 2023, Vol. 43
2023, Vol. 43文章信息
- 杨扬, 王嵘, 陈小勇
- YANG Yang, WANG Rong, CHEN Xiaoyong
- 榕-传粉小蜂化学通讯机制研究进展
- Chemical communication between figs and their pollinating wasps
- 生态学报. 2023, 43(11): 4400-4410
- Acta Ecologica Sinica. 2023, 43(11): 4400-4410
- http://dx.doi.org/10.5846/stxb202111073120
-
文章历史
- 收稿日期: 2021-11-07
- 网络出版日期: 2023-02-07

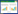




昆虫与植物是自然界中具有极高物种多样性的两大类群[1—2]。两者漫长的协同进化历史和相互间生态适应形成了丰富的种间关系[3], 不仅对维持生态系统的稳定性有重要作用, 而且与农业生产等方面休戚相关[4—6]。化学通讯是被子植物与昆虫间最为高效和普遍的互作方式之一[7—8]。植物释放的挥发性有机物(VOCs)为传粉昆虫与植食性昆虫识别宿主的提供了关键信息[9—10]。因此, 植物VOCs的变化被认为是驱动植物与昆虫协同进化与多样化的重要因素[11—12]。然而, 目前对化学通讯介导成种的理解尚不全面, 这主要是由于难以确定植物与昆虫在化学通讯中相互适应的关键性状[8, 13], 尤其在相对泛化的互作体系中[14]。此外, 某些昆虫对植物而言既是取食者(植食性幼虫)又是传粉者(成虫), 因此植物的VOCs常常同时受到来自取食者和传粉者的选择[15—18]。但这一复杂过程在一般的植物-传粉者共生体系中难以很好地分离。因此, 要厘清化学通讯的过程并探究背后的机制, 需要选取具有紧密协同进化历史和明显相互适应性状的植物和传粉昆虫作为研究体系[19—23]
桑科(Moraceae)榕属植物(Ficus, 下简称榕)是热带及亚热带重要的植物类群, 拥有极高的物种多样性(约800种)[24]。榕因其特殊的隐头花序(俗称榕果), 必须依赖于专一性的传粉榕小蜂(Agaonidae, Chalcidoidea, 下简称榕小蜂)进入隐头花序内部为其传粉[25];而雌性榕小蜂将卵产在雌花的子房内形成虫瘿, 其幼虫依靠取食虫瘿中的胚乳等植物组织方能完成生长发育[26]。紧密的种间互作使榕和榕小蜂构成了高度专一的专性共生体系(榕-蜂共生系统)[27], 而化学通讯则是维系二者专性共生关系的重要方式——雌性榕小蜂通过榕果在接受期(雌花成熟)释放的VOCs对宿主进行特异性识别[28—32]。尽管榕-蜂专性共生体系是探索植物-昆虫化学通讯模式与机理的理想研究对象, 但榕-蜂间的化学通讯机制及其对两者协同多样化的贡献仍不明确。近年来, 随着生理、多组学等技术的发展与普及, 这些问题得到了较深入的研究并取得重要进展。因此, 本文将对榕和榕小蜂蜂间化学通讯(下简称榕-蜂化学通讯)机制进行系统梳理, 总结化学通讯对于该共生关系的生态学与进化意义, 并对未来的研究进行展望。
1 榕-蜂化学通讯的基础榕在接受期释放的VOCs以及榕小蜂的嗅觉识别系统是榕-蜂化学通讯的重要基础, 阐明二者的基本特征是进一步分析榕-蜂特异性识别机制的重要前提。目前已通过大量研究揭示了榕气味谱与榕小蜂嗅觉系统的特征, 概括如下。
1.1 榕气味谱特征根据Knudsen的分类标准[33], 可大致将榕释放的VOCs分为脂肪族类、萜烯类、苯及苯系物、含氮化合物以及杂环化合物等。通过对比不同种榕以及同种榕不同发育时期、不同组织器官的气味谱, 可归纳出三个主要特征, 作为榕小蜂准确识别、定位提供化学信息基础。首先, 榕属植物接受期释放的VOCs存在种间异质性, 能够使榕小蜂准确识别专一性宿主。物种特异性化学物质的合成是这一特征产生的主要原因[29, 32, 34]。Grison-Pigé等[35]发现20种榕属植物的接受期气味谱中, 物种特异性VOCs的比例较高(37%), 但其相对含量通常较低(< 1%), 而泛化程度较高的物质相对含量往往较高。其次, 同种榕属植物VOCs的组成存在明显的物候变化[36—37], 这一特征能帮助传粉榕小蜂识别处于接受期的榕果以便顺利地完成授粉/产卵。如相比于接受前期(雌花未成熟), 对叶榕(Ficus hispida)接受期气味释放总量明显上升, 而间花期(雌花完成授粉)的释放量则急剧下降, 吸引专性传粉榕小蜂的活性物质芳樟醇的释放量在接受期显著提升是导致这一变化的主要原因[38]。类似地, 在鸡嗉子榕(Ficus semicordata)中发现对鸡嗉子榕小蜂(Ceratosolen gravelyi)起吸引作用的4-甲基苯甲醚仅在接受期大量释放, 在接受前期和间花期几乎不释放[28]。最后, 榕果不同组织释放的VOCs组成不同, 特别是苞片组织与其他组织/器官(如, 榕果外壁、叶片等)。这能帮助引导榕小蜂准确定位苞片口, 并由此进入榕果完成传粉/产卵。如Ficus burtt-davyi仅苞片口释放的VOCs对榕小蜂具有吸引作用[39]。此外, Wang等发现薜荔(Ficus pumila)接受期榕果的苞片组织释放的吸引性VOC浓度显著高于同时期雌花组织[21]。这一现象可能与苞片组织有许多释放VOCs的腺体以及吸引物合成的转录调控有关[40—41]。
以上介绍的榕果气味谱特征是形成榕-蜂特异性识别的重要代谢基础, 在研究中通过比较这些特征可有效缩小特异性吸引物的筛选范围, 为揭示两者相互适应的机制提供理想的切入点[21, 28, 30, 34]。
1.2 榕小蜂嗅觉系统特征榕小蜂对榕属植物特异性吸引物的准确鉴定是实现两者专性识别的另一重要基础。触角是榕小蜂接收化学信息的主要器官, 其上密布许多嗅觉感受器, 每一个嗅觉感受器内部都含有数个浸润在淋巴液内的嗅觉受体神经元[42]。当VOCs进入嗅觉感受器内后, 先溶解于淋巴液中, 随后与相应的气味结合蛋白(OBPs)结合形成配体, 这些配体与相应嗅觉受体结合引发神经冲动, 最终完成从气味分子的捕获到信号转化的过程[42—43]。
榕小蜂嗅觉感受器种类繁多, 根据形态差异可以分为毛状、锥状、板状、刺状、钟形、腔锥状和栓锥状等;根据功能可分为气味分子类别感受器、浓度感受器等[44—47]。不同种类榕小蜂的触角嗅觉感受器分布与组成存在明显差异(即物种特异性), 这与榕小蜂气味偏好性的产生有关, 且与其宿主榕释放的特异性吸引物质/物质组合的类别、浓度等相匹配[48—49]。如, 对5种鸡嗉子榕小蜂雌蜂的研究则表明雌蜂多孔板形感器的比表面积与宿主榕挥发物释放量之间存在显著正相关关系[50]。另外, 嗅觉感受器的组成在榕小蜂中存在明显的性二型性:雌性个体的触角更加发达, 感受器数量、种类以及结构多样化程度更高;而雄性个体触角退化, 感受器数量极少[51]。这与不同性别榕小蜂的生活史特征差异有关, 雌性须飞出榕果完成宿主识别并产卵/传粉, 而雄性大多终身生活在榕果内, 帮助雌性离开虫瘿并完成交配。
尽管上述榕果气味谱特征与榕小蜂嗅觉系统特征是形成榕-蜂特专性识别的重要基础, 但阐明榕小蜂特异性鉴别接受期榕果释放的VOCs的机制需依靠大量生物学和生态学实验的支持。近期结合基因组组学、昆虫行为学、电生理与分子生物学技术对榕属植物气味谱与榕小蜂气味感受过程进行深入研究, 已在两者识别机制领域取得了突破性进展。
2 榕-蜂特异性吸引-识别机制基于榕-蜂化学通讯的基础, 相关研究从吸引物鉴定出发深入分子机制层面, 深化了对于榕-蜂共生系统特异性化学识别机制的理解。
2.1 吸引物鉴定榕-蜂完成特异性吸引-识别过程仅通过化学通讯即可完成[52]。这种由单一类型信号介导的吸引-识别模式称之为私有通道[53—54]。目前仅在5种榕属植物中鉴定出了吸引榕小蜂的VOCs(表 1), 结果表明私有通道的建立可以依赖于某些特定的VOCs[54]。
| 榕属植物 Ficus species |
活性物质数量 Number of active VOCs |
活性物质种类 Categories of active VOCs |
吸引机制 Mechanism of attraction |
参考文献 References |
| 无花果 Ficus carica | 5种 | 苯甲醇、芳樟醇以及3种芳樟醇氧化物 | 关键物质苯甲醇与辅助物质芳樟醇产生协同效应 | [30] |
| 薜荔 Ficus pumila | 3种 | 癸醛、壬醛以及芳樟醇 | 关键物质癸醛单独吸引 | [21] |
| 对叶榕 Ficus hispida | 4种 | 苯甲酸苄酯以及3种未知倍半萜类物质 | 未知 | [23] |
| 大果榕 Ficus auriculate | 3种 | 3种未知倍半萜类物质 | 未知 | [23] |
| 鸡嗉子榕 Ficus semicordata | 1种 | 4-甲基苯甲醚 | 关键物质4-甲基苯甲醚单独吸引 | [23] |
| GC-EAD: 气谱-触角电位仪联动体系 Gas chromatography-electroantennogram detection; VOCs: 挥发性有机物 Volatile organic compounds | ||||
已有研究认为接受期榕果释放的VOCs中存在吸引榕小蜂的关键物质, 这些特定的VOC能够单独对榕小蜂起到特异性吸引作用, 如鸡嗉子榕释放的4-甲基苯甲醚[23, 28]。薜荔接受期榕果释放的VOCs中, 3种物质能引起薜荔榕小蜂的电生理反应, 后续的行为学实验证实仅癸醛起到吸引作用, 且并未观察到癸醛与其他VOCs存在协同效应[21]。
一些研究发现, 除特定的吸引物质外, 其他VOC成分与这些关键物质的组合可引发协同效应, 从而增强对榕小蜂的吸引力[55—56]。协同效应的效果与强度依赖于VOCs的具体组合以及浓度范围。如无花果的特异性吸引物为苯甲醇, 而该物质对无花果小蜂(Blastophaga psenes)的吸引作用能够被中性物质芳樟醇强化, 且两种物质间不同比例的组合对无花果传粉小蜂的吸引效果不同[30]。
最近, Yang等[57]采用信息论模型分析, 证实了只需1—2种化合物就可以维持榕-蜂共生关系的特异性。但在实际研究中, 准确判断榕-蜂化学通讯的关键物质较为困难, 这主要是由于榕果气味谱中低浓度VOCs常难以准确鉴定, 并且不易提取完成相关电生理实验。因此, 需要结合高精度气相色谱分析技术与高灵敏昆虫信息素测试系统形成气谱-触角电位仪联动体系(GC-EAD), 通过积累大量实证研究以系统、全面地验证这一气味识别机制在榕-蜂共生系统中的普适性。
2.2 分子机制确定榕的特异性吸引物为挖掘调控二者化学通讯的分子机制奠定基础。通常认为榕合成特异性吸引物质的关键基因与榕小蜂识别特异性气味的嗅觉基因均会经历较强的纯化选择, 因此筛选受选择基因能够缩小候选关键基因的范围。如在聚果榕亚属(Sycomorus)中, 萜烯类合成以及苯系化合物合成相关通路上共有18个基因受到纯化选择, 而参与萜烯类碳骨架合成的甲羟戊酸途径以及参与苯及苯丙素类合成的莽草酸途径中有9个基因受到选择清除作用[23];在对叶榕小蜂的77个嗅觉相关基因中则发现7个嗅觉受体蛋白基因(ORs)受到了强烈的选择清除[23]。这极大地缩小了候选关键基因的范围, 并表明少量的关键基因控制着榕-蜂特异性化学通讯。
这一推测在薜荔榕小蜂(Wiebesia pumilae)的研究中得到了验证。在薜荔榕小蜂、对叶榕小蜂及小叶榕小蜂(Eupristina verticillate)基因组中, 与嗅觉相关的两个重要基因家族-气味结合蛋白(OBPs)基因家族和气味受体蛋白(ORs)基因家族发生了显著的收缩。后续的物质亲和实验(SPR)发现, 薜荔榕小蜂仅通过单一气味结合蛋白与特异性的吸引物癸醛结合从而完成识别[21]。而在薜荔中, 与特异性吸引物癸醛合成直接相关的关键基因仅有4个, 这些关键基因在接受期的差异表达受到4个关键转录因子调控, 证实了少数关键基因控制特异性吸引物的合成[21]。而在粗叶榕(Ficus hirta)及其近缘种Ficus triloba中也证实了转录调控模式差异导致了二者的气味谱分化[41]。
因此, 特定转录调控模式下关键基因的特异性表达可能是榕果合成特异性吸引物质的重要分子机制(图 1);而嗅觉相关基因家族收缩使榕小蜂嗅觉识别能力特化, 同时个别OR及OBP基因功能特化导致榕小蜂能够识别宿主榕释放特异性吸引物(图 1), 从而实现榕-蜂的特异性化学通讯[21, 23, 58]。这些发现表明榕-蜂特化的化学通讯是二者在分子层面上相互适应的结果。

|
| 图 1 榕特异性吸引与榕小蜂特异性识别机制图解 Fig. 1 Illustration of the mechanism of specific attraction of figs and identification of fig wasps |
榕-蜂特异性的化学通讯模式对维持种间关系的专一性与稳定性有着十分重要的意义。特化的化学通讯在很大程度上避免了榕-蜂错配, 从而保障了双方的适合度。如鸡嗉子榕通过4-甲基苯甲醚吸引传粉者, 薜荔通过癸醛吸引传粉者[21, 23, 28]。而榕小蜂仅被宿主榕释放的气味所吸引[21, 23, 43]。正因如此, 特化的化学通讯还能在一定程度上缓解不同榕属植物争夺传粉者的竞争压力, 利于物种共存[25—27]。在不同榕VOCs存在较大差异的同时, 同种榕的VOCs则保持高度的相似性, 使种群中每个个体均有授粉机会, 这也是稳定两者种群规模的关键因素[59—60]。
更为重要的是, 榕-蜂特异性的化学通讯形成了一种推拉机制, 提高了共生双方有性繁殖成功率[61—62]。“拉”效应是指榕小蜂因对宿主榕接受期榕果释放的气味有着强烈的偏好性而被其特异性地吸引[28—30], 从而携带花粉进入接受期榕果完成授粉。“推”效应表现为处于非接受期榕果会释放排斥物使雌性榕小蜂携带花粉飞离。接受前期榕果雌花尚未完全成熟或苞片口紧闭[62—63], 而间花期雌花已被授粉/产卵, 故“推拉”机制能够很好地避免无效传粉的发生, 同时降低由于花序不成熟或过度竞争导致榕小蜂幼虫死亡的概率。目前, 已有研究发现了造成“推拉”机制的具体物质。如, 薜荔接受期榕果释放癸醛吸引榕小蜂, 而接受前期榕果则释放大量排斥榕小蜂的壬醛, 起到“推拉”作用[21]。
此外, 对于功能性雌雄异株榕, 榕-蜂种群维持与共生关系的稳定性主要依赖于性别间特异性吸引物/物质组合的一致性[64]。功能性雄株榕榕果中的雌花花柱较短, 雌株榕果中的雌花花柱较长。由于产卵器长度的限制, 雌性榕小蜂只能在功能性雄株的榕果内产卵, 而进入雌株榕果的榕小蜂无法产生后代。若榕小蜂能够区分不同性别植株, 将不会有榕小蜂进入雌株榕果内传粉, 导致共生关系崩溃[65]。实际研究中发现同种榕不同性别榕果的VOCs不存在显著差异, 而性别间化学拟态假说是目前解释这一现象的主流观点[36, 38, 65]。由于大量研究均显示雌雄异株的榕属植物拥有性别间化学拟态的三个典型特征[61]:(1)在感知能力和学习能力极强的或极度依赖气味信号的传粉体系中最强[27];(2)在欺骗性传粉体系中加强[65];(3)在不同性别花期同步时表现得更加明显, 而在花期异步时减弱[64, 66], 因此普遍认为榕属植物VOCs性别间的相似性是性别间化学拟态的结果。
4 榕-蜂化学通讯驱动协同进化化学通讯关系到榕和榕小蜂的繁殖适合度, 因此与化学通讯相关性状的分化可能会使共生双方失配, 导致灭绝[67];但也可能引发新物种诞生[68—69]。目前认为榕-蜂协同进化的模式主要有2种:协同成种和宿主转移。其中协同成种模式一度被认为是主导榕蜂协同多样化的主要模式[70—71]。对局域环境的适应导致的化学通讯分化是促使同域协同成种的重要模式。但随着大量宿主转移现象以及榕属植物种间杂交事件的发现, 宿主转移或已成为导致榕-蜂协同多样化的又一重要机制[72—74]。
4.1 协同成种严格的协同成种是解释榕-蜂协同多样化的经典理论。“三阶段”假说被用来描述协同成种发生的具体过程:第一阶段, 榕与榕小蜂遵循传统的“一对一”模式, 此时榕小蜂种群与宿主榕种群均未出现明显的遗传分化。第二阶段, 由于榕小蜂世代周期短, 更快地适应局域环境从而产生遗传分化与遗传结构;并且由于传粉者扩散能力的限制, 逐渐影响到宿主榕种群间的基因交流, 从而使榕也逐渐形成显著的种群遗传结构。此时榕小蜂与宿主榕的化学通讯尚未完全分化, 宿主榕的种群间基因流仍能维持, 宿主榕种群间遗传分化在较长时间内维持在较低的水平。第三阶段, 完全分化的榕小蜂间的VOCs产生差异, 进一步限制了宿主榕种群间基因流, 最终促使宿主榕分化成种, 新生成的物种间仍为“一对一”关系[75—76]。在这种成种模式下, 榕与榕小蜂在系统发育上可表现出一致的进化格局[70, 77]。
在协同成种模式中, 化学通讯扮演着重要角色。异域物种形成过程中, 地理隔离直接限制基因流, 而随之产生的化学通讯分化可能进一步促进了生殖隔离的产生, 限制乃至杜绝不同种群日后二次接触时产生基因交流(图 2)。如主要分布于我国台湾的薜荔变种爱玉子(F. pumila var. awkeotsang), 其接受期榕果的气味已经与薜荔产生了差异, 这与二者榕小蜂的偏好性相对应[78]。在地理隔离和化学通讯分化的共同作用下, 爱玉子与薜荔及其榕小蜂正处于协同成种的第二阶段, 即宿主榕的遗传分化程度并未达到种的水平, 但榕小蜂已经完全分化成种[79]。协同成种也可能导致同域物种形成, 化学通讯方式分化则可能是促使榕小蜂遗传分化并进一步引发宿主榕遗传分化的重要驱动力。如分布于巴布新几内亚不同海拔高度的几种近缘榕与传粉榕小蜂协同成种的案例中, 伴随着明显的宿主榕VOCs与榕小蜂气味偏好性的分化, 可能起到了促进二者遗传分化的作用[32]。

此外, 榕对榕小蜂的选择作用也能在两者协同成种过程中起决定性作用。如某些情况下榕能通过种子传播或者引种等先扩散至新生境[80], 并由于环境可塑性而发生气味谱改变。那么, 跟随宿主而来的榕小蜂则需要适应这种新的化学气味才能识别宿主。这种由榕产生的选择作用可能使榕小蜂迅速地发生气味偏好变化, 从而与其他种群产生隔离分化, 最终导致协同成种[69]。
4.2 宿主转移在某些特殊的情况下, 榕小蜂也会为非宿主榕传粉, 甚至进一步成为其他榕的专性传粉者[81—82], 称之为宿主转移。随着宿主转移事件被频繁报导, 加之经典的协同成种假说难以解释在各个分类水平榕-蜂双方系统进化树上普遍存在的不一致情况[72—74], 宿主转移介导的成种假说逐渐受到关注。
化学通讯中发生的“误配”是宿主转移事件发生的重要原因。不同榕属植物, 特别是近缘种之间, 可能会释放较为相似的化学物质, 导致榕小蜂发生误判进入非宿主榕中。如Wang等针对同域分布的5种榕:鸡嗉子榕及其变种(Ficus semicordata var. montana)和另外3种亲缘关系较近的大果榕(Ficus auriculate)、苹果榕(Ficus oligodon)及海南榕(Ficus hainanensis)的研究表明, 亲缘关系更近的榕拥有更相似的气味谱, 同时种间基因流更强;行为学实验也表明5种榕的传粉者平均共享率达24.52%[83]。因此, 当具有相似气味谱的榕同域分布时, 宿主转移便有可能发生。
然而杂交并不仅仅发生在近缘种之间。对已发表的59种榕的叶绿体基因组数据的进化分析表明, 榕的系统发育关系呈现明显的地理聚集格局, 这与其原本的亲缘关系并不相符[84—85]。对6个榕亚属15种榕及对应榕小蜂基因组的系统进化重构分析则发现杂交事件在榕的进化历史中有着较高的发生频率, 这与其榕小蜂频繁发生的宿主转移事件相对应[85]。杂交导致的基因重组为自然选择提供了原材料:杂交后代基因组上形成的“杂交区域”有可能成为基因分化组岛[86], 即这些区域在杂交后代和亲本间的基因流较其他区域弱, 在一定的选择压力下, 基因分化组岛能够不断扩大, 最终使杂交后代与亲本产生生殖隔离, 最终引发成种[86—88](图 2)。如果基因重组区域中包含控制关键性状的基因[89—90], 则可能会导致杂交后代产生与亲本截然不同的气味谱[69]。而榕小蜂的嗅觉偏好可能会因对杂交榕气味的适应而分化, 榕小蜂的分化进一步促进了榕的分化, 最终产生新种[91—92](图 2)。此外, 榕小蜂的偏好性的改变也可能是表型可塑性的结果[93], 而这种嗅觉偏好改变能否最终成为可遗传的性状并最终导致物种的分化目前尚不明确。
5 总结与展望化学通讯是榕-蜂互作的重要纽带。榕VOCs的种间差异性、物候变化特征以及组织间差异性与榕小蜂多样化的气味感知系统为构建专一性的传粉关系打下基础[35, 40, 37, 94]。特化的化学通讯性状由关键基因及特化的转录调控模式决定, 这是榕与榕小蜂间化学通讯模式形成的分子机制[21—23]。化学通讯既维持着榕-蜂种间关系的稳定, 也促进了这两个类群中不同物种的共存, 更可能是协同成种与宿主转移事件发生的催化剂[32, 52, 85, 95]。
尽管对于榕-蜂体系化学通讯机制以及化学通讯介导下物种形成的研究取得了一定的进展, 但仍有许多谜团有待破解。首先, 大部分榕特异性吸引物质未知[21, 23]。同时, 目前对于榕特异性吸引物质以及其合成途径以及相关调控模式的研究十分有限[21, 41, 96]。相应的, 具体、完整的榕小蜂特异性气味识别过程也尚不明确。此外, 物质组合和单一特异性物质如何在化学通讯过程中让榕小蜂产生偏好仍不清楚。要获得上述问题的答案, 需要结合多组学与神经生物学等相关技术加以深入研究[97—98]。
阐明榕-蜂化学通讯机制能够深刻理解化学通讯对二者协同多样化的影响。然而, 目前尚不清楚两者的化学通讯究竟是如何分化并最终引发协同成种[32, 76, 85]。另外, 在化学通讯“误配”介导的宿主转移过程中, 榕小蜂气味偏好发生变化以及与新宿主建立稳定互作关系的机制也不明确。在这些方面取得突破能够揭示触发宿主转移的条件并预测其后果。在全球气候变化的背景下, 物种分布的改变以及物候的改变均会改变传粉者与宿主植物的种间关系[99]。在这种情况下, 榕和榕小蜂有可能发生快速进化或通过宿主转移建立并维持新的传粉关系[100—102]。因此, 深入研究两者间化学通讯的机制能够为预测榕-蜂在全球变化下的命运提供理论依据, 并为理解、保护其他传粉体系提供研究案例示范。
| [1] |
Strong D R, Lawton J H, Southwood T R E. Insects on Plants. Oxford: Blackwell Scientific Publications, 1984.
|
| [2] |
Brummitt N, Araújo A C, Harris T. Areas of plant diversity—What do we know?. Plants, People, Planet, 2021, 3(1): 33-44. DOI:10.1002/ppp3.10110 |
| [3] |
Crepet W L, Niklas K J. Darwin′s second "abominable mystery": Why are there so many angiosperm species?. American Journal of Botany, 2009, 96(1): 366-381. DOI:10.3732/ajb.0800126 |
| [4] |
Steffan-Dewenter I, Westphal C. The interplay of pollinator diversity, pollination services and landscape change. Journal of Applied Ecology, 2008, 45(3): 737-741. |
| [5] |
Johnson S D. The pollination niche and its role in the diversification and maintenance of the southern African flora. Philosophical Transactions of the Royal Society B: Biological Sciences, 2010, 365(1539): 499-516. DOI:10.1098/rstb.2009.0243 |
| [6] |
Vasiliev D, Greenwood S. Pollinator biodiversity and crop pollination in temperate ecosystems, implications for national pollinator conservation strategies: mini review. Science of the Total Environment, 2020, 774: 140880. |
| [7] |
Bouwmeester H, Schuurink R C, Bleeker P M, Schiestl F. The role of volatiles in plant communication. The Plant Journal, 2019, 100(5): 892-907. DOI:10.1111/tpj.14496 |
| [8] |
Raguso R A, Thompson J N, Campbell D R. Improving our chemistry: challenges and opportunities in the interdisciplinary study of floral volatiles. Natural Product Reports, 2015, 32(7): 893-903. DOI:10.1039/C4NP00159A |
| [9] |
Piskorski R, Kroder S, Dorn S. Can pollen headspace volatiles and pollenkitt lipids serve as reliable chemical cues for bee pollinators?. Chemistry & Biodiversity, 2011, 8(4): 577-586. |
| [10] |
Burdon R C F, Raguso R A, Kessler A, Parachnowitsch A L. Spatiotemporal floral scent variation of Penstemon digitalis. Journal of Chemical Ecology, 2015, 41(7): 641-650. DOI:10.1007/s10886-015-0599-1 |
| [11] |
Palmqvist B, Brazeau H A, Parachnowitsch A L. Differences in floral scent and petal reflectance between diploid and tetraploid Chamerion angustifolium. Frontiers in Ecology and Evolution, 2021, 9: 734128. DOI:10.3389/fevo.2021.734128 |
| [12] |
Gervasi D D L, Schiestl F P. Real-time divergent evolution in plants driven by pollinators. Nature Communications, 2017, 8: 14691. DOI:10.1038/ncomms14691 |
| [13] |
Raguso R A. More lessons from linalool: insights gained from a ubiquitous floral volatile. Current Opinion in Plant Biology, 2016, 32: 31-36. DOI:10.1016/j.pbi.2016.05.007 |
| [14] |
Kantsa A, Raguso R A, Lekkas T, Kalantzi O I, Petanidou T. Floral volatiles and visitors: a meta-network of associations in a natural community. Journal of Ecology, 2019, 107(6): 2574-2586. DOI:10.1111/1365-2745.13197 |
| [15] |
Pichersky E, Raguso R A. Why do plants produce so many terpenoid compounds?. New Phytologist, 2018, 220(3): 692-702. DOI:10.1111/nph.14178 |
| [16] |
Zu P J, Boege K, Del-Val E, Schuman M C, Stevenson P C, Zaldivar-Riverón A, Saavedra S. Information arms race explains plant-herbivore chemical communication in ecological communities. Science, 2020, 368(6497): 1377-1381. DOI:10.1126/science.aba2965 |
| [17] |
Jacobsen D J, Raguso R A. Leaf induction impacts behavior and performance of a pollinating herbivore. Frontiers in Plant Science, 2021, 12: 791680. DOI:10.3389/fpls.2021.791680 |
| [18] |
Schiestl F P, Kirk H, Bigler L, Cozzolino S, Desurmont G A. Herbivory and floral signaling: phenotypic plasticity and tradeoffs between reproduction and indirect defense. New Phytologist, 2014, 203(1): 257-266. DOI:10.1111/nph.12783 |
| [19] |
Armbruster W S. The specialization continuum in pollination systems: diversity of concepts and implications for ecology, evolution and conservation. Functional Ecology, 2017, 31(1): 88-100. DOI:10.1111/1365-2435.12783 |
| [20] |
Segar S T, Volf M, Sisol M, Pardikes N A, Souto-Vilarós D. Chemical cues and genetic divergence in insects on plants: conceptual cross pollination between mutualistic and antagonistic systems. Current Opinion in Insect Science, 2019, 32: 83-90. DOI:10.1016/j.cois.2018.11.009 |
| [21] |
Wang R, Yang Y, Jing Y, Segar S T, Zhang Y, Wang G, Chen J, Liu Q F, Chen S, Chen Y, Cruaud A, Ding Y Y, Dunn D W, Gao Q, Gilmartin P M, Jiang K, Kjellberg F, Li H Q, Li Y Y, Liu J Q, Liu M, Machado C A, Ming R, Rasplus J Y, Tong X, Wen P, Yang H M, Yang J J, Yin Y, Zhang X T, Zhang Y Y, Yu H, Yue Z, Compton S G, Chen X Y. Molecular mechanisms of mutualistic and antagonistic interactions in a plant-pollinator association. Nature Ecology & Evolution, 2021, 5(7): 974-986. |
| [22] |
Whitehead M R, Peakall R. Integrating floral scent, pollination ecology and population genetics. Functional Ecology, 2009, 23(5): 863-874. DOI:10.1111/j.1365-2435.2009.01620.x |
| [23] |
Zhang X T, Wang G, Zhang S C, Chen S, Wang Y B, Wen P, Ma X K, Shi Y, Qi R, Yang Y, Liao Z Y, Lin J, Lin J S, Xu X M, Chen X Q, Xu X D, Deng F, Zhao L H, Lee Y L, Wang R, Chen X Y, Lin Y R, Zhang J S, Tang H B, Chen J, Ming R. Genomes of the banyan tree and pollinator wasp provide insights into fig-wasp coevolution. Cell, 2020, 183(4): 875-889. DOI:10.1016/j.cell.2020.09.043 |
| [24] |
Harrison R D. Figs and the diversity of tropical rainforests. BioScience, 2005, 55(12): 1053-1064. DOI:10.1641/0006-3568(2005)055[1053:FATDOT]2.0.CO;2 |
| [25] |
Janzen D H. How to be a fig. Annual Review of Ecology and Systematics, 1979, 10: 13-51. DOI:10.1146/annurev.es.10.110179.000305 |
| [26] |
Weiblen G D. How to be a fig wasp. Annual Review of Entomology, 2002, 47: 299-330. DOI:10.1146/annurev.ento.47.091201.145213 |
| [27] |
Cook J M, Rasplus J Y. Mutualists with attitude: coevolving fig wasps and figs. Trends in Ecology & Evolution, 2003, 18(5): 241-248. |
| [28] |
Chen C, Song Q S, Proffit M, Bessière J M, Li Z B, Hossaert-McKey M. Private channel: a single unusual compound assures specific pollinator attraction in Ficus semicordata. Functional Ecology, 2009, 23(5): 941-950. DOI:10.1111/j.1365-2435.2009.01622.x |
| [29] |
Proffit M, Chen C, Soler C, Bessière J M, Schatz B, Hossaert-McKey M. Can chemical signals, responsible for mutualistic partner encounter, promote the specific exploitation of nursery pollination mutualisms? -The case of figs and fig wasps. Entomologia Experimentalis et Applicata, 2009, 131(1): 46-57. DOI:10.1111/j.1570-7458.2009.00823.x |
| [30] |
Proffit M, Lapeyre B, Buatois B, Deng X X, Arnal P, Gouzerh F, Carrasco D, Hossaert-McKey M. Chemical signal is in the blend: bases of plant-pollinator encounter in a highly specialized interaction. Scientific Reports, 2020, 10(1): 10071. DOI:10.1038/s41598-020-66655-w |
| [31] |
Rodriguez L J, Bain A, Chou L S, Conchou L, Cruaud A, Gonzales R, Hossaert-McKey M, Rasplus J Y, Tzeng H Y, Kjellberg F. Diversification and spatial structuring in the mutualism between Ficus septica and its pollinating wasps in insular South East Asia. BMC Evolutionary Biology, 2017, 17(1): 207. DOI:10.1186/s12862-017-1034-8 |
| [32] |
Souto-Vilarós D, Proffit M, Buatois B, Rindos M, Sisol M, Kuyaiva T, Isua B, Michalek J, Darwell C T, Hossaert-McKey M, Weiblen G D, Novotny V, Segar S T. Pollination along an elevational gradient mediated both by floral scent and pollinator compatibility in the fig and fig-wasp mutualism. Journal of Ecology, 2018, 106(6): 2256-2273. DOI:10.1111/1365-2745.12995 |
| [33] |
Knudsen J T, Eriksson R, Gershenzon J, Ståhl B. Diversity and distribution of floral scent. The Botanical Review, 2006, 72(1): 1-120. DOI:10.1663/0006-8101(2006)72[1:DADOFS]2.0.CO;2 |
| [34] |
Proffit M, Johnson S D. Specificity of the signal emitted by figs to attract their pollinating wasps: comparison of volatile organic compounds emitted by receptive syconia of Ficus sur and F. sycomorus in Southern Africa. South African Journal of Botany, 2009, 75(4): 771-777. DOI:10.1016/j.sajb.2009.08.006 |
| [35] |
Grison-Pigé L, Hossaert-McKey M, Greeff J M, Bessière J M. Fig volatile compounds——a first comparative study. Phytochemistry, 2002, 61(1): 61-71. DOI:10.1016/S0031-9422(02)00213-3 |
| [36] |
Grison-Pigé L, Bessière J M, Turlings T C J, Kjellberg F, Roy J, Hossaert-McKey M. Limited intersex mimicry of floral odour in Ficus carica. Functional Ecology, 2001, 15(4): 551-558. DOI:10.1046/j.0269-8463.2001.00553.x |
| [37] |
Proffit M, Bessière J M, Schatz B, Hossaert-McKey M. Can fine-scale post-pollination variation of fig volatile compounds explain some steps of the temporal succession of fig wasps associated with Ficus racemosa?. Acta Oecologica, 2018, 90: 81-90. DOI:10.1016/j.actao.2017.08.009 |
| [38] |
Proffit M, Schatz B, Bessière J M B, Chen C, Soler C, Hossaert-McKey M. Signalling receptivity: comparison of the emission of volatile compounds by figs of Ficus hispida before, during and after the phase of receptivity to pollinators. Symbiosis, 2008, 45(1/3): 15-24. |
| [39] |
Van Noort S, Ware A B, Compton S G. Pollinator-specific volatile attractants released from the figs of Ficus burtt-davyi. South African Journal of Science, 1989, 85(5): 323-324. |
| [40] |
Souza C D, Pereira R A S, Marinho C R, Kjellberg F, Teixeira S P. Diversity of fig glands is associated with nursery mutualism in fig trees. American Journal of Botany, 2015, 102(10): 1564-1577. DOI:10.3732/ajb.1500279 |
| [41] |
Hu R, Sun P, Yu H, Cheng Y F, Wang R, Chen X Y, Kjellberg F. Similitudes and differences between two closely related Ficus species in the synthesis by the ostiole of odors attracting their host-specific pollinators: a transcriptomic based investigation. Acta Oecologica, 2020, 105: 103554. DOI:10.1016/j.actao.2020.103554 |
| [42] |
王楠. 对叶榕传粉榕小蜂气味结合蛋白和化学感受蛋白基因的进化和表达[D]. 泰安: 山东农业大学, 2014.
|
| [43] |
Wang N, Wang N X, Niu L M, Bian S N, Xiao J H, Huang D W. Odorant-binding protein (OBP) genes affect host specificity in a fig-pollinator mutualistic system. Insect Molecular Biology, 2014, 23(5): 621-631. DOI:10.1111/imb.12110 |
| [44] |
Li Z B, Yang P, Peng Y Q, Yang D R. Ultrastructure of antennal sensilla of female Ceratosolen solmsi marchali (Hymenoptera: Chalcidoidea: Agaonidae: Agaoninae). The Canadian Entomologist, 2009, 141(5): 463-477. DOI:10.4039/n09-011 |
| [45] |
李宗波, 杨培, 彭艳琼, 杨大荣. 木瓜榕传粉榕小蜂雌蜂触角感器的分布和超微形态. 昆虫学报, 2012, 55(11): 1272-1281. DOI:10.16380/j.kcxb.2012.11.010 |
| [46] |
李宗波, 杨培, 杨大荣. 钝叶榕三种进果榕小蜂雌性触角的结构、感器及其适生意义. 环境昆虫学报, 2013, 35(2): 217-225. DOI:10.3969/j.issn.1674-0858.2013.02.13 |
| [47] |
杨培, 李宗波, 杨大荣, 彭艳琼. 木瓜榕食果榕小蜂雌蜂触角感器的扫描电镜观察. 生物学杂志, 2017, 34(4): 11-16. |
| [48] |
孟晶, 卜文俊, 肖金花, 黄大卫. 中国榕小蜂触角感受器形态特征及进化适应性分析. 昆虫学报, 2015, 58(7): 800-810. DOI:10.16380/j.kcxb.2015.07.013 |
| [49] |
李宗波, 杨培, 赵健, 杨大荣. 传粉榕小蜂雌蜂多孔板形感器的形态与适应进化. 应用昆虫学报, 2014, 51(4): 1086-1097. |
| [50] |
刘志祥, 杨培, 李宗波. 鸡嗉子榕蜂群落中雌蜂触角感器及适生意义. 应用昆虫学报, 2021, 58(3): 736-746. |
| [51] |
Meng J, Huang D W, Xiao J H, Bu W J. Antennal sensilla of fig wasps (Hymenoptera: Agaonidae): function-driven elaboration in females and degeneration in males. Annals of the Entomological Society of America, 2016, 109(1): 99-105. DOI:10.1093/aesa/sav084 |
| [52] |
Soler C, Proffit M, Chen C, Hossaert-McKey M. Private channels in plant-pollinator mutualisms. Plant Signaling & Behavior, 2010, 5(7): 893-895. |
| [53] |
Schaefer H M, Schaefer V, Levey D J. How plant-animal interactions signal new insights in communication. Trends in Ecology & Evolution, 2004, 19(11): 577-584. |
| [54] |
Raguso R A. Wake up and smell the roses: the ecology and evolution of floral scent. Annual Review of Ecology, Evolution, and Systematics, 2008, 39: 549-569. DOI:10.1146/annurev.ecolsys.38.091206.095601 |
| [55] |
陈友玲, 方丽娜, 吴文珊. 信息化合物对薜荔榕小蜂选择行为的影响. 生态学报, 2010, 30(11): 2949-2957. |
| [56] |
Warren M, Robertson M P, Greeff J M. A comparative approach to understanding factors limiting abundance patterns and distributions in a fig tree-fig wasp mutualism. Ecography, 2010, 33(1): 148-158. DOI:10.1111/j.1600-0587.2009.06041.x |
| [57] |
Yang Y, Zhang Y Y, Zhang Y, Chen S, Li Q Y, Wang R, Chen X Y. Selection to attract pollinators and to confuse antagonists specializes fig-pollinator chemical communications. Journal of Systematics and Evolution, in press.
|
| [58] |
Xiao J H, Yue Z, Jia L Y, Yang X H, Niu L H, Wang Z, Zhang P, Sun B F, He S M, Li Z, Xiong T L, Xin W, Gu H F, Wang B, Werren J H, Murphy R W, Wheeler D, Niu L M, Ma G C, Tang T, Bian S N, Wang N X, Yang C Y, Wang N, Fu Y G, Li W Z, Yi S V, Yang X Y, Zhou Q, Lu C X, Xu C Y, He L J, Yu L L, Chen M, Zheng Y, Wang S W, Zhao S, Li Y H, Yu Y Y, Qian X J, Cai Y, Bian L L, Zhang S, Wang J Y, Yin Y, Xiao H, Wang G H, Yu H, Wu W S, Cook J M, Wang J, Huang D W. Obligate mutualism within a host drives the extreme specialization of a fig wasp genome. Genome Biology, 2013, 14(12): R141. DOI:10.1186/gb-2013-14-12-r141 |
| [59] |
Soler C, Hossaert-McKey M, Buatois B, Bessière J M, Schatz B, Proffit M. Geographic variation of floral scent in a highly specialized pollination mutualism. Phytochemistry, 2011, 72(1): 74-81. DOI:10.1016/j.phytochem.2010.10.012 |
| [60] |
Soler C C L, Schatz B, Bessière J M, Hossaert-McKey M. Geographic variation of fruit scents in a dispersion mutualism, the case of Ficus lutea. Acta Oecologica, 2018, 90: 46-48. DOI:10.1016/j.actao.2017.11.014 |
| [61] |
Ashman T L. Sniffing out patterns of sexual dimorphism in floral scent. Functional Ecology, 2009, 23(5): 852-862. DOI:10.1111/j.1365-2435.2009.01590.x |
| [62] |
Gu D, Compton S G, Peng Y Q, Yang D R. ′Push′ and ′pull′ responses by fig wasps to volatiles released by their host figs. Chemoecology, 2012, 22(4): 217-227. DOI:10.1007/s00049-012-0108-8 |
| [63] |
Verkerke W. Structure and function of the fig. Experientia, 1989, 45(7): 612-622. DOI:10.1007/BF01975678 |
| [64] |
Hossaert-McKey M, Proffit M, Soler C C L, Chen C, Bessière J M, Schatz B, Borges R M. How to be a dioecious fig: chemical mimicry between sexes matters only when both sexes flower synchronously. Scientific Reports, 2016, 6(1): 21236. DOI:10.1038/srep21236 |
| [65] |
Yang J, Niu Y, Sun W B, Cai X H, Chen G. Intersexual mimicry and imperfect deceit of a threatened aquatic herb Ottelia acuminata. Journal of Systematics and Evolution, 2022, 60(2): 377-385. DOI:10.1111/jse.12657 |
| [66] |
Soler C C L, Proffit M, Bessière J M, Hossaert-Mckey M, Schatz B. Evidence for intersexual chemical mimicry in a dioecious plant. Ecology Letters, 2012, 15(9): 978-985. DOI:10.1111/j.1461-0248.2012.01818.x |
| [67] |
Sachs J L, Simms E L. Pathways to mutualism breakdown. Trends in Ecology & Evolution, 2006, 21(10): 585-592. |
| [68] |
Faucci A, Toonen R J, Hadfield M G. Host shift and speciation in a coral-feeding nudibranch. Proceedings of the Royal Society B: Biological Sciences, 2007, 274(1606): 111-119. DOI:10.1098/rspb.2006.3685 |
| [69] |
Herre E A, Jandér K C, Machado C A. Evolutionary ecology of figs and their associates: recent progress and outstanding puzzles. Annual Review of Ecology, Evolution, and Systematics, 2008, 39: 439-458. DOI:10.1146/annurev.ecolsys.37.091305.110232 |
| [70] |
Rønsted N, Weiblen G D, Cook J M, Salamin N, Machado C A, Savolainen V. 60 million years of co-divergence in the fig-wasp symbiosis. Proceedings of the Royal Society B: Biological Sciences, 2005, 272(1581): 2593-2599. DOI:10.1098/rspb.2005.3249 |
| [71] |
Cruaud A, Rønsted N, Chantarasuwan B, Chou L S, Clement W L, Couloux A, Cousins B, Genson G, Harrison R D, Hanson P E, Hossaert-McKey M, Jabbour-Zahab R, Jousselin E, Kerdelhué C, Kjellberg F, Lopez-Vaamonde C, Peebles J, Peng Y Q, Pereira R A S, Schramm T, Ubaidillah R, Van Noort S, Weiblen G D, Yang D R, Yodpinyanee A, Libeskind-Hadas R, Cook J M, Rasplus J Y, Savolainen V. An extreme case of plant-insect codiversification: figs and fig-pollinating wasps. Systematic Biology, 2012, 61(6): 1029-1047. DOI:10.1093/sysbio/sys068 |
| [72] |
Machado C A, Robbins N, Gilbert M T P, Herre E A. Critical review of host specificity and its coevolutionary implications in the fig/fig-wasp mutualism. Proceedings of the National Academy of Sciences of the United States of America, 2005, 102(S1): 6558-6565. |
| [73] |
Cook J M, Segar S T. Speciation in fig wasps. Ecological Entomology, 2010, 35(S1): 54-66. |
| [74] |
Yang L Y, Machado C A, Dang X D, Peng Y Q, Yang D R, Zhang D Y, Liao W J. The incidence and pattern of copollinator diversification in dioecious and monoecious figs. Evolution, 2015, 69(2): 294-304. DOI:10.1111/evo.12584 |
| [75] |
陈艳, 李宏庆, 刘敏, 陈小勇. 榕-传粉榕小蜂间的专一性与协同进化. 生物多样性, 2010, 18(1): 1-10. |
| [76] |
Souto-Vilarós D, Machac A, Michalek J, Darwell C T, Sisol M, Kuyaiva T, Isua B, Weiblen G D, Novotny V, Segar S T. Faster speciation of fig-wasps than their host figs leads to decoupled speciation dynamics: snapshots across the speciation continuum. Molecular Ecology, 2019, 28(17): 3958-3976. DOI:10.1111/mec.15190 |
| [77] |
Cruaud A, Jabbour-Zahab R, Genson G, Cruaud C, Couloux A, Kjellberg F, Van Noort S, Rasplus J Y. Laying the foundations for a new classification of Agaonidae (Hymenoptera: Chalcidoidea), a multilocus phylogenetic approach. Cladistics, 2010, 26(4): 359-387. |
| [78] |
杨升, 陈友铃, 吴文珊, 陈晓娟, 包甜甜, 叶洁芬, 宋天宇, 吴怡祾, 周莲香, 曾喜育. 薜荔榕小蜂对薜荔和爱玉子雌花期榕果挥发物的行为反应. 生态学报, 2017, 37(21): 7161-7169. |
| [79] |
Liu M, Zhao R, Chen Y, Zhang J, Compton S G, Chen X Y. Competitive exclusion among fig wasps achieved via entrainment of host plant flowering phenology. PLoS One, 2014, 9(5): e97783. DOI:10.1371/journal.pone.0097783 |
| [80] |
Wang R, Chen X Y, Chen Y, Wang G, Dunn D W, Quinnell R J, Compton S G. Loss of top-down biotic interactions changes the relative benefits for obligate mutualists. Proceedings of the Royal Society B Biological Sciences, 2019, 286(1897): 20182501. DOI:10.1098/rspb.2018.2501 |
| [81] |
Molbo D, Machado C A, Sevenster J G, Keller L, Herre E A. Cryptic species of fig-pollinating wasps: implications for the evolution of the fig-wasp mutualism, sex allocation, and precision of adaptation. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100(10): 5867-5872. DOI:10.1073/pnas.0930903100 |
| [82] |
Tsai L, Hayakawa H, Fukuda T, Yokoyama J. A breakdown of obligate mutualism on a small island: An interspecific hybridization between closely related fig species (Ficus pumila and Ficus thunbergii) in Western Japan. American Journal of Plant Sciences, 2015, 6(1): 126-131. DOI:10.4236/ajps.2015.61014 |
| [83] |
Wang G, Cannon C H, Chen J. Pollinator sharing and gene flow among closely related sympatric dioecious fig taxa. Proceedings of the Royal Society B Biological Sciences, 2016, 283(1828): 20152963. |
| [84] |
Bruun-Lund S, Clement W L, Kjellberg F, Rønsted N. First plastid phylogenomic study reveals potential cyto-nuclear discordance in the evolutionary history of Ficus L. (Moraceae). Molecular Phylogenetics and Evolution, 2017, 109: 93-104. |
| [85] |
Wang G, Zhang X T, Herre E A, McKey D, Machado C A, Yu W B, Cannon C H, Arnold M L, Pereira R A S, Ming R, Liu Y F, Wang Y B, Ma D N, Chen J. Genomic evidence of prevalent hybridization throughout the evolutionary history of the fig-wasp pollination mutualism. Nature Communications, 2021, 12(1): 718. |
| [86] |
Abbott R, Albach D, Ansell S, Arntzen J W, Baird S J E, Bierne N, Boughman J, Brelsford A, Buerkle C A, Buggs R, Butlin R K, Dieckmann U, Eroukhmanoff F, Grill A, Cahan S H, Hermansen J S, Hewitt G, Hudson A G, Jiggins C, Jones J, Keller B, Marczewski T, Mallet J, Martinez-Rodriguez P, Möst M, Mullen S, Nichols R, Nolte A W, Parisod C, Pfennig K, Rice A M, Ritchie M G, Seifert B, Smadja C M, Stelkens R, Szymura J M, Väinölä R, Wolf J B W, Zinner D. Hybridization and speciation. Journal of Evolutionary Biology, 2013, 26(2): 229-246. |
| [87] |
Mallet J. Hybrid speciation. Nature, 2007, 446(7133): 279-283. |
| [88] |
Payseur B A, Rieseberg L H. A genomic perspective on hybridization and speciation. Molecular Ecology, 2016, 25(11): 2337-2360. |
| [89] |
Capblancq T, Després L, Rioux D, Mavárez J. Hybridization promotes speciation in Coenonympha butterflies. Molecular Ecology, 2015, 24(24): 6209-6222. |
| [90] |
MacLeod A, Rodríguez A, Vences M, Orozco-terWengel P, García C, Trillmich F, Gentile G, Caccone A, Quezada G, Steinfartz S. Hybridization masks speciation in the evolutionary history of the Galápagos marine iguana. Proceedings of the Royal Society B Biological Sciences, 2015, 282(1809): 20150425. |
| [91] |
Vereecken N J, Cozzolino S, Schiestl F P. Hybrid floral scent novelty drives pollinator shift in sexually deceptive orchids. BMC Evolutionary Biology, 2010, 10(1): 103. |
| [92] |
Ayasse M, Stökl J, Francke W. Chemical ecology and pollinator-driven speciation in sexually deceptive orchids. Phytochemistry, 2011, 72(13): 1667-1677. |
| [93] |
Proffit M, Khallaf M A, Carrasco D, Larsson M C, Anderson P. 'Do you remember the first time?′ Host plant preference in a moth is modulated by experiences during larval feeding and adult mating. Ecology Letters, 2015, 18(4): 365-374. |
| [94] |
Yang P, Li Z B, Yang D R, Peng Y Q, Kjellberg F. Comparison of the antennal sensilla of females of four fig-wasps associated with Ficus auriculata. Acta Oecologica, 2018, 90: 99-108. |
| [95] |
Gu D, Yang D R, Yang P, Peng Y Q, Wang Z J. Work division of floral scent compounds in mediating pollinator behaviours. Chemistry and Ecology, 2016, 32(8): 733-741. |
| [96] |
Yu H, Nason J D, Zhang L, Zheng L N, Wu W, Ge X J. De novo transcriptome sequencing in Ficus hirta Vahl. (Moraceae) to investigate gene regulation involved in the biosynthesis of pollinator attracting volatiles. Tree Genetics & Genomes, 2015, 11(3): 91. |
| [97] |
Zeng F F, Xu P X, Leal W S. Odorant receptors from Culex quinquefasciatus and Aedes aegypti sensitive to floral compounds. Insect Biochemistry and Molecular Biology, 2019, 113: 103213. |
| [98] |
Lahondère C, Vinauger C, Okubo R P, Wolff G H, Chan J K, Akbari O S, Riffell J A. The olfactory basis of orchid pollination by mosquitoes. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(1): 708-716. |
| [99] |
王晴晴, 高燕, 王嵘. 全球变化对食物网结构影响机制的研究进展. 植物生态学报, 2021, 45(10): 1064-1074. |
| [100] |
Compton S G. A collapse of host specificity in some African fig wasps. South African Journal of Science, 1990, 86(1): 39-40. |
| [101] |
William R B. Hybridization of Ficus religiosa with F. septica and F. aurea (Moraceae). Revista de Biologia Tropical, 1994, 42(1/2): 339-342. |
| [102] |
Wei Z D, Kobmoo N, Cruaud A, Kjellberg F. Genetic structure and hybridization in the species group of Ficus auriculata: can closely related sympatric Ficus species retain their genetic identity while sharing pollinators?. Molecular Ecology, 2014, 23(14): 3538-3550. |