 2023, Vol. 43
2023, Vol. 43文章信息
- 王怡萌, 段磊磊, 陈聪, 王铭, 王升忠, 赵婧
- WANG Yimeng, DUAN Leilei, CHEN Cong, WANG Ming, WANG Shengzhong, ZHAO Jing
- 不同水位管理对恢复泥炭地土壤CO2、CH4排放的影响
- Effects of different hydrologic managements on soil CO2 and CH4 emissions of the restored peatlands
- 生态学报. 2023, 43(11): 4583-4593
- Acta Ecologica Sinica. 2023, 43(11): 4583-4593
- http://dx.doi.org/10.5846/stxb202111013067
-
文章历史
- 收稿日期: 2021-11-01
- 网络出版日期: 2023-02-07

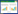




2. 长白山地理过程与生态安全教育部重点实验室, 长春 130024;
3. 东北师范大学国家环境保护湿地生态与植被恢复重点实验室, 长春 130024
2. Key Laboratory of Geographical Progresses and Ecological Security of Changbai Mountains, Ministry of Education, Changchun 130024, China;
3. Key Laboratory of Wetland Ecology and Vegetation Restoration of National Environmental Protection, Changchun 130024, China
泥炭地是重要的沼泽湿地类型, 占陆地面积的2.84%[1], 但存储了大于1/3的陆地土壤碳[2]。泥炭地泥炭的累积过程是大气向陆地的碳汇过程, 因而泥炭地对控制大气温室气体含量和减缓温室效应起着重要作用[3]。然而, 近年来, 受农业开垦、泥炭开采、过度放牧和基础设施建设等人为干扰, 全球泥炭地面积不断缩减[4]。21世纪初全球约12.5%的泥炭沼泽已经消失, 而农业开垦是导致泥炭沼泽退化的主要原因[5]。泥炭地排水改造后, 泥炭好氧分解, 可能使泥炭地从“碳汇”转变为“碳源”, 加剧全球变暖[6—7]。随着对泥炭地生态价值及其在全球气候变化中重要性认识的提高, 退化泥炭地生态恢复工程在全球范围内逐步开展[8—9]。
泥炭地的生态恢复包括植被恢复、基质恢复与水文恢复等, 其中水文恢复被认为是泥炭地恢复与重建的重要前提[10]。泥炭地水文条件影响泥炭地生物地球化学循环, 控制和维持着泥炭地生态系统的结构和功能[11]。对退化泥炭地进行水文恢复可以减弱泥炭地对于消极气候变化的敏感性, 对于其碳汇功能的恢复尤为重要[12]。当前对于退化泥炭地水文恢复的研究主要集中在土壤基本理化性质、植被群落结构以及微生物活性的响应等方面[13—14]。而对于不同水位条件下, 恢复泥炭地土壤CH4和CO2如何响应尚存在争议。一般认为, 较低的水位有利于土壤微生物好氧呼吸, 促进土壤CO2排放[15];同时较低的地下水位导致CH4被氧化消耗, 减少CH4的排放[16]。而水位的升高为产甲烷菌生长繁殖提供有利条件, 导致CH4的大量产生[17]。然而, Kannenberg[18]的研究发现, 与长期淹水的湿地土壤相比, 经历短期干旱后再湿润的恢复, 湿地土壤会产生更多的CH4。同时, 有研究发现退化泥炭地水位抬升后土壤CO2排放减少带来的降温效果可能会被CH4排放增加所抵消[19], 但也有研究发现水位抬升后土壤CH4排放量并未显著升高[20]。由此可见, 不同水位条件下恢复泥炭地碳排放还存在较多的不确定性, 尚需较多的野外监测明确恢复泥炭地土壤CO2、CH4排放对不同水文条件的响应及机制。
长白山区泥炭地是我国泥炭地集中分布区之一, 区域内泥炭地面积463.31 km2, 有机碳储量高达47.64 Tg[21]。自20世纪50年代, 该区泥炭地受农业开垦、排水造林等干扰, 泥炭地面积大幅减少[22]。以长白山区通化县天然、退化及不同水位管理的恢复泥炭地为研究对象, 通过野外原位监测, 分析天然泥炭地、不同水位管理恢复泥炭地(高水位、低水位及高低交替水位)及退化泥炭地土壤CO2、CH4排放差异, 揭示恢复泥炭地土壤CO2、CH4排放对不同水位管理的响应及机制, 探究具有较低增温潜能的最佳水位, 以期为我国泥炭地的生态水文恢复及水位管理提供科学参考。
1 研究区概况与研究方法 1.1 研究区概况研究区位于吉林省通化市四棚乡(125°34′44"E, 41°51′22"N, 海拔514 m, 图 1), 地处长白山西南麓, 属中温带湿润气候区, 年平均气温5.5 ℃, 年平均降水量870 mm。独特的地形、气候环境使该区发育有草本泥炭沼泽, 泥炭层厚度为0.8—1.2 m。2000年左右, 部分泥炭地被开垦为水田, 2010年在当地政府号召下退耕。但由于农业垦殖、农田灌溉及地下水的大量使用, 该处开垦泥炭沼泽的原生植被已遭到破坏, 沼泽水位变浅, 杂草丛生, 地表植被以灯芯草(Juncus bufonius)、野稗(Echinochloa crusgali)、鬼针草(Bidens pilosa)等非典型泥炭地植物为主。在未进行人为干扰的情况下, 退化泥炭地难以进行自我恢复。退化泥炭地生态恢复工程自2019年4月开始进行, 植被恢复以瘤囊苔草移栽为主, 移栽间隔为50 cm, 苔草移栽后植被盖度为30%—50%。恢复泥炭地设置3个水文管理梯度, 分别为高水位区、高-低水位交替区及低水位区。

|
| 图 1 研究区位置及样地分布 Fig. 1 The location of the study site and distribution of sampling plots NP:天然泥炭地Natural peatland;DP:退化泥炭地Degraded peatland;LR:低水位恢复泥炭地Low water level restored peatland;H-LR:高低水位交替恢复泥炭地High-low alternating water level restored peatland;HR:高水位恢复泥炭地High water level restored peatland |
本研究设置5个研究样地:天然区(NP)、高水位恢复区(HR)、高低水位交替恢复区(H-LR)、低水位恢复区(LR)及退化区(DP)(图 1)。天然区、恢复区(高水位、高-低水位交替、低水位)及退化区面积均>700 m2。天然区:未进行任何人为扰动, 为天然苔草群落, 面积约1.8 hm2。恢复区:2019年4月进行恢复, 首先对地表进行枯落物去除-松耙-灌水-搅耙-清除表层残体, 使恢复区统一形成镜面田, 而后对其进行苔草移栽, 5月初对恢复区进行不同水位梯度控制。研究区北部的蓄水池可为恢复区补水。高水位区:对其进行补水, 使其水位控制在5—10 cm;高-低水位交替区:对其进行15 d一个间隔的高-低水位循环处理;低水位区:不进行补水, 使其水位控制在-10—0 cm。退化区:为泥炭地开垦后的弃耕地, 退耕后未进行人为扰动, 不进行补水。水文管理从生长季5月进行至10月, 生长季平均水位见表 1。在每个研究样地随机设置3个2 m×2 m的研究小区, 各小区间间隔>20 m, 在研究小区内设置静态箱并进行土壤样品的采集。
| 样地 Samples |
总有机碳 TOC/(g/kg) |
总氮 TN/(g/kg) |
总磷 TP/(g/kg) |
可溶性有机碳 DOC/(mg/kg) |
酸碱度 pH |
土壤含水量 SWC/% |
水位 WL/cm |
| NP天然区 | 333.48±1.25a | 22.35±0.21a | 1.15±0.01c | 774.53±27.36a | 5.92±0.01c | 615.47±27.91a | -1.03±0.47b |
| DP退化区 | 57.34±1.01c | 5.29±0.02b | 1.59±0.01a | 319.69±2.21b | 6.34±0.04a | 95.01±2.77b | -4.25±0.60c |
| LR低水位区 | 60.65±0.87c | 5.07±0.08b | 1.41±0.01b | 213.19±3.34c | 6.03±0.04b | 83.89±0.73c | -3.67±0.70c |
| H-LR高低水位交替区 | 60.68±0.84c | 4.95±0.03b | 1.72±0.04a | 157.97±9.79e | 6.11±0.02b | 102.56±1.89b | -0.07±0.70b |
| HR高水位区 | 66.10±1.08b | 5.37±0.12b | 1.72±0.01a | 193.34±5.19d | 6.27±0.01ab | 105.41±4.55b | 7.49±0.08a |
| TOC:总有机碳Total organic carbon;TN:总氮Total nitrogen;TP:总磷Total phosphorus;DOC:可溶性有机碳Dissolved organic carbon;pH:酸碱度;SWC:土壤含水量Soil water content;WL:水位Water level表中数值为平均值±标准误(n=3); 同列不同小写字母表示同一指标在不同样地间差异显著(P<0.05) | |||||||
利用静态箱-气相色谱法进行土壤CO2和CH4气体样品采集, 采样时间为2020年6月—10月。静态箱由集气箱和不锈钢底座两部分组成。底座由内圈和外圈组成, 直径分别为36 cm和40 cm, 高分别为19 cm和9 cm。内圈和外圈之间有一个宽4 cm的凹槽。集气箱是一个圆形聚氯乙烯(PVC)管, 有顶无底, 其直径和高分别为38 cm和50 cm。为了减小对土壤的扰动, 于6月第一次采样前一周安装静态箱底座, 直至10月最后一次采样完成后取出。采样频率为两星期一次, 在晴朗无云的上午9:00—11:00采集气体样品, 气体样品采集完成后即刻对需要补水或排水的样地进行补水或排水。采样前一天清除地表植被以及凋落物, 采样时将静态箱放置于底座的凹槽上, 加水密封后用60 mL注射器立即采集气体, 间隔10 min采集一次气体样品, 30 min内共采集4个样品, 并记录近地表空气温度。采集的气体在一周内使用气相色谱仪(GC7890B)完成分析, 气体排放速率计算公式:
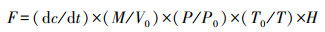
|
式中:F表示气体排放速率(mg m-2 h-1), dc/dt 表示静态箱内气体浓度变化率, M为气体摩尔质量(g/mol), V0为标准状态下的所测气体摩尔体积(mL/mol), P0、T0分别为标准状态下空气绝对气压(Pa)和温度(℃), H为静态箱内空气高度(m)。
5—10月气体累积碳排放量计算公式为:
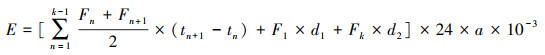
|
其中:E为气体累积碳排放量(g/m2), k为总的采样次数, n为第n次采样, Fn为第n次的气体排放速率(mg m-2 h-1), tn+1-tn为第n+1次与第n次采样的间隔天数。F1和Fk分别为第一次和最后一次采样日的气体排放速率, d1为5月1日至第一次采样日期的间隔天数, d2为最后一次采样日期至10月31日的间隔天数。a为气体分子量中的碳原子占比(CO2为0.273, CH4为0.75)。
CH4和CO2的增温效果用全球增温潜势表示:
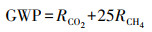
|
式中:GWP为全球增温潜势(t/hm2), RCO2、RCH4表示观测期间CO2、CH4累积排放量(t/hm2), 100 a时间尺度上, CH4的增温潜势为CO2的25倍[23]。
1.4 土壤样品采集与分析在每个样地中安装水位计(HOBO U20L-02水位计, Oneset公司, 美国;Odyssey ODYWL10水位计, Dataflow公司, 新西兰)监测地下水位(WL), 并在5月、7月、10月采集样方表层0—10 cm土壤, 带回实验室后冷藏储存。在实验室, 将采集的土壤过2 mm筛, 去除石块及植物根系并混合均匀, 用于理化指标测定。
土壤含水量(SWC)使用烘干称重法测量;土壤pH使用奥豪斯ST3100酸度计测定;全磷(TP)测定使用微波消解钼锑抗比色法。土壤有机碳(TOC)及总氮(TN)使用EA3000元素分析仪测定[24];可溶性有机碳(Dissolved organic carbon, DOC)使用Aurora 1030C有机碳分析仪测定。
1.5 统计分析在分析不同样地间土壤理化指标及CO2、CH4累积碳排放量、增温潜势的差异之前, 使用Shapiro. test函数以及Levene Test函数检验数据是否符合正态性、方差齐性, 满足条件的数据使用方差分析检验组间差异并使用Tukey HSD函数进行比较。不满足上述条件时, 使用Kruckal. test函数检验组间差异, 利用T. test函数进行比较。使用线性回归模型(R=a+bW), 拟合CO2、CH4排放速率与水位的关系;使用指数模型(R=aebT)拟合CO2排放速率与温度的关系, 并计算温度敏感性Q10(Q10=e10b)。采用Pearson相关分析法, 分析水位与CO2、CH4累积碳排放量的相关性。统计分析和制图使用R语言4.0.3和Origin 2021完成。
2 结果分析 2.1 土壤理化性质研究区NP土壤TOC、TN、DOC和SWC显著高于恢复区和DP, 但其TP和pH显著低于恢复区和DP(P < 0.05)(表 1)。DP的TOC显著低于NP和恢复区, 但其pH和DOC保持较高水平。恢复区中, HR和H-LR的TP和SWC显著高于LR, HR的TOC显著高于H-LR和LR, LR的DOC显著高于HR和H-LR, 三者的pH和TP不存在显著差异(P>0.05)(表 1)。
生长季, NP、DP、LR、HR、H-LR水位的变化模式不同(图 2), HR一直处于淹水状态, 且水位波动较小;NP、DP、LR呈现相似的水位变化, 受降雨的影响, 6月下旬至8月上旬水位波动剧烈, NP水位波动(-12.86 cm—4.35 cm)小于DP(-28.16 cm—1.01 cm)和LR(-35.92 cm—2.04 cm), 9月之后水位趋于平稳。H-LR的水位除了受降雨影响外, 还呈现出明显的人为调控模式。生长季HR的水位显著高于其它样地(P < 0.05), DP和LR水位较低, 分别为(-4.25±0.60)cm和(-3.67±0.70)cm(表 1)。

|
| 图 2 研究区水位及近地表温度的季节动态 Fig. 2 Seasonal dynamics of water table and air temperature |
研究区平均近地表温度为(25.13±0.07)℃。6月开始气温逐渐上升, 最高气温(31.21±0.33)℃出现在8月中旬;9月之后, 气温迅速降低, 最低温为(17.52±0.80) ℃(图 2)。各样地之间近地表温度无显著差异(P>0.05)。DP的平均温最高为(25.31±0.60)℃, HR的平均温最低为(24.92±0.65)℃。
2.2 不同水位管理泥炭地土壤CO2、CH4排放特征研究区土壤CO2排放速率呈明显的季节动态(图 3), NP、DP、LR表现出相似的单峰模式, 峰值出现在7月中旬(850.68—1135.57 mg CO2 m-2h-1);9月之后排放速率较低, 趋于稳定。HR的土壤CO2排放速率也呈现单峰型, 但峰值出现在8月中旬(309.86 mg CO2 m-2 h-1)。H-LR 6—8月土壤CO2排放速率波动较大, 低水位期排放速率较高, 高水位期排放速率低, 9—10月CO2排放速率较低, 且波动较小(图 3)。各样地间, DP的土壤CO2累积碳排放量最高(675.1 g CO2-C/m2), 显著高于各恢复区(P < 0.05);HR的土壤CO2累积碳排放量最低;NP、LR和H-LR间CO2-C累积排放量无显著差异(P>0.05, 图 3)。

|
| 图 3 研究区土壤CO2排放速率季节变化及CO2-C排放量变化 Fig. 3 Seasonal variation in soil CO2 emission and changes in soil CO2-C emissions 不同小写字母表示同一指标在不同样地间差异显著(P < 0.05) |
研究区土壤CH4排放速率呈现出与CO2不同的模式(图 4)。生长季, LR与DP的土壤CH4排放速率较低, 无明显的季节变化特征(图 4);H-LR与NP的土壤CH4排放速率随时间先升高后降低, 峰值出现在8月中旬;HR的土壤CH4排放速率呈现出双峰模式。各样地间, HR的土壤CH4累积碳排放量最高, 达到91.68 g CH4-C/m2, 显著高于其它区(P < 0.05);DP排放量最低, 其次为LR, H-LR和NP间CH4-C累积排放量无显著差异。

|
| 图 4 研究区土壤CH4排放速率季节变化及CH4-C排放量变化 Fig. 4 Seasonal variation in soil CH4 emission and changes in soil CH4-C emissions |
研究区除HR外, 土壤CO2排放速率与水位显著负相关(P < 0.05)(表 2)。表明随水位的升高, 土壤CO2排放速率显著降低。HR的水位波动对CO2排放速率没有显著影响(P>0.05)。LR水位与CH4排放速率显著正相关(P < 0.05), 在其它区不显著(P>0.05)。近地表温度与土壤CO2排放速率呈显著的指数回归关系(表 3), 其中HR的相关性最高(P < 0.01)。LR的温度敏感性Q10为4.62, 高于其它样地(2.46—2.63)。H-LR中温度与CH4排放速率呈显著的指数相关(P < 0.05), 在其它区不显著(P>0.05)(表 3)。
| 样地 Samples |
CO2 | CH4 | |||||||
| 拟合方程 Fitted equation |
R2 | F | P | 拟合方程 Fitted equation |
R2 | F | P | ||
| NP天然区 | R=-31.11W+423.18 | 0.49 | 6.85 | 0.03 | R=0.14W+6.34 | 0.04 | 0.30 | 0.60 | |
| DP退化区 | R=-68.17W+362.97 | 0.66 | 13.72 | <0.01 | R=0.0004W+0.45 | <0.01 | <0.01 | 0.99 | |
| LR低水位区 | R=-55.33W+289.03 | 0.70 | 16.47 | <0.01 | R=-0.35W+0.75 | 0.58 | 9.80 | 0.02 | |
| H-LR高低水位交替区 | R=-34.07W+383.67 | 0.56 | 8.82 | 0.02 | R=0.03W+7.12 | <0.01 | 0.02 | 0.90 | |
| HR高水位区 | R=-4.733W+206.69 | <0.01 | 0.02 | 0.89 | R=14.11W-70.62 | 0.12 | 0.98 | 0.36 | |
| R: CO2、CH4排放速率; W: 水位 | |||||||||
| 样地 Samples |
CO2 | CH4 | ||||||||
| 拟合方程 Fitted equation |
Q10 | R2 | F | P | 拟合方程 Fitted equation |
R2 | F | P | ||
| NP天然区 | R=44.35e0.090T | 2.46 | 0.55 | 8.50 | 0.02 | R=7.29e-0.006T | 0.002 | 0.02 | 0.90 | |
| DP退化区 | R=43.33e0.099T | 2.69 | 0.50 | 7.00 | 0.03 | R=0.13e0.048T | 0.10 | 0.77 | 0.41 | |
| LR低水位区 | R=6.82e0.153T | 4.62 | 0.60 | 10.4 | 0.01 | R=0.34e0.053T | 0.03 | 0.22 | 0.65 | |
| H-LR高低水位交替区 | R=0.07e0.306T | - | 0.51 | 7.36 | 0.03 | R=0.14e0.147T | 0.56 | 8.78 | 0.02 | |
| HR高水位区 | R=15.71e0.093T | 2.53 | 0.71 | 17.5 | <0.01 | R=4.14e0.084T | 0.15 | 1.23 | 0.30 | |
| R: CO2、CH4排放速率; T: 温度; Q10: 温度敏感性; 高低水位交替恢复区因水位管理, CO2排放速率波动较大, 未能计算Q10值 | ||||||||||
本研究中, 土壤CO2累积碳排放量与TOC、WL显著负相关, 与DOC显著正相关(P < 0.05);土壤CH4累积碳排放量与SWC、TOC和WL显著正相关。土壤CO2、CH4累积碳排放量受pH、TN、TP影响不显著(图 5)。

|
| 图 5 研究区土壤CO2累积碳排放量、CH4累积碳排放量与土壤理化指标、水位相关分析 Fig. 5 Relationships between soil physicochemical properties, water level and CO2-C, CH4-Cemission CO2-C:CO2累积碳排放量;CH4-C:CH4累积碳排放量;WL:水位Water level;SWC:土壤含水量Soil water content;pH:酸碱度;TP:总磷Total phosphorus;TOC:总有机碳Total organic carbon;TN:总氮Total nitrogen;DOC:可溶性有机碳Dissolved organic carbon |
生长季, 研究区各样地累积碳排放量差异显著(P < 0.05), DP的累积碳排放量最高, 达到6.77 t/hm2, HR的累积碳排放量最低, LR与H-LR排放量接近, 低于NP(表 4)。在增温潜势方面, 由于HR具有较高的CH4增温潜势, 其综合增温潜势显著高于其它样地;H-LR、DP与NP增温潜势接近, LR增温潜势最低。
| 样地 Samples |
累积碳排放量/(t/hm2) Cumulative carbon emissions |
综合增温潜势Warming potential(t/hm2) | ||
| GWP(CO2) | GWP(CH4) | ΣGWP | ||
| NP天然区 | 5.55±0.48ab | 19.70±1.86ab | 5.82±0.95b | 25.52±0.91b |
| DP退化区 | 6.77±0.43a | 24.75±1.58a | 0.47±0.13d | 25.22±1.49b |
| LR低水位区 | 4.54±0.37b | 16.47±1.37b | 1.75±0.24c | 18.22±1.24c |
| H-LR高低水位交替区 | 4.54±0.15b | 15.86±0.63b | 7.26±1.04b | 23.12±0.67b |
| HR高水位区 | 3.00±0.25c | 7.63±0.44c | 30.56±4.61a | 38.19±5.00a |
| GWP:全球增温潜势Global warming potential;同列不同小写字母表示同一指标在不同样地间差异显著(P<0.05) | ||||
本研究中, NP、DP、LR与HR的土壤CO2排放速率随季节变化呈现单峰型, 峰值出现在7—8月, 且CO2排放速率与近地表温度显著相关(表 3), 这与他人研究结果相似[25—26]。这主要是因为温度升高能促进微生物新陈代谢, 加速土壤不稳定底物分解;而另一方面, 7、8月份植物生长旺盛, 根系自养呼吸作用强烈[27], 也可导致土壤CO2排放速率升高。H-LR在6—8月份CO2排放速率波动剧烈, 大致表现为随水位增高CO2排放速率降低, 这是由于较高水位对土壤微生物活性产生了抑制作用[28]。在生长季末期, 温度降低, 植物、微生物生理代谢减慢, 导致研究区各样地CO2排放速率降低且趋于平稳。
水位可以通过改变土壤氧化还原电位、透气性、pH、微生物活性以及土壤中温室气体向大气扩散速率来影响温室气体的产生与排放[29]。本研究中, 除HR外, 其它样地的土壤CO2排放速率都与水位有显著的负相关关系(表 2), 这与其它研究的结论较一致[30—31]。较低的地下水位增加土壤好氧层厚度, 促进植物根系及微生物好氧呼吸, 促进土壤CO2的释放。HR一直处于淹水环境下, 土壤中氧气含量处于较低水平, 土壤微生物活性受到抑制, 土壤CO2排放速率较低;且HR水位波动小, 因此CO2排放速率与水位相关性不显著(P>0.05)。
温度敏感性Q10指温度每升高10 ℃土壤有机碳分解速率所增加的倍数, 可以表征土壤呼吸对温度变化的响应[32]。本研究中, 退化和恢复泥炭地Q10均高于天然泥炭地。表明泥炭地受破坏后, 土壤CO2排放速率对于温度变化更为敏感。马维伟等[33]的研究也发现重度退化泥炭地Q10值大于未退化泥炭地。这可能是因为泥炭地退化后, 土壤中易分解的有机碳被快速分解, 而剩余有机碳中顽固有机碳(如木质素类化合物)相对较多。根据酶动力学理论, 低质量有机质具有更高的活化能, 应当具有更高的温度敏感性。在恢复区中, HR的Q10最低, LR的Q10最高。这是由于HR较高水位抑制了土壤呼吸, 降低了其温度的敏感性[34];LR较低的水位使得土壤通气量增加, 协同加快土壤有机碳的矿化速率。而在一定的温度范围内, 土壤有机碳的矿化速率随温度升高而增加, 土壤呼吸强度与温度成正比[26], 从而导致LR的温度敏感性更高。
研究区土壤CO2累积碳排放量与WL、SOC含量呈显著的负相关关系, 与DOC呈显著正相关(P < 0.05)(图 5), 说明水位和底物的有效性是调控研究区不同样地土壤CO2累积碳排放量差异的关键因素。Evans等[35]通过对比全球65个不同地点的气体通量数据, 指出水位是引起不同地点CO2净生态系统交换差异的唯一因素, 而在水位变化较小的生态系统中, 植被、养分、pH、微地形等其它因素对碳循环影响较大。较多的研究[36]表明, 土壤SOC含量与土壤CO2排放呈正相关关系。主要是由于土壤有机碳库是土壤微生物分解的主要碳源, 其含量高低直接影响着土壤微生物的活性, 从而影响了土壤的碳排放过程[37]。但在本研究中较高的水位抑制了微生物的活性, 降低了微生物对有机质的分解速率, 促进了有机质的累积, 因此水位和SOC都表现出与土壤CO2-C负相关。土壤中DOC的浓度是土壤有机碳分解产生DOC和土壤微生物分解消耗DOC之间的平衡。DOC作为微生物异养呼吸的底物, 其较高的浓度可促进微生物的呼吸作用[38—39]。本研究中水位较低的NP、DP和LR样地均具有相对较高的DOC浓度(表 1), 这是由于在低水位条件下, 生物代谢加强, 很多高分子化合物被分解, 产生的DOC更多地被分解以CO2的形式释放[40—41], 而该过程对甲烷释放影响较弱[42]。因此本研究中, DOC浓度与CO2释放显著正相关, 但与CH4排放未有显著相关性(图 5)。
3.2 土壤CH4排放特征及其影响因素本研究中, H-LR的土壤CH4排放速率呈现出单峰型季节变化, 排放峰值出现在8月中旬, 与温度的季节变化一致。其主要原因是夏季温度较高, 产CH4菌活性增强, 促进CH4释放[42];而NP、LR与DP的土壤CH4排放速率无明显的季节变化模式, 仅在H-LR发现土壤CH4排放速率与温度显著相关(P < 0.05)。在已有的研究中, 温度与CH4排放速率既存在显著相关也存在不显著相关[26, 43]。土壤CH4排放是CH4产生、氧化与传输综合作用的结果, 温度不仅影响CH4产生, 而且影响其氧化过程[25, 44]。NP、LR、DP平均水位低于地表, 土壤上部有较厚的氧化层, 因此甲烷氧化菌生长活动旺盛。较高温度有利于甲烷氧化菌对CH4氧化, 导致CH4排放速率较低[45]。HR的土壤CH4排放速率呈现出双峰型模式, 陈槐等[46]在若尔盖高原湿地木里苔草群落的研究中也发现类似的现象。这可能是因为长期积水条件下产甲烷菌活性较强, 在6月下旬苔草分蘖盛期, 根系分泌物增加[47];而在8月下旬, 植物凋落物逐渐输入, 根系分泌物和凋落物为产CH4菌提供大量的不稳定底物[44], 促进土壤CH4的再次大量排放。
尽管较高地下水位营造的厌氧环境是土壤CH4大量产生的前提, 然而在野外环境下, 地下水位变化对土壤CH4排放的影响是复杂因素相互作用的结果[48]。本研究中, NP、DP、H-LR与HR的土壤CH4排放速率与水位没有显著相关性(P>0.05), 周文昌[49]在若尔盖高原自然和排水泥炭地中也得出类似结论。这是因为水位对土壤CH4排放速率的影响存在滞后效应, 土壤好氧层因水位抬升被淹没后, 土壤CH4排放不会立刻增加, 直到土壤中的氧分子耗尽[50]。已有研究[51]也发现在地下水位相对较高(接近或超过地表 10 cm)的湿地中, 水位的正常季节波动对土壤CH4排放速率影响较小, 这与本研究中水位对HR的CH4排放速率影响一致。值得注意的是, 在LR观测到土壤CH4排放速率与水位有显著的负相关关系(P < 0.01)。这是因为地下水位下降后, 压力梯度降低、温度升高导致的CH4释放作用大于水位升高限制微生物生产CH4、氧化CH4的作用[52]。
本研究中, 各样地土壤CH4累积碳排放量差异十分显著(图 4), 表现为HR>H-LR=NP>LR>DP。相关分析表明, SWC、WL与CH4累积碳排放量有显著的正相关关系(P < 0.05)(图 5)。较高的土壤含水量和水位为CH4产生提供有利的厌氧环境, 有利于产CH4菌释放CH4并抑制CH4氧化菌消耗CH4, 从而促进CH4释放[39]。此外, 土壤CH4累积碳排放量与TOC显著正相关(P < 0.01)(图 5)。高水位环境促进了有机质的累积, 较高的SOC含量为产甲烷菌提供充足底物, 促进CH4排放。
本研究中, 土壤CO2和CH4的累积碳排放量与土壤pH、TP及TN均不具有显著的相关关系, 表明pH、TP、TN不是研究区土壤CO2和CH4的累积碳排放量产生差异的主要原因。这可能是由于恢复泥炭地进行差异性水位管理的时间较短, 使得不同水位管理恢复区土壤pH、TP及TN差异并不显著(表 1)。因此, 上述因子并未对研究区土壤CO2和CH4的累积排放量产生主要影响。
3.3 含碳温室气体排放量以及全球增温潜势通过对研究区土壤含碳温室气体的累积碳排放量及综合增温潜势(100 a尺度)的分析发现, DP的累积碳排放量显著高于NP和恢复泥炭地(LR、HR、H-LR), 增温潜势也高于恢复区。这表明人为排水导致更多温室气体排放, 加剧了全球增温的潜在风险。而对于被农业开垦的退化泥炭地, 植被和水文条件的恢复可有效地控制土壤温室气体排放带来的增温效应。恢复泥炭地中, HR的碳累积排放量低于LR和H-LR(表 4), 表明对恢复泥炭地进行持续的淹水可降低总碳的输出量。但值得注意的是, HR的综合增温潜势显著高于其他水位恢复区, 甚至高于退化和天然泥炭地(表 4), 这主要是由于其较高的CH4增温潜势造成的。而LR与HR的累积碳排放量相差不多, 均显著低于DP, LR增温潜势最低(表 4)。此外, 研究区的恢复泥炭地处于恢复初期阶段, 该阶段移栽的优势植物尚处于快速扩繁建群阶段。已有的研究表明, 相较于积水环境, 低水位更能促进苔草地下根系的繁殖, 促进其地下生物量的累积[53—54]。因此, 建议在泥炭地恢复初期将低水位管理作为短期策略, 以更好地恢复泥炭地碳汇功能、减弱其增温潜势。
4 结论(1) 不同水位管理下恢复泥炭地土壤CO2、CH4排放差异显著。H-LR受水位管理的影响, CO2排放速率波动较大, LR、HR土壤CO2排放速率随季节变化呈现单峰型;HR的土壤CH4排放速率最高, 呈双峰模式;LR与H-LR土壤CH4排放无明显的时间变化特征。温度和水位变化是研究区泥炭地土壤CO2、CH4排放季节变化的主控因子。
(2) 通过对研究区土壤含碳温室气体的累积排放量及综合增温潜势(100 a尺度)的分析发现, 退化泥炭地的累积碳排放量显著高于天然和恢复泥炭地(LR、HR、H-LR), 表明泥炭地开垦导致更多温室气体排放, 而对于退化泥炭地进行植被和水文条件恢复可有效地控制土壤温室气体排放带来的增温效应。恢复泥炭地中, 高水位管理恢复泥炭地的累积碳排放量显著低于低水位和高-低水位交替恢复泥炭地, 但其综合增温潜势显著高于其他水位恢复区。低水位恢复泥炭地的累积碳排放量显著低于退化泥炭地, 且其增温潜势最低。因此, 本文建议在泥炭地恢复初期将低水位管理作为短期策略, 以更好地恢复泥炭地碳汇功能, 减弱其增温潜势。
致谢: 感谢实验室周新华、刘莎莎、刘自平老师对实验的耐心帮助。| [1] |
Xu J R, Morris P J, Liu J G, Holden J. PEATMAP: Refining estimates of global peatland distribution based on a meta-analysis. CATENA, 2018, 160: 134-140. DOI:10.1016/j.catena.2017.09.010 |
| [2] |
Yu Z C, Loisel J, Brosseau D P, Beilman D W, Hunt S J. Global peatland dynamics since the Last Glacial Maximum. Geophysical Research Letters, 2010, 37(13): L13402. |
| [3] |
张乐. 长白山地区泥炭沼泽湿地碳累积及其地下部温室气体排放影响机制研究[D]. 南京: 南京信息工程大学, 2021.
|
| [4] |
Li C J, Grayson R, Holden J, Li P F. Erosion in peatlands: Recent research progress and future directions. Earth-Science Reviews, 2018, 185: 870-886. DOI:10.1016/j.earscirev.2018.08.005 |
| [5] |
Frolking S, Talbot J, Jones M C, Treat C C, Kauffman J B, Tuittila E S, Roulet N. Peatlands in the Earth's 21st century climate system. Environmental Reviews, 2011, 19: 371-396. DOI:10.1139/a11-014 |
| [6] |
Maljanen M, Komulainen V M, Hytönen J, Martikainen P J, Laine J. Carbon dioxide, nitrous oxide and methane dynamics in boreal organic agricultural soils with different soil characteristics. Soil Biology and Biochemistry, 2004, 36(11): 1801-1808. DOI:10.1016/j.soilbio.2004.05.003 |
| [7] |
Moore T R, Roulet N T, Waddington J M. Uncertainty in predicting the effect of climatic change on the carbon cycling of Canadian peatlands. Climatic Change, 1998, 40(2): 229-245. DOI:10.1023/A:1005408719297 |
| [8] |
Bonn A, Allott T, Evans M, Joosten H, Stoneman R. Peatland Restoration and Ecosystem Services: Science, Policy and Practice. Cambridge: Cambridge University Press, 2016: 339-357.
|
| [9] |
Parry L E, Holden J, Chapman P J. Restoration of blanket peatlands. Journal of Environmental Management, 2014, 133: 193-205. DOI:10.1016/j.jenvman.2013.11.033 |
| [10] |
Fennessy M S, Mitsch W J. Effects of hydrology on spatial patterns of soil development in created riparian wetlands. Wetlands Ecology and Management, 2001, 9(2): 103-120. DOI:10.1023/A:1011104902410 |
| [11] |
孟宪民. 湿地与全球环境变化. 地理科学, 1999, 19(5): 385-391. |
| [12] |
Planas-Clarke A M, Chimner R A, Hribljan J A, Lilleskov E A, Fuentealba B. The effect of water table levels and short-term ditch restoration on mountain peatland carbon cycling in the Cordillera Blanca, Peru. Wetlands Ecology and Management, 2020, 28(1): 51-69. DOI:10.1007/s11273-019-09694-z |
| [13] |
赵婧, 段磊磊, 王铭, 王升忠, 陈聪, 李鸿凯, 李兴丽. 水文管理措施对长白山区恢复泥炭地土壤酶活性的影响. 生态学杂志, 2022, 41(5): 948-954. |
| [14] |
Kokkonen N A K, Laine A M, Laine J, Vasander H, Kurki K, Gong J N, Tuittila E S. Responses of peatland vegetation to 15-year water level drawdown as mediated by fertility level. Journal of Vegetation Science, 2019, 30(6): 1206-1216. DOI:10.1111/jvs.12794 |
| [15] |
Henneberg A, Brix H, Sorrell B K. The interactive effect of Juncus effusus and water table position on mesocosm methanogenesis and methane emissions. Plant Soil, 2016, 400(1): 45-54. |
| [16] |
Jerman V, Danevčič T, Mandic-Mulec I. Methane cycling in a drained wetland soil profile. Journal of Soils and Sediments, 2017, 17(7): 1874-1882. DOI:10.1007/s11368-016-1648-2 |
| [17] |
Rask H, Schoenau J, Anderson D. Factors influencing methane flux from a boreal forest wetland in Saskatchewan, Canada. Soil Biology and Biochemistry, 2002, 34(4): 435-443. DOI:10.1016/S0038-0717(01)00197-3 |
| [18] |
Kannenberg S A, Dunn S T, Ludwig S M, Spawn S A, Schade J D. Patterns of potential methanogenesis along soil moisture gradients following drying and rewetting in Midwestern prairie pothole wetlands. Wetlands, 2015, 35(4): 633-640. DOI:10.1007/s13157-015-0653-3 |
| [19] |
Cooper M D A, Evans C D, Zielinski P, Levy P E, Gray A, Peacock M, Norris D, Fenner N, Freeman C. Infilled ditches are hotspots of landscape methane flux following peatland re-wetting. Ecosystems, 2014, 17(7): 1227-1241. DOI:10.1007/s10021-014-9791-3 |
| [20] |
Green S M, Baird A J, Evans C D, Peacock M, Holden J, Chapman P J, Smart R P. Methane and carbon dioxide fluxes from open and blocked ditches in a blanket bog. Plant and Soil, 2018, 424(1/2): 619-638. |
| [21] |
马学慧. 中国泥炭地碳储量与碳排放. 北京: 中国林业出版社, 2013: 48.
|
| [22] |
李英俊. 吉林省泥炭资源现状及保护管理对策. 湿地科学, 2013, 11(3): 404-406. |
| [23] |
谢立勇, 许婧, 郭李萍, 徐玉秀, 孙雪, 赵洪亮, 郭飞, 赵迅. 水肥管理对稻田CH4排放及其全球增温潜势影响的评估. 中国生态农业学报, 2017, 25(7): 958-967. |
| [24] |
Batson J, Noe G B, Hupp C R, Krauss K W, Rybicki N B, Schenk E R. Soil greenhouse gas emissions and carbon budgeting in a short-hydroperiod floodplain wetland. Journal of Geophysical Research: Biogeosciences, 2015, 120(1): 77-95. DOI:10.1002/2014JG002817 |
| [25] |
Li Y Y, Dong S K, Liu S L, Zhou H K, Gao Q Z, Cao G M, Wang X X, Su X K, Zhang Y, Tang L, Zhao H D, Wu X Y. Seasonal changes of CO2, CH4 and N2O fluxes in different types of alpine grassland in the Qinghai-Tibetan Plateau of China. Soil Biology and Biochemistry, 2015, 80: 306-314. DOI:10.1016/j.soilbio.2014.10.026 |
| [26] |
沙晨燕. 不同类型河滨湿地甲烷和二氧化碳排放的研究[D]. 上海: 华东师范大学, 2010.
|
| [27] |
Song Y Y, Song C C, Hou A X, Ren J S, Wang X W, Cui Q, Wang M Q. Effects of temperature and root additions on soil carbon and nitrogen mineralization in a predominantly permafrost peatland. CATENA, 2018, 165: 381-389. DOI:10.1016/j.catena.2018.02.026 |
| [28] |
Musarika S, Atherton C E, Gomersall T, Wells M J, Kaduk J, Cumming A M J, Page S E, Oechel W C, Zona D. Effect of water table management and elevated CO2 on radish productivity and on CH4 and CO2 fluxes from peatlands converted to agriculture. Science of the Total Environment, 2017, 584-585: 665-672. DOI:10.1016/j.scitotenv.2017.01.094 |
| [29] |
李海防, 夏汉平, 熊艳梅, 张杏锋. 土壤温室气体产生与排放影响因素研究进展. 生态环境, 2007, 16(6): 1781-1788. |
| [30] |
Hirano T, Kusin K, Limin S, Osaki M. Carbon dioxide emissions through oxidative peat decomposition on a burnt tropical peatland. Global Change Biology, 2014, 20(2): 555-565. DOI:10.1111/gcb.12296 |
| [31] |
Silvola J, Alm J, Ahlholm U, Nykanen H, Martikainen P J. CO2 fluxes from peat in boreal mires under varying temperature and moisture conditions. The Journal of Ecology, 1996, 84(2): 219-228. DOI:10.2307/2261357 |
| [32] |
朱珠, 徐侠, 杨赛兰, 彭凡茜, 张惠光, 蔡斌. 陆地生态系统土壤有机碳分解温度敏感性研究进展. 南京林业大学学报: 自然科学版, 2022, 46(1): 33-39. |
| [33] |
马维伟, 孔同伟, 宋元君, 朱正清, 梁鹏飞. 尕海湿地植被退化过程中土壤有机碳矿化特征. 水土保持学报, 2020, 34(3): 342-348. |
| [34] |
Conant R T, Dalla-Betta P, Klopatek C C, Klopatek J M. Controls on soil respiration in semiarid soils. Soil Biology and Biochemistry, 2004, 36(6): 945-951. DOI:10.1016/j.soilbio.2004.02.013 |
| [35] |
Evans C D, Peacock M, Baird A J, Artz R R E, Burden A, Callaghan N, Chapman P J, Cooper H M, Coyle M, Craig E, Cumming A, Dixon S, Gauci V, Grayson R P, Helfter C, Heppell C M, Holden J, Jones D J, Kaduk J, Levy P, Matthews R, McNamara N P, Misselbrook T, Oakley S, Page S E, Rayment M, Ridley L M, Stanley K M, Williamson J L, Worrall F, Morrison R. Overriding water table control on managed peatland greenhouse gas emissions. Nature, 2021, 593(7860): 548-552. DOI:10.1038/s41586-021-03523-1 |
| [36] |
刘意立. 我国亚热带季风气候区湿地土壤CO2、CH4排放规律研究[D]. 杭州: 浙江大学, 2014.
|
| [37] |
Wagner D, Pfeiffer E M. Two temperature optima of methane production in a typical soil of the Elbe river marshland. FEMS Microbiology Ecology, 1997, 22(2): 145-153. DOI:10.1111/j.1574-6941.1997.tb00366.x |
| [38] |
Strack M, Waddington J M. Response of peatland carbon dioxide and methane fluxes to a water table drawdown experiment. Global Biogeochemical Cycles, 2007, 21(1): GB1007. |
| [39] |
Treat C C, Wollheim W M, Varner R K, Grandy A S, Talbot J, Frolking S. Temperature and peat type control CO2 and CH4 production in Alaskan permafrost peats. Global Change Biology, 2014, 20(8): 2674-2686. DOI:10.1111/gcb.12572 |
| [40] |
Van Den Berg L J L, Shotbolt L, Ashmore M R. Dissolved organic carbon (DOC) concentrations in UK soils and the influence of soil, vegetation type and seasonality. Science of the Total Environment, 2012, 427-428: 269-276. DOI:10.1016/j.scitotenv.2012.03.069 |
| [41] |
Wang J Y, Song C C, Zhang J, Wang L L, Zhu X Y, Shi F X. Temperature sensitivity of soil carbon mineralization and nitrous oxide emission in different ecosystems along a mountain wetland-forest ecotone in the continuous permafrost of Northeast China. CATENA, 2014, 121: 110-118. DOI:10.1016/j.catena.2014.05.007 |
| [42] |
刘亮锋, 陈槐, 彭长辉, 朱求安, 李宝鑫. 若尔盖高原不同深度泥炭在增温条件下CH4释放. 应用与环境生物学报, 2016, 22(1): 1-7. |
| [43] |
Zhu X Y, Song C C, Guo Y D, Sun X X, Zhang X H, Miao Y Q. Methane emissions from temperate herbaceous peatland in the Sanjiang Plain of Northeast China. Atmospheric Environment, 2014, 92: 478-483. DOI:10.1016/j.atmosenv.2014.04.061 |
| [44] |
Lai D Y F. Methane dynamics in northern peatlands: a review. Pedosphere, 2009, 19(4): 409-421. DOI:10.1016/S1002-0160(09)00003-4 |
| [45] |
Berglund Ö, Kätterer T, Meurer K H E. Emissions of CO2, N2O and CH4 from cultivated and set aside drained peatland in central Sweden. Frontiers in Environmental Science, 2021, 9: 630721. DOI:10.3389/fenvs.2021.630721 |
| [46] |
陈槐, 高永恒, 姚守平, 吴宁, 王艳芬, 罗鹏, 田建卿. 若尔盖高原湿地甲烷排放的时空异质性. 生态学报, 2008, 28(7): 3425-3437. |
| [47] |
龚振平, 颜双双, 闫超, 王家睿, 战厚强. 寒地水稻秸秆还田和温度对稻田甲烷排放的影响. 东北农业大学学报, 2015, 46(12): 8-15. |
| [48] |
Long K D, Flanagan L B, Cai T B. Diurnal and seasonal variation in methane emissions in a northern Canadian peatland measured by eddy covariance. Global Change Biology, 2010, 16(9): 2420-2435. |
| [49] |
周文昌. 人类活动对若尔盖高原泥炭地碳通量和碳储量的影响[D]. 北京: 中国林业科学研究院, 2015.
|
| [50] |
Suyker A E, Verma S B, Clement R J, Billesbach D P. Methane flux in a boreal fen: Season-long measurement by eddy correlation. Journal of Geophysical Research: Atmospheres, 1996, 101(D22): 28637-28647. |
| [51] |
Christensen T R, Ekberg A, Ström L, Mastepanov M, Panikov N, Öquist M, Svensson B H, Nykänen H, Martikainen P J, Oskarsson H. Factors controlling large scale variations in methane emissions from wetlands. Geophysical Research Letters, 2003, 30(7): 1414. |
| [52] |
Treat C C, Bubier J L, Varner R K, Crill P M. Timescale dependence of environmental and plant-mediated controls on CH4 flux in a temperate fen. Journal of Geophysical Research: Biogeosciences, 2007, 112(G1): G01014. |
| [53] |
王秋林, 陈静蕊, 程平生. 湿地植物灰化苔草对淹水的生态响应. 水生态学杂志, 2017, 38(1): 24-29. |
| [54] |
李亚芳, 陈心胜, 项文化, 谢永宏. 不同高程短尖苔草对水位变化的生长及繁殖响应. 生态学报, 2016, 36(7): 1959-1966. |