 2023, Vol. 43
2023, Vol. 43文章信息
- 郭线茹, 刘晓光, 汤清波, 丁识伯, 周洲, 李为争, 马继盛
- GUO Xianru, LIU Xiaoguang, TANG Qingbo, DING Shibo, ZHOU Zhou, LI Weizheng, MA Jisheng
- 植食性昆虫寄主植物嗅觉搜寻述介
- An introduction to phytophagous insect searching for host-plants through olfaction
- 生态学报. 2023, 43(11): 4411-4422
- Acta Ecologica Sinica. 2023, 43(11): 4411-4422
- http://dx.doi.org/10.5846/stxb202202100334
-
文章历史
- 收稿日期: 2022-02-10
- 网络出版日期: 2023-02-07





2. 河南省害虫绿色防控国际a联合实验室, 郑州 450046;
3. 中华人民共和国扬州海关, 扬州 225009;
4. 信阳农林学院农学院, 信阳 464000
2. Henan International Laboratory for Green Pest Control, Zhengzhou 450046, China;
3. Customs of Yangzhou, People's Republic of China, Yangzhou 225009, China;
4. College of Agriculture, Xinyang Agriculture and Forestry University, Xinyang 464000, China
昆虫搜寻其所需资源是主动而重要的生命活动, 所需的资源主要包括食物, 配偶, 产卵、筑巢和藏匿的场所等[1]。这些资源均是实体, 凡实体都包含和/或传递着信息, 昆虫就是借助这些信息搜寻资源的。寄主植物[2]是植食性昆虫-取食植物活体组织或汁液的昆虫-所需的食物, 因此, 搜寻食物就是搜寻寄主植物的活动。
植食性昆虫的寄主搜寻行为具时序性, 时序性分为寄主的栖境定向、定位、接受和利用等4个连续性步骤[3]。其中定向和定位是搜寻过程, 期间起主要作用的是昆虫的嗅觉, 亦兼及视觉;接受和利用是择定过程, 其中味觉及触觉起主导作用[4]。本文仅就植食性昆虫寄主嗅觉搜寻方面的一些资料进行了梳理、总结和讨论, 希冀为相关研究的深入开展、利用植物气味人为调控植食性昆虫寄主选择行为等提供可资参考的资料。
1 植物气味植食性昆虫搜寻寄主的嗅觉媒介是植物气味。植物气味包括构成性气味和诱发性气味这两类气味。构成性气味是指绿色植物正常生长发育进程中释放的挥发性有机物;诱发性气味指的是植物因受昆虫食害、机械致伤、病原物侵染等生物和非生物胁迫因子的影响, 构成性气味中释放量发生显著变化(提高或下降)的有机物和新释放出的构成性气味中不曾出现的有机物。构成性气味和诱发性气味都是植物的次级代谢物。
1.1 构成性气味绿色植物正常生长发育期间会释放多种挥发性有机物, 这些有机物即构成性气味的组分, 大多是分子量在300以下的亲脂性物质, 其中多数是脂肪酸衍生物、类异戊二烯、类苯和类苯丙烷以及一些含氮或硫的化合物。这些物质多是植物地上部营养组织和花释放的, 例如花释放的约1,000种(其中已鉴定的有700多种)[5], 也有些是根释放的[6]。
气味的组分数与构成因植物分类地位而不同。例如禾本科玉米Zea mays、锦葵科陆地棉Gossypium hirsutum、豆科豇豆Vigna unguiculata和埃及车轴草Trifolium alexandrinum以及十字花科甘蓝Brassica oleracea等植物的营养组织释放的气味组分有48种, 这些化合物包括单萜、倍半萜、同源萜(不规则萜)、脂肪酸衍生物以及其他化合物等5大类物质。但不同种类植物的气味组分数及构成差异很大:玉米和陆地棉的气味组分分别有16种和22种, 均见于上述5大类物质中;豇豆和甘蓝分别有17种和14种, 均是单萜最多, 倍半萜阙如, 但α-异佛尔酮和β-异佛尔酮为甘蓝所特有。以上5种植物释放的这些特定组分的气味混合物, 都会引发棉贪夜蛾Spodoptera littoralis成虫触角产生显著不同的反应。除玉米外, 其他4种植物气味混合物组分的比例, 在植物种内是保守的, 棉贪夜蛾成虫就是利用这样的混合物搜寻到寄主的[7];
槭树科槭属的复叶槭Acer negundo、五角枫A. pictum、华北五角枫A. truncatum和挪威槭A. platanoides, 气味组分计有84种, 但它们的共有组分只有乙酸乙酯、2-己烯醛、α-蒎烯、β-蒎烯等8种。复叶槭气味中以酯、醛和烷烃化合物为主, 五角枫以酯和醛为主, 华北五角枫以酯、萜和醇较多, 挪威槭最多的是酯。研究发现, 不同树种对光肩星天牛Anoplophora glabripennis的抗性与其气味组分有关, 感虫的复叶槭气味中未检测到醇和酮, 4种树木中醛、醇和酮的相对含量顺序是复叶槭<五角枫<华北五角枫<挪威槭, 这与4种槭树抗虫性高低顺序相一致, 光肩星天牛即是借助气味中这些组分的含量搜寻这些槭树的[8];
茄科烟草属的黄花烟草Nicotiana rustica、香甜烟草N. suaveolens等9种烟草, 整个生长发育期间营养器官和花释放的气味组分多达120余种, 其中包括单萜和含氧单萜、不规则萜、倍半萜和含氧倍半萜、芳香醇和醛、芳香酯、脂肪酸衍生物以及含氮化合物等。但这些烟草气味组分的数量差异很大:具翼烟草N. alata组分数最多(66种), 蓝格斯多夫烟草N. langsdorffii最少(仅14种), 且后者几乎都是单萜。这些组分的释放在白天特异性不明显, 在夜间具有种特异性, 这种特异性与传粉昆虫烟草天蛾Manduca sexta的活动有关, 利用天蛾授粉的花在夜间释放含氮的醛肟和苯酯。对烟草天蛾成虫具有引诱活性的芳樟醇、橙花叔醇和法尼醇异构体、醛肟和苯酯, 具翼烟草的释放量最大, 而蓝格斯多夫烟草气味中, 有的组分缺乏, 有的释放量少。烟草气味的这种表型变异, 是传粉者和植食性昆虫介导的选择、生物合成途径和共同的进化历史等因素决定的[9]。
气味中具有植物种特征性的组分或组分组合。怀特曼陀罗Datura wrightii和渐狭叶烟草N. attenuata均为烟草天蛾的寄主, 但烟草天蛾偏好选择后者产卵, 这种偏好仅是借助于渐狭叶烟草气味中的3-甲基-2-丁烯-1-醇实现的, 表明这种化合物是渐狭叶烟草的特征性组分[10]。陆地棉、甘蓝、埃及车轴草和豇豆的气味中, 则都具有能表征各自“身份”的组分组合:甘蓝气味组分的特征性组合是香叶烯和1,8-桉叶油素, 陆地棉是反-4,8-二甲基-1,3,7-壬三烯(E-DMNT)和芳樟醇, 埃及车轴草是乙酸顺-3-己烯酯和顺-3-己烯-1-醇, 豇豆是香叶烯、1,8-桉叶油素、顺-β-罗勒烯和反-β-罗勒烯。各组合相关组分的释放速率之间呈显著正相关。由这些特征组分组成的混合物, 均能使棉贪夜蛾成虫产生显著的行为反应[7]。
气味中一些组分的释放具节律性。在夜出性昆虫为传粉媒介的一些植物中, 夜间植株的花香味更强, 如在光/暗导引条件下, 林烟草N. sylvestris和香甜烟草花释放的苯甲醇、苯甲醛和苯甲酸甲酯等主要组分的最大释放量皆出现在暗期[11];晚上开花的夜香树Cestrum nocturnum在暗期开始后2 h花香释放量达到高峰, 正好与传粉的夜出性蛾的取食高峰吻合;陆地棉枝叶构成性气味中的反-β-罗勒烯、E-DMNT和反,反-4,8,12-三甲基-1,3,7,11-十三碳四烯[(E, E)-TMTT]的释放量白天显著高于夜间, 被甜菜夜蛾Spodoptera exigua幼虫食害后, 诱发性气味中萜的释放量也呈现明显的节律性[12];甘蓝中硫代葡萄糖苷含量具有24 h变化的昼夜节律性, 这种节律性除与温度和光照等因素有关外, 还与植食性昆虫的昼夜活动周期有关[13]。
1.2 诱发性气味植物在昆虫食害等胁迫因子的作用下所释放的气味包括构成性和诱发性这两类气味[14]。构成性气味组分, 如顺-3-己烯醛、顺-3-己烯醇和一些储存于溶生腺中的单萜和倍半萜等[14—15], 释放量会在受到食害或机械致伤后快速发生变化, 诱发性气味则释放较晚[14]。诱发性气味组分及构成不仅受胁迫因子的影响, 还与植物本身属性有关。
分类地位不同的植物, 气味的组分与构成差别很大。研究发现, 受同种昆虫食害后, 不同植物释放的气味物质差异很大。如受二斑叶螨Tetranychus urticae食害后, 豆科的大豆Glycine max叶片释放的气味组分有7种, 毒豆Laburnum anagyroides 16种, 刺槐Robinia pseudoacacia 26种, 豇豆9种;茄科的普通烟草N. tabacum 20种, 茄子Solanum melalonga 12种, 曼陀罗Datura stramonium 13种, 甜椒Capsicum annuum 16种;桑科的啤酒花Humulus lupulus和葡萄科的葡萄Vitis vinifera叶释放的气味组分分别有13种和14种[16]。这些组分中, 有些是构成性组分, 有些是新出现的组分。新组分出现是气味“质”的变化, 如豇豆气味中出现的2-丁酮、Z-DMNT、反-β-法尼烯和(E, E)-TMTT等, 甜椒释放的β-榄香烯、右旋大根香叶烯和(E, E)-TMTT等。同时, 构成性气味某些组分的释放量也发生了很大变化, 如葡萄叶被食害后E-DMNT的释放量提高了4倍多[16]。这种“量”变会改变气味组分间的比例, 从而会影响植食性昆虫的寄主搜寻行为。
气味的构成因植物基因型而不同。玉米是棉贪夜蛾的主要寄主, 但棉贪夜蛾幼虫对不同基因型玉米植株气味的诱发性有差异。将棉贪夜蛾幼虫反吐液注射入31个玉米自交系幼苗后, 不同自交系释放的气味组分明显不同, 如在比较重要的23个组分中, 自交系F1852气味中缺乏其他自交系气味中都存在的乙酸顺-3-己烯-1-酯等4种酯, lo3、F7001、F2、CM174、A188和la5pop气味中缺乏在其他自交系中都存在的环异洒剔烯和α-依兰烯。同时, 各自交系气味总释放量也呈现显著差异:F1852、lo4、F476和Du101等的气味释放量较大, 而W401、F7、MIS3、F113、F752等都很小。23个组分中绝对释放量的广义遗传力平均为0.84, 大于0.8的组分多达15个, 表明气味组分的大部分变异是遗传因素决定的。实践中, 即可据此性状辨别自交系的来源, 如根据典型的反-β-石竹烯的释放量差异, 可以区分开欧洲系和美洲系[17]。
气味中的有些组分是由植物属、种特征性成分衍生的。在机械致伤和昆虫食害后, 十字花科植物释放的异硫氰酸酯[13], 百合科葱蒜类植物释放的含硫化合物[18]等, 分别是由非挥发性硫代葡萄糖苷和S-丙烯基半胱氨酸亚砜在植物体内酶的作用下生成的。诱发性气味的这种性状与植物自身防卫密切相关, 如对葱蝇Delia antiqua敏感的洋葱Allium cepa根系分泌物中含硫化合物含量高于抗性品种, 敏感品种释放的产卵引诱物质甲基丙基二硫醚和二丙基二硫醚较多[18]。
气味组分随植物生长发育阶段或器官不同而变化。陆地棉幼苗被谷实夜蛾Helicoverpa zea幼虫食害后, 0—2 h释放的气味组分有18种, 其中构成性组分13种, 诱发性组分5种。这些组分中多为萜和醛。被食害后16—19 h释放的气味组分有22种, 其中构成性组分13种, 诱发性组分9种[19];现蕾株的棉蕾被食害后, 即刻释放出的气味组分有12种, 其中诱发性组分3种(丁酸顺-3-己烯酯、异丁酸顺-3-己烯酯和乙酸顺-3-己烯酯);食害后24 h的气味组分有26种, 其中乙酸己酯、橙花叔醇等为新释放的组分。即刻释放的3种诱发性组分, 也是烟草被烟芽夜蛾Heliothis virescens幼虫食害后夜间释放的特有组分。这些组分可能也像烟草对烟芽夜蛾成虫产卵的驱避作用一样, 驱避谷实夜蛾成虫在其幼虫食害的棉蕾上继续产卵, 以避免种内竞争[20]。
气味释放随昆虫食害方式而不同。白芥Sinapis alba是萝卜蚜Lipaphis erysimi、烟蚜Myzus persicae和菜蛾Plutella xylostella的寄主。白芥韧皮部被两种蚜虫刺吸食害后, 白芥花气味中苯甲醛、茴香醛、水杨酸甲酯等的释放量较之构成性气味显著下降, 而被菜蛾幼虫咀嚼食害后, 这些组分的释放量则未发生变化。有意义的是, 以白芥花气味定向寄主的这两种蚜虫的天敌仍能保持对受害白芥植株的嗅觉定向, 表明刺吸类昆虫取食能使寄主繁殖器官的防卫功能增强。据推测, 防卫功能的提升可能是白芥韧皮部汁液中含有某些信号分子, 蚜虫取食引起韧皮部汁液的移动, 从而引发防卫物质释放[21]。与蚜虫食害方式相同的褐飞虱Nilaparvata lugens和与菜蛾幼虫食害方式相同的二化螟Chilo suppressalis食害水稻Oryza sativa后, 两者单独食害稻株和联合食害稻株以及未受害稻株所释放的气味组分计有61种, 与未受害稻株相比, 双重被害稻株不仅释放的气味组分较多, 而且对二化螟成虫产卵的引诱性也强。由于褐飞虱取食使稻株释放的气味组分构成发生了变化, 稻株对稻螟赤眼蜂Trichogramma japonicum的引诱性下降, 从而降低了二化螟卵被寄生的风险[22]。还有研究发现, 同种昆虫对某种寄主致害方式不同, 也会诱发寄主产生不同的气味物质。如山核桃Juglans mandshurica植株受星天牛Anoplophora chinensis雌虫产卵、取食和幼虫钻蛀致害时, 释放的气味组分数均较未受害株多, 并且未受害株气味中仅有3种组分能引发雄虫产生反应, 而成虫取食株、产卵株和幼虫钻蛀株分别有6、7和5种;气味组分中, 罗勒烯对雌虫的引诱作用较强, 蒎烯和罗勒烯对雄虫的引诱作用较强, 丙烯酸-2-乙基己酯对雌虫具驱避作用[23]。
昆虫产卵影响气味释放。草地贪夜蛾Spodoptera frugiperda产卵会使玉米植株芳樟醇等萜的释放量显著降低。如在玉米叶片受伤处涂抹其幼虫口腔反吐液后, 1.5 h时释放的气味组分有反-2-己烯醛、E-DMNT、吲哚等6种, 而涂抹反吐液且又承接雌虫产卵的叶片, 释放的气味组分只有前述气味中的5种。处理6 h后, 虽然两种处理叶片释放的气味均为10种相同的组分, 但涂抹反吐液又承接雌虫产卵的叶片中, 单萜及同源萜等8种组分的释放量均较未承接产卵的叶片显著下降[24]。萜是一些天敌昆虫定位猎物或寄主的信号, 其释放量减少能提高植食性昆虫的防卫性能[25]。
病原物其或其他微生物定殖会改变气味组分构成。马铃薯Solanum tuberosum叶构成性气味组分有2-己烯-1-醇、庚醛和壬醛等21种, 而受马铃薯卷叶病毒(PLRV)侵染后, 这21种组分总释放量提高了1.9倍;受马铃薯X病毒(PVX)或马铃薯Y病毒(PVY)侵染后, 虽然气味释放总量与构成性气味差异不显著, 但感染PVX的植株, 2-己烯-1-醇和壬醛的释放量分别提高了3倍和5倍, 荜澄茄油烯释放量显著下降;感染PVY的植株, 香叶烯的释放量显著提高, 荜澄茄油烯显著下降[26];拟南芥Arabidopsis thaliana受甘蓝夜蛾Mamestra brassicae幼虫食害后, 释放反-α-香柑油烯、水杨酸甲酯和铃兰醛等诱发性气味组分, 但定殖于拟南芥根际的细菌, 能使这些组分的释放量下降[27]。球孢白僵菌Beauveria bassiana在玉米幼苗根部定殖后, 能使玉米植株对亚洲玉米螟Ostrinia furnacalis产卵具有引诱作用的顺-3-己烯醇和2-乙基己醇的释放量提高, 产卵驱避素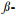
病菌侵染且同时被昆虫食害会改变寄主气味的构成。花生Arachis hypogaea叶释放的构成性气味组分仅有19种, 但受白绢病菌Sclerotium rolfsii侵染同时又被甜菜夜蛾幼虫食害后, 气味组分多达28种(反-2-己烯醛、3-辛酮、橙花叔醇)等, 其中一些组分的释放量, 尤其是3-辛酮(单独受白绢病菌侵染的叶释放的特征性组分)、β-罗勒烯和芳樟醇等, 较之白绢病菌单独侵染时显著提高。而单独受甜菜夜蛾幼虫食害的叶释放的组分中未见3-辛酮、水杨酸甲酯和反-2-己烯醛[29]。
机械致伤、温度、光照、紫外线、湿度等非生物因子也影响植物气味的组分数及构成。如玉米幼苗受植食性昆虫食害产生的诱发性气味物质主要是萜和吲哚, 但它们的释放量随非生物因子而变化:干旱土壤中的释放量大于湿土, 空气相对湿度60%左右时释放量最大, 气温22—27℃时释放量高, 黑暗中则不释放[30];渐狭叶烟草和长花烟草N. longiflora幼苗经紫外线-B处理后, 绿原酸和二咖啡亚精胺的异构体积累提高, 但云香苷的积累只有经紫外线-B诱发才会产生[31]。
2 气味扩散模型自然环境中, 气味物质被风从其释放源头吹散开时, 便会形成气缕。气缕是分子扩散、湍流扩散[32]和平流扩散等这3个物理学过程综合作用的结果。气味扩散状态显著影响昆虫的嗅觉定向行为。
气味扩散状态可以用数学模型表征。气体在空气中扩散的第一个数学模型, 是基于对硝烟自然扩散的研究构建起来的, 这个模型给出了气味源下风向气味平均浓度的估算方程。后来, 地面气缕源扩散状态的半经验方程即Sutton模型被构建出来。该模型假定垂直于气缕中心线所有方向上的化合物平均浓度呈高斯正态分布, 且设定湍流或大气稳定度决定的扩散系数为一常数[32, 33], 则气缕中心线上气味浓度距气味源由近到远时的下降情形符合特定的幂指数定律。
此后, 应用Sutton模型构建了估测昆虫信息化合物在释放源下风向各处浓度的方程:在空气静止的空间, 若设时间t=0时气味化合物Q从笛卡尔直角坐标系的原点瞬间释放, 同时又快速消失, 则其分布密度函数U(mol/cm3)随时间推进而变化的状态可用公式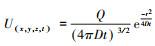
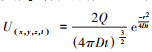
近一二十年来, 一些描述气味扩散的模型陆续刊出, 如以化学信息介导的食根昆虫寄主植物定位的概念模型[34], 基于高斯模型构建的气缕追踪行为模型[35], 植食性昆虫在林木实时气味环境中的气味检测以及气缕追踪最佳路线模型[36]等。
实践中, 表征气味扩散状态时, 应用比较普遍的是预测大气污染物平均浓度的基于高斯气缕分布的模型。该模型假定气味沿x轴平行扩散,且气味全总部被地表反射,则该模型的基本形式是C(x, y, z)=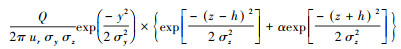
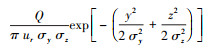
植物生长发育进程中释放气味是正常的生命活动现象。对植食性昆虫来说, 植物气味无论是具驱避作用, 还是具引诱作用, 都会影响成虫和/或幼虫的嗅觉搜寻和定向行为[37]。
3.1 地上植食性昆虫的嗅觉搜寻 3.1.1 成虫搜寻成虫嗅觉搜寻寄主借助于植物气味来完成[38, 39]。植物气味具有特异性, 这种特异性是针对植食性昆虫来说的, 特异性成因有2个可供选择的假说:Ⅰ、植物气味具有植物属种特定的特异性, 形成这种特异性的成分是它类植物所不具有的, 或Ⅱ、植物气味具有特异性, 这种特异性是由广泛存在于植物中的普通成分间的特定比例形成的[40]。
假说Ⅰ所称的特异性气味, 是植物次级代谢物的分解产物, 例如十字花科植物释放的异硫氰酸酯, 百合科葱蒜类释放的硫醚类等, 这些物质在植物受到生物或非生物因子胁迫(如被昆虫食害或机械致伤)后的释放量更多[13, 40]。假说Ⅱ所称的普通成分, 是植物中普遍存在的合成途径形成的, 如不饱和脂肪酸(亚麻酸、亚油酸等)生成所谓绿叶性气味的己烯醛、己醛及相对应的醇类[41], 异戊二烯焦磷酸聚合得到的萜(单萜、倍半萜等)[42]。
对于假说Ⅰ, 支持的证据基本都是来自对异硫氰酸酯的研究。研究表明, 昆虫触角上具有专事调谐这类化合物的嗅感神经元(ORNs), 例如, 十字花科专食者甘蓝蚜Brevicoryne brassicae[43]、种子象Ceutorhynchus assimilis[44]都有这类ORNs。异硫氰酸酯是这两种昆虫搜寻寄主的关键化合物。但不以十字花科植物为寄主的昆虫, 如忽布疣蚜Phorodon humuli和豆卫矛蚜Aphis fabae, 也具有对异硫氰酸酯高度特异的ORNs[43]。研究发现, 有些昆虫能主动回避营养质量不良的寄主或非寄主[45], 因此, 对于上述这两种蚜虫, 异硫氰酸酯是驱避性物质, 可能就是借此避开十字花科植物的。还要注意的是, 在这个假说背景中, 专食者和多食者似应进化出不同的ORNs, 但实际情况并非如此。例如烟青虫Helicoverpa assulta和棉铃虫H. armigera这两近缘种, 前者是寡食性种, 后者是高度多食性种, 但两者鉴别植物气味的ORNs却相似[46];烟青虫、棉铃虫和烟芽夜蛾这3种实夜蛾亚科昆虫成虫触角中, 都存在“气味受体-共受体”OR12/Orco, 用61种挥发性化合物(其中大多数是植物气味组分)进行测试后发现, 3者的OR12/Orco皆被调谐至β-香茅醇、香叶醇、3,7-二甲基-3-辛醇、(-)-芳樟醇、芳樟醇、乙酸反-2-己烯酯等6种挥发性化合物[47], 说明近缘种功能相似的ORNs或气味受体(OR)具有保守性或重现性[46—47]。多食性的白条天蛾Hyles lineata与专食性的节天蛾Sphinx pereglans[48]及多食性的稻纵卷叶螟Cnaphalocrocis medinalis与寡食性的刷须野螟Marasmia patnalis对寄主气味的电生理反应相当[49], 也呈现出同科而食性不同种类嗅觉识别的相似性。
假说Ⅱ是假说Ⅰ的替代性假说。对许多植食性昆虫的电生理学研究发现, 昆虫的外周受体及其参与的信号通路, 能与植物气味中普遍存在的组分结合并被激活[45], 这些组分如脂肪酸衍生物、类苯丙烷和类异戊二烯化合物等[5]。如果昆虫大多数外周受体检测到的气味组分并非是其寄主所特有(这是最常见的), 那么, 植物气味某些组分的比例就成为嗅觉信号的关键特征。这时, 则必须检测特定气味组分的混合物。支持这种认识的实例是对马铃薯叶甲Leptinotarsa decemlineata搜寻行为的研究。普遍存在于植物气味中的反-3-己烯醇、反-2-己烯醇、顺-2-己烯醇和反-2-己烯醛的比例发生些微变化, 就会显著影响马铃薯叶甲成虫的定向行为, 而这些组分非自然比例的混合物对成虫不具引诱活性[50]。
对寄主的嗅觉搜寻, 植物气味的组分混合至关重要, 这是因为混合物具有涌现性-系统具有组成它的单个要素所不具有的特性-这种性状, 会引发昆虫产生不同于单个组分所引发的行为反应, 而昆虫对混合物的感知就是建立在这一特性之上的[51]。因此, 当寄主气味混合物的单个组分独立存在时, 可能不会被昆虫认为是寄主气味混合物的组分而被检测到。例如, 桃Prunus persica新枝气味中的乙酸顺-3-己烯酯、顺-3-己烯醇和苯甲醛, 若以4 ∶ 1 ∶ 1比例混合时, 能引诱梨小食心虫Grapholitha molesta雌虫, 而单独任何一种组分均无引诱活性[52]。而且混合物中各组分须以适当比例存在才会对昆虫产生嗅觉导向作用。如小麦Triticum aestivum花普通气味组分的适当配比, 能引诱寡食性的麦红吸浆虫Sitodiplosis mosellana成虫, 而组分比例不适当的混合物则无引诱性[53];食害葡萄树叶的葡萄花翅小卷蛾Lobesia botrana雌蛾对葡萄植株表现出定向飞行反应, 引发这种行为的关键信号物质是葡萄叶片释放的反-β-石竹烯、反-β-法尼烯和E-DMNT, 三者100 ∶ 78 ∶ 9比例(葡萄树叶气味中的比例)的混合物对雌蛾的引诱效果显著高于37 ∶ 17 ∶ 100比例(非寄主植物苹果树叶气味的比例)的混合物[54]。一般地, 昆虫搜寻寄主植物, 通常利用植物气味中3—10种重要组分[51]。
由上可知, 寄主植物嗅觉搜寻, 或是需检测含有植物种特征性的气味组分, 或是需检测含有某些组分比例特定的混合物, 而后一种情况更为复杂, 因为昆虫既要检测组分的比例, 又要确定它们是否来自寄主植物。自然情况下, 寄主植物栖息地中的其他植物也都会释放气味, 因此昆虫都是生活在许多植物的气味组成的“空气汤”中[45]。如何从空气汤中挑拣出寄主植物的信号, 昆虫对植物气味特异的ORNs提供了一种机制, 通过这种机制, 昆虫可以根据植物气味中普遍存在的某些化合物的比例界定寄主, 因为不同化合物向中枢神经系统(CNS)输入的型式或标记线型不同, 从而可以区分气味组分的比例。但是, 除非ORNs在空间上的位置相同, 否则, 两个差异调谐的ORNs并不能探测出同时到达的不同化合物[45]。研究发现, 触角上同一嗅觉感器中存在着对植物气味产生共同反应的共表达嗅感神经元, 这些神经元有频密协调的功能性结构[44], 这种结构能向CNS输入相关化合物尺度精细的时空分辨率。这种情况在谷实夜蛾对雌性信息素与其拮抗素的感知研究中已得以证实[55]。昆虫具有这种结构, 便赋有了这样一种机制:通过确定两种气味是否以正确的比例出自同一源点, 来鉴别寄主植物特有的气缕[45]。
通过上述这些研究结果可知, 昆虫中成对或聚集的ORNs, 能对气味中普遍存在的化合物进行时空精细分辨率方面的整理, 即进行重合检测。按照重合检测原理, 植食性昆虫能够在弥混着非寄主植物气味的复杂环境中, 通过对植物气味嗅觉信息的中央处理, 而不是对其所做的最初检测, 来鉴别寄主的气味[55]。
3.1.2 幼虫搜寻分析多种鳞翅目昆虫幼虫的寄主搜寻研究结果发现, 幼虫也能够利用植物气味信息趋向或回避植物。葡萄花翅小卷蛾饥饿的幼虫遇到普通气流(对照)时, 做无方向性缓慢爬行运动, 但当遇到葡萄气味(含有水杨酸甲酯、1-己醇、E-DMNT等近10种组分)中的某些组分或其混合物的气流时, 爬行形式即变成近似直线的目标取向性方式。这种化学性靶向位移现象表明, 食物气味中的某些组分能激起饥饿幼虫的取食欲, 且幼虫对气味组分的反应是浓度梯度性的, 因而能有效定向于食物源[56];欧洲玉米螟Ostrinia nubilalis初孵幼虫对其寄主植物玉米与非寄主植物菠菜Spinacea oleraceae的气味反应也明显不同:玉米气味流量40 mL/min与气味脉冲2 s及6 s的组合, 能诱使幼虫产生正趋向反应, 脉冲频率低至9 s和10 s时幼虫会避离, 而对菠菜气味, 气味流量为40 mL/min时, 无论脉冲状态如何, 幼虫都会避离[57]。
被苹果小卷蛾Laspeyresia pomonella幼虫食害5 d后的苹果与未被食害的苹果相比, 食害后的苹果对这种卷蛾初孵幼虫的吸引力大于未被食害的苹果。分析发现, 受害苹果中幼虫引诱素(E, E)-α-法尼烯含量显著高于未受害苹果[58];草地贪夜蛾初孵幼虫在不同处理豇豆幼苗上的扩散行为不同, 留居于被其幼虫食害或经Inceptin处理的植株上的数量, 高于未被幼虫食害株、机械致伤株、新受害株或口腔分泌物中缺乏Inceptin的幼虫食害的植株。Inceptin是豇豆叶绿体三磷酸腺苷(ATP)合成酶部分水解而形成的一种含二硫键的多肽。豇豆叶片被草地贪夜蛾6龄幼虫取食后,Inceptin能触发豇豆苗产生诱发性气味,而且只有先前摄入过含叶绿体组织的幼虫的口腔分泌物中才有这种多肽[59]。草地贪夜蛾幼虫食害能诱发豇豆释放E-DMNT, 用这种化合物处理未受害豇豆植株, 可显著提高幼虫的选择偏好性[60]。玉米苗受草地贪夜蛾幼虫食害6 h后芳樟醇的释放量显著高于未受害苗, 而芳樟醇对第6龄幼虫具显著引诱性[61]。这些结果表明, 食害诱发的气味能改变幼虫对受害植株的嗅觉搜寻行为, 同时也是幼虫搜寻寄主的化学信息。
菊科千里光属的Senecio gregorii与万头菊属的Myriocephalus stuartii均为斑实夜蛾Heliothis punctifera的寄主, 在沙漠中两种植物的斑块状混合分布区, 幼虫在前一种寄主上的丰度高于后一种。室内检验结果表明, 幼虫在这两种植物上的分布不均匀现象, 是幼虫的寄主定向搜寻行为所致, 而且取食经历在定向搜寻中具有重要作用[62]。
上述这些研究表明, 鳞翅目昆虫幼虫依靠嗅觉对植物气味的感知搜寻寄主, 尽管视觉也会参与其中。迄今所知, 幼虫触角和下颚须上的嗅觉感器中, 约有78个神经元投射到触角叶上, 其中有些神经元用来检测植物气味。与成虫相比, 幼虫的嗅觉感器构造相对简单, 但鳞翅目一些昆虫初孵幼虫仍能定向于寄主气味源, 且表现出特定的选择性, 进而搜寻到寄主[57, 63]。
3.2 地下植食性昆虫的嗅觉搜寻地下植食性昆虫搜寻寄主, 与食害植物地上部的昆虫相似, 也主要是利用化学信息[64]。
植物根会释放或分泌初级和次级代谢物[6, 34]。初级代谢物(CO2、氨基酸、糖类等)和次级代谢物都是地下植食性昆虫搜寻定位根系的信息物质, 而次级代谢物往往具有植物种特定性, 正因为如此, 才使昆虫得以区分出寄主植物和非寄主植物[34]。如洋葱的含硫化合物烯丙基硫醚、正丙基硫醚、甲基二硫醚和丙基硫醇、异丙基硫醇等, 对葱蝇幼虫具引诱作用, 这种幼虫正是根据这些信息化合物定位寄主的[37];早竹Phyllostachys praecox竹笋气味组分α--蛇麻烯对筛胸梳爪叩甲Melanotus cribricollis幼虫具有明显的引诱作用, 且引诱作用随α-蛇麻烯浓度增加而增强[65]。
在业经研究的与地下植食性昆虫寄主搜寻有关的74种化合物中, 80%以上的种类能诱使这类昆虫产生行为反应, 其中分子量低的化合物如醇、酯和醛(占化合物总数的37%)多具引诱性, 而碳氢化合物通常有驱避性[34, 64]。搜寻寄主地下组织时, 寡食性昆虫多倾向于利用寄主特异性次级代谢物, 而多食性昆虫大多利用普通的信息化合物[34]。
CO2是根系释放于土壤中最多的气味物质, 其与地下植食性昆虫搜寻寄主的行为密切相关。如西方玉米根叶甲Diabrotica virgifera virgifera对植物根定位的唯一特定物质是CO2[66];专食性的胡萝卜茎蝇Psilae rosae、根瘤象Sitona lepidus、筛胸梳爪叩甲幼虫以及多食性的步甲Evarthrus sodalis, 也都会对CO2产生浓度依赖性趋向反应[64, 65];大麦Hordeum vulgare根系释放的反-2-己烯醛、己醛等4种醛的混合物对叩甲Agriotes sordidus幼虫定向作用会因CO2的缺失而减弱[67];西洋蒲公英Taraxacum sectio ruderalia根系水溶性提取物能“掩蔽”CO2的释放, 从而能规避被五月鳃金龟Melolontha melolontha幼虫发现的风险[68]。
最近的研究表明, 玉米幼苗释放于土壤中的CO2浓度随着距根的距离增大而渐次降低。在距玉米幼苗根32 cm或更近距离释放的西方玉米根叶甲幼虫, 能借助CO2浓度梯度正向搜寻幼苗位置, 而在更远距离释放的幼虫, 其移动是随机的。这种叶甲幼虫有DvvGr1、DvvGr2和DvvGr3等3组CO2受体, 在玉米根际生理活性范围内, 幼虫被富集CO2的环境吸引, 而沉默DvvGr2受体基因则会特异性地抑制幼虫对CO2的响应[69]。但CO2缺乏特异性, 且根系释放入土壤中的CO2的垂直梯度往往比水平梯度更强, 故而无助于土壤植食性昆虫在水平方向上定位寄主[34], 因此在地下植食性昆虫寄主搜寻中, CO2可能充当“搜寻触发器”角色, 或者作为周围存在寄主植物的信号, 从而引导昆虫频密地在其潜在资源区内觅食[64]。
4 结束语植食性昆虫与寄主植物关系的本质是化学。植物构成性气味与诱发性气味之间不存在天然鸿沟。气味的形成与释放, 是植物自身及其与环境因素相互作用的结果, 是复杂的生理生态学现象。植物气味具有多样性, 其构成不仅具有植物科、属、种或品种特性的“化学指纹图”, 而且与环境因子和胁迫因素等密切相关。植物气味之于植食性昆虫及其天敌、乃至气味释放植株的毗邻植株等, 都是一种重要的化学信息[24], 强烈影响它们之间的相互作用, 尤其是植物与植食性昆虫的相互作用[70]。
构成性气味的组分和构成随植物分类地位而差异很大。对分类地位不同或经济价值不同等植物的构成性气味, 至今已进行了许多研究[7—9], 而对食害这些植物的昆虫如何利用这种气味搜寻到寄主的研究相对较少。昆虫食害、病原物侵染、机械致害、气象因素等生物和非生物因子胁迫引发诱发性气味生成和释放的研究, 近二三十年来开展得很多, 其原因与诱发性气味中的某些组分或混合物能招引植食性昆虫的天敌, 以致赋予植物间接防卫功能有关。研究表明, 无论是诱发性气味还是构成性气味, 作用于植食性昆虫时, 它们或单独或共同参与或影响这类昆虫对寄主的嗅觉识别、寄主定向[18]、寄主选择[22]、产卵场所选择[20, 23, 27]、雌雄个体间信息交流[71]、天敌昆虫的寄主定向[21, 72]以及植物的直接防卫或间接防卫[29], 有的还会抑制植物自身的生长发育[73]等。限于本文设定的范围, 对于植物气味的这些生态学以及生理学功能, 文中未予或未多论述。
在寄主搜寻方面, 成虫的搜寻比幼虫重要得多, 因为成虫的搜寻行为关系下代存活乃至种群繁衍问题。植物构成性气味是成虫搜寻寄主的媒介, 对植食性昆虫而言, 这种气味有特异性, 特异性成因有2个假说, 但将植物气味以这样的特异性来划分, 显得有些粗浅。近些年对植物气味的深入研究发现, 植物气味与其分类地位或系统进化程度的吻合度差异较大, 以“特征性挥发物”即“化学指纹图”中的主组分及对昆虫嗅觉感受贡献较大的组分来界定植物气味[74], 或许更为准确和实用。
与成虫相比, 幼虫活动能力有限, 嗅觉搜寻寄主能力较弱, 但嗅觉搜寻在幼虫的寄主转换和植株间转移方面有一定作用[59, 62], 同样应予重视。
地下植食性昆虫寄主搜寻也主要是利用嗅觉。与搜寻行为密切关联的因素是植物地下组织释放或分泌的初级和次级代谢物, 其中作为“搜寻触发器”的CO2, 使这类昆虫的搜寻更为有效[34, 64]。气味物质难以收集和昆虫活动状态不易观察, 是地下植食性昆虫寄主搜寻行为研究的瓶颈, 因此, 结论比较明晰的研究资料比较少。随着气味收集新技术及现代监测手段与观测仪器设备的出现, 这类研究将会深入开展。
植食性昆虫寄主嗅觉搜寻研究中需要特别重视的问题是, 如何将昆虫触角电位、单细胞记录等电生理研究或室内行为研究与野外测试以至实践应用结合起来。电生理研究仅仅是第一步, 但若不与野外测试应用研究结合, 仅凭电生理研究的结果, 还不能确切断言一种气味的某些组分或其混合物与特定昆虫行为反应或电生理反应相对应的因果关系, 因为实施测试时的室内小微环境, 与野外自然环境或特定环境(如仓库)的差别非常大, 在室内观察到的植物气味的某些组分特定比例的混合物能诱使一种昆虫产生趋向行为, 但在野外未必能观察到这样的现象[75], 只有进行多次野外试验, 才能获得准确的可用于实践的结果。相应地, 研究植食性昆虫对植物气味的嗅觉感受机制, 也应与野外试验研究相结合。还需注意的是, 在研究植食性昆虫嗅觉搜寻寄主时, 应重视和借鉴化学生态学在探究物种相互作用、分布、丰度和多样性的成因和结果时所秉持的观念和所采用的机理性研究方法[76], 近年来昆虫系统发育学、比较基因组学、系统生物学、代谢工程学和神经生物学等的发展, 也为拓深植食性昆虫与植物之间的化学信息介导的相互作用研究提供了新技术、新工具, 这也必将有力推动相关研究的系统、深入开展。
致谢: 加拿大康科迪亚大学(Concordia University)陈新博士在文献收集和本文撰写过程中提出诸多建议和修改意见, 中国西北农林科技大学花保祯博士在昆虫名称厘定等方面、河南农业大学那日松博士和杨西发博士在化合物名称厘定方面给予帮助, 在此一并致谢。| [1] |
Bell W J. Searching behavior patterns in insects. Annual Review of Entomology, 1990, 35(1): 447-467. DOI:10.1146/annurev.en.35.010190.002311 |
| [2] |
郭线茹, 李为争, 董钧锋, 丁识伯, 周洲, 宋南, 马继盛. 植食性昆虫寄主植物选择假说述介. 应用昆虫学报, 2021, 58(6): 1245-1256. |
| [3] |
Pentzold S, Burse A, Boland W. Contact chemosensation of phytochemicals by insect herbivores. Natural Product Reports, 2017, 34(5): 478-483. DOI:10.1039/C7NP00002B |
| [4] |
陆宴辉, 张永军, 吴孔明. 植食性昆虫的寄主选择机理及行为调控策略. 生态学报, 2008, 28(10): 5113-5122. DOI:10.3321/j.issn:1000-0933.2008.10.059 |
| [5] |
Knudsen J T, Tollsten L, Bergström L G. Floral scents—a checklist of volatile compounds isolated by head-space techniques. Phytochemistry, 1993, 33(2): 253-280. DOI:10.1016/0031-9422(93)85502-I |
| [6] |
Steeghs M, Bais H P, de Gouw J, Goldan P, Kuster W, Northway M, Fall R, Vivanco J M. Proton-transfer-reaction mass spectrometry as a new tool for real time analysis of root-secreted volatile organic compounds in Arabidopsis. Plant Physiology, 2004, 135(1): 47-58. DOI:10.1104/pp.104.038703 |
| [7] |
Conchou L, Anderson P, Birgersson G. Host plant species differentiation in a polyphagous moth: olfaction is enough. Journal of Chemical Ecology, 2017, 43(8): 794-805. DOI:10.1007/s10886-017-0876-2 |
| [8] |
张风娟, 金幼菊, 陈华君, 武晓颖. 光肩星天牛对4种不同槭树科寄主植物的选择机制. 生态学报, 2006, 26(3): 870-877. |
| [9] |
Raguso R A, Levin R A, Foose S E, Holmberg M W, McDade L A. Fragrance chemistry, nocturnal rhythms and pollination "syndromes" in Nicotiana. Phytochemistry, 2003, 63(3): 265-284. DOI:10.1016/S0031-9422(03)00113-4 |
| [10] |
Späthe A, Reinecke A, Olsson S B, Kesavan S, Knaden M, Hansson B S. Plant species- and status-specific odorant blends guide oviposition choice in the moth Manduca sexta. Chemical Senses, 2013, 38(2): 147-159. DOI:10.1093/chemse/bjs089 |
| [11] |
Loughrin J H, Hamilton-Kemp T R, Andersen R A, Hildebrand D F. Circadian rhythm of volatile emission from flowers of Nicotiana sylvestris and N. suaveolens. Physiologia Plantarum, 1991, 83(3): 492-496. DOI:10.1111/j.1399-3054.1991.tb00125.x |
| [12] |
Loughrin J H, Manukian A, Heath R R, Turlings T C, Tumlinson J H. Diurnal cycle of emission of induced volatile terpenoids by herbivore-injured cotton plant. Proceedings of the National Academy of Sciences of the United States of America, 1994, 91(25): 11836-11840. DOI:10.1073/pnas.91.25.11836 |
| [13] |
Del Carmen Martínez-Ballesta M, Moreno D A, Carvajal M. The physiological importance of glucosinolates on plant response to abiotic stress in Brassica. International Journal of Molecular Sciences, 2013, 14(6): 11607-11625. DOI:10.3390/ijms140611607 |
| [14] |
Röse U S R, Manukian A, Heath R R, Tumlinson J H. Volatile semiochemicals released from undamaged cotton leaves: a systemic response of living plants to caterpillar damage. Plant Physiology, 1996, 111(2): 487-495. DOI:10.1104/pp.111.2.487 |
| [15] |
Elzen G W, Williams H J, Bell A A, Stipanovic R D, Vinson S B. Quantification of volatile terpenes of glanded and glandless Gossypium hirsutum L. cultivars and lines by gas chromatography. Journal of Agricultural and Food Chemistry, 1985, 33(6): 1079-1082. DOI:10.1021/jf00066a015 |
| [16] |
Van Den Boom C E M, Van Beek T A, Posthumus M A, De Groot A, Dicke M. Qualitative and quantitative variation among volatile profiles induced by Tetranychus urticae feeding on plants from various families. Journal of Chemical Ecology, 2004, 30(1): 69-89. DOI:10.1023/B:JOEC.0000013183.72915.99 |
| [17] |
Degen T, Dillmann C, Marion-Poll F, Turlings T C J. High genetic variability of herbivore-induced volatile emission within a broad range of maize inbred lines. Plant Physiology, 2004, 135(4): 1928-1938. DOI:10.1104/pp.104.039891 |
| [18] |
Ikeshoji T. S-propenylcysteine sulfoxide in exudates of onion roots and its possible decompartmentalization in root cells by bacteria into attractant of the onion maggot, Hylemya antiqua (Diptera: Anthomyiidae). Applied Entomology and Zoology, 1984, 19(2): 159-169. DOI:10.1303/aez.19.159 |
| [19] |
McCall P J, Turlings T C J, Loughrin J, Proveaux A T, Tumlinson J H. Herbivore-induced volatile emissions from cotton (Gossypium hirsutum L.) seedlings. Journal of Chemical Ecology, 1994, 20(12): 3039-3050. DOI:10.1007/BF02033709 |
| [20] |
Röse U S R, Tumlinson J H. Volatiles released from cotton plants in response to Helicoverpa zea feeding damage on cotton flower buds. Planta, 2004, 218(5): 824-832. DOI:10.1007/s00425-003-1162-9 |
| [21] |
Pareja M, Qvarfordt E, Webster B, Mayon P, Pickett J, Birkett M, Glinwood R. Herbivory by a phloem-feeding insect inhibits floral volatile production. PLoS One, 2012, 7(2): e31971. DOI:10.1371/journal.pone.0031971 |
| [22] |
Liu Q S, Hu X Y, Su S L, Ning Y S, Peng Y F, Ye G Y, Lou Y G, Turlings T C J, Li Y H. Cooperative herbivory between two important pests of rice. Nature Communications, 2021, 12(1): 6772. DOI:10.1038/s41467-021-27021-0 |
| [23] |
马艳, 史黎央, 赵艺, 徐华潮. 星天牛不同为害状态下山核桃挥发物成分的GC-EAD和行为反应. 应用昆虫学报, 2019, 56(3): 530-538. |
| [24] |
Peñaflor M F G V, Erb M, Robert C A M, Miranda L A, Werneburg A G, Dossi F C A, Turlings T C J, Bento J M S. Oviposition by a moth suppresses constitutive and herbivore-induced plant volatiles in maize. Planta, 2011, 234(1): 207-215. DOI:10.1007/s00425-011-1409-9 |
| [25] |
Schnee C, Köllner T G, Held M, Turlings T C J, Gershenzon J, Degenhardt J. The products of a single maize sesquiterpene synthase form a volatile defense signal that attracts natural enemies of maize herbivores. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(4): 1129-1134. DOI:10.1073/pnas.0508027103 |
| [26] |
Eigenbrode S D, Ding H J, Shiel P, Berger P H. Volatiles from potato plants infected with potato leafroll virus attract and arrest the virus vector, Myzus persicae (Homoptera: Aphididae). Proceedings of the Royal Society B: Biological Sciences, 2002, 269(1490): 455-460. DOI:10.1098/rspb.2001.1909 |
| [27] |
Pangesti N, Weldegergis B T, Langendorf B, van Loon J J A, Dicke M, Pineda A. Rhizobacterial colonization of roots modulates plant volatile emission and enhances the attraction of a parasitoid wasp to host-infested plants. Oecologia, 2015, 178(4): 1169-1180. DOI:10.1007/s00442-015-3277-7 |
| [28] |
付军. 球孢白僵菌(Beauveria bassiana)对亚洲玉米螟(Ostrinia furnacalis)产卵选择行为的调控机制[D]. 长春: 东北师范大学, 2018.
|
| [29] |
Cardoza Y J, Alborn H T, Tumlinson J H. In vivo volatile emissions from peanut plants induced by simultaneous fungal infection and insect damage. Journal of Chemical Ecology, 2002, 28(1): 161-174. DOI:10.1023/A:1013523104853 |
| [30] |
Gouinguene S P, Turlings T C J. The effects of abiotic factors on induced volatile emissions in corn plants. Plant Physiology, 2002, 129(3): 1296-1307. DOI:10.1104/pp.001941 |
| [31] |
Izaguirre M M, Mazza C A, Svatoš V A, Baldwin I T, Ballare C L. Solar ultraviolet-B radiation and insect herbivory trigger partially overlapping phenolic responses in Nicotiana attenuata and Nicotiana longiflora. Annals of Botany, 2007, 99(1): 103-109. DOI:10.1093/aob/mcl226 |
| [32] |
Murlis J, Elkinton J S, Cardé R T. Odor plumes and how insects use them. Annual Review of Entomology, 1992, 37(1): 505-532. DOI:10.1146/annurev.en.37.010192.002445 |
| [33] |
Bossert W H, Wilson E O. The analysis of olfactory communication among animals. Journal of Theoretical Biology, 1963, 5(3): 443-469. DOI:10.1016/0022-5193(63)90089-4 |
| [34] |
Johnson S N, Gregory P J. Chemically-mediated host-plant location and selection by root-feeding insects. Physiological Entomology, 2006, 31(1): 1-13. DOI:10.1111/j.1365-3032.2005.00487.x |
| [35] |
Grünbaum D, Willis M A. Spatial memory-based behaviors for locating sources of odor plumes. Movement Ecology, 2015, 3(1): 11. DOI:10.1186/s40462-015-0037-6 |
| [36] |
Shao X L, Cheng K, Wang Z W, Zhang Q, Yang X T. Use of odor by host-finding insects: the role of real-time odor environment and odor mixing degree. Chemoecology, 2021, 31(3): 149-158. DOI:10.1007/s00049-021-00342-8 |
| [37] |
Sutherland O R W. Plant chemicals influencing insect behaviour. New Zealand Entomologist, 1977, 6(3): 222-228. DOI:10.1080/00779962.1977.9722253 |
| [38] |
江丽容, 刘守安, 韩宝瑜, 欧阳立明. 7种寄主和非寄主植物气味对茶尺蠖成虫行为的调控效应. 生态学报, 2010, 30(18): 4993-5000. |
| [39] |
盛子耀, 李为争, 原国辉. 植物气味多样性与昆虫关系的研究进展. 应用昆虫学报, 2019, 56(4): 652-661. |
| [40] |
Visser J H. Host odor perception in phytophagous insects. Annual Review of Entomology, 1986, 31(1): 121-144. DOI:10.1146/annurev.en.31.010186.001005 |
| [41] |
Visser J H, Van Straten S, Maarse H. Isolation and identification of volatiles in the foliage of potato, Solanum tuberosum, a host plant of the Colorado beetle, Leptinotarsa decemlineata. Journal of Chemical Ecology, 1979, 5(1): 13-25. DOI:10.1007/BF00987684 |
| [42] |
Ma W C, Visser J H. Single unit analysis of odour quality coding by the olfactory antennal receptor system of the Colorado beetle. Entomologia Experimentalis et Applicata, 1978, 24(3): 520-533. DOI:10.1111/j.1570-7458.1978.tb02813.x |
| [43] |
Nottingham S F, Hardie J, Dawson G W, Hick A J, Pickett J A, Wadhams L J, Woodcock C M. Behavioral and electrophysiological responses of aphids to host and nonhost plant volatiles. Journal of Chemical Ecology, 1991, 17(6): 1231-1242. DOI:10.1007/BF01402946 |
| [44] |
Blight M M, Pickett J A, Wadhams L J, Woodcock C M. Antennal perception of oilseed rape, Brassica napus (Brassicaceae), volatiles by the cabbage seed weevil Ceutorhynchus assimilis (Coleoptera, Curculionidae). Journal of Chemical Ecology, 1995, 21(11): 1649-1664. DOI:10.1007/BF02033667 |
| [45] |
Bruce T J A, Wadhams L J, Woodcock C M. Insect host location: a volatile situation. Trends in Plant Science, 2005, 10(6): 269-274. DOI:10.1016/j.tplants.2005.04.003 |
| [46] |
Stranden M, Røstelien T, Liblikas I, Almaas T J, Borg-Karlson A K, Mustaparta H. Receptor neurones in three heliothine moths responding to floral and inducible plant volatiles. Chemoecology, 2003, 13(3): 143-154. DOI:10.1007/s00049-003-0242-4 |
| [47] |
Cao S, Liu Y, Guo M B, Wang G R. A conserved odorant receptor tuned to floral volatiles in three Heliothinae species. PLoS One, 2016, 11(5): e0155029. DOI:10.1371/journal.pone.0155029 |
| [48] |
Raguso R A, Light D M. Electroantennogram responses of male Sphinx perelegans hawkmoths to floral and 'green-leaf volatiles'. Entomologia Experimentalis et Applicata, 1998, 86(3): 287-293. DOI:10.1046/j.1570-7458.1998.00291.x |
| [49] |
Ramachandran R, Khan Z R, Caballero P, Juliano B O. Olfactory sensitivity of two sympatric species of rice leaf folders (Lepidoptera: Pyralidae) to plant volatiles. Journal of Chemical Ecology, 1990, 16(9): 2647-2666. DOI:10.1007/BF00988076 |
| [50] |
Visser J H, Avé D A. General green leaf volatiles in the olfactory orientation of the Colorado beetle, Leptinotarsa decemlineata. Entomologia Experimentalis et Applicata, 1978, 24(3): 738-749. DOI:10.1111/j.1570-7458.1978.tb02838.x |
| [51] |
Bruce T J A, Pickett J A. Perception of plant volatile blends by herbivorous insects-finding the right mix. Phytochemistry, 2011, 72(13): 1605-1611. DOI:10.1016/j.phytochem.2011.04.011 |
| [52] |
Natale D, Mattiacci L, Hern A, Pasqualini E, Dorn S. Response of female Cydia molesta (Lepidoptera: Tortricidae) to plant derived volatiles. Bulletin of Entomological Research, 2003, 93(4): 335-342. DOI:10.1079/BER2003250 |
| [53] |
Birkett M A, Bruce T J A, Martin J L, Smart L E, Oakley J, Wadhams L J. Responses of female orange wheat blossom midge, Sitodiplosis mosellana, to wheat panicle volatiles. Journal of Chemical Ecology, 2004, 30(7): 1319-1328. DOI:10.1023/B:JOEC.0000037742.05022.9f |
| [54] |
Tasin M, Bäckman A C, Bengtsson M, Ioriatti C, Witzgall P. Essential host plant cues in the grapevine moth. Naturwissenschaften, 2006, 93(3): 141-144. DOI:10.1007/s00114-005-0077-7 |
| [55] |
Baker T C, Fadamiro H Y, Cosse A A. Moth uses fine tuning for odour resolution. Nature, 1998, 393(6685): 530-530. DOI:10.1038/31131 |
| [56] |
Becher P G, Guerin P M. Oriented responses of grapevine moth larvae Lobesia botrana to volatiles from host plants and an artificial diet on a locomotion compensator. Journal of Insect Physiology, 2009, 55(4): 384-393. DOI:10.1016/j.jinsphys.2009.01.006 |
| [57] |
Piesik D, Rochat D, van der Pers J, Marion-Poll F. Pulsed odors from maize or spinach elicit orientation in European corn borer neonate larvae. Journal of Chemical Ecology, 2009, 35(9): 1032-1042. DOI:10.1007/s10886-009-9676-7 |
| [58] |
Landolt P J, Brumley J A, Smithhisler C L, Biddick L L, Hofstetter R W. Apple fruit infested with codling moth are more attractive to neonate codling moth larvae and possess increased amounts of (E, E)-α-farnesene. Journal of Chemical Ecology, 2000, 26(7): 1685-1699. DOI:10.1023/A:1005595014589 |
| [59] |
Schmelz E A, Carroll M J, LeClere S, Phipps S M, Meredith J, Chourey P S, Alborn H T, Teal P E A. Fragments of ATP synthase mediate plant perception of insect attack. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(23): 8894-8899. DOI:10.1073/pnas.0602328103 |
| [60] |
Carroll M J, Schmelz E A, Teal P E A. The attraction of Spodoptera frugiperda neonates to cowpea seedlings is mediated by volatiles induced by conspecific herbivory and the elicitor inceptin. Journal of Chemical Ecology, 2008, 34(3): 291-300. DOI:10.1007/s10886-007-9414-y |
| [61] |
Carroll M J, Schmelz E A, Meagher R L, Teal P E A. Attraction of Spodoptera frugiperda larvae to volatiles from herbivore-damaged maize seedlings. Journal of Chemical Ecology, 2006, 32(9): 1911-1924. DOI:10.1007/s10886-006-9117-9 |
| [62] |
Cunningham J P, Lange C L, Walter G H, Zalucki M P. Host location behaviour in the desert caterpillar, Heliothis punctifera. Entomologia Experimentalis et Applicata, 2011, 141(1): 1-7. DOI:10.1111/j.1570-7458.2011.01163.x |
| [63] |
Binyameen M, Ali Q, Roy A, Schlyter F. Plant volatiles and their role in insect olfaction//Singh I K, Singh A, eds. Plant-Pest Interactions: From Molecular Mechanisms to Chemical Ecology. Singapore: Springer, 2021: 127-156.
|
| [64] |
Johnson S N, Nielsen U N. Foraging in the dark-chemically mediated host plant location by belowground insect herbivores. Journal of Chemical Ecology, 2012, 38(6): 604-614. DOI:10.1007/s10886-012-0106-x |
| [65] |
孟海林. 竹林金针虫寄主搜寻机制初步研究[D]. 北京: 中国林业科学研究院, 2017.
|
| [66] |
Bernklau E J, Bjostad L B. Reinvestigation of host location by western corn rootworm larvae (Coleoptera: Chrysomelidae): CO2 is the only volatile attractant. Journal of Economic Entomology, 1998, 91(6): 1331-1340. DOI:10.1093/jee/91.6.1331 |
| [67] |
Barsics F, Delory B M, Delaplace P, Francis F, Fauconnier M L, Haubruge É, Verheggen F J. Foraging wireworms are attracted to root-produced volatile aldehydes. Journal of Pest Science, 2017, 90(1): 69-76. DOI:10.1007/s10340-016-0734-y |
| [68] |
Reinecke A, Müller F, Hilker M. Attractiveness of CO2 released by root respiration fades on the background of root exudates. Basic and Applied Ecology, 2008, 9(5): 568-576. DOI:10.1016/j.baae.2007.10.002 |
| [69] |
Arce C C, Theepan V, Schimmel B C, Jaffuel G, Erb M, Machado R A. Plant-associated CO2 mediates long-distance host location and foraging behaviour of a root herbivore. eLife, 2021, 10: e65575. DOI:10.7554/eLife.65575 |
| [70] |
Krieger J, Breer H. Olfactory reception in invertebrates. Science, 1999, 286(5440): 720-723. DOI:10.1126/science.286.5440.720 |
| [71] |
McCormick A C, Heyer J, Sims J W, Mescher M C, De Moraes C M. Exploring the effects of plant odors, from tree species of differing host quality, on the response of Lymantria dispar males to female sex pheromones. Journal of Chemical Ecology, 2017, 43(3): 243-253. DOI:10.1007/s10886-017-0825-0 |
| [72] |
De Lange E S, Laplanche D, Guo H J, Xu W, Vlimant M, Erb M, Ton J, Turlings T C J. Spodoptera frugiperda caterpillars suppress herbivore-induced volatile emissions in maize. Journal of Chemical Ecology, 2020, 46(3): 344-360. DOI:10.1007/s10886-020-01153-x |
| [73] |
宁可真, 袁学松, 杨忠仁, 郝丽珍, 张凤兰, 孙晶洁. 邻苯二甲酸和肉桂酸对沙芥属植物生长及保护酶活性的影响. 西北农林科技大学学报: 自然科学版, 2021, 49(11): 97-104. |
| [74] |
付国需, 李为争, 刘珂, 王莫慧, 安靖靖, 吴少英, 原国辉. 不同科芳香植物特征挥发物的生源及其对昆虫行为的影响//雷朝亮, 王满囷. 华中昆虫研究(第五卷). 武汉: 湖北科学技术出版社, 2008.
|
| [75] |
方宇凌, 张钟宁. 植物气味化合物对棉铃虫产卵及田间诱蛾的影响. 昆虫学报, 2002, 45(1): 63-67. DOI:10.16380/j.kcxb.2002.01.012 |
| [76] |
Raguso R A, Agrawal A A, Douglas A E, Jander G, Kessler A, Poveda K, Thaler J S. The raison d′être of chemical ecology. Ecology, 2015, 96(3): 617-630. DOI:10.1890/14-1474.1 |