 2023, Vol. 43
2023, Vol. 43文章信息
- 吕凯, 诸爽爽, 谷磊, 孙运菲, 张露, 杨州
- LÜ Kai, ZHU Shuangshuang, GU Lei, SUN Yunfei, ZHANG Lu, YANG Zhou
- 舍曲林对蚤状溞诱导型反捕食防御代内与代际可塑性的干扰效应
- Interference effect of sertraline on phenotypic plasticity within- and trans-generations of induced anti-predator defense in Daphnia pulex
- 生态学报. 2023, 43(11): 4604-4618
- Acta Ecologica Sinica. 2023, 43(11): 4604-4618
- http://dx.doi.org/10.5846/stxb202201290276
-
文章历史
- 收稿日期: 2022-01-29
- 网络出版日期: 2023-02-17




表型可塑性指同一基因型的物种应对异质生境而表达多样化表型性状的能力。表型可塑性是生物适应环境变化的基本能力之一, 因此受到生态学领域研究的广泛关注[1—3]。很多研究证实, 环境变化能够诱导生物在生长发育过程中性状的改变, 这属于代内可塑性或者发育可塑性。常见的生态因子(如温度、资源以及竞争)都会诱导个体的大小、寿命以及繁殖等性状发生改变[4—5]。除此之外, 环境变化还能导致表型变化持续多个世代。另一方面, 亲代的环境经历影响下一子代甚至连续多个子代的表型, 这一现象称为代际可塑性。代际可塑性既可以表现为亲代经历的环境胁迫降低了子代的适合度, 进而可能增加种群消亡的风险[6—7];也可以表现为亲代经历胁迫环境后, 产下的子代具有较强的胁迫耐受能力[8—10]。代内/代际表型可塑性机制赋予生物在不改变遗传信息的情况下对环境波动做出快速适应性响应的能力, 因此代内/代际表型可塑性的方向以及程度对于种群的维持具有重要的影响[11]。
枝角类是淡水湖泊池塘常见的浮游动物类群, 也是淡水食物网高营养级生物的主要捕食对象[12—13]。当面对捕食者时, 枝角类可以产生多样化的诱导型防御响应特征, 概括地分为形态、行为和生活史变化三个方面。由于枝角类的诱导型防御在维持枝角类与捕食者的种间关系平衡中起着重要作用, 因此“捕食者-枝角类”是研究水域生态系统中种间关系的良好模型。在一个世代内, 枝角类通过加速生长、提前产卵、后代数增多、生殖模式转变为有性生殖等生活史特征变化, 体现了代内表型可塑性响应来自鱼类的捕食风险[14—15]。在代际可塑性方面, 亲代受到鱼类捕食风险影响后, 其产下的子代表现出更强烈的诱导型防御特征[16]。然而, 也有报道认为, 在枝角类受到捕食风险胁迫的同时, 如果环境中存在其它胁迫因素, 如温度升高[17]、溶解氧下降[18]、食物质量不佳[19—20]、杀虫剂污染[21], 反而干扰枝角类诱导型防御的正常表达。因此, 这些诱导型防御结果的多样性表明, 需要进一步开展枝角类与捕食者相互作用研究并且加强对表型可塑性代内代际作用的认识。
由于诱导型防御表达在一定程度上受到自身生理代谢稳态调节, 因此, 普遍认为枝角类的诱导型防御特征容易受到外界环境胁迫影响[22—23]。一个潜在的可塑性干扰因素是人类活动排放的化学药品污染, 尤其是具有调节生物神经信号传导作用的抗抑郁类药品。现代生活节奏的加快与压力的加大, 失眠、抑郁和焦虑等疾病的发病率逐年增加, 使得抑郁症这类疾病受到人们的重视, 抗抑郁药的年使用量也逐渐增加[24], 其中舍曲林是世界上最常用的抗抑郁药之一。自然水体中舍曲林的浓度范围从1 ng/L到0.6 μg/L[25—26], 但在未经处理的污水中达到17 μg/L[27]。此外, 也有研究报道舍曲林在地表水中的浓度为33—49 μg/L[28]。虽然舍曲林在水环境中的浓度很低, 但是由于其较高的生物活性和难以被降解的特性, 仍然会对水域生态系统中的浮游生物产生影响[29]。例如, 舍曲林会降低小球藻中叶绿素a浓度从而降低光合效率;舍曲林还会刺激特定蓝藻的生长, 干扰蓝藻群落之间的平衡[30—31]。对于枝角类, 舍曲林减慢其生长和游动速度[32]。另外, 枝角类连续两代暴露于舍曲林促进亲代(F1)的繁殖量, 然而却抑制子代(F2)繁殖量, 说明舍曲林会干扰连续谱系的繁殖输出稳定性[33]。
枝角类诱导型反捕食防御(以下简称“诱导型防御”)是由触角化学感知捕食者信息素而触发, 随后包括诸如神经元信号整合和转化为内分泌物质等一系列内部生理反应。参与诱导型防御表型可塑性的主要关键物质是乙酰胆碱[34]和多巴胺[35]。多巴胺可以调节保幼激素的合成和降解, 从而控制枝角类发育繁殖等性状的多态性。另一方面, 舍曲林具有直接抑制多巴胺再摄取的作用而提高多巴胺功能, 因此对枝角类的诱导型防御有影响[36]。尽管舍曲林对角突网纹溞没有直接的致死效应, 但是会抑制角突网纹溞的诱导型防御在形态方面的表达, 进而导致捕食者-浮游动物种群动态发生改变[37]。
由于舍曲林在环境中持续存在, 而枝角类发育快、世代周期短, 导致枝角类连续多个世代可能都会受到舍曲林的影响, 因此需要评估连续多代暴露于舍曲林对枝角类反捕食防御的影响[33]。基于上述分析, 本研究提出的科学问题是:连续2个世代暴露于舍曲林是否会干扰枝角类诱导型防御代内/代际可塑性?通过回答这一科学问题, 将揭示以舍曲林为代表的神经活性药物残留对浮游动物诱导型防御的长期影响, 同时强调生物表型可塑性在“生态-演化-人类社会相互关系”中的作用。
1 材料与方法 1.1 实验材料本研究使用的蚤状溞(Daphnia pulex)采自江苏镇江某池塘, 经鉴定分离后, 在稳定的实验室条件下孤雌单克隆培养。采用枝角类通用COMBO培养基, 每周更换3—5次培养基, 每天定时定量投喂足够的食物(蛋白核小球藻, 浓度为1.5 mg C/L)以保持种群稳定, 便于后续实验的开展。培养温度为25℃, 光照强度为500 Lux, 光暗比为14 h∶10 h(L∶D)。
使用BG-11藻类培养基获得蛋白核小球藻(Chlorella pyrenoidosa), 无菌培养至对数期, 经离心(4000×g, 15 min), 后用分光光度计测定藻类吸光度(OD)值, 换算为浓度后作为食物储存于4 ℃冷藏保存。
舍曲林购自源叶生物(中国上海), 化学品注册号(CAS)为79559-97-0, 纯度>99%。使用前, 用超纯水溶解, 未使用其他载体溶剂。
鱼类信息素物质尚无分离确定, 国内外通用做法是采用添加滤液的方式[15, 18, 21]。鱼类信息素(以下简称“信息素”)的制备方法:高体鳑鲏鱼在我国自然水体中分布广泛且易获得, 因此本研究中采用野生鳑鲏鱼来制备信息素。获得的野生鳑鲏鱼在25 ℃、500 Lux光照强度和14 h∶10 h的光暗比下进行实验室驯养, 在实验室保存过程中每周更新一次养殖水, 每天定时投喂饲料红虫, 以保证鳑鲏鱼的活力。信息素的制备过程参考先前的研究[19], 具体如下:
(1) 在开始制备信息素前对其进行饥饿处理24 h, 以确保肠道内不再含有其他物质;
(2) 挑选20条活力较好、体长均为3—5 cm的鳑鲏鱼在6 h内投喂6000只蚤状溞;
(3) 随后将其放入20 L新制备的COMBO培养基中, 在不投喂任何食物的情况下培养18 h;
(4) 将培养溶液用0.22 μm玻璃纤维过滤器(Millipore)过滤而获得信息素, 将其分装于无菌瓶中, 储存于-20 ℃冰箱;
(5) 实验时将信息素稀释20倍, 即1条鱼/10 L的浓度。
1.2 实验方法F1代实验设置了三个舍曲林浓度, 分别为:0、0.2和20 μg/L, 并设置对照组(C)和信息素(F)处理组, 共计6个不同的处理, 每个处理5个重复, 将每个处理下F1代的第三窝幼体进行F2代两部分实验。实验1(长期暴露于舍曲林对诱导型防御代内可塑性的影响):分别将F1代C-0、C-0.2和C-20组产下的第三窝(F1代暴露的第9天)子代分为两个实验组:与F1代C-0、C-0.2、C-20组相同的暴露条件, 标记为CC-0、CC-0.2和CC-20;在对照组的基础上添加信息素, 标记为CF-0、CF-0.2和CF-20。实验2(长期暴露于舍曲林对诱导型防御代际可性的影响):分别将F1代F-0、F-0.2和F-20组产下的第三窝(F1代暴露的第9天)子代分为两个实验组:与F1代F-0、F-0.2、F-20组相同的暴露条件, 标记为FF-0、FF-0.2和FF-20;将3个舍曲林浓度的FF去除信息素, 标记为FC-0、FC-0.2和FC-20。实验处理如图 1。

|
| 图 1 实验处理流程图 Fig. 1 Schematic diagram of the experiment “C”和“F”分别表示对照组(C)和信息素(F)处理组; “0”、“0.2”、“20”分别表示舍曲林的浓度为: 0、0.2和20 μg/L |
实验在250 mL玻璃烧杯中进行, 每个烧杯中含200 mL COMBO培养基并放置5只蚤状溞刚出生幼体进行实验, 每天更换新鲜培养基和食物, 食物浓度为1.5 mg C/L蛋白核小球藻。各处理组放置于500 Lux光照培养箱中进行培养, 光暗比(L∶D)设置为14 h∶10 h。
1.3 研究参数的定义与计算方法F1代和F2代均测量性成熟和第9天(所有实验溞都在9天内产完第三窝)时的体长和尾刺长、记录个体幼溞数(随即从烧杯中移除)、繁殖时间。根据公式[38]计算相对尾刺长、种群净繁殖率(R0)、世代时间(T)以及种群内禀增长率(r):
净繁殖率: 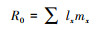
世代时间: 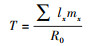
其中x为特定龄期(天), lx是特定龄期的存活率, mx是特定龄期的繁殖量。根据Euler-Lotka方程迭代估算种群的内禀增长率“r“:1=∫e-rxlxmxdx。
相对尾刺长的计算公式:相对尾刺长=尾刺长/体长;使用相对变化率定量分析不同处理下的响应变化情况:变化率=(T-C)/C, 其中C和T分别表示在对照和信息素处理下的个体防御特征值。
1.4 数据分析采用Sigmaplot 14.0对实验数据进行统计学分析以及作图展示。所有数据均使用平均值±标准误(Mean ± SE)的形式进行表示。使用双因素方差分析评估F1代中舍曲林和信息素对所有特征的交互作用, 然后进行Tukey检验进行显著性差异分析。计算每个指标的相对变化率并进行单因素方差分析, 以进一步评估舍曲林对蚤状溞响应信息素的影响。
对F2代的体长、尾刺长及种群相关指标在每个处理中进行双因素方差分析, 并计算每个指标的相对变化率。对每个指标的相对变化率进行双因素方差分析, 评估F2代中舍曲林和信息素对所有指标相对变化率的交互作用, 然后进行Tukey检验进行显著性差异分析。
2 结果 2.1 舍曲林对F1代诱导型防御以及种群指标的影响 2.1.1 性成熟阶段舍曲林和信息素处理对蚤状溞的性成熟体长均有显著影响, 且二者存在交互作用(图 2;表 1)。在任一个舍曲林浓度下, 相比于无信息素组, 添加信息素均显著减小蚤状溞的性成熟体长。在性成熟体长相对变化率方面, 舍曲林浓度上升显著加强性成熟体长减小的程度(P < 0.05)。

|
| 图 2 不同舍曲林浓度和信息素处理下蚤状溞亲代(F1)性成熟时相关特征及其相对变化率 Fig. 2 Key traits and relative change rate at maturity of Daphnia pulex F1 generation under different sertraline concentrations with or without fish kairomone 不同大写或小写字母表示同一颜色的数据柱之间存在差异显著(P < 0.05), *表示同一舍曲林浓度下信息素处理间存在差异显著(***:P < 0.001) |
| 指标Traits | df | F | P | 指标Traits | df | F | P | |
| 性成熟体长BL at maturity | 第9天相对尾刺长SB on 9th day | |||||||
| 信息素处理FK | 1 | 1393.200 | <0.001 | 信息素处理FK | 1 | 329.202 | <0.001 | |
| 舍曲林浓度SC | 2 | 3.401 | 0.036 | 舍曲林浓度SC | 2 | 9.580 | <0.001 | |
| FK×SC | 2 | 10.723 | <0.001 | FK×SC | 2 | 2.843 | 0.062 | |
| 性成熟尾刺长SL at maturity> | 净繁殖率Net reproduction rate | |||||||
| 信息素处理FK | 1 | 17.355 | <0.001 | 信息素处理FK | 1 | 39.474 | <0.001 | |
| 舍曲林浓度SC | 2 | 0.839 | 0.434 | 舍曲林浓度SC | 2 | 25.180 | <0.001 | |
| FK×SC | 2 | 2.045 | 0.133 | FK×SC | 2 | 19.397 | <0.001 | |
| 性成熟相对尾刺长SB at maturity | 世代时间Generation time | |||||||
| 信息素处理FK | 1 | 311.933 | <0.001 | 信息素处理FK | 1 | 132.618 | <0.001 | |
| 舍曲林浓度SC | 2 | 0.600 | 0.550 | 舍曲林浓度SC | 2 | 89.835 | <0.001 | |
| FK×SC | 2 | 0.103 | 0.902 | FK×SC | 2 | 76.079 | <0.001 | |
| 第9天体长BL on 9th day | 种群内禀增长率IRP | |||||||
| 信息素处理FK | 1 | 338.006 | <0.001 | 信息素处理FK | 1 | 0.339 | 0.566 | |
| 舍曲林浓度SC | 2 | 33.264 | <0.001 | 舍曲林浓度SC | 2 | 3.521 | 0.047 | |
| FK×SC | 2 | 4.283 | 0.016 | FK×SC | 2 | 2.050 | 0.153 | |
| 第9天尾刺长SL on 9th day | ||||||||
| 信息素处理FK | 1 | 92.909 | <0.001 | |||||
| 舍曲林浓度SC | 2 | 0.273 | 0.762 | |||||
| FK×SC | 2 | 0.187 | 0.829 | |||||
| BL:体长Body length; SL:尾刺长Spine length; SB:相对尾刺长Spine length/body length; IRP:种群内禀增长率Intrinsic rate of population; FK:信息素处理Fish kairomone;SC:舍曲林浓度Sertraline concentration | ||||||||
舍曲林和信息素处理对蚤状溞的性成熟尾刺长和相对尾刺长(尾刺长与体长之比)不存在交互作用(图 2;表 1)。舍曲林浓度分别在0和0.2 μg/L, 相比于无信息素组, 添加信息素显著增加蚤状溞的性成熟尾刺长(P < 0.05);但是在20 μg/L时, 性成熟尾刺长无显著性差异(P>0.05)。相对变化率结果显示, 当舍曲林浓度为20 μg/L时, 在该指标上对鱼类信息素的响应强度减小(图 2)。在任一个舍曲林浓度下, 相比于无信息素组, 添加信息素均显著增加蚤状溞的性成熟相对尾刺长。而且相对变化率结果显示, 舍曲林没有对该指标的响应强度产生影响(图 2)。
2.1.2 个体发育至第9天(产完第三窝)舍曲林和信息素处理对蚤状溞第9天的体长均有显著影响, 且二者存在交互作用(图 3;表 1)。在各舍曲林浓度下, 相比于无信息素组, 添加信息素均显著减小蚤状溞的性成熟体长。分别在有或无信息素组中, 20 μg/L的舍曲林可以显著减小第9天的体长。相对变化率的结果显示, 舍曲林浓度上升显著加强第9天的体长减小的程度(P < 0.05)。

|
| 图 3 不同舍曲林浓度和信息素处理下蚤状溞F1代9日龄的相关特征及其相对变化率 Fig. 3 Key traits and relative change rate of Daphnia pulex F1 generation at 9th day under different sertraline concentrations with or without fish kairomone 不同大写或小写字母表示同一颜色的数据柱之间存在差异显著(P < 0.05), *表示同一舍曲林浓度下信息素处理间存在差异显著(***:P < 0.001) |
舍曲林和信息素处理对蚤状溞第9天的尾刺长和相对尾刺长不存在交互作用(图 3;表 1)。在三个舍曲林浓度下, 相比于无信息素组, 添加信息素均显著增加尾刺长和相对尾刺长(P < 0.05);在信息素组中, 20 μg/L的舍曲林显著增加第9天的相对尾刺长(P < 0.05)。相对变化率结果显示:舍曲林对第9天的绝对尾刺长响应强度不会产生影响(P>0.05), 但可以显著增强蚤状溞的相对尾刺长的响应强度(P < 0.05)。
2.1.3 种群适合度相关指标舍曲林浓度和信息素处理对蚤状溞的净繁殖率和世代时间存在交互作用, 对内禀增长率不存在交互作用(图 4;表 1)。舍曲林浓度在0和0.2 μg/L时, 相比于无信息素组, 添加信息素显著抑制种群净繁殖率, 缩短种群世代时间(P < 0.05);但是在20 μg/L舍曲林时, 发现信息素对种群净繁殖率无显著性影响(P>0.05), 并且信息素显著推迟了种群世代时间(P < 0.05)。相对变化率结果显示:0.2 μg/L的舍曲林分别显著加强种群净繁殖率减小以及世代时间缩短的程度(P < 0.05)。在种群内禀增长率方面, 舍曲林浓度分别在0 μg/L时, 相比于无信息素组, 添加信息素显著增加蚤状溞的种群内禀增长率(P < 0.05);但是在0.2和20 μg/L时, 种群内禀增长率无显著性差异(P>0.05)。

|
| 图 4 不同舍曲林浓度和信息素处理下蚤状溞F1代种群相关指标及其相对变化率 Fig. 4 Population traits and relative change rate of Daphnia pulex F1 generation under different sertraline concentrations with or without fish kairomone *表示同一舍曲林浓度下信息素处理间存在差异显著(*:P < 0.05;***:P < 0.001) |
信息素处理和连续两代舍曲林暴露对F2代的蚤状溞性成熟体长均有显著影响, 且二者存在交互作用(图 5;表 2)。在三个舍曲林浓度下, 相比于无信息素组, 添加信息素均显著减小蚤状溞的性成熟体长。同时, 相对变化率的结果显示:随着连续两代舍曲林浓度上升, 性成熟体长减小的程度显著性加强(图 5;P < 0.05)。

|
| 图 5 不同舍曲林浓度和信息素(FK)处理下蚤状溞子代(F2)性成熟体长、性成熟尾刺长、相对尾刺长及其相对变化率 Fig. 5 Body length, spine length and relative spine length at maturity of Daphnia pulex F2 generation and relative change rate under different sertraline concentrations with or without fish kairomone |
| 指标 Trait |
代内可塑性 Within-generational plasticity |
代际可塑性 Trans-generational plasticity |
|||||
| df | F | P | df | F | P | ||
| 性成熟体长BL at maturity | |||||||
| 信息素处理FK | 1 | 239.320 | <0.001 | 1 | 757.798 | <0.001 | |
| 舍曲林浓度SC | 2 | 267.935 | <0.001 | 2 | 35.790 | <0.001 | |
| FK×SC | 2 | 268.481 | <0.001 | 2 | 41.707 | <0.001 | |
| 性成熟尾刺长SL at maturity | |||||||
| 信息素处理FK | 1 | 30.579 | <0.001 | 1 | 3.612 | 0.059 | |
| 舍曲林浓度SC | 2 | 48.452 | <0.001 | 2 | 0.784 | 0.458 | |
| FK×SC | 2 | 41.707 | 0.940 | 2 | 0.564 | 0.570 | |
| 性成熟相对尾刺长SB at maturity | |||||||
| 信息素处理FK | 1 | 501.621 | <0.001 | 1 | 127.379 | <0.001 | |
| 舍曲林浓度SC | 2 | 36.953 | <0.001 | 2 | 18.514 | <0.001 | |
| FK×SC | 2 | 8.382 | <0.001 | 2 | 8.341 | <0.001 | |
| 第9天体长BL on 9th day | |||||||
| 信息素处理FK | 1 | 331.488 | 0.070 | <0.001 | 664.872 | <0.001 | |
| 舍曲林浓度SC | 2 | 32.365 | <0.001 | <0.001 | 45.935 | <0.001 | |
| FK×SC | 2 | 8.433 | 0.577 | <0.001 | 10.978 | <0.001 | |
| 第9天尾刺长SL on 9th day | |||||||
| 信息素处理FK | 1 | 9.394 | 0.003 | 1 | 55.354 | <0.001 | |
| 舍曲林浓度SC | 2 | 35.755 | <0.001 | 2 | 0.0656 | 0.937 | |
| FK×SC | 2 | 1.469 | 0.236 | 2 | 0.664 | 0.517 | |
| 第9天相对尾刺长SB on 9th day | |||||||
| 信息素处理FK | 1 | 137.613 | <0.001 | 1 | 450.901 | <0.001 | |
| 舍曲林浓度SC | 2 | 34.868 | <0.001 | 2 | 14.355 | <0.001 | |
| FK×SC | 2 | 14.541 | <0.001 | 2 | 2.988 | 0.05 | |
| 净繁殖率Net reproduction rate | |||||||
| 信息素处理FK | 1 | 160.692 | <0.001 | 1 | 59.174 | <0.001 | |
| 舍曲林浓度SC | 2 | 38.151 | <0.001 | 2 | 0.000196 | 1.000 | |
| FK×SC | 2 | 2.139 | 0.14 | 2 | 4.242 | 0.026 | |
| 世代时间Generation time | |||||||
| 信息素处理FK | 1 | 80.495 | <0.001 | 1 | 60.764 | <0.001 | |
| 舍曲林浓度SC | 2 | 6.640 | 0.005 | 2 | 2.139 | 0.140 | |
| FK×SC | 2 | 0.914 | 0.414 | 2 | 1.196 | 0.320 | |
| 种群内禀增长率IRP | |||||||
| 信息素处理FK | 1 | 118.147 | <0.001 | 1 | 46.245 | <0.001 | |
| 舍曲林浓度SC | 2 | 52.171 | <0.001 | 2 | 2.039 | 0.152 | |
| FK×SC | 2 | 0.347 | 0.711 | 2 | 7.906 | 0.002 | |
信息素处理和连续两代舍曲林胁迫对F2代的性成熟尾刺长不存在交互作用(图 5;表 2)。在三个舍曲林浓度下, 相比于无信息素组, 添加信息素均显著增加尾刺长(P < 0.05);而且相对变化率结果显示, 连续两代舍曲林胁迫对该指标的相对变化程度没有影响(图 5;P>0.05)。在相对尾刺长度方面(图 5), 在三个舍曲林浓度下, 相比于无信息素组, 添加信息素均显著增加性成熟相对尾刺长度(P < 0.05), 同时相对尾刺长度的变化率也随着连续两代舍曲林胁迫浓度增加而上升(图 5;P < 0.05)。
2.2.2 个体发育至第9天(产完第三窝)信息素处理和连续两代舍曲林暴露对F2代9日龄体长均有显著影响, 且二者存在交互作用(图 6;表 2)。在三个舍曲林浓度下, 相比于无信息素组, 添加信息素均显著减小9日龄体长。同时, 相对变化率的结果显示:连续两代暴露于20 μg/L舍曲林导致性成熟体长减小的程度显著性加强(图 6;P < 0.05)。

|
| 图 6 不同舍曲林浓度和信息素(FK)处理下蚤状溞F2代9日龄体长、性成熟尾刺长、相对尾刺长及其相对变化率 Fig. 6 Body length, spine length and relative spine length at the 9th day of Daphnia pulex F2 generation and relative change rate under different sertraline concentrations with or without fish kairomone |
在尾刺长方面(图 6), 舍曲林浓度在20 μg/L时, 相比于无信息素组, 添加信息素显著增加9日龄的尾刺长(P < 0.05);但是在0和0.2 μg/L时, 尾刺长无显著性差异(P>0.05)。在相对尾刺长度方面(图 6), 分别在三个连续两代舍曲林浓度下, 相比于无信息素组, 添加信息素均显著增加9日龄相对尾刺长度(P < 0.05), 同时相对尾刺长度的变化率也随着连续两代舍曲林胁迫浓度增加而上升(图 6;P < 0.05)。
2.2.3 种群适合度相关指标连续两代暴露于舍曲林和信息素处理对F2代的蚤状溞的净繁殖率和世代时间无交互作用, 但是对内禀增长率存在交互作用(图 7;表 2)。在三个舍曲林浓度下, 相比于无信息素组, 添加信息素显著缩短种群世代时间, 抑制种群净繁殖率, 以及种群内禀增长率(P < 0.05)。然而, 相对变化率结果:连续两代暴露于舍曲林对种群净繁殖率、种群内禀增长率减小以及世代时间缩短的程度无显著性影响(P>0.05)。

|
| 图 7 不同舍曲林浓度和信息素(FK)处理下蚤状溞F2代种群净繁殖率、世代时间、种群内禀增长率及其相对变化率 Fig. 7 Net reproduction rate, generation time and intrinsic rate of population growth of Daphnia pulex F2 generation and relative change rate under different sertraline concentrations with or without fish kairomone |
连续两代的信息素处理和舍曲林暴露对F2代的蚤状溞性成熟体长均有显著影响, 且二者存在交互作用(图 5;表 2)。在三个舍曲林浓度下, 相比于无信息素组, 添加信息素均显著减小蚤状溞的性成熟体长。同时, 相对变化率的结果显示:相比于从未经历舍曲林胁迫, 连续两代暴露于20 μg/L舍曲林浓度导致性成熟阶段的反捕食防御体长减小的程度显著加强(图 5;P < 0.05)。
连续两代的信息素处理和舍曲林胁迫对F2代的性成熟尾刺长不存在交互作用(图 5;表 2)。分别在三个舍曲林浓度下, 相比于无信息素组, 连续两代的添加信息素处理均没有显著增加尾刺长(P>0.05);在相对尾刺长度方面(图 5), 在三个舍曲林浓度下, 相比于无信息素组, 在连续两代中添加信息素均显著增加F2性成熟相对尾刺长度(P < 0.05), 同时相对尾刺长度的变化率也随着连续两代舍曲林胁迫浓度增加而上升(图 5;P < 0.05)。同时, 相对变化率的结果显示:相比于从未经历舍曲林胁迫, 连续两代暴露于20 μg/L舍曲林浓度导致性成熟阶段的反捕食防御相对尾刺长减小的程度显著加强(图 5;P < 0.05)。
2.3.2 个体发育至第9天(产完第三窝)连续两代的信息素处理和舍曲林暴露对F2代的蚤状溞9日龄体长均有显著影响, 且二者存在交互作用(图 6;表 2)。在三个舍曲林浓度下, 相比于无信息素组, 在连续两代中添加信息素显著减小9日龄个体体长。同时, 相对变化率的结果显示:相比于从未经历舍曲林胁迫, 连续两代暴露于20 μg/L舍曲林浓度导致9日龄阶段的反捕食防御体长减小的程度显著加强(图 6;P < 0.05)。
连续两代的信息素处理和舍曲林胁迫对F2代的性成熟尾刺长不存在交互作用(图 6;表 2)。在三个舍曲林浓度下, 相比于无信息素组, 连续两代的添加信息素处理均显著增加尾刺长(P < 0.05);在相对尾刺长度方面(图 6), 在三个舍曲林浓度下, 相比于无鳑信息素组, 在连续两代中添加信息素均显著增加F2性成熟相对尾刺长度(P < 0.05)。同时, 相对变化率的结果显示:相比于从未经历舍曲林胁迫, 随着连续两代舍曲林胁迫浓度增加, 9日龄阶段的反捕食防御相对尾刺长增加的程度也显著加强(图 6;P < 0.05)。
2.3.3 种群适合度相关指标连续两代暴露于舍曲林和信息素处理对F2代的蚤状溞的种群净繁殖率和内禀增长率存在交互作用, 但是对世代时间无交互作用(图 7;表 2)。在三个舍曲林浓度下, 相比于无信息素组, 添加信息素显著缩短种群世代时间, 抑制种群净繁殖率, 以及种群内禀增长率(P < 0.05)。然而, 相对变化率结果:相比于从未经历舍曲林胁迫, 连续两代暴露于20 μg/L舍曲林浓度加强了反捕食防御干扰种种群净繁殖率和内禀增长率的作用(P < 0.05)。
3 讨论诱导型反捕食防御对维持种间互作的结构与功能稳定具有重要的作用。诱导型反捕食防御的表达与被捕食者发育阶段、生理内稳态有关, 因此包括舍曲林在内的化学药物能够干扰浮游动物的反捕食防御的触发和维持[35, 37, 39—41]。本研究分别从代内可塑性与代际可塑性层面探讨连续两代舍曲林暴露对蚤状溞响应鱼类信息素的诱导型防御的影响。研究结果表明, 连续两代舍曲林暴露导致诱导型防御代内可塑性发生变化:舍曲林暴露放大了鱼类信息素诱导的蚤状溞体长减小趋势以及相对尾刺长度增大趋势, 并且对鱼类信息素作用的种群适合度参数(种群净繁殖率、种群世代时间、种群内禀增长率)无显著性影响效果。在代际可塑性方面, 随着连续两代舍曲林的浓度上升, 亲代经历鱼类信息素产下的子代继续响应鱼类信息素时, 体长减小、相对尾刺长度增大、种群净繁殖量以及种群内禀增长率下降的趋势得到进一步加强。结合舍曲林使用量大并且在水中难以被降解的特征, 长期舍曲林污染可能会干扰以蚤状溞为代表的浮游动物响应鱼类的捕食风险行为与种群维持策略[29]。由于适应性表型可塑性(如繁殖输出、个体发育大小)上的细小变化都会对小型猎物(尤其是r-对策者)种群增长率产生明显的影响[42—43], 因此环境中的捕食风险以及污染胁迫因素通常会明显增加猎物适应环境的难度。另外, 枝角类对捕食者的响应(包括形态、行为和生理)通常存在克隆依赖性分化[18, 23]。诱导型防御的克隆变异可以促使种群产生局部适应, 从而维持捕食者和猎物之间共同进化的军备竞赛关系。因此, 基于本研究的架构, 建议可以进一步开展包含更多克隆、更多物种的研究, 分析比较舍曲林干扰诱导型防御的克隆保守性程度。
3.1 诱导型防御代内可塑性的响应代内可塑性赋予单一基因型生物在发育过程中改变表型的能力, 来应对短期内环境变化的影响[44]。枝角类的反捕食防御在表型上的调整涉及形态、行为以及生活史方面, 这些防御表型的改变能够有效降低猎物被捕食的概率。在本研究中, F1代受鱼类信息素的影响下, 个体体长缩短、相对尾刺增加, 世代周期缩短(繁殖时间提前)以及种群适合度上升(图 2—4), 这些结果与先前的报道相似[18, 20, 45], 说明鱼类信息素有效诱导蚤状溞产生形态与种群(生活史)方面的防御特征。在此基础上, 舍曲林的存在改变F1代的诱导型防御特征:一方面, 舍曲林放大了信息素诱导的体长缩小以及相对尾刺变长的趋势;另一方面舍曲林干扰个体繁殖输出、世代时间, 抑制种群内禀增长率(图 4), 由此降低蚤状溞种群(生活史)防御作用。该结果表明, 当存在鱼类捕食风险以及舍曲林联合胁迫时, 蚤状溞的诱导型防御以削弱种群(生活史)防御为代价, 加强个体形态防御效果。先前的研究发现, 低浓度的舍曲林能够强化网纹溞的形态防御特征[37], 推测是由于低浓度的神经干扰物影响化学信息的传导并导致了信号系统和接收器的适应性响应[35]。Bell等[20]观察食物中磷元素含量对蚤状溞诱导型防御的影响, 结果发现磷含量比例下降导致鱼类信息素诱导的性成熟体长缩小趋势增强, 同时抑制了种群内禀增长率。不同的是, Mirza等[46]报道了铜离子污染导致蚤状溞形态防御受损, 但对生活史防御无明显影响。另外, 研究发现, 体长缩小的趋势分别在个体性成熟以及第九日龄阶段皆与舍曲林的浓度呈正相关。不同的是, 在性成熟阶段不存在信息素诱导尾刺长和相对尾刺长增长的趋势被舍曲林放大的现象(图 2), 而在个体达到九日龄时, 舍曲林放大了信息素诱导尾刺长和相对尾刺长增长的趋势(图 3), 这说明由舍曲林触发的形态防御效果的增强与个体发育进程有关, 继而表现出舍曲林对诱导型防御代内可塑性的调控作用。
当F1代在舍曲林干扰下产下的幼体(即F2代)面临鱼类信息素与舍曲林联合作用时, 鱼类信息素诱导的体长缩小、相对尾刺变长的趋势被连续两代舍曲林暴露所放大(图 5和6), 同时对种群(生活史)防御无明显干扰影响(图 7), 这个结果与F1代诱导型防御受到舍曲林作用存在一定的差别:舍曲林的单一代内暴露方式以及连续两代暴露方式都导致蚤状溞形态防御效果的增强, 但是, 单一代内舍曲林暴露同时对种群(生活史)防御造成干扰;连续两代舍曲林暴露并没有表现对种群(生活史)防御干扰效果。根据诱导型防御代价理论:诱导型防御的形成以及维持是一种耗能过程, 诱导型防御形成的背后往往存在着某一些形式的代价[47—48]。然而, 有些情况代价的形式可能难以被检测。所以, 连续两代舍曲林暴露在强化形态防御的同时并没有削弱种群(生活史)防御, 并不能表示防御强化所产生的代价不存在。生理研究发现, 舍曲林能够抑制乙酰胆碱酯酶(AChE)的活性[49], 而AChE活性与枝角类的游泳能力正相关[50]。因此, 需要更多的后续研究验证连续两代舍曲林暴露在强化形态防御的同时是否会干扰蚤状溞的游动能力, 由此判断形态防御的强化是否以行为防御代价为基础。
3.2 诱导型防御代际可塑性的响应目前表型可塑性研究认为, 如果当前环境条件可以准确反映未来环境的变化时, 代际可塑性通常表现为对环境的适应倾向:亲代修饰子代的功能性状, 以帮助子代尽可能地适应环境。枝角类连续世代暴露鱼类信息素时, 后代依然能表现诱导型防御特征[16], 甚至形态防御随着亲代暴露代数增加而得到加强[22], 体现出诱导型防御的代间可塑性对捕食风险的适应特征。在这些研究结果的基础上, 进一步发现连续两代的舍曲林暴露导致子代的形态防御加强, 表现为随着舍曲林浓度上升, 子代的性成熟以及第九日龄的体长缩小以及相对尾刺长度增加, 但是绝对尾刺长度不发生变化;同时, 结果显示子代的种群净繁殖率受抑制程度被放大(图 7), 由此形成了形态防御增强的代价体现在抑制种群适合度方面(图 7)。基于转录组技术, Hales等[51]证实枝角类诱导型防御的代际可塑性可能与外骨骼组分上调表达相关。由于枝角类的形态防御的体现需要外骨骼不断地蜕皮与重生来维持, 因此, 枝角类的形态防御效果直接受到外骨骼组分功能基因表达的影响。有研究发现舍曲林处理组导致甲壳动物小龙虾发生22次蜕皮(对照组为15次)[52], 提示舍曲林可能促进外骨骼的重生过程。因此, 推测这种形态防御代际可塑性的加强可能与舍曲林进一步上调外骨骼组分功能基因表达有关。相似的研究结果也见于农药对轮虫反捕食防御的代际可塑性影响。Heine-Fuster等[53]发现农药甲胺磷不会引起亲代响应捕食者信息素时种群增长率下降, 却引起子代在增强形态防御的同时, 种群内禀增长率受到抑制。然而, 水体二氧化碳上升不仅抑制蚤状溞子代的形态防御, 同时对子代生活史防御的表达也产生干扰效应, 这可能是由于二氧化碳抑制了蚤状溞嗅觉感知能力, 使得蚤状溞对鱼类信息素的敏感性下降[54]。
3.3 舍曲林对相对尾刺长度的影响尾刺是枝角类重要的形态防御“武器”[45]。尾刺的作用是在捕食事件发生之前, 枝角类利用较长的尾刺“警告”捕食者, 而不是在被捕食之后阻止鱼类消化过程[55—56]。一些较大的浮游动物在响应鱼类信息素时, 可以减小它们的体型, 同时会拉长尾刺的长度, 由此增加相对尾刺长[23]。本研究发现, 连续两代舍曲林暴露增强蚤状溞的形态防御, 即强化体长缩小和相对尾刺长度增大的程度, 但是并没有强化绝对尾刺长度增长的变化趋势, 表明尾刺的长度存在天花板阈值。这与以下几个方面原因有关:首先, 枝角类防御形态的维持需要不断的物质和能量来源。相对于缩小个体体长, 延伸尾刺需要投入更多的资源, 比如甲壳素(构建基本骨架)、钙离子(提升硬度)。同时, 如前所述, 尾刺的作用仅仅是为了“警告”捕食者, 并不能作为真正的防御武器去阻挡捕食者进攻, 造成尾刺延伸的“受益-投入”比过低。其次, 研究表明处于孵育囊中的胚胎已经开始分化形态防御特征[57]。延伸的尾刺增加折叠难度, 尾刺也容易发生断裂, 还可能在拥挤的孵育囊中“戳”到近邻幼体。所以, 蚤状溞面对捕食者信息素以及舍曲林的联合暴露, 选择在保持尾刺一定长度的同时, 通过缩小个体体长的方式提高相对尾刺长度, 尽管缩小的个体可能会带来繁殖输出受限, 如本研究中种群净繁殖率下降。
3.4 舍曲林胁迫对胚胎的干扰效应在3.1中提到, 诱导型防御代内可塑性在连续两代舍曲林暴露与单代舍曲林暴露之间存在一定的差别:在加强形态防御效果的同时, 后者同时削弱了种群(生活史)防御效果, 而前者没有。推测产生这种差异的主要原因与亲代(即F1代)受到舍曲林的影响有关, 导致子代(即F2代)的诱导型防御表型产生了分化。但是, 值得注意的一点是, 产生这种差异可能也与子代(即F2代)在胚胎发育阶段直接受到舍曲林的干扰有关。具体地说, 在连续两代舍曲林暴露实验中, F2代在“正式”受到舍曲林作用之前, 其作为胚胎在F1的孵育囊中已经受到舍曲林的影响。同时, 收集的实验个体是24 h之内出生的幼体。有研究认为, 处于胚胎发育时期以及离开母体阶段的枝角类对环境变化十分敏感[8, 58]。因此, F2代在胚胎期以及出生后的24 h之内受到舍曲林的影响, 也是导致连续两代舍曲林暴露与单代舍曲林暴露之间形成诱导型防御代内可塑性差别的原因之一。不过, 更倾向于支持“母体受影响”因素, 因为亲代(F1)受到舍曲林暴露持续了9天(第三窝), 而F2代在胚胎期以及出生后的24 h内, 其受舍曲林影响只持续了其中小部分时期。
4 结论生态学家们逐渐意识到发生在表型可塑性、遗传变异以及环境污染三者间的交联互作的重要性[59—60]。然而, 代内以及代际表型可塑性调控生态-演化响应进程的潜在作用仍缺乏足够的认识。本文分析了神经活性药品代表污染物舍曲林对常见枝角类的诱导型防御特征分别在代内以及代间维度的干扰作用。结果发现:(1)在代内可塑性方面, 连续两代的舍曲林暴露放大了鱼类信息素诱导的蚤状溞体长减小趋势以及相对尾刺长度增大趋势, 并且对鱼类信息素作用的种群适合度参数无显著性影响效果;(2)在代际可塑性方面, 随着连续两代舍曲林的浓度上升, 亲代经历鱼类信息素后产下的子代继续响应鱼类信息素时, 体长减小、相对尾刺长度增大, 说明形态防御得到加强;同时种群净繁殖量以及种群内禀增长率下降的趋势被放大, 由此可能抑制鱼类捕食风险下蚤状溞的种群丰度。上述结果表明, 尽管神经药品污染物对浮游动物没有直接的致死效应, 但是神经活性污染物长期停留于水环境, 会干扰浮游动物的反捕食防御行为, 从而间接影响浮游动物与其捕食者种群互作。因此, 本论文的研究结果强调了代内以及代际表型可塑性在“生态-演化-人类活动”之间互作进程中的作用。
| [1] |
Agrawal A A. Phenotypic plasticity in the interactions and evolution of species. Science, 2001, 294(5541): 321-326. DOI:10.1126/science.1060701 |
| [2] |
王小艺, 杨忠岐, 魏可, 唐艳龙. 昆虫翅型分化的表型可塑性机制. 生态学报, 2015, 35(12): 3988-3999. |
| [3] |
王姝, 周道玮. 植物表型可塑性研究进展. 生态学报, 2017, 37(24): 8161-8169. |
| [4] |
Uller T. Developmental plasticity and the evolution of parental effects. Trends in Ecology & Evolution, 2008, 23(8): 432-438. |
| [5] |
Van Donk E, Ianora A, Vos M. Induced defences in marine and freshwater phytoplankton: a review. Hydrobiologia, 2011, 668(1): 3-19. DOI:10.1007/s10750-010-0395-4 |
| [6] |
Dao T S, Do-Hong L C, Wiegand C. Chronic effects of cyanobacterial toxins on Daphnia magna and their offspring. Toxicon, 2010, 55(7): 1244-1254. DOI:10.1016/j.toxicon.2010.01.014 |
| [7] |
Reis P, Pereira R, Carvalho F P, Oliveira J, Malta M, Mendo S, Lourenço J. Life history traits and genotoxic effects on Daphnia magna exposed to waterborne uranium and to a uranium mine effluent-a transgenerational study. Aquatic Toxicology, 2018, 202: 16-25. DOI:10.1016/j.aquatox.2018.06.009 |
| [8] |
Walsh M R, Gillis M K. Transgenerational plasticity in the eye size of Daphnia. Biology Letters, 2021, 17(6): 20210143. DOI:10.1098/rsbl.2021.0143 |
| [9] |
Jiang X D, Yang W, Zhao S Y, Liang H S, Zhao Y L, Chen L Q, Li R. Maternal effects of inducible tolerance against the toxic cyanobacterium Microcystis aeruginosa in the grazer Daphnia carinata. Environmental Pollution, 2013, 178: 142-146. DOI:10.1016/j.envpol.2013.03.017 |
| [10] |
Galloway L F, Etterson J R. Transgenerational plasticity is adaptive in the wild. Science, 2007, 318(5853): 1134-1136. DOI:10.1126/science.1148766 |
| [11] |
Donelson J M, Salinas S, Munday P L, Shama L N S. Transgenerational plasticity and climate change experiments: where do we go from here?. Global Change Biology, 2018, 24(1): 13-34. DOI:10.1111/gcb.13903 |
| [12] |
马剑敏, 靳萍, 郭萌, 代克岩, 徐婷婷, 杨程, 蔺庆伟. 磷浓度对铜绿微囊藻、大型溞和金鱼藻三者相互作用的影响. 生态学报, 2014, 34(6): 1520-1526. |
| [13] |
魏利军, 汤龙升, 李胜男, 周建, 孔繁翔, 史小丽. CO2升高对枝角类群落结构影响的原位模拟. 生态学报, 2016, 36(7): 1846-1853. |
| [14] |
Reede T. Life history shifts in response to different levels of fish kairomones in Daphnia. Journal of Plankton Research, 1995, 17(8): 1661-1667. DOI:10.1093/plankt/17.8.1661 |
| [15] |
Pijanowska J, Stolpe G. Summer diapause in Daphnia as a reaction to the presence of fish. Journal of Plankton Research, 1996, 18(8): 1407-1412. DOI:10.1093/plankt/18.8.1407 |
| [16] |
Walsh M R, Cooley F, Biles K, Munch S B. Predator-induced phenotypic plasticity within-and across-generations: a challenge for theory?. Proceedings of the Royal Society B: Biological Sciences, 2015, 282(1798): 20142205. DOI:10.1098/rspb.2014.2205 |
| [17] |
Qin S S, Ma L L, Li D, Huang J, Zhang L, Sun Y F, Yang Z. Rising temperature accelerates the responses of inducible anti-predator morphological defenses of Ceriodaphnia cornuta but decreases the responsive intensity. Ecological Indicators, 2021, 120: 106919. DOI:10.1016/j.ecolind.2020.106919 |
| [18] |
Wilczynski W, Dynak P, Babkiewicz E, Bernatowicz P, Leniowski K, Maszczyk P. The combined effects of hypoxia and fish kairomones on several physiological and life history traits of Daphnia. Freshwater Biology, 2019, 64(12): 2204-2220. DOI:10.1111/fwb.13407 |
| [19] |
Gu L, Qin S S, Zhu S S, Lu N, Sun Y F, Zhang L, Huang Y, Lyu K, Chen Y F, Yang Z. Microcystis aeruginosa affects the inducible anti-predator responses of Ceriodaphnia cornuta. Environmental Pollution, 2020, 259: 113952. DOI:10.1016/j.envpol.2020.113952 |
| [20] |
Bell A C, Murray D L, Prater C, Frost P C. Fear and food: effects of predator-derived chemical cues and stoichiometric food quality on Daphnia. Limnology and Oceanography, 2019, 64(4): 1706-1715. DOI:10.1002/lno.11145 |
| [21] |
Coors A, De Meester L. Synergistic, Antagonistic and additive effects of multiple stressors: predation threat, parasitism and pesticide exposure in Daphnia magna. Journal of Applied Ecology, 2008, 45(6): 1820-1828. DOI:10.1111/j.1365-2664.2008.01566.x |
| [22] |
Pötter L, Krebs N, Horstmann M, Tollrian R, Weiss L C. Long-term effects of elevated pCO2 levels on the expression of Chaoborus-induced defences in Daphnia pulex. Zoology, 2021, 146: 125909. DOI:10.1016/j.zool.2021.125909 |
| [23] |
Diel P, Kiene M, Martin-Creuzburg D, Laforsch C. Knowing the enemy: inducible defences in freshwater zooplankton. Diversity, 2020, 12(4): 147. DOI:10.3390/d12040147 |
| [24] |
Calisto V, Esteves V I. Psychiatric pharmaceuticals in the environment. Chemosphere, 2009, 77(10): 1257-1274. DOI:10.1016/j.chemosphere.2009.09.021 |
| [25] |
Metcalfe C D, Chu S G, Judt C, Li H X, Oakes K D, Servos M R, Andrews D M. Antidepressants and their metabolites in municipal wastewater, and downstream exposure in an urban watershed. Environmental Toxicology and Chemistry, 2010, 29(1): 79-89. DOI:10.1002/etc.27 |
| [26] |
Schultz M M, Furlong E T. Trace analysis of antidepressant pharmaceuticals and their select degradates in aquatic matrixes by LC/ESI/MS/MS. Analytical Chemistry, 2008, 80(5): 1756-1762. DOI:10.1021/ac702154e |
| [27] |
Styrishave B, Halling-Sørensen B, Ingerslev F. Environmental risk assessment of three selective serotonin reuptake inhibitors in the aquatic environment: a case study including a cocktail scenario. Environmental Toxicology and Chemistry, 2011, 30(1): 254-261. DOI:10.1002/etc.372 |
| [28] |
Brooks B W, Chambliss C K, Stanley J K, Ramirez A, Banks K E, Johnson R D, Lewis R J. Determination of select antidepressants in fish from an effluent-dominated stream. Environmental Toxicology and Chemistry, 2005, 24(2): 464-469. DOI:10.1897/04-081R.1 |
| [29] |
Gornik T, Vozic A, Heath E, Trontelj J, Roskar R, Zigon D, Vione D, Kosjek T. Determination and photodegradation of sertraline residues in aqueous environment. Environmental Pollution, 2020, 256: 113431. DOI:10.1016/j.envpol.2019.113431 |
| [30] |
Richmond E K, Rosi-Marshall E J, Lee S S, Thompson R M, Grace M R. Antidepressants in stream ecosystems: influence of selective serotonin reuptake inhibitors (SSRIs) on algal production and insect emergence. Freshwater Science, 2016, 35(3): 845-855. DOI:10.1086/687841 |
| [31] |
Yang Z P, Lu T, Zhu Y C, Zhang Q, Zhou Z G, Pan X L, Qian H F. Aquatic ecotoxicity of an antidepressant, sertraline hydrochloride, on microbial communities. Science of the Total Environment, 2019, 654: 129-134. DOI:10.1016/j.scitotenv.2018.11.164 |
| [32] |
Chevalier J, Harscoët E, Keller M, Pandard P, Cachot J, Grote M. Exploration of Daphnia behavioral effect profiles induced by a broad range of toxicants with different modes of action. Environmental Toxicology and Chemistry, 2015, 34(8): 1760-1769. DOI:10.1002/etc.2979 |
| [33] |
Minguez L, Ballandonne C, Rakotomalala C, Dubreule C, Kientz-Bouchart V, Halm-Lemeille M P. Transgenerational effects of two antidepressants (sertraline and venlafaxine) on Daphnia magna life history traits. Environmental Science & Technology, 2015, 49(2): 1148-1155. |
| [34] |
Weiss L C, Kruppert S, Laforsch C, Tollrian R. Chaoborus and gasterosteus anti-predator responses in Daphnia pulex are mediated by independent cholinergic and gabaergic neuronal signals. Plos One, 2012, 7(5): e36879. DOI:10.1371/journal.pone.0036879 |
| [35] |
Weiss L C, Leese F, Laforsch C, Tollrian R. Dopamine is a key regulator in the signalling pathway underlying predator-induced defences in Daphnia. Proceedings of the Royal Society B: Biological Sciences, 2015, 282(1816): 20151440. DOI:10.1098/rspb.2015.1440 |
| [36] |
郭慧荣, 仇剑崟, 徐俊冕. 舍曲林与舒必利合用治疗早期精神分裂症3例. 上海精神医学, 2004, 16(2): 100-102. |
| [37] |
Zhu S S, Wu G J, Gu L, Sun Y F, Zhang L, Huang Y, Lyu K, Yang Z. Antidepressant sertraline impairs the induced morphological defense of Ceriodaphnia cornuta in response to Chaoborus larvae kairomone. Environmental Pollution, 2020, 266: 115092. DOI:10.1016/j.envpol.2020.115092 |
| [38] |
Lyu K, Zhang L, Gu L, Zhu X X, Wilson A E, Yang Z. Cladoceran offspring tolerance to toxic Microcystis is promoted by maternal warming. Environmental Pollution, 2017, 227: 451-459. DOI:10.1016/j.envpol.2017.04.095 |
| [39] |
Bedrossiantz J, Fuertes I, Raldua D, Barata C. Pharmacological modulation of fish-induced depth selection in D. magna: the role of cholinergic and GABAergic signalling. Scientific Reports, 2021, 11(1): 19407. DOI:10.1038/s41598-021-98886-w |
| [40] |
Van Donk E, Peacor S, Grosser K, De Senerpont Domis L N, Lürling M. Pharmaceuticals may disrupt natural chemical information flows and species interactions in aquatic systems: ideas and perspectives on a hidden global change//Gunther F A, de Voogt P, eds. Reviews of Environmental Contamination and Toxicology. Cham: Springer, 2016: 91-105.
|
| [41] |
Hedgespeth M L, Nilsson P A, Berglund O. Ecological implications of altered fish foraging after exposure to an antidepressant pharmaceutical. Aquatic Toxicology, 2014, 151: 84-87. DOI:10.1016/j.aquatox.2013.12.011 |
| [42] |
Hansen L K, Frost P C, Larson J H, Metcalfe C D. Poor elemental food quality reduces the toxicity of fluoxetine on Daphnia magna. Aquatic Toxicology, 2008, 86(1): 99-103. DOI:10.1016/j.aquatox.2007.10.005 |
| [43] |
Frost P C, Ebert D, Larson J H, Marcus M A, Wagner N D, Zalewski A. Transgenerational effects of poor elemental food quality on Daphnia magna. Oecologia, 2010, 162(4): 865-872. DOI:10.1007/s00442-009-1517-4 |
| [44] |
Moczek A P, Sultan S, Foster S, Ledón-Rettig C, Dworkin I, Nijhout H F, Abouheif E, Pfennig D W. The role of developmental plasticity in evolutionary innovation. Proceedings of the Royal Society B: Biological Sciences, 2011, 278(1719): 2705-2713. DOI:10.1098/rspb.2011.0971 |
| [45] |
Gu L, Xu Y, Yang T T, Qin S S, Zhang L, Sun Y F, Huang Y, Yang Z. Predator-induced allometric changes in the tail spine length of Daphnia: a distinct resource allocation strategy. Journal of Plankton Research, 2021, 43(6): 884-893. DOI:10.1093/plankt/fbab063 |
| [46] |
Mirza R S, Pyle G G. Waterborne metals impair inducible defences in Daphnia pulex: morphology, life-history traits and encounters with predators. Freshwater Biology, 2009, 54(5): 1016-1027. DOI:10.1111/j.1365-2427.2008.02148.x |
| [47] |
Black A R, Dodson S I. Demographic costs of Chaoborus-induced phenotypic plasticity in Daphnia pulex. Oecologia, 1990, 83(1): 117-122. DOI:10.1007/BF00324642 |
| [48] |
Riessen H P. Costs of predator-induced morphological defences in Daphnia. Freshwater Biology, 2012, 57(7): 1422-1433. DOI:10.1111/j.1365-2427.2012.02805.x |
| [49] |
Müller T C, Rocha J B T, Morsch V M, Neis R T, Schetinger M R C. Antidepressants inhibit human acetylcholinesterase and butyrylcholinesterase activity. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease, 2002, 1587(1): 92-98. |
| [50] |
Ren Z M, Zhang X, Wang X G, Qi P P, Zhang B, Zeng Y, Fu R S, Miao M S. AChE inhibition: one dominant factor for swimming behavior changes of Daphnia magna under DDVP exposure. Chemosphere, 2015, 120: 252-257. DOI:10.1016/j.chemosphere.2014.06.081 |
| [51] |
Hales N R, Schield D R, Andrew A L, Card D C, Walsh M R, Castoe T A. Contrasting gene expression programs correspond with predator-induced phenotypic plasticity within and across generations in Daphnia. Molecular Ecology, 2017, 26(19): 5003-5015. DOI:10.1111/mec.14213 |
| [52] |
Hossain S, Kubec J, Grabicová K, Grabic R, Randák T, Guo W, Kouba A, Buřič M. Environmentally relevant concentrations of methamphetamine and sertraline modify the behavior and life history traits of an aquatic invertebrate. Aquatic Toxicology, 2019, 213: 105222. DOI:10.1016/j.aquatox.2019.105222 |
| [53] |
Heine-Fuster I, Aránguiz-Acuña A, Ramos-Jiliberto R. Pesticide increases transgenerational cost of inducible defenses in a freshwater rotifer. Hydrobiologia, 2017, 799(1): 249-260. DOI:10.1007/s10750-017-3221-4 |
| [54] |
Weiss L C, Pötter L, Steiger A, Kruppert S, Frost U, Tollrian R. Rising pCO2 in freshwater ecosystems has the potential to negatively affect predator-induced defenses in Daphnia. Current Biology, 2018, 28(2): 327-332. DOI:10.1016/j.cub.2017.12.022 |
| [55] |
Engel K, Schreder T, Tollrian R. Morphological defences of invasive Daphnia lumholtzi protect against vertebrate and invertebrate predators. Journal of Plankton Research, 2014, 36(4): 1140-1145. DOI:10.1093/plankt/fbu023 |
| [56] |
Engel K, Tollrian R. Inducible defences as key adaptations for the successful invasion of Daphnia lumholtzi in North America?. Proceedings of the Royal Society B: Biological Sciences, 2009, 276(1663): 1865-1873. DOI:10.1098/rspb.2008.1861 |
| [57] |
Laforsch C, Tollrian R. Embryological aspects of inducible morphological defenses in Daphnia. Journal of Morphology, 2004, 262(3): 701-707. DOI:10.1002/jmor.10270 |
| [58] |
Garbutt J S, Scholefield J A, Vale P F, Little T J. Elevated maternal temperature enhances offspring disease resistance in Daphnia magna. Functional Ecology, 2014, 28(2): 424-431. |
| [59] |
Oostra V, Saastamoinen M, Zwaan B J, Wheat C W. Strong phenotypic plasticity limits potential for evolutionary responses to climate change. Nature Communications, 2018, 9(1): 1005. |
| [60] |
Hendry A P. Key questions on the role of phenotypic plasticity in eco-evolutionary dynamics. Journal of Heredity, 2016, 107(1): 25-41. |