 2023, Vol. 43
2023, Vol. 43文章信息
- 肖倩茹, 江蓝, 贾梅花, 何中声, 陈博, 刘金福, 朱德煌, 陈文伟, 李文周
- XIAO Qianru, JIANG Lan, JIA Meihua, HE Zhongsheng, CHEN Bo, LIU Jinfu, ZHU Dehuang, CHEN Wenwei, LI Wenzhou
- 戴云山黄山松群落生态(火用)空间异质性
- Eco-exergy spatial heterogeneity of Pinus taiwanensis community in Daiyun Mountain
- 生态学报. 2023, 43(11): 4689-4699
- Acta Ecologica Sinica. 2023, 43(11): 4689-4699
- http://dx.doi.org/10.5846/stxb202107262026
-
文章历史
- 收稿日期: 2021-07-26
- 网络出版日期: 2023-02-07

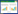




2. 福建农林大学海峡自然保护区研究中心, 福州 350002;
3. 生态与资源统计福建省高校重点实验室, 福州 350002;
4. 武夷学院, 武夷山 354300;
5. 戴云山国家级自然保护区管理局, 泉州 362500
2. Cross-Strait Nature Reserve Research Center, Fujian Agriculture and Forestry University, Fuzhou 350002, China;
3. Key Laboratory of Fujian Universities for Ecology and Resource Statistics, Fuzhou 350002, China;
4. Wuyi University, Wuyi Mountain 354300, China;
5. Daiyun Mountain National Nature Reserve, Quanzhou 362500, China
森林生态系统为开放性系统[1], 是由森林群落能量流与环境相互作用形成的具有一定自组织、结构和功能的复杂网络[2], 任何生态系统过程均伴随着能量转换, 并遵循热力学定律[2—3]。因而, 热力学原理可能影响森林生态系统结构和功能的发展[3]。将热力学指标应用于森林生态系统, 可反映森林群落与环境之间物质能量的转化传递, 体现系统结构组成与功能状态[4]。森林生态系统具有一定空间结构, 表现出明显的时空特征[1]。运用热力学指标研究森林生态系统空间结构, 能以整体方式反映群落生物热力学空间结构, 为准确量化生态系统状况提供了新思路[5]。
热力学指标生态火用(Ex)为特定生物体或生态系统偏离热力学平衡状态的距离[2], 通过计算物种包含的遗传信息和生物量得到, 可在时间尺度上度量生态系统的发展状况, 反映系统的结构和稳定性[6—7]。单位生物量的生态火用值为结构火用(SpEx), 是对单位生物量所包含物种遗传信息的度量[8]。由于高等生物每单位生物量中携带了更多的遗传信息(单倍体基因组DNA的物理总长度C-value更大), 因而结构火用值可反映物种组成的优势, 其大小主要取决于物种组成和群落性质。相较于传统群落结构研究, 热力学指标生态火用和结构火用结合了物种生物量, 考虑了不同物种的进化历史和遗传信息, 对群落结构变化敏感, 能全面量化群落结构[3, 6, 9], 衡量生态系统的稳定性[10], 并且计算方便、可行性强, 有助于丰富和完善经典群落结构指数[11]。生态火用理论最初应用于水生生态与湿地生态系统健康评价研究, 而森林生态系统的生态火用研究较少, 主要集中在自组织、健康水平和生态恢复评估等[7, 12—14]。生态火用和结构火用相结合能更好地描述森林生态系统健康状况, 准确客观的评估生态系统现状[9—10]。
森林生态系统中, 由于随机性和结构性因子在时空上的综合作用, 影响了群落物种组成与结构, 群落生态火用在空间分布上也存在异质性, 为区域化变量[15—16]。空间异质性研究可用于反映区域化变量生态学过程和格局的变异性和复杂性[17]。目前群落空间异质性研究多集中于林木个体、灌木层植物多样性和土壤理化性质[18], 综合群落生态火用与空间异质性理论可以分析不同尺度的生态火用空间异质性程度以及变异格局, 对拓展生态火用理论的应用领域具有重要意义。
戴云山自然保护区为研究中亚热带地区植被演替和生物多样性保护提供良好平台, 主要保护对象是我国东南沿海保存最完好及面积最大的原生性黄山松(Pinus taiwanensis)林[19—20]。黄山松为我国特有种, 作为中山地区植被恢复更新的优良造林树种, 具有马尾松(P. massoniana)无法达到的生态演替功能[21]。研究表明戴云山黄山松在群落中占据优势地位[22], 目前黄山松群落研究主要集中在物种多样性[23]、种群生态位[22]、空间异质性和植被维持机制[17]等方面。苏松锦[17]对戴云山黄山松群落空间结构进行了研究, 但未考虑群落中不同物种的进化情况和遗传信息, 而将生态火用理论应用于黄山松群落空间结构研究, 可有效衡量黄山松群落整体的稳定性和发展状况[24]。为此, 以黄山松集中分布的戴云山国家级自然保护区的陈板岭头1 hm2样地为研究对象, 主要研究:(1)确定黄山松在戴云山植物群落中的生物热力学地位, 阐明黄山松种群在维系群落结构稳定中发挥的作用;(2)通过研究不同尺度生态火用和结构火用的空间异质性程度及其变异格局, 揭示戴云山黄山松群落生物热力学空间结构。
1 研究区自然概况戴云山国家级自然保护区位于福建省泉州市德化县境内(北纬25°38′07″—25°43′40″, 东经118°05′22″—118°20′15″), 总面积13472.4 hm2, 其森林覆盖度达93.4%, 区内拥有6400 hm2的原生性黄山松森林群落[19, 25]。保护区地处中亚热带和南亚热带过渡区域, 属于典型海洋季风气候区, 年平均气温15.6—19.5℃, 年均日照1875.4 h, 年平均雾日220 d, 无霜期260 d, 年均降水量在1700—2000 mm, 年相对湿度高达80%以上。土壤呈酸性, 主要有5亚类:山地红壤、山地黄壤、赤红壤、山地黄红壤和泥炭沼泽土[26—27]。研究样地位于保护区内陈板岭头(25°40′14″N、118°13′9″E), 冠层郁闭度93.4%, 平均海拔1415 m, 坡度5—20°[28]。结合研究区地形图与林业基本图, 通过实地踏查, 在地形相对平缓且黄山松集中分布区域, 设置一块面积为1 hm2(100 m×100 m)样地(图 1)。

|
| 图 1 研究区域和采样点的位置 Fig. 1 Location of the study area and the sampling points |
采用规则网格法, 将样地分为100个10 m×10 m的小样方。乔木层起测树高为5 m, 灌木层为样地内高度<5 m的木本植物, 包括乔木的幼树和灌木, 记录样方中树种胸径、树高、基径、冠幅等[29]。研究区群落层次明显, 乔木层主要有黄山松、江南山柳(Clethra cavaleriei)、鹿角杜鹃(Rhododendron latoucheae)、红楠(Machilus thunbergiisieb)和大萼两广黄瑞木(Adinandra glischroloma);灌木层主要有岗柃(Eurya groffii)、中华野海棠(Bredia sinensis), 映山红(R. pulchrum)和短尾越桔(Vaccinium carlesii)等[30—31]。
3 数据处理与分析结合群落物种多度、生物量和重要值, 分析戴云山黄山松群落物种组成。为了确定黄山松在群落中的生物热力学地位以及在维系群落结构稳定中发挥的作用, 计算群落中各物种在不同冠层中的生态火用值和结构火用值, 以及各物种在不同冠层中的生态火用贡献率。采用GS+10.0进行半方差函数计算, 绘制半方差函数图, 拟合半方差函数模型, 描述不同尺度的生态火用和结构火用在空间上的变异程度[32]。最后, 依据半方差函数拟合的最佳模型, 使用ArcGIS 10.1中克里金插值进行空间插值分析, 揭示戴云山黄山松群落不同尺度的生态火用和结构火用值在空间上的变异格局。
3.1 生物量和重要值计算黄山松最适生物量由模型M=0.092×(D2.09×H)[33]估算得到;木荷生物量为地上生物量模型M=0.12045×(D2.06446×H)0.38265和地下生物量模型M=0.081769×D2.32395×H-0.24289的总和[34];其他乔木树种生物量由混合物种生物量模型M=0.0772×(D2×H)0.9196[35]估算得到;其他灌木树种生物量由模型M=0.0042×H1.9323[36]估算得到。模型中M为生物量, D为胸径, H为树高。群落中树种优势度可用重要值来衡量[37]:
重要值=(相对多度+相对频度+相对显著度)/3
3.2 生态火用和结构火用的计算生态火用(Ex)和结构火用(SpEx)的计算公式为:
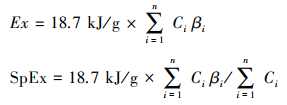
|
其中, 18.7 kJ/g为碎屑物所含生态火用的平均值, Ci代表i种物种的生物量(g/m2), βi代表i种物种相对于碎屑物的权重转换因子[12, 38]。
生态火用值与权重因子β密切相关。目前研究中, 植物和土壤真核生物生态火用常采用C-value法[3, 39]。本文主要研究对象为植物, 故采用C-value法计算β, 即使用种水平上的C-value作为参数推断β权重转换因子, C-value是单倍核中包含的DNA数量, 用碱基对数表示[40—41]。公式为:
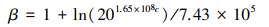
|
其中, C代表生物单倍体基因组DNA的物理总长度(C-value), 单位pg或10-12g。C-value在英国皇家植物园C-value库中查询[42](http://cvalues.science.kew.org)。为了尽可能精确β值, C值尽可能精确至种水平。若库中缺失相应种, 则取同属植物的C-value计算, 如不能精确至属, 则以同科植物C-value平均值计算[38, 43]。
变异系数CV为样本数据标准差与平均值的比值, 使用其来表示生态火用和结构火用的离散程度, CV≤0.1时为弱变异, 0.1<CV≤1时为中等变异, CV>1时为强变异, CV越大表明数据差异程度越大[44]。
3.3 群落生态火用贡献率与冠层生态火用贡献率的计算物种在群落中会占据一定资源空间, 具有特定群落地位。为了更好的从热力学角度反映物种在群落中的地位, 分别采用群落生态火用贡献率PC和冠层生态火用贡献率PL来量化物种在群落和冠层中的地位, 明确群落结构组成[38]。
|
式中n为群落包含的冠层数, 本文中冠层数为2, 即乔木层和灌木层。Exij为冠层i物种j的生态火用, mi为i冠层所含有的物种数。
3.4 空间异质性 3.4.1 半方差函数分析半方差函数是地统计学中研究空间变异性的工具函数, 常被用来表征变量的空间变异结构[32]。生态火用值为区域化变量, 具有随机性和结构性特征, 可采用半方差函数对生态火用的空间异质性尺度进行分析, 数学表达式为:
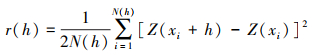
|
上述式中h为步长, Z(xi)和Z(xi+h)为变量Z在空间位置xi和xi+h上的观测值。其中i=1, 2, …, N(h);N(h)指步长为h时的样点对数。
为防止半方差函数因特异值发生畸变, 采用阈值法对数据进行预处理, 即将样本均值加减3倍标准差外的样点剔除, 并用均值代替[45]。考虑到约翰逊(Johnson)变换为双向对称变换, 对峰度的改变效果明显, 转换能力强, 在应用中整体效果较好, 成功率超过99.5%[46—47]。因而, 本文各层生态火用和结构火用值采用约翰逊(Johnson)变换, 转换后数据均符合正态分布。采用SPSS 22.0对转换后的数据进行单样本柯尔莫可洛夫-斯米洛夫(Kolmogorov-Smirnov)检验以判断数据是否服从正态分布。
通常用半方差函数相关参数来描述其空间分布结构, 其中块金值(Co)表示随机性因素引起的空间异质性;结构系数(C)表示由结构性因素引起的空间异质性[45];有效变程(Ao)表示可变的空间自相关范围, 即聚集格局的规模[48];基台值(Co+C)反映总的空间异质性程度;结构比C/(Co+C)的大小反映了空间相关性强度, 该值<25%、25%—75%和>75%时分别表示变量的空间相关性较弱、中等和较强[44]。半方差函数模型的最优选择根据决定系数(R2)及残差平方和(RSS)决定, R2越大RSS越小, 表明实测值与回归线越靠近, 拟合曲线效果越好[49]。
3.4.2 空间插值分析采用地统计学结合地理信息系统(GIS)技术进行生态火用和结构火用值插值以分析其空间异质性。由于数据数学期望未知, 样方为连续区域, 选用普通克里金模型得到戴云山黄山松林各样方生态火用值和结构火用值克里金等值线图[50]。插值预测结果评价标准为:均值误差(ME)趋近0, 均方根误差(RMSE)越小越好, 平均标准误差(ASE)趋近均方根误差, 标准化均方根误差(SRMSE)趋近1[16, 45]。
4 结果 4.1 群落概况研究区内黄山松群落共有110个树种, 黄山松在乔木层中多度最高(1616株), 重要值最高(32.11%), 在乔木层和灌木层中生物量均最大(40780.23 g/m2, 14951.26 g/m2)(表 1)。黄山松为乔木和灌木层的优势种。岗柃在灌木层中多度最高(7456株), 重要值9.29%, 灌木层生物量84.58 g/m2, 为灌木层优势种。
| 物种 Species |
多度Abundances | 重要值/% Important value |
生物量Biomass/(g/m2) | |||
| 群落 Community |
乔木层 Tree layer |
灌木层 Shrub layer |
乔木层 Tree layer |
灌木层 Shrub layer |
||
| 黄山松Pinus taiwanensis | 5607 | 1616 | 3991 | 32.11 | 40780.23 | 14951.26 |
| 岗柃Eurya groffii | 7460 | 4 | 7456 | 9.29 | 1.97 | 84.58 |
| 中华野海棠Bredia sinensis | 5612 | 0 | 5612 | 6.79 | 0.00 | 12.03 |
| 映山红Rhododendron pulchrum | 4331 | 0 | 4331 | 6.16 | 0.00 | 74.97 |
| 短尾越桔Vaccinium carlesii | 2470 | 9 | 2461 | 3.77 | 6.20 | 10.56 |
| 江南山柳Clethra cavaleriei | 1265 | 195 | 1070 | 3.75 | 1777.79 | 32.55 |
| 鹿角杜鹃R. latoucheae | 1424 | 54 | 1370 | 3.66 | 871.53 | 36.57 |
| 岩柃E. saxicola | 1564 | 0 | 1564 | 2.87 | 0.01 | 11.04 |
| 羊舌树Symplocos glauca | 475 | 18 | 457 | 1.92 | 339.15 | 100.11 |
| 窄基红褐柃E. rubiginosa var. attenuata | 692 | 0 | 692 | 1.73 | 0.00 | 8.64 |
| 红楠Machilus thunbergiisieb | 223 | 45 | 178 | 1.68 | 1285.53 | 4.61 |
| 大萼两广黄瑞木Adinandra glischroloma var. macrosepala | 319 | 29 | 290 | 1.58 | 341.51 | 9.42 |
| 华丽杜鹃R. eudoxum | 224 | 0 | 224 | 1.35 | 0.00 | 7.33 |
| 矩圆叶鼠刺Itea omeiensis | 412 | 1 | 411 | 1.03 | 3.25 | 5.33 |
| 满山红R. mariesii | 284 | 0 | 284 | 1.05 | 0.00 | 3.67 |
| 小叶赤楠Syzygium grijsii | 271 | 0 | 271 | 1.04 | 0.00 | 5.94 |
| 钝齿冬青Ilex crenata | 221 | 12 | 209 | 1.04 | 5.83 | 3.08 |
各层生态火用和结构火用变异系数在4.17%—66.77%之间, 具有中等强度的空间变异性(表 2), 表明本研究数据差异性适中。各层生态火用与结构火用分析结果按生态火用贡献率大小排序。群落生态火用总值为1.71×1010 kJ/m2(表 3), 乔木层生态火用总值为1.26×1010 kJ/m2(表 4), 群落生态火用值集中于乔木层。黄山松在群落、乔木层和灌木层中贡献率均高于97%, 在群落(1.68×1010 kJ/m2)、乔木层(1.23×1010 kJ/m2)和灌木层(4.52×109 kJ/m2)中生态火用值均最高, 黄山松在群落中占有绝对优势。木荷在群落和乔木层中生态火用值仅次于黄山松, 分别为9.00×107 kJ/m2和8.78×107 kJ/m2(表 3、4、5)。乔木层结构火用总值为1.83×106 kJ/g(表 4), 灌木层结构火用总值为2.45×106 kJ/g(表 5), 灌木层在结构火用中占据较大比重。黄山松群落结构火用值最高(3.02×105 kJ/g)。综上表明黄山松种群对维持群落结构的复杂性和稳定性具有关键作用。乔木层是群落生态火用的主要贡献者, 灌木层是群落结构火用的主要贡献者。
| 种类 Types of exergy |
最小值 Minimum |
最大值 Maximum |
平均值 Average |
标准差 Standard deviation |
变异系数CV | 柯尔莫可洛夫-斯米洛夫检验 K-S test (P=0.05) |
| 群落生态火用The community Ex, CEx | 2.54×107kJ/m2 | 3.74×108kJ/m2 | 1.71×108kJ/m2 | 6.85×107kJ/m2 | 40.00% | 0.20* |
| 群落结构火用The community SpEx, CSpEx | 1.34×105kJ/m | 3.02×105kJ/m | 2.57×105kJ/m | 3.45×104kJ/m | 13.42% | 0.20* |
| 乔木层生态火用The tree layer Ex, TLEx | 4.27×104kJ/m2 | 3.21×108kJ/m2 | 1.25×108kJ/m2 | 7.46×107kJ/m2 | 59.73% | 0.20* |
| 乔木层结构火用The tree layer SpEx, TLSpEx | 7.11×104kJ/m | 3.02×105kJ/m | 2.39×105kJ/m | 5.30×104kJ/m | 22.14% | 0.20* |
| 灌木层生态火用The shrub layer Ex, SLEx | 7.61×105kJ/m2 | 1.21×108kJ/m2 | 4.29×107kJ/m2 | 2.86×107kJ/m2 | 66.77% | 0.20* |
| 灌木层结构火用The shrub layer SpEx, SLSpEx | 2.37×105kJ/m | 3.01×105kJ/m | 2.90×105kJ/m | 1.21×104kJ/m | 4.17% | 0.20* |
| *:数据经过约翰逊(Johnson)转换Data is Johnson transformed | ||||||
| 物种 Species |
生态火用Ex/ (kJ/m2) |
结构火用SpEx/ (kJ/g) |
群落贡献率 The community contribution rate, PC/% |
| 黄山松P. taiwanensis | 1.68×1010 | 3.02×105 | 98.397 |
| 木荷Schima superba | 9.00×107 | 4.52×104 | 0.526 |
| 红楠M. thunbergiisieb | 2.71×107 | 2.10×104 | 0.159 |
| 江南山柳C. cavaleriei | 2.57×107 | 1.42×104 | 0.150 |
| 大萼两广黄瑞木A. glischroloma var. macrosepala | 1.59×107 | 4.52×104 | 0.093 |
| 深山含笑Michelia maudiae | 1.17×107 | 4.53×104 | 0.068 |
| 密花山矾S. congesta | 1.00×107 | 1.99×104 | 0.059 |
| 山苍子Litsea cubeba | 9.18×106 | 3.51×104 | 0.054 |
| 羊舌树S. glauca | 8.75×106 | 1.99×104 | 0.051 |
| 鹿角杜鹃R. latoucheae | 8.38×106 | 9.22×103 | 0.048 |
| 总计Total | 1.71×1010 | 2.66×106 | 1 |
| 物种 Species |
生态火用Ex/ (kJ/m2) |
结构火用SpEx/ (kJ/g) |
乔木层贡献率 The tree layer contribution rate, PL/% |
| 黄山松P. taiwanensis | 1.23×1010 | 3.02×105 | 97.927 |
| 木荷S. superba | 8.78×107 | 4.52×104 | 0.698 |
| 红楠M. thunbergiisieb | 2.71×107 | 2.10×104 | 0.215 |
| 江南山柳C. cavaleriei | 2.52×107 | 1.42×104 | 0.201 |
| 大萼两广黄瑞木A. glischroloma var. macrosepala | 1.54×107 | 4.52×104 | 0.123 |
| 深山含笑M. maudiae | 1.14×107 | 4.53×104 | 0.091 |
| 密花山矾S. congesta | 1.00×107 | 1.99×104 | 0.079 |
| 山苍子L. cubeba | 8.76×106 | 3.51×104 | 0.069 |
| 福建柏Fokienia hodginsii | 8.35×106 | 1.56×105 | 0.066 |
| 鹿角杜鹃R. latoucheae | 8.04×106 | 9.22×103 | 0.064 |
| 总计Total | 1.26×1010 | 1.83×106 | 1 |
| 物种 Species |
生态火用Ex/ (kJ/m2) |
结构火用SpEx/ (kJ/g) |
灌木层贡献率 The shrub layer contribution rate, PL/% |
| 黄山松P. taiwanensis | 4.52×109 | 3.02×105 | 99.704 |
| 岗柃E. groffii | 4.11×106 | 4.85×104 | 0.091 |
| 木荷S. superba | 2.15×106 | 4.52×104 | 0.047 |
| 羊舌树S. glauca | 1.99×106 | 1.99×104 | 0.044 |
| 映山红R. pulchrum | 6.92×105 | 9.22×103 | 0.015 |
| 岩柃E. saxicola | 5.36×105 | 4.85×104 | 0.012 |
| 江南山柳C. cavaleriei | 4.62×105 | 1.42×104 | 0.010 |
| 大萼两广黄瑞木A. glischroloma var. macrosepala | 4.26×105 | 4.52×104 | 0.009 |
| 窄基红褐柃E. rubiginosa var. attenuata | 4.20×105 | 4.85×104 | 0.009 |
| 山苍子L. cubeba | 4.16×105 | 3.51×104 | 0.009 |
| 总计Total | 4.53×109 | 2.45×106 | 1 |
从表 6和图 2可知, 各生态火用和结构火用R2范围为0.490—0.950, 曲线精度较高, 各生态火用和结构火用残差和均接近于0, 拟合效果均较好, 表明模型可反映研究区生态火用和结构火用的空间分布特征。各生态火用和结构火用块金值均较小, 接近于0, 表明由实验误差和小于最小取样尺度引起的随机变异较小。乔木层结构火用基台值最大(1.070), 空间异质性程度最高;群落结构火用基台值最小(0.850), 空间异质性程度最低。研究区生态火用和结构火用有效变程波动幅度为15.20—29.10 m, 其中灌木层结构火用变程最大(29.10 m), 表明其空间连续性好。研究区内灌木层结构火用、乔木层结构火用、群落生态火用、乔木层生态火用、灌木层生态火用和群落结构火用分别在29.10 m、20.61 m、19.10 m、18.53 m、18.30 m和15.20 m范围内存在空间自相关。
| 种类 Types of exergy |
Moran指数 Moran I |
块金值 Co |
基台值 (Co+C) |
有效变程 Ao(m) |
结构比 C/(Co+C) |
理论模型 Theoretical model |
决定系数 R2 |
残差和 RSS |
| 群落生态火用CEx | 0.155 | 0.049 | 0.954 | 19.10 | 95.0% | Spherical球状模型 | 0.721 | 0.009 |
| 群落结构火用CSpEx | 0.102 | 0.001 | 0.850 | 15.20 | 99.9% | Spherical球状模型 | 0.625 | 0.011 |
| 乔木层生态火用TLEx | 0.245 | 0.135 | 0.943 | 18.53 | 85.7% | Gaussian高斯模型 | 0.868 | 0.006 |
| 乔木层结构火用TLSpEx | 0.122 | 0.182 | 1.070 | 20.61 | 83.0% | Gaussian高斯模型 | 0.950 | 0.004 |
| 灌木层生态火用SLEx | 0.247 | 0.024 | 0.938 | 18.30 | 97.4% | Exponential指数模型 | 0.490 | 0.011 |
| 灌木层结构火用SLSpEx | 0.300 | 0.146 | 1.055 | 29.10 | 86.2% | Exponential指数模型 | 0.766 | 0.013 |

|
| 图 2 生态火用和结构火用半方差函数曲线 Fig. 2 Semivariograms of eco-exergy and specific eco-exergy |
戴云山黄山松林各层生态火用和结构火用结构比均大于75%(表 6), 其在研究尺度上均具有强烈的空间自相关。在大于取样尺度小于有效变程范围内, 引起各层生态火用和结构火用空间变异的结构性因子占83.0%—99.9%, 随机性因子占0.1%—17%, 表明在群落形成过程中由土壤质地、地形、植物组成和微环境等结构性因子引起的空间异质性占主导地位, 说明本研究采样密度相对合理, 由人类干扰等随机性因子引起的空间异质性较弱。
4.4 生态火用和结构火用空间分布特征表 7中各生态火用和结构火用均值误差均较小, 趋近于0, 均方根误差和平均标准误差接近, 标准均方根预测误差趋近于1, 即各生态火用和结构火用指标插值图精度较高, 可用于生态火用和结构火用空间分布预测。从图 3可知, 各层生态火用和结构火用均呈明显条带状和斑块状分布, 等值线弯曲密集, 空间异质性程度较高, 在研究区东北角均出现低值区。各层生态火用值在研究区东边中部、西南部和东南部均分布有高值区。群落和乔木层的结构火用值均在西南部和东边中部出现高值区, 在西边中部出现低值区。灌木层结构火用值在西北边和东南边出现高值区。
| 指标Index | 群落生态火用CEx | 群落结构火用CSpEx | 乔木层生态火用TLEx | 乔木层结构火用TLSpEx | 灌木层生态火用SLEx | 灌木层结构火用SLSpEx |
| 均方根误差Root mean square error | 9.31×10-1 | 9.36×10-1 | 8.50×10-1 | 1.00×100 | 8.94×10-1 | 8.68×10-1 |
| 均值误差Mean error | -7.42×10-3 | 1.52×10-2 | -1.58×10-3 | 1.67×10-2 | 2.29×10-3 | 3.20×10-3 |
| 标准均方根误差Root mean square standardized error | 9.98×10-1 | 1.00×100 | 1.01×100 | 9.93×10-1 | 9.69×10-1 | 9.54×10-1 |
| 平均标准误差Average standard error | 9.33×10-1 | 9.32×10-1 | 8.46×10-1 | 1.01×100 | 9.52×10-1 | 9.13×10-1 |

|
| 图 3 黄山松林各层生态火用和结构火用空间分布图 Fig. 3 Spatial distribution map of eco-exergy and specific eco-exergy in P. taiwanensis forestry shrub layer |
戴云山黄山松在群落、乔木层和灌木层生态火用贡献率均高于97%, 在各层结构火用中也保持最高, 表明黄山松在群落结构稳定性中发挥着关键作用, 对构成复杂稳定的群落结构贡献最大。原因在于戴云山1400—1600 m海拔处, 黄山松种群优势明显(陈板岭头平均海拔1415 m)[19], 在群落中各物种资源利用共性差, 强烈排斥其他物种[21]。戴云山黄山松群落中大部分生态火用集中于乔木层, 结构火用与生态火用不同, 灌木层在群落中占据较大比重。Lu等[2]对中国东南部四种森林进行生态火用评估后发现, 亚热带人工林生态火用的主要贡献者为乔木层, 草本和灌木层是群落结构火用的主要贡献者;李浩等[38]对鹤山红木荷人工林群落结构的研究发现, 乔木层和草本层分别在生态火用和结构火用中占据较大比重;与本文研究结果保持一致。由于生态火用值高低主要取决于生物量, 乔木层拥有较高生物量, 促使乔木层成为生态火用的主要贡献者[51]。结构火用值的大小主要取决于物种和群落性质, 灌草层遗传信息较丰富, 因而灌草层是结构火用的主要贡献者[7]。
在生态系统演替过程中, 系统自发地向着生态火用最大化方向发展[24]。两个生态系统的生态火用和结构火用可以进行比较[5], 良好的生态系统应具有相对较高的生态火用和结构火用, 较高的生态火用意味系统保持其稳定方面效率更高, 较高的结构火用表示系统保持其复杂性方面效率更高[11, 41]。戴云山黄山松群落生态火用和结构火用总值为1.71×1010 kJ/m2和2.66×106 kJ/g, 位于鹤山国家森林生态系统野外研究站的28年林龄针叶人工林群落生态火用和结构火用总值均低于戴云山黄山松群落[2], 由于人工林物种丰富度低于天然林, 林分组成的复杂性较低, 且较发达的成熟生态系统具有相对较高的生态火用值[13]。相较于鹤山28年林龄针叶人工林, 戴云山黄山松林在保持其稳定和复杂性方面效率更高。
空间异质性由结构性因素和随机性因素共同作用产生[28]。秦倩倩等[52]发现结构性因素能增强空间相关性, 而随机性因素会减弱空间相关性, 与本文研究结果一致。戴云山黄山松群落形成过程中由结构性因子引起的空间异质性占主导地位, 生态火用和结构火用在研究尺度上均具有强烈空间自相关。苏松锦等[53]发现戴云山黄山松林土壤水分—物理性质的空间变异主要受控于地形。结合样地内地形、土壤含水量、土壤养分含量和黄山松种群空间分布发现[25, 28], 研究区东边中部、西南部和东南部土壤含水量、毛管孔隙度和氮元素含量均较高, 黄山松幼树在此区域呈聚集分布, 导致各层生态火用在研究地东边中部、西南部和东南部均分布有高值区。生态火用格局与土壤理化性质格局一致, 原因在于土壤养分影响了植物群落的种类组成与生活型特征[54—55]。乔木优势树种聚集分布的区域中, 物种多样性较高时, 植物生物量逐渐积累, 土壤养分含量增加, 群落生态火用值上升, 进而影响生态系统的结构和功能[56—57]。综上, 戴云山黄山松林群落生态火用空间异质性受土壤因子驱动。有效变程反映了区域变量空间自相关性的范围[44], 研究区生态火用和结构火用有效变程波动幅度差异较小, 为15.2—29.1 m, 表明生态火用和结构火用的空间异质性尺度较小。
生态火用理论还可用于生态系统发展动态变化分析, 通过定期监测系统各组分的生态火用值, 判断生态系统发展方向[8]。Lu等[3]发现20年后广东鹤山三种人工林群落生态火用均增加3倍以上, 群落结构趋于稳定。李浩等[38]发现15年后红木荷群落生态火用和结构火用均上升, 群落健康程度进一步提升。因此, 未来可定期监测戴云山黄山松群落生态火用值, 评估其生态系统健康状况, 为进一步科学管理戴云山黄山松群落提供依据。
6 结论戴云山黄山松群落生态火用值集中于乔木层;除优势树种黄山松外, 其他矮灌木和小乔木结构火用值也较可观。黄山松在群落中具有更高竞争能力和生存概率, 对维持群落结构稳定具有关键作用, 在群落中占有绝对优势。乔灌层生态火用和结构火用具有中等强度的空间变异性。各层生态火用和结构火用均在研究尺度上均具有强烈空间自相关。在群落形成过程中, 由土壤质地、地形、植物组成和微环境等结构性因子引起的空间异质性占主导地位。各层生态火用和结构火用均呈明显条带状及斑块状分布, 空间异质性程度较高, 在研究区东北角均出现低值区。各层生态火用值在研究区东边中部、西南部和东南部均分布有高值区, 表明研究区东边中部、西南部和东南部乔木层和灌木层生物量积累较高。黄山松种群在群落中具有较高热力学地位, 是维系其群落结构稳定和健康的重要物种。
| [1] |
王雄宾, 谷建才, 朱建刚, 鲁绍伟, 余新晓, 李永杰. 熵值计算及其在森林生物量分布中的应用. 北京林业大学学报, 2008, 30(S2): 160-164. |
| [2] |
Lu H F, Wang Z H, Campbell D E, Ren H, Wang J. Emergy and eco-exergy evaluation of four forest restoration modes in southeast China. Ecological Engineering, 2011, 37(2): 277-285. DOI:10.1016/j.ecoleng.2010.11.003 |
| [3] |
Lu H F, Fu F Y, Li H, Campbell D E, Ren H. Eco-exergy and emergy based self-organization of three forest plantations in lower subtropical China. Scientific Reports, 2015, 5: 15047. DOI:10.1038/srep15047 |
| [4] |
康鹏, 陈卫平, 王美娥. 基于生态系统服务的生态风险评价研究进展. 生态学报, 2016, 36(5): 1192-1203. |
| [5] |
Jørgensen S E. Application of holistic thermodynamic indicators. Ecological Indicators, 2006, 6(1): 24-29. DOI:10.1016/j.ecolind.2005.08.003 |
| [6] |
Tian Y, Lu H F, Wang J, Lin Y B, Campbell D E, Jian S G. Effects of canopy and understory nitrogen addition on the structure and eco-exergy of a subtropical forest community. Ecological Indicators, 2019, 106: 105459. DOI:10.1016/j.ecolind.2019.105459 |
| [7] |
Jørgensen S E. Application of exergy and specific exergy as ecological indicators of coastal areas. Aquatic Ecosystem Health & Management, 2000, 3(3): 419-430. |
| [8] |
Cai H Y, Lu H F, Tian Y, Liu Z F, Huang Y, Jian S G. Effects of invasive plants on the health of forest ecosystems on small tropical coral islands. Ecological Indicators, 2020, 117: 106656. DOI:10.1016/j.ecolind.2020.106656 |
| [9] |
Santos G M D, Linares M S, Callisto M, Marques J C. Two tropical biodiversity hotspots, two different pathways for energy. Ecological Indicators, 2019, 106: 105495. DOI:10.1016/j.ecolind.2019.105495 |
| [10] |
Ye S F, Zhang L P, Feng H. Ecosystem intrinsic value and its evaluation. Ecological Modelling, 2020, 430: 109131. DOI:10.1016/j.ecolmodel.2020.109131 |
| [11] |
Zhang J J, Gurkan Z, Jørgensen S E. Application of eco-exergy for assessment of ecosystem health and development of structurally dynamic models. Ecological Modelling, 2010, 221(4): 693-702. DOI:10.1016/j.ecolmodel.2009.10.017 |
| [12] |
Chen Q, Zhao Q, Chen P M, Lu H F, Jian S G. Eco-exergy based self-organization of the icrobenthic faunal assemblage during mangrove succession in Zhanjiang, China. Ecological Indicators, 2018, 95: 887-894. DOI:10.1016/j.ecolind.2018.08.044 |
| [13] |
Jørgensen S E, Nielsen S N, Mejer H. Emergy, environ, exergy and ecological modelling. Ecological Modelling, 1995, 77(2/3): 99-109. |
| [14] |
Sun J F, Yuan X Z, Liu G D, Tian K. Emergy and eco-exergy evaluation of wetland restoration based on the construction of a wetland landscape in the northwest Yunnan Plateau, China. Journal of Environmental Management, 2019, 252: 109499. DOI:10.1016/j.jenvman.2019.109499 |
| [15] |
O'Neill R V, Milne B T, Turner M G, Gardner R H. Resource utilization scales and landscape pattern. Landscape Ecology, 1988, 2(1): 63-69. DOI:10.1007/BF00138908 |
| [16] |
司建华, 冯起, 鱼腾飞, 常宗强, 席海洋, 苏永红. 额济纳绿洲土壤养分的空间异质性. 生态学杂志, 2009, 28(12): 2600-2606. |
| [17] |
苏松锦. 戴云山高海拔黄山松林的空间异质性与植被维持机制. 福州: 福建农林大学, 2015.
|
| [18] |
闫东锋, 贺文, 杨喜田. 栓皮栎人工林灌木层植物多样性的空间分布及其与光环境的关系. 应用生态学报, 2020, 31(11): 3605-3613. |
| [19] |
刘金福, 朱德煌, 兰思仁, 洪伟, 郑世群, 何中声, 徐道炜. 戴云山黄山松群落与环境的关联. 生态学报, 2013, 33(18): 5731-5736. |
| [20] |
江蓝, 魏晨思, 何中声, 朱静, 邢聪, 王雪琳, 刘金福, 沈彩霞, 施友文. 格氏栲天然林林窗植物群落功能性状的变异. 植物生态学报, 2022, 46(3): 267-279. |
| [21] |
苏松锦, 刘金福, 兰思仁, 洪伟, 李文周. 黄山松研究综述(1960-2014)及其知识图谱分析. 福建农林大学学报: 自然科学版, 2015, 44(5): 478-486. |
| [22] |
朱德煌, 刘金福, 洪伟, 郑世群, 何中声, 徐道炜, 李文周. 戴云山黄山松群落主要树种更新生态位研究. 热带亚热带植物学报, 2012, 20(6): 561-565. |
| [23] |
任国学, 刘金福, 徐道炜, 洪伟, 郑世群, 黄志森. 戴云山国家级自然保护区黄山松群落类型与物种多样性分析. 植物资源与环境学报, 2011, 20(3): 82-88. |
| [24] |
李晴新, 冯剑丰, 朱琳. 生态能质(eco-exergy)在水生生态系统建模和评价中的应用. 生态学杂志, 2011, 30(2): 376-383. |
| [25] |
Liu J F, Su S J, He Z S, Jiang L, Gu X G, Xu D W, Ma R F, Hong W. Relationship between Pinus taiwanensis seedling regeneration and the spatial heterogeneity of soil nitrogen in Daiyun Mountain, southeast China. Ecological Indicators, 2020, 115: 106398. DOI:10.1016/j.ecolind.2020.106398 |
| [26] |
何中声, 陈佳嘉, 朱静, 王紫薇, 谷新光, 江蓝, 陈博, 徐道炜, 吴则焰, 刘金福. 戴云山南坡不同海拔森林土壤微生物功能多样性特征及影响因素. 生态学报, 2022, 42(9): 3504-3515. |
| [27] |
何中声, 王紫薇, 朱静, 陈佳嘉, 谷新光, 江蓝, 陈博, 吴则焰, 刘金福, 陈文伟. 戴云山南坡不同海拔森林土壤微生物群落结构特征和影响因素. 环境科学, 2022, 43(5): 2802-2811. |
| [28] |
苏松锦, 刘金福, 马瑞丰, 洪伟, 陈志芳, 刘伟, 祁丽霞. 戴云山黄山松幼苗更新与土壤有机碳空间异质性的关系. 应用与环境生物学报, 2014, 20(6): 986-991. |
| [29] |
徐道炜, 刘金福, 何中声, 郑世群. 毛竹向杉木林扩张后的群落物种多样性特征. 森林与环境学报, 2019, 39(1): 37-41. |
| [30] |
郑世群, 刘金福, 任国学, 李文周, 洪伟, 徐道炜. 戴云山黄山松群落乔木层主要种群生态位研究. 福建林学院学报, 2012, 32(3): 193-198. |
| [31] |
江蓝, 何中声, 刘金福, 冯雪萍, 刘艳会, 陈文伟. 戴云山黄山松种群径级结构的海拔分布格局. 福建农林大学学报: 自然科学版, 2019, 48(5): 585-590. |
| [32] |
董果. 太行山南麓荆条灌丛土壤氮磷空间变化特征研究. 郑州: 河南农业大学, 2014.
|
| [33] |
Lü M, Sun M K, Peñuelas J, Sardans J, Sun J, Chen X P, Zhong Q L, Cheng D L. Temperature controls growth of Pinus taiwanensis along an elevational gradient. Trees, 2021, 35(2): 433-440. |
| [34] |
国家林业局. LY/T 2660—2016立木生物量模型及碳计量参数木荷. 北京: 中国标准出版社, 2016.
|
| [35] |
左舒翟, 任引, 翁闲, 丁洪峰, 罗云建. 亚热带常绿阔叶林9个常见树种的生物量相对生长模型. 应用生态学报, 2015, 26(2): 356-362. |
| [36] |
张倩媚, 温达志, 叶万辉, 孔国辉. 南亚热带常绿阔叶林林下层植物的生物量及其测定方法的探讨. 生态科学, 2000, 19(4): 62-66. |
| [37] |
孙杰杰, 江波, 吴初平, 袁位高, 朱锦茹, 黄玉洁, 焦洁洁, 沈爱华. 浙江省檫木林生境与生态位研究. 生态学报, 2019, 39(3): 884-894. |
| [38] |
李浩, 曾宪曙, 陆宏芳. 基于生态(Eco-exergy)的鹤山红木荷人工林群落结构发展动态. 生态环境学报, 2012, 21(11): 1822-1829. |
| [39] |
Wang J, Lu H F, Lin Y B, Campbell D E, Cai H Y, Ren H. Dynamics of community structure and bio-thermodynamic health of soil organisms following subtropical forest succession. Journal of Environmental Management, 2021, 280: 111647. |
| [40] |
Chen Q, Yuan H R, Chen P M. Short-term effects of artificial reef construction on the taxonomic diversity and eco-exergy of the icrobenthic faunal community in the Pearl River Estuary, China. Ecological Indicators, 2019, 98: 772-782. |
| [41] |
Xu F, Yang Z F, Chen B, Zhao Y W. Ecosystem health assessment of the plant-dominated Baiyangdian Lake based on eco-exergy. Ecological Modelling, 2011, 222(1): 201-209. |
| [42] |
Royal Botanic Gardens. Kew: Plant DNA C-values database. [2021-4-28]. http://cvalues.science.kew.org.
|
| [43] |
Fonseca J C, Marques J C, Paiva A A, Freitas A M, Madeira V M C, Jørgensen S E. Nuclear DNA in the determination of weighing factors to estimate exergy from organisms biomass. Ecological Modelling, 2000, 126(2/3): 179-189. |
| [44] |
鱼腾飞, 冯起, 司建华, 席海洋, 陈丽娟. 黑河下游额济纳绿洲植物群落物种多样性的空间异质性. 应用生态学报, 2011, 22(8): 1961-1966. |
| [45] |
苏松锦, 刘金福, 何中声, 洪伟, 张金彪. 格氏栲天然林土壤养分空间异质性. 生态学报, 2012, 32(18): 5673-5682. |
| [46] |
王跃, 张鹏新, 党跃武. Box-Cox和Johnson方法在油气井数据预处理中的研究. 云南民族大学学报: 自然科学版, 2018, 27(4): 340-347. |
| [47] |
李长江, 邓文平, 曹元元, 鲍宇. 基于Box-Cox变换与Johnson变换非正态过程能力分析. 齐齐哈尔大学学报, 2015, 31(1): 66-70. |
| [48] |
李艳丽, 杨华, 亢新刚, 王妍, 岳刚, 沈林. 长白山云冷杉针阔混交林天然更新空间分布格局及其异质性. 应用生态学报, 2014, 25(2): 311-317. |
| [49] |
富金赤, 李晓莎, 许中旗, 郭延鹏, 赵娱, 李校. 冀北山地阳坡草本植物物种多样性的空间异质性研究. 草地学报, 2018, 26(6): 1298-1304. |
| [50] |
李俊晓, 李朝奎, 殷智慧. 基于ArcGIS的克里金插值方法及其应用. 测绘通报, 2013(9): 87-90, 97-97. |
| [51] |
Jørgensen S E, Nielsen S N. Thermodynamic orientors: exergy as a goal function in ecological modeling and as an ecological indicator for the description of ecosystem development//Müller F, Leupelt M, eds. Eco Targets, Goal Functions, and Orientors. Berlin, Heidelberg: Springer, 1998: 63-86.
|
| [52] |
秦倩倩, 王海燕, 李翔, 雷相东, 解雅麟, 郑永林, 耿琦. 东北天然针阔混交林凋落物磷素空间异质性及其影响因素. 生态学报, 2019, 39(12): 4519-4529. |
| [53] |
苏松锦, 刘金福, 陈文伟, 旷开金, 唐蓉, 洪伟. 戴云山黄山松林土壤水分物理性质空间变异特征与格局. 资源科学, 2014, 36(11): 2423-2430. |
| [54] |
金章利, 刘高鹏, 周明涛, 许文年. 喀斯特山地草地群落多样性海拔特征及土壤理化性质特征. 生态环境学报, 2019, 28(4): 661-668. |
| [55] |
Verdú J R, Lobo J M, Sánchez-Piñero F, Gallego B, Numa C, Lumaret J P, Cortez V, Ortiz A J, Tonelli M, García-Teba J P, Rey A, Rodríguez A, Durán J. Ivermectin residues disrupt dung beetle diversity, soil properties and ecosystem functioning: an interdisciplinary field study. Science of the Total Environment, 2018, 618: 219-228. |
| [56] |
舒韦维, 卢立华, 李华, 农友, 何日明, 陈海, 黄彪. 林分密度对杉木人工林林下植被和土壤性质的影响. 生态学报, 2021, 41(11): 4521-4530. |
| [57] |
Muvengwi J, Ndagurwa H G T, Nyenda T. Enhanced soil nutrient concentrations beneath-canopy of savanna trees infected by mistletoes in a southern African savanna. Journal of Arid Environments, 2015, 116: 25-28. |