时间尺度吗?植物LUE的季节动态能被关键叶性状所捕捉吗?这一科学问题需要得到大量田间观测数据的进一步验证。
荒漠生态系统占陆地面积的20%—30%[26], 是对气候变化最为敏感的陆地生态系统类型之一。由于全球气候变暖进程加快(极端降水, 持续高温、干旱等), 荒漠植物的持续生存状况面临着巨大挑战。油蒿(Artemisia ordosica), 菊科蒿属, 具有极强的环境适应能力, 是半干旱区具有代表性的沙生植物, 广泛分布于中国北部及西北部。目前学术界围绕油蒿的生长动态、蒸腾耗水及光合特性等方面展开了大量研究[27—30], 然而有关油蒿LUE季节动态的研究还鲜有报道。本文假设, 长期干旱环境下, 典型荒漠灌木油蒿光能利用效率日均值(daily mean light use efficiency, LUEday)的动态变化可能跟叶片结构或化学性状的季节调整有关。为验证该假设, 通过对油蒿叶片气体交换特征和8个叶性状指标进行生长季定期测定, 从叶性状角度, 探究影响油蒿LUEday季节变异的生物因素。
1 材料与方法 1.1 研究区概况研究区位于宁夏盐池毛乌素沙地生态系统国家定位观测研究站(37.07°—38.17° N, 106.5°—107.78° E, 海拔1550 m)。该站点位于毛乌素沙地南缘, 气候类型为典型的中温带大陆性季风气候。年均温8.1℃, 年均潜在蒸发量2024 mm[12]。年均降水量(precipitation, PPT)约287 mm, 主要集中于生长季, 其中7—9月约占全年降水的60%以上[15]。研究区内以旱生植物为主, 灌木优势物种为油蒿(Artemisia ordosica)、杨柴(Hedysarum mongolicum)和北沙柳(Salix psammophila)等, 草本优势种为赖草(Leymus secalinus)和针茅(Stipa glareosa)等。
1.2 研究方法研究于2021年5—9月在宁夏盐池荒漠生态系统定位研究站内开展。在10 m × 10 m的油蒿固定样地内, 以每10 d为1个测量周期, 对油蒿叶片生长季的气体交换、功能性状进行同步测定, 通过样地附近的通量塔和土壤水分传感器对空气温度(air temperature, Ta)、相对湿度(relative humidity, RH)、光合有效辐射(photosynthetically active radiation, PAR)、土壤体积含水量(Soil volume water content, VWC)进行连续监测。根据以往研究[31—32], 当地油蒿叶片萌芽期大致在3月下旬, 展叶期在4—5月, 落叶期在9—10月, 每年的物候期稍有变动。本文对于监测样地的油蒿样株进行了两天一次的物候定位观测, 结果表明, 油蒿展叶期为4月9日至5月30日(第100—150天), 完全展叶期为5月31日—8月31日(第151—243天), 落叶期为9月1日至10月7日(第244—280天)。
1.2.1 叶片气体交换参数的测定生长季期间, 在上述样地选取3株标准株(长势一致, 株高、冠幅接近)作为重复, 采用配有2 cm × 3 cm透明叶室的LI-6400XT便捷式光合仪(LI-COR, Lincoln, NE, USA)对油蒿叶片全天的实时光合进行测量。测量日设置为每月的上、中、下旬, 选择晴朗无风天气的日子进行测量。测量时段为当天的8:00—18:00, 间隔为每2 h, 取全天6次时段的平均值作为测量日均值。选择冠层顶部当年生小枝末端的一簇向阳叶片(约5—10片)进行测定, 当测量数值小数点后一位稳定时, 每个叶片连续记录多次读数。主要测量指标含净光合速率(net photosynthetic rate, Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间二氧化碳浓度(Ci)、叶表面温度(Tl)及叶面光合有效辐射(photosynthetically active radiation on leaf surface, PARi)等。后续将测量叶片带回室内进行叶面积拍照, 利用Image J软件计算的实际叶面积对气体交换数据进行校准。光能利用效率日均值计算公式如下:
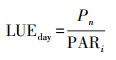
|
由于油蒿叶片较小, 测量气体交换的叶片无法满足后续叶性状测量样本需求, 为最大化减少周期性采样对监测标准株生长的干扰, 本文参考王子奇等[32]和Jiang等[33]对于油蒿叶片的采样方法, 选择在标准株周围选择三个相似株, 采取与气体交换测量样品接近的叶片(叶位、叶片大小较一致)作为油蒿叶性状的测试叶片, 带回实验室进行测定。每个周期的单个测量指标(除叶厚度以外)的重复数设置为3个, 总计15个测量周期。
(1) 叶厚度(leaf thickness, Lt)测定:用精度为0.02 mm的电子游标卡尺测量叶片主脉两侧的厚度, 重复测量10个叶片, 用其平均值代表叶片厚度。
(2) 叶片相对含水量(leaf relative water content, LRWC)测定:取部分叶片称取鲜重, 浸入装有水的烧杯中避光静置5 h使其充分吸水。完成后用吸水纸吸取叶片多余水分, 用万分之一电子天平上称重记录叶片饱和鲜重。称重后将其置于80℃的烘箱烘干48 h, 称取叶片干质量。叶片相对含水量为(鲜重-干重)/(饱和鲜重-干重)。
(3) 比叶面积(specific leaf area, SLA)测定:将叶片平铺在A4纸上进行拍照, 利用Image J软件计算其实际叶面积。将拍过照的叶片装入信封, 置于80℃的烘箱中烘干48 h, 用万分之一电子天平称取其叶片干质量。比叶面积为叶面积/叶干质量。
1.2.3 叶片化学性状的测定(1) 叶片光合色素含量测定:称取2.5 g鲜重叶片, 充分研磨后加入95%酒精, 待其溶于酒精后使用紫外分光光度计(UV-2100, UNIC, USA)分别测量其在665 mm、649 mm和470 mm下的吸光度, 按以下公式计算叶绿素a含量(chlorophyll a, Chl a)、叶绿素b含量(chlorophyll b, Chl b)和类胡罗卜素含量(carotenoid content, Car)的色素浓度。
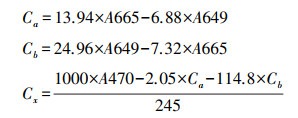
|
式中:Ca、Cb、Cx分别为叶绿素a、b和类胡萝卜素的浓度(mg/L)。A665、A649、A470分别为叶绿体溶液在波长665 nm、649 nm和470 nm下的吸光度。
求得色素浓度后, 计算组织中各色素的含量, 通过比叶面积和叶片干/鲜重比将基于质量的光合色素指标转化为基于面积的Chl a、Chl b和Car, 具体公式如下所示:

|
式中:Chl a、Chl b、Car分别为叶绿素a、b和类胡萝卜素含量(μg/cm2);V为提取液体积(mL);N为稀释倍数;m为样品鲜重(g);DW为干鲜重比(g/g);SLA为比叶面积(cm2/g);总叶绿素含量(total chlorophyll content, Chl)为Chl a与Chl b之和。
(2) 叶片碳、氮含量测定:将采集的植物叶片分别装入信封置于烘箱中, 在105℃下杀青30 min, 70℃下烘干48 h至恒质量, 烘干后的样叶经过粉碎、研磨制成粉样, 利用凯氏定氮法测定叶氮含量(leaf nitrogen content, LNC)。采用元素分析仪(Vario Max CN Element analyzer, Elementar, Germany)测定叶碳含量(leaf carbon content, LCC)。
1.3 数据处理与分析使用变异系数CV(标准偏差/平均值)×100%, 表征季节尺度油蒿LUEday和8个叶片功能性状的变异程度。参照Burnett等人的研究[20], 采用相对偏差(%)描述LUEday和叶性状在不同物候期的变异幅度(相对整个生长季平均水平), 相对偏差计算公式为:
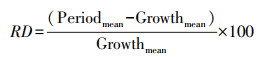
|
式中:RD为具体测量指标的相对偏差, Periodmean为展叶期、完全展叶期和落叶期中任一物候期间测量指标的平均值, Growthmean为测量指标在生长季中的平均值。
采用单因素方差分析(one-way ANOVA)检验油蒿LUEday和8个叶性状在不同物候期间是否存在显著差异, 并采用LSD法进行多重比较。采用Pearson相关分析检验8个叶性状间的相关性。通过线性回归分析检验季节尺度, LUEday和叶性状(Lt、SLA、LRWC、Chl、Chl a/b、Car、LNC和LCC)间的线性关系。在明确影响LUEday季节动态的关键叶性状后, 进一步采用线性回归分析探究了叶性状与环境因子间的关系。本文的数据统计分析均在R 3.6.1中完成, 所有统计分析的显著水平均设置为P=0.05, 绘图则使用Origin 8.2软件完成。
2 结果与分析 2.1 环境因子的季节动态如图 1所示, 生长季PAR范围58.68—610.45 μmol m-2 s-1, 平均值419.10 μmol m-2 s-1, 最大值和最小值分别出现在第160天和第258天;Ta最大值27.22℃(第195天), 最小值7.64℃(第135天), 生长季平均温度18.76℃。7月是生长季中最热的月份(平均Ta 23.41℃), 5月和9月月均温度均低于17℃;饱和水汽压差(vapor pressure deficit, VPD)的季节趋势与RH相反, 观测期内VPD平均值1.34 kPa, 最大值2.88 kPa(第212天), 最小值0.14 kPa(第258天);5—9月累积PPT为135.47 mm, 最大降水月份为9月(54.73 mm), 占生长季PPT的40.40%。最大单次降雨事件发生在第246天, 降水量约为25 mm。7月是整个生长季最干旱的月份, 降水量仅为0.67 mm;VWC的季节动态反映了其对PPT的响应。5至8月中旬VWC整体呈下降趋势, 8月下旬至9月期间由于降水的补给VWC随之增大。10 cm VWC对于降水的响应快于30cm VWC, 但30cm VWC的变幅要明显大于10 cm VWC。其中10 cm VWC平均值0.061 m3/m3, 波动范围0.049—0.089 m3/m3。30cm VWC平均值0.086 m3/m3, 范围为0.059—0.15 m3/m3。

|
| 图 1 生长季(5—9月)环境因子的季节动态 Fig. 1 Seasonal dynamics of environmental factors in May to September during growing season |
图 2反映了生长季5—9月期间, 油蒿叶片发育过程中的环境特征差异。展叶期和落叶期Ta平均值均在15℃左右, 两者之间并无显著差异。完全展叶期Ta平均值为20.86℃, 显著高于展叶期和落叶期37.46%和30.28%;展叶期和完全展叶期PAR平均值分别为460.97 μmol m-2 s-1和442.12 μmol m-2 s-1, 落叶期PAR显著降低至305.23 μmol m-2 s-1;VPD在3个叶物候期均存在显著差异。VPD平均值在完全展叶期最高(1.52 kPa), 落叶期VPD平均值相比展叶期和完全展叶期时降低了33.50%和44.18%;10 cm VWC和30 cm VWC在落叶期平均值最高, 分别为0.082 m3/m3和0.13 m3/m3, 完全展叶期10 cm VWC和30 cm VWC均显著降低至0.054 m3/m3和0.065 m3/m3。10 cm VWC和30 cm VWC平均值在3个物候期间均存在显著差异。综合以上信息可知, 油蒿叶片各物候期的环境特征可描述为, 展叶期温度适中、干燥和强辐射, 完全展叶期高温、强辐射, 并且伴随着严重的土壤水分亏缺, 而落叶期环境特征为温度适中、湿润且辐射低。

|
| 图 2 油蒿不同物候期的环境因子差异特征 Fig. 2 Characteristics of environmental factors in different leaf phenological stages of Artemisia ordosica 图中不同字母代表各不同物候期间环境因子存在显著差异(P < 0.05) |
生长季Pn平均值8.26 μmol m-2 s-1, 最大值15.09 μmol m-2 s-1 (第126天), 最小值1.54 μmol m-2 s-1(第212天;图 3)。完全展叶期Pn相比展叶期和落叶期分别降低39.82%和31.71;PARi波动范围340.77—1239.06 μmol m-2 s-1, 最大值出现在第179天, 最小值为269 d。不同物候期间PARi差异不明显;LUEday平均值0.010 mol/mol, 最大值0.017 mol/mol (第126天), 最小值0.003 mol/mol(第212天)。整个观测周期, LUEday表现出了较大的季节波动, 变异系数为38.75%。从物候期来看, 油蒿叶片在展叶期和落叶期拥有较高的光能利用效率,其值分别高于生长季平均值26.37%和25.73%(图 4)。完全展叶期期间,LUEday呈先降低后增加趋势,LUEday平均值相比生长季平均值降低了17.37%,比展叶期和落叶期时降低30%左右。

|
| 图 3 油蒿叶片气体交换参数的季节动态 Fig. 3 Seasonal dynamics of leaf gas-exchange parameters of Artemisia ordosica 图中虚线将整个生长季划分为展叶期、完全展叶期和落叶期三个物候期, 横坐标所显示的天数即为测量的实际日期(天数) |
由图 4展示了8个叶性状的季节动态。不同叶性状间的季节变异幅度差异较大(图 5), 其中Chl、Car和LNC均表现出较大的季节变异性(CV ≥ 20%), 而LCC和LRWC的变异程度最低(CV < 7%)。结构性状方面, 生长季Lt平均值为0.29 mm, 总体趋势上Lt随着叶龄增加而逐渐增加。展叶期Lt均值显著低于完全展叶期和落叶期, 完全展叶期和落叶期间Lt无显著差异;SLA在展叶期平均值为93.06 cm2/g, 显著低于落叶期平均值(124.48 cm2/g), 完全展叶期时SLA在生长季均值上下波动;LRWC在展叶期和落叶期均维持在75%以上, 完全展叶期LRWC发生了显著降低, 第212天时LRWC达到最小值66.42%。

|
| 图 4 油蒿叶片结构性状和化学性状的季节动态 Fig. 4 Seasonal dynamics of leaf structural traits and biochemical traits of Artemisia ordosica 图中虚线将整个生长季划分为展叶期、完全展叶期和落叶期三个物候期, 横坐标所显示的天数即为测量的实际日期(天数) |

|
| 图 5 不同物候期油蒿叶片LUEday和叶性状指标的相对偏差 Fig. 5 Relative deviation of LUEday and leaf functional traits of Artemisia ordosica in different leaf phenological periods 图中不同字母代表各不同物候期间LUEday和叶性状指标存在显著差异(P < 0.05) |
叶片化学性状方面, Chl在展叶期间快速增加(平均值30.28 μg/cm2), 完全展叶期间Chl波动较小(26.26—31.38 μg/cm2), 落叶期Chl迅速增加, 其平均值显著高于展叶期和完全展叶期(图 5);Chl a/b的变化与Chl相反, 即Chl a/b在展叶初期达到最大值(3.80, 第126天)后逐渐减小, 于完全展叶期末期达到最小值2.66后逐渐增加。展叶期Chl a/b均值显著高于完全展叶期和落叶期;Car的季节趋势与Chl类似, 生长季均值为6.77 μg/cm2, 落叶期Car均值显著高于展叶期和完全展叶期。LNC的季节动态与LRWC和Chl a/b的趋势相似, 其在展叶期和落叶期无显著差异, 完全展叶期LNC平均值发生了显著降低, 低于生长季均值13.10%;LCC在展叶期间维持在较低水平(43.88%), 其平均值显著低于完全展叶期均值, 而落叶期LCC逐渐降低。
2.4 季节尺度LUEday与叶性状间的相关性油蒿LUEday与叶性状参数的相关性结果表明(图 6和图 7), 结构性状方面, LUEday与Lt和SLA之间关系不显著, 但与LRWC显著正相关(R2=0.56, P < 0.001), 表明季节尺度下LUEday随着LRWC的增大而增大;化学性状方面, LUE的变化与Chl、Car和LNC均不相关, 但LUEday与Chl a/b和LNC显著正相关, Chl a/b和LNC分别解释了LUEday季节变化的38%和36%。

|
| 图 6 油蒿叶片LUEday与结构性状间的相关性 Fig. 6 Correlation between LUEday of Artemisia ordosica and leaf structural traits |

|
| 图 7 油蒿LUEday与化学性状间的相关性 Fig. 7 Correlation between LUEday of Artemisia ordosica and leaf biochemical traits |
在一些大型的LUE生产力模型建模方案中, 不同植被类型和区域间的植物LUE取值差异较大[6—8], 对于特定生态系统中, LUE的动态模拟一直是研究热点和难点。本文的田间测量结果发现, 油蒿光能利用效率具有很强的季节变异性, 这与之前在旱生植物的研究结果一致[15, 33—34]。这些基于典型优势种的叶片研究共同强调, 在旱地生态系统中, 采取LUE固定值或者某一时期的测量值估算生态系统生产力可能会产生较大的季节偏差, 例如根据本文的研究结果, 采用叶片成熟期LUE值进行生产力模型建模时, 将会严重低估旱地生态系统的年度生产力。
研究表明油蒿叶片LUEday的季节波动范围为0.003—0.017 mol/mol, 该变化区间在之前高寒地区物种和北方森林物种所报道的LUE变异范围内[35—37], 但就生长季平均值而言, 本文的结果明显低于以往关于油蒿的报道[15, 33]。相关研究表明, 该区域降水特征存在严重的时空分布不均, 干旱年份非生物因子(如土壤水和温度等)对于冠层碳水交换过程和旱生植物生长动态的限制较大[38—39]。本文推测, 这种植物资源利用差异可能是由于场地资源有效性的强烈年际差异引起的, 测量期发生的严重夏季干旱事件一定程度上抑制了油蒿对光能的利用, 导致年度LUEday的下调。
3.2 叶性状的季节变异针对地中海8个木本植物的短期观测实验发现, 夏季干旱诱导下叶片结构性状(LRWC和SLA)均发生了显著降低[19], 在本文的研究中, LRWC的动态变化符合这一描述特征, 但完全展叶期SLA在干旱影响下仍维持在生长季平均水平左右。SLA的差异表现可能与植物的叶片构型特征有关, 油蒿叶片为退化的椭圆肉质叶, 其所允许的叶面积变化幅度较为有限。在叶片发育过程中, 油蒿叶厚度Lt的总体趋势是逐渐增大的(图 4和图 5), 对夏季干旱并不敏感。本文猜测Lt的季节变化更倾向于是一种生长季碳同化量累积的结果, 其对环境的可塑性程度较低, 这一结论需要得到进一步验证。另外, 目前学术界对于叶片化学性状季节变异的认识存在一定分歧, 在没有环境胁迫的条件下, 固氮藤本植物葛根的LNC和Chl季节波动非常小[40], 而针对玉米等作物的研究表明随着LNC和Chl随着叶龄的而增加[41]。本文的结果表明, 在长期干旱环境中, 展叶期Chl逐渐增加而LNC逐渐降低, 完全展叶期两者均呈现出低有效性, 落叶期在持续降水的作用下得到一定恢复。这些结果共同揭示了叶片性状季节变异特征取决于物种、物候学特征、生态系统类型和气候效应的综合影响。
叶片性状间的相互关系为阐明植物适应环境的生物途径提供了便利性, 丰富了人们对于植物生活史策略的理解。许多群落研究均发现空间尺度上叶片性状间存在强烈的关联性[42—44], 本文的研究表明, 生长季叶性状间的这种相关性仍十分显著。LRWC与所有叶片化学性状(Chl、Chl a/b、Car、LNC和LCC)均存在显著相关(表 1), 这说明LRWC是调节油蒿叶片生物化学水平的关键变量, 其与叶片的养分吸收、光合色素合成以及碳同化的运输过程密切相关。另一方面, 有些基于物种的研究指出LNC与SLA存在显著的正相关[45], 本文在个体发育尺度的发现似乎并不支持这一结果, LNC和SLA的不相关与展叶期和完全展叶期时两者的差异化表现有关。
| 变量 Variable |
叶厚度 Leaf thickness/mm |
比叶面积 Specific leaf area/(cm2/g) |
叶片相对 含水量 leaf relative water content/% |
总叶绿素 含量 Total chlorophyll content/ (μg/cm2) |
叶绿素a/b值 Ratio of chlorophyll a and b |
类胡萝卜素 含量Carotenoid content/ (μg/cm2) |
叶氮含量 Leaf nitrogen content/% |
| 比叶面积/(cm2/g) Specific leaf area |
0.40 | ||||||
| 叶片相对含水量/% leaf relative water content |
-0.02 | 0.25 | |||||
| 总叶绿素含量/(μg/cm2) Total chlorophyll content |
0.39 | 0.45 | 0.55* | ||||
| 叶绿素a/b值 Ratio of chlorophyll a and b |
-0.40 | -0.24 | 0.70** | 0.09 | |||
| 类胡萝卜素含量/(μg/cm2) Carotenoid content |
0.38 | 0.37 | 0.57* | 0.97** | 0.23 | ||
| 叶氮含量/% Leaf nitrogen content |
-0.38 | 0.20 | 0.74** | 0.39 | 0.65** | 0.40 | |
| 叶碳含量/% Leaf carbon content |
0.51 | 0.33 | -0.55* | -0.05 | -0.59* | -0.03 | -0.48 |
| *和**分别表示叶性状在P < 0.05和P < 0.01水平显著相关 | |||||||
叶经济谱理论提供了一个很好的方式将叶片性状与叶片资源获取行为联系在一起, 这为深入理解植物组织变化对气候变化的快速响应奠定了基础[17, 25]。目前大部分研究聚焦于叶经济谱在植物组织尺度(如由叶延伸至根、茎器官)和空间尺度(个体、群落及冠层)的理论延伸[43—45], 而从个体发育角度, 叶性状指标对于叶级资源利用动态是否具有指示性作用仍了解甚少。研究表明, LUEday的季节变异与叶性状的波动密切相关, LRWC、Chl a/b和LNC分别解释了LUEday变异的56%和38%和36%, LRWC是反映旱生灌木油蒿LUEday季节波动的关键生物变量。该结果支持了本文最初的假设, 即叶性状能够预测生长季植物在资源获取效率上的动态变化。由于本研究仅限于旱生灌木油蒿这单个物种, 并且考虑到这一研究结果是初步的, 未来扩大物种范围和资源利用效率的指标类型将会是研究工作的重要方向。
本研究中, 尽管LRWC在季节尺度的变异性较低, 但LUEday对于LRWC的变化最为敏感, 这跟长岛混交林栓皮栎光合参数的季节波动与叶片含水量联系紧密的结果相似[20]。这种强耦合可能是由于叶片含水量降低导致光合作用产物(可溶性糖或淀粉)的运输受到阻滞, 过多的同化物积累会对叶片光合产生抑制作用[46—47]。而当叶面大量脱水时, 气孔导度和光合酶数量、活性的降低均会影响叶片净光合速率, 进一步引起资源利用效率的降低[48—49]。另一方面, 叶绿素和叶氮直接参与叶片光能吸收和光化学反应过程[50], 叶片生物化学性状的有效性与植物资源整合的能力密切相关。当光合色素的合成速度低于分解速度时, 色素含量随之降低[51]。Chl a和Chl b之间的相互调节依赖叶绿素循环[52]。本研究中LUEday与光合色素的关系取决于叶绿素a/b值而不是Chl, 说明Chl a在干旱环境中的过度水解一定程度上抑制了植物对光能的吸收, 油蒿叶片在光能吸收和能量转运方面的相对变化影响了其对光能的利用[53]。一些研究表明叶氮通过影响叶片发育过程和生理功能来改变其资源利用效率[10, 54], 本文的研究证实了LNC与LUEday呈显著正相关, 但这种关联性非常弱, 表明叶氮对于LUE的限制可能是次要的。
最近一项干旱胁迫实验表明, 随着土壤水分有效性的降低, 旱生植物沙米的叶片SLA、LRWC和Chl均发生显著降低[55]。本文发现自然环境下油蒿叶片LRWC和LNC受空气温度Ta和土壤含水量VWC的共同调节, 其中油蒿主根系所在的30 cm土壤含水量相比浅层土壤水影响更大, 而Chl a/b的季节波动主要由浅层土壤含水量控制(图 8), 这一结果与之前在同一场地观察到旱生灌木叶氮含量的季节动态主要由土壤水变化驱动的结论较为一致[56]。以上发现强调, 这些能够调节旱生植物资源利用效率动态的关键叶性状对环境温度和土壤水分变化的敏感性较高, 在未来预计极端的气候事件(极端干旱或持续热浪事件)发生更频繁的旱地场景中, 植物叶片性状的适应性调整应当被充分考虑到旱地生态系统的通量建模方案中。

|
| 图 8 叶性状与环境因子间的关系 Fig. 8 Relationship between leaf functional traits and environmental factors |
生长季油蒿叶片LUEday存在较大的季节变异, 完全展叶期LUEday在干旱和高温的影响下发生显著降低;不同叶性状的季节变异幅度差异很大, LRWC虽然季节变异幅度低, 但LUEday对于LRWC的变化最为敏感;与LUEday关系密切的叶性状中, LRWC和LNC受空气温度和土壤含水量的共同调节, 而Chl a/b的季节波动主要由浅层土壤含水量控制。研究结果表明, 在利用光能利用模型进行旱地生态系统生产力估算时, 关键叶性状的相对变化一定程度上可以反映旱生植物的LUEday季节动态。
| [1] |
Potter C S, Davidson E A, Klooster S A, Nepstad D C, De Negreiros G H, Brooks V. Regional application of an ecosystem production model for studies of biogeochemistry in Brazilian Amazonia. Global Change Biology, 1998, 4(3): 315-333. DOI:10.1046/j.1365-2486.1998.00154.x |
| [2] |
李贵才. 基于MODIS数据和光能利用率模型的中国陆地净初级生产力估算研究[D]. 北京: 中国科学院研究生院(遥感应用研究所), 2004.
|
| [3] |
Wu C Y, Chen J M, Desai A R, Hollinger D Y, Arain M A, Margolis H A, Gough C M, Staebler R M. Remote sensing of canopy light use efficiency in temperate and boreal forests of North America using MODIS imagery. Remote Sensing of Environment, 2012, 118: 60-72. DOI:10.1016/j.rse.2011.11.012 |
| [4] |
Lin S R, Coops N C, Tortini R, Jia W, Nesic Z, Beamesderfer E, Arain M A, Li J, Liu Q H. Species and stand-age driven differences in photochemical reflectance index and light use efficiency across four temperate forests. International Journal of Applied Earth Observation and Geoinformation, 2021, 98(4): 102308. |
| [5] |
Yuan W P, Liu S G, Zhou G S, Zhou G Y, Tieszen L L, Baldocchi D, Bernhofer C, Gholz H, Goldstein A H, Goulden M L, Hollinger D Y, Hu Y M, Law B E, Stoy P C, Vesala T, Wofsy S C. Deriving a light use efficiency model from eddy covariance flux data for predicting daily gross primary production across biomes. Agricultural and Forest Meteorology, 2007, 143(3/4): 189-207. |
| [6] |
伍卫星, 王绍强, 肖向明, 于贵瑞, 伏玉玲, 郝彦宾. 利用MODIS影像和气候数据模拟中国内蒙古温带草原生态系统总初级生产力. 中国科学D辑: 地球科学, 2008, 38(8): 993-1004. DOI:10.3321/j.issn:1006-9267.2008.08.008 |
| [7] |
隋雪梅, 辛晓平, 张宏斌, 海全胜, 包刚, 闫瑞瑞, 包玉海. 呼伦贝尔贝加尔针茅草甸草原光能利用率变化规律分析. 中国农业资源与区划, 2013, 34(5): 27-35. |
| [8] |
仇宽彪, 张志强, 康满春, 查同刚, 牛勇, 蔡永茂, 赵广亮. 北京永定河沿河沙地杨树人工林光能利用效率. 生态学报, 2016, 36(6): 1571-1581. |
| [9] |
Green D S, Erickson J E, Kruger E L. Foliar morphology and canopy nitrogen as predictors of light-use efficiency in terrestrial vegetation. Agricultural and Forest Meteorology, 2003, 115(3/4): 163-171. |
| [10] |
同小娟, 李俊, 于强. 农田生态系统光能利用效率及其影响因子分析. 自然资源学报, 2009, 24(8): 1393-1401. DOI:10.3321/j.issn:1000-3037.2009.08.008 |
| [11] |
Li X L, Liang S L, Yu G R, Yuan W P, Cheng X, Xia J Z, Zhao T B, Feng J M, Ma Z G, Ma M G, Liu S M, Chen J Q, Shao C L, Li S G, Zhang X D, Zhang Z Q, Chen S P, Ohta T, Varlagin A, Miyata A, Takagi K, Saiqusa N, Kato T. Estimation of gross primary production over the terrestrial ecosystems in China. Ecological Modelling, 2013, 261-262: 80-92. DOI:10.1016/j.ecolmodel.2013.03.024 |
| [12] |
李成, 马景永, 张财, 王奔, 查天山, 贾昕. 油蒿灌木荒漠光能利用效率季节动态研究. 北京林业大学学报, 2019, 41(9): 99-107. DOI:10.13332/j.1000-1522.20180217 |
| [13] |
康婷婷, 高苹, 居为民, 黄金龙. 江苏省农作物最大光能利用率时空特征及影响因子. 生态学报, 2014, 34(2): 410-420. |
| [14] |
Alton P B, North P R, Los S O. The impact of diffuse sunlight on canopy light-use efficiency, gross photosynthetic product and net ecosystem exchange in three forest biomes. Global Change Biology, 2007, 13(4): 776-787. DOI:10.1111/j.1365-2486.2007.01316.x |
| [15] |
李鑫豪, 闫慧娟, 卫腾宙, 周文君, 贾昕, 查天山. 油蒿资源利用效率在生长季的相对变化及对环境因子的响应. 植物生态学报, 2019, 43(10): 889-898. DOI:10.17521/cjpe.2019.0214 |
| [16] |
李群, 赵成章, 王继伟, 赵连春, 徐婷, 韩玲. 张掖湿地芦苇比叶面积和水分利用效率的关系. 生态学报, 2017, 37(15): 4956-4962. |
| [17] |
Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets V, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821-827. DOI:10.1038/nature02403 |
| [18] |
Grassi G, Vicinelli E, Ponti F, Cantoni L, Magnani F. Seasonal and interannual variability of photosynthetic capacity in relation to leaf nitrogen in a deciduous forest plantation in northern Italy. Tree Physiology, 2005, 25(3): 349-360. DOI:10.1093/treephys/25.3.349 |
| [19] |
Bongers F J, Olmo M, Lopez-Iglesias B, Anten N P R, Villar R. Drought responses, phenotypic plasticity and survival of Mediterranean species in two different microclimatic sites. Plant Biology, 2017, 19(3): 386-395. DOI:10.1111/plb.12544 |
| [20] |
Burnett A C, Serbin S P, Lamour J, Anderson J, Davidson K J, Yang D D, Rogers A. Seasonal trends in photosynthesis and leaf traits in scarlet oak. Tree Physiology, 2021, 41(8): 1413-1424. DOI:10.1093/treephys/tpab015 |
| [21] |
Niinemets V, Tenhunen J D. A model separating leaf structural and physiological effects on carbon gain along light gradients for the shade-tolerant species Acer saccharum. Plant, Cell and Environment, 1997, 20(7): 845-866. DOI:10.1046/j.1365-3040.1997.d01-133.x |
| [22] |
Niinemets V, Valladares F, Ceulemans R. Leaf-level phenotypic variability and plasticity of invasive Rhododendron ponticum and non-invasive Ilex aquifolium co-occurring at two contrasting European sites. Plant, Cell & Environment, 2003, 26(6): 941-956. |
| [23] |
史作民, 唐敬超, 程瑞梅, 罗达, 刘世荣. 植物叶片氮分配及其影响因子研究进展. 生态学报, 2015, 35(18): 5909-5919. |
| [24] |
Limousin J M, Yepez E A, McDowell N G, Pockman W T. Convergence in resource use efficiency across trees with differing hydraulic strategies in response to ecosystem precipitation manipulation. Functional Ecology, 2015, 29(9): 1125-1136. DOI:10.1111/1365-2435.12426 |
| [25] |
陈莹婷, 许振柱. 植物叶经济谱的研究进展. 植物生态学报, 2014, 38(10): 1135-1153. |
| [26] |
Thomas D S G. Arid Zone Geomorphology: Process, Form and Change in Drylands. 3rd ed. Hoboken: John Wiley and Sons, 2011.
|
| [27] |
杨洪晓, 张金屯, 吴波, 李晓松, 张友炎. 毛乌素沙地油蒿种群点格局分析. 植物生态学报, 2006, 30(4): 563-570. DOI:10.3321/j.issn:1005-264X.2006.04.004 |
| [28] |
李思静, 查天山, 秦树高, 钱多, 贾昕. 油蒿(Artemisia ordosica)茎流动态及其环境控制因子. 生态学杂志, 2014, 33(1): 112-118. |
| [29] |
张景波, 张金鑫, 卢琦, 辛智鸣, 李新乐, 刘庆新, 李宏亮. 乌兰布和沙漠油蒿叶片PSⅡ叶绿素荧光动力学参数及其光响应曲线动态. 草业科学, 2019, 36(3): 713-719. |
| [30] |
何莹莹, 于明含, 丁国栋, 高广磊, 刘伟, 周子渊. 油蒿(Artemisia ordosica)幼苗生长及生物量分配对降雨量和降雨间隔的响应. 中国沙漠, 2021, 41(5): 183-191. |
| [31] |
周文君, 查天山, 贾昕, 田赟, 卫腾宙, 靳川. 宁夏盐池油蒿叶片水分利用效率的生长季动态变化及对环境因子的响应. 北京林业大学学报, 2020, 42(7): 98-105. |
| [32] |
王子奇, 查天山, 贾昕, 吴雅娟, 张明艳, 穆家伟. 油蒿光合参数季节动态及其与叶氮含量和比叶面积的关系. 生态学杂志, 2017, 36(4): 916-924. |
| [33] |
Jiang Y, Tian Y, Zha T S, Jia X, Bourque C P A, Liu P, Jin C, Jiang X Y, Li X H, Wei N N, Gao S J. Dynamic changes in plant resource use efficiencies and their primary influence mechanisms in a typical desert shrub community. Forests, 2021, 12(10): 1372. DOI:10.3390/f12101372 |
| [34] |
付涛, 蒋志荣, 鲍婧婷, 朱瑞清, 鲁玉超. 沙坡头地区3种沙生植物水分及光能利用效率特征比较分析. 甘肃农业大学学报, 2015, 50(2): 106-110. DOI:10.3969/j.issn.1003-4315.2015.02.017 |
| [35] |
Tarvainen L, Räntfors M, Wallin G. Seasonal and within-canopy variation in shoot-scale resource-use efficiency trade-offs in a Norway spruce stand. Plant, Cell & Environment, 2015, 38(11): 2487-2496. |
| [36] |
Field C, Mooney H A. Leaf age and seasonal effects on light, water, and nitrogen use efficiency in a California shrub. Oecologia, 1983, 56(2): 348-355. |
| [37] |
周紫鹃, 苏培玺, 侍瑞, 解婷婷. 不同高寒生态系统植物光能利用效率及其环境影响. 生态学杂志, 2017, 36(6): 1570-1577. |
| [38] |
Jia X, Zha T S, Gong J N, Wang B, Zhang Y Q, Wu B, Qin S G, Peltola H. Carbon and water exchange over a temperate semi-arid Shrubland during three years of contrasting precipitation and soil moisture patterns. Agricultural and Forest Meteorology, 2016, 228-229: 120-129. |
| [39] |
Iqbal S, Zha T S, Jia X, Hayat M, Qian D, Bourque C P A, Tian Y, Bai Y J, Liu P, Yang R Z, Khan A. Interannual variation in sap flow response in three xeric shrub species to periodic drought. Agricultural and Forest Meteorology, 2021, 297: 108276. |
| [40] |
Way D A, Stinziano J R, Berghoff H, Oren R. How well do growing season dynamics of photosynthetic capacity correlate with leaf biochemistry and climate fluctuations?. Tree Physiology, 2017, 37(7): 879-888. |
| [41] |
Miner G L, Bauerle W L. Seasonal responses of photosynthetic parameters in maize and sunflower and their relationship with leaf functional traits. Plant, Cell & Environment, 2019, 42(5): 1561-1574. |
| [42] |
宋彦涛, 周道玮, 王平, 李强. 松嫩草地66种草本植物叶片性状特征. 生态学报, 2013, 33(1): 79-88. |
| [43] |
王钊颖, 陈晓萍, 程英, 王满堂, 钟全林, 李曼, 程栋梁. 武夷山49种木本植物叶片与细根经济谱. 植物生态学报, 2021, 45(3): 242-252. |
| [44] |
刘润红, 白金连, 包含, 农娟丽, 赵佳佳, 姜勇, 梁士楚, 李月娟. 桂林岩溶石山青冈群落主要木本植物功能性状变异与关联. 植物生态学报, 2020, 44(8): 828-841. |
| [45] |
宋贺, 于鸿莹, 陈莹婷, 许振柱, 周广胜. 北京植物园不同功能型植物叶经济谱. 应用生态学报, 2016, 27(6): 1861-1869. |
| [46] |
Anadon-Rosell A, Hasibeder R, Palacio S, Mayr S, Ingrisch J, Ninot J M, Nogués S, Bahn M. Short-term carbon allocation dynamics in subalpine dwarf shrubs and their responses to experimental summer drought. Environmental and Experimental Botany, 2017, 141: 92-102. |
| [47] |
邓秀秀, 肖文发, 曾立雄, 雷蕾, 施征. 马尾松幼苗光合产物的运输与分配特征. 林业科学, 2019, 55(7): 27-34. |
| [48] |
安玉艳, 梁宗锁. 植物应对干旱胁迫的阶段性策略. 应用生态学报, 2012, 23(10): 2907-2915. |
| [49] |
Winkler D E, Belnap J, Duniway M C, Hoover D, Reed S C, Yokum H, Gill R. Seasonal and individual event-responsiveness are key determinants of carbon exchange across plant functional types. Oecologia, 2020, 193(4): 811-825. |
| [50] |
许 大全. 光合作用学. 北京: 科学出版社, 2013: 94-95.
|
| [51] |
崔西甜, 袁凤辉, 王安志, 关德新, 吴家兵, 金昌杰. 蒙古栎叶片光合作用随叶龄的变化及其与叶片功能性状的关系. 生态学杂志, 2017, 36(11): 3160-3167. |
| [52] |
Yasumura Y, Hikosaka K, Hirose T. Seasonal changes in photosynthesis, nitrogen content and nitrogen partitioning in Lindera umbellata leaves grown in high or low irradiance. Tree Physiology, 2006, 26(10): 1315-1323. |
| [53] |
Zhang D W, Yuan S, Xu F, Zhu F, Yuan M, Ye H X, Guo H Q, Lv X, Yin Y H, Lin H H. Light intensity affects chlorophyll synthesis during greening process by metabolite signal from mitochondrial alternative oxidase in Arabidopsis. Plant, Cell & Environment, 2016, 39(1): 12-25. |
| [54] |
Albrizio R, Steduto P. Resource use efficiency of field-grown sunflower, sorghum, wheat and chickpea: Ⅰ. Radiation use efficiency. Agricultural and Forest Meteorology, 2005, 130(3/4): 254-268. |
| [55] |
张家琦, 郑冬梅, 朱凯. 干旱胁迫对沙米叶片结构性状与光合色素含量的影响. 林业资源管理, 2022(2): 157-163. |
| [56] |
Chen Z H, Zha T S, Jia X, Wu Y J, Wu B, Zhang Y Q, Guo J B, Qin S G, Chen G P, Peltola H. Leaf nitrogen is closely coupled to phenophases in a desert shrub ecosystem in China. Journal of Arid Environments, 2015, 122: 124-131. |