 2023, Vol. 43
2023, Vol. 43文章信息
- 梁元也, 张玲雪, 周小龙, 范连连, 毛洁菲, 李耀明
- LIANG Yuanye, ZHANG Lingxue, ZHOU Xiaolong, FAN Lianlian, MAO Jiefei, LI Yaoming
- 天山高寒草原土壤团聚体结构与养分含量对模拟氮磷沉降的响应
- Response of soil aggregate structure and nutrient content to the simulated nitrogen and phosphorus deposition in an alpine steppe of Tianshan Mountains
- 生态学报. 2023, 43(10): 4130-4141
- Acta Ecologica Sinica. 2023, 43(10): 4130-4141
- http://dx.doi.org/10.5846/stxb202206081632
-
文章历史
- 收稿日期: 2022-06-08
- 采用日期: 2023-02-06





2. 新疆大学生态与环境学院, 乌鲁木齐 830046;
3. 新疆大学绿洲生态教育部重点实验室, 乌鲁木齐 830046;
4. 中国科学院中亚生态与环境研究中心, 乌鲁木齐 830011;
5. 中国科学院巴音布鲁克草原生态系统研究站, 巴音布鲁克 841314;
6. 中国科学院大学, 北京 100049
2. College of Ecology and Environment, Xinjiang University, Urumqi 830046, China;
3. Key Laboratory of Oasis Ecology of Ministry of Education, Xinjiang University, Urumqi 830046, China;
4. Research Center for Ecology and Environment of Central Asia, Chinese Academy of Sciences, Urumqi 830011, China;
5. Bayinbuluk Grassland Ecosystem Research Station, Chinese Academy of Sciences, Bayinbuluk 841314, China;
6. University of Chinese Academy of Sciences, Beijing 100049, China
草地是陆地生态系统的重要组成部分, 具有涵养水源、保持水土、固碳、维持生物多样性和生态系统稳定等重要生态功能, 草地土壤在生态系统功能和动态中起着主导作用[1—3]。土壤团聚体是土壤颗粒在有机与无机胶结物质作用下形成的聚合体[4], 目前通常以 < 0.053 mm、0.053—0.25 mm、0.25—2 mm和>2 mm作为土壤团聚体常用粒径分区[5]。土壤团聚体作为土壤结构的基本单元[6], 其稳定性在一定程度上反映了土壤的孔隙度、渗透性、持水性、抗侵蚀能力等物理性质[7—8]。土壤团聚体还可以改变不同土壤组分间的养分分布, 主导土壤养分积累的变化, 尤其是作为土壤固碳功能的载体而影响土壤肥力[7—8]。因此, 土壤团聚体的稳定性是土壤质量的重要评价标准之一, 提高土壤团聚体的含量和稳定性是改善土壤结构和肥力的关键途径[6]。
近几十年来, 全球人口的增长和工业化的发展等一系列问题极大增加了大气氮、磷沉降[9]。在全球范围内, 自20世纪初至今大气氮沉降使陆地生态系统氮素增加了3到5倍[10], 而2000年的磷沉积是其自然本底值的2.5到5倍[11]。其中, 中国的氮磷沉降增长幅度最大[9]。土壤团聚体的结构和稳定性受到气候因素、土壤碳含量、土壤关键营养元素可利用性、土壤微生物、植物根系等因素的共同影响[12]。氮磷沉降通过改变土壤理化性质、土壤微生物群落结构、植物群落特征与植物功能性状以及全球气候来影响土壤团聚体的结构和养分承载能力, 进而影响土壤质量[12—14]。
国内外已广泛开展有关氮磷添加对草地生态系统土壤团聚体影响的研究, 然而关于氮添加对土壤团聚体影响的研究结果并不一致[15]。有研究发现氮添加显著提高了>0.25 mm团聚体比例与稳定性及其有机碳含量[12, 16—17];也有研究表明氮添加并未显著影响草地土壤团聚体结构或土壤碳库[18—19]。有关磷添加和氮磷交互添加对草地土壤团聚体影响的研究与氮添加相比相对较少, 研究表明磷添加和氮磷交互添加也会对草地土壤团聚体产生积极[20]、消极[12]或中性[15]的影响。相比于农田生态系统和森林生态系统[21—23], 氮磷添加对草地生态系统土壤团聚体影响的研究相对较少。在农田或氮素限制的生态系统中, 氮添加对有机碳的促进作用主要通过植物生产力的增加;而在森林和草地生态系统中, 微生物对有机质的分解作用以及植物输入是调节土壤碳库发生改变的主要因素[15]。关于氮磷添加对草地土壤团聚体的研究目前主要集中在典型草原[17, 20]、荒漠草原[24]两种草地类型, 对高寒草原的研究较少。因此, 氮磷添加对高寒草原土壤团聚体的结构、稳定性与养分含量有何影响有待于进一步开展相关研究。
巴音布鲁克草原位于新疆天山南麓, 是中国著名的亚高山高寒草原[25]。它是全疆尤其是南疆的重要生态屏障, 承载着重要的生态功能, 也是新疆最大的畜牧业基地, 决定着当地的经济发展[26]。同时, 又因特殊的地理位置与高寒的气候条件, 巴音布鲁克草原是生态脆弱区和全球变化的敏感区[27]。氮磷沉降导致的植物、土壤、微生物以及气候变化对巴音布鲁克草原土壤团聚体的影响尚不明确。因此, 本实验以巴音布鲁克高寒草原为研究对象, 以全球营养网络(https://nutnet.org/)为实验设计的基础, 研究短期(4年)氮磷添加对不同土壤层团聚体结构及其养分含量的影响, 为预测未来草地生态系统的生产力、承载力、土壤质量等的变化趋势提供数据支持与理论基础, 以期实现草地可持续利用。
1 材料与方法 1.1 研究区概况研究区位于新疆巴音郭楞蒙古自治州和静县内的中国科学院巴音布鲁克草原生态系统研究站(42°52′—42°53′ N, 83°42′—83°43′ E)(图 1), 研究地点海拔约2470 m, 年平均气温约-4.8 ℃, 年平均降水量约260 mm, 年蒸发量1022—1247 mm, 全年积雪天数150—180 d, 属典型的高寒气候, 主要土壤类型为栗钙土。研究地点是以羊茅(Festuca ovina)为优势种的高寒草原, 其他主要伴生物种为冰草(Agropyron cristatum)、二裂委陵菜(Potentilla bifurca)、莓叶委陵菜(Potentilla fragarioides)等。

|
| 图 1 研究区位置图 Fig. 1 Location map of the study area |
本研究依托全球营养网络在巴音布鲁克站点的实验开展。于2018年选择常年禁牧、生境均匀和植被长势良好的高寒草地进行围封作为研究对象, 通过使用尿素[CO(NH2)2]、磷酸二氢钙[Ca(H2PO4)2], 换算为实际氮、磷含量, 以10 g m-2 a-1为养分添加量单位进行添加。共设置4个处理, 分别为对照(CK), N添加、P添加、NP交互添加处理, 每个处理设置3次重复, 共计12个样方。实验布局采用随机区组的实验设计方法, 共设有3个面积为25 m2(5 m×5 m)的处理样区, 每个样区包含4个面积为4 m2(2 m×2 m)的样方, 样方间留有1 m间隔。于每年5月中旬, 选择在阴雨天气, 将预先称好的颗粒状肥料一次性均匀洒在样方内。该地区2013年N的平均沉降量为0.8 g N m-2 a-1, P的平均沉降量为0.01 g P m-2 a-1[9]。
1.2.2 植物样品采集、处理与测定2021年8月初在植物生长旺盛期采集植物地上部分样品。在每个样方内随机设置一个0.4 m×0.4 m的小样方, 采集小样方内所有植物的地上部分, 装入信封带回实验室于105 ℃烘箱中杀青30 min, 然后将温度调至65 ℃烘干至恒重, 用精度为0.001 g的天平称重获取植物干重, 通过换算获取植物地上生物量(Aboveground biomass, AGB), 单位为g/m2。
1.2.3 土壤样品采集、处理与测定采集植物样品的同时获取土壤样品。在每个2 m×2 m样方内按“S”型随机取样法选取3个点, 用直径为10 cm的土钻采集0—10 cm土层与10—20 cm土层的土壤样品, 并将每层土壤的三个样品混匀为一个混合土样。随后去除土壤样品中的根、石块等杂质, 把每个混合土样分为3份保存:1份保存于4 ℃的冰箱中用于后续土壤微生物量碳、氮的测定;1份自然风干后过筛, 用于测定土壤化学性质;1份自然风干后, 用于土壤团聚体筛分。
土壤团聚体的测定采用湿筛法[28]:称取约50 g的风干土放置于土壤团聚体湿筛分析仪套筛(从上到下依次为2 mm、0.25 mm和0.053 mm孔径)的最上层, 水浸15 min后震荡15 min(振幅为5 cm, 频率为每分钟30次), 以实现团聚体的分级。震荡结束后将每个筛上保留的水稳定性团聚体冲洗到容器中, 静置2 h, 小心去掉上清液后将其放置于60 ℃烘箱中烘干至恒重。烘干后用精度为0.001 g的天平称重, 以获取 < 0.053 mm、0.053—0.25 mm、0.25—2 mm和>2 mm四个粒级的团聚体重量。称重完成后测定0.053—0.25 mm、0.25—2 mm、>2 mm三个粒级土壤团聚体有机碳(Soil organic carbon, SOC)、总氮(Total nitrogen, TN)、总磷(Total phosphorus, TP)、速效氮(Available nitrogen, AN)、速效磷(Available phosphorus, AP)含量。
土壤pH采用电极法测定, SOC采用重铬酸钾-外加热法测定, 土壤TN采用凯氏定氮法测定, 土壤TP采用高氯酸-硫酸-钼锑抗比色法测定, 土壤AN采用碱解扩散法测定, 土壤AP采用碳酸氢钠-钼锑抗比色法测定[29]。土壤微生物量碳(Microbial biomass carbon, MBC)与土壤微生物量氮(Microbial biomass nitrogen, MBN)采用氯仿熏蒸法测定[30]。
1.3 数据处理与分析 1.3.1 数据处理各粒级土壤团聚体百分比含量(Wi)计算方法为公式(1)[31];土壤团聚体的稳定性用平均重量直径(Mean weight diameter, MWD)来表示, 计算方法为公式(2)[32]:
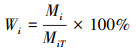
|
(1) |
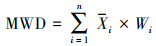
|
(2) |
式中, Wi为各粒级土壤团聚体百分比含量(%), Mi为不同粒级土壤团聚体质量(g), MiT为土壤样品总重量(g);Xi为土壤团聚体平均直径(mm)。
N有效率和P有效率计算方法分别为公式(3)和公式(4)[33]:
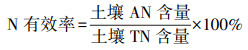
|
(3) |

|
(4) |
利用单因素方差分析及多重比较(LSD)研究氮磷添加对土壤全土理化性质、土壤团聚体结构及其养分含量的影响。利用Pearson相关分析来研究AGB与团聚体AN、AP含量间的相关性以及氮磷添加背景下团聚体养分含量与其N、P有效率间的相关性。本研究运用IBM Statistics SPSS 25.0以及Origin 2022软件进行统计分析, 使用Origin 2022软件作图。所有数值采用平均值±标准误表示。
2 结果与分析 2.1 氮磷添加对土壤理化性质和地上生物量的影响通过图 2可知, 连续4年氮磷添加后, 巴音布鲁克高寒草原土壤理化性质发生了显著变化。0—10 cm土壤层SOC浓度范围为28.53—63.54 g/kg, TN为3.62—4.52 g/kg, TP为0.71—0.92 g/kg;10—20 cm土壤层SOC浓度范围为19.70—42.15 g/kg, TN为2.38—4.03 g/kg, TP为0.63—0.80 g/kg。氮磷添加对两土壤层SOC、TN、TP及AN含量影响整体上不显著。与对照相比, 10—20 cm土壤层N添加处理下土壤SOC含量显著降低31.60%(P < 0.05);P添加和NP交互添加显著提高了0—10 cm土壤层AP含量, P添加显著提高了10—20 cm土壤层AP含量(P < 0.05)。氮磷添加没有显著改变两土壤层pH值。

|
| 图 2 氮磷添加处理对土壤理化性质、土壤微生物量以及植物地上生物量的影响 Fig. 2 Effects of N and P addition on soil physical and chemical properties, soil microbial biomass and aboveground biomass CK:对照处理;N:氮添加;P:磷添加;NP:氮磷交互添加; 不同小写字母表示同一土层不同处理间差异显著(P < 0.05) |
氮磷添加对土壤微生物量碳、氮含量有显著影响(P < 0.05)。在0—10 cm土壤层, 与对照相比, P添加显著降低了土壤MBC含量(P < 0.05);在10—20 cm土壤层, P添加和NP交互添加处理下土壤MBC含量均显著高于对照(P < 0.05)。与对照相比, 氮磷添加处理降低了两土壤层MBN含量。N添加处理下植物AGB高于对照, 但在统计学上并不显著。
2.2 氮磷添加对土壤团聚体粒级组成及稳定性的影响通过图 3可知, 氮磷添加显著影响土壤团聚体粒级组成(P < 0.05)。0—10 cm与10—20 cm土壤层团聚体粒级组成相似, 两土层各粒级团聚体比例平均值从低到高依次为:0.053—0.25 mm(9.23%)、< 0.053 mm(14.24%)、0.25—2 mm(15.76%)和>2 mm(60.75%)。在0—10 cm土壤层中, N添加处理下>2 mm团聚体比例显著低于对照和其他处理, 其中P添加处理下>2 mm团聚体比例比N添加高36.7%(P < 0.05)。在10—20 cm土壤层中, P添加处理下>2 mm团聚体比例显著低于对照和其他处理(P < 0.05)。

|
| 图 3 土壤团聚体组成及其稳定性对氮磷添加的响应 Fig. 3 The response of soil aggregate structure and stability to N and P addition |
图 3表明, 氮磷添加显著影响土壤团聚体稳定性(P < 0.05)。相比于对照, N添加处理下0—10 cm土壤层团聚体MWD显著降低;P添加处理下10—20 cm土壤层团聚体MWD显著降低(P < 0.05)。
2.3 氮磷添加对土壤团聚体养分含量与氮、磷有效率的影响通过图 4可知, 氮磷添加处理显著影响土壤团聚体养分含量(P < 0.05)。相比于对照, 0—10 cm土壤层中, N添加使>2 mm、0.25—2 mm与0.053—0.25 mm粒级团聚体SOC含量分别提高了19.33%、19.77%和11.69%, P添加和NP交互添加降低了SOC含量, 但在统计学上并不显著。10—20 cm土壤层各粒级团聚体SOC含量显著低于0—10 cm土壤层(P < 0.05)。氮磷添加对0—10 cm土壤层团聚体TN含量没有显著影响;相比于对照, N添加、P添加显著提高了10—20 cm土壤层团聚体TN含量, NP处理显著降低了10—20 cm土壤层团聚体TN含量(P < 0.05)。氮磷添加对团聚体TP影响总体上不显著。与对照相比, N添加显著提高了0—10 cm土壤层各粒级团聚体AN含量, 降低了AP含量(P < 0.05)。

|
| 图 4 氮磷添加处理对土壤团聚体有机碳、总氮、总磷、速效氮、速效磷的影响 Fig. 4 Effects of N and P addition on aggregate-associated organic carbon, total nitrogen, total phosphorus, available nitrogen and available phosphorus 不同大写字母表示同一处理不同土层间差异显著(P < 0.05) |
通过表 1可知, 0—10 cm土壤层N添加处理下各粒级团聚体N有效率显著高于对照和其他处理(P < 0.05);两土层中N添加处理下各粒级团聚体P有效率均低于对照;NP交互添加处理下10—20 cm土壤层各粒级团聚体P有效率显著低于对照(P < 0.05)。
| 处理 Treatment |
氮有效率 Percentage of available N in total N/% |
磷有效率 Percentage of available P in total P/% |
|||||
| >2 mm | 0.25—2 mm | 0.053—0.25 mm | >2 mm | 0.25—2 mm | 0.053—0.25 mm | ||
| 0—10 cm | |||||||
| CK | 3.36±0.27bA | 3.45±0.23bA | 3.75±0.12bA | 2.17±0.53aA | 2.74±0.39aA | 2.89±0.50aA | |
| N | 4.35±0.57aA | 4.24±0.62aA | 4.17±0.58aA | 1.17±0.09aA | 1.24±0.05bA | 1.28±0.06aA | |
| P | 3.30±0.02bA | 3.33±0.19bA | 3.42±0.19bA | 2.51±0.15aA | 2.39±0.15aA | 2.65±0.42aA | |
| NP | 3.43±0.64bA | 3.76±0.24bA | 3.40±0.13bA | 2.81±0.56aA | 3.03±0.44aA | 2.74±0.64aA | |
| 10—20 cm | |||||||
| CK | 3.36±0.09aA | 3.37±0.02aA | 3.08±0.12aB | 2.89±0.02aA | 2.85±0.05aA | 2.86±0.04aA | |
| N | 1.91±0.45bB | 2.43±0.49aB | 2.95±0.19aB | 1.19±0.21cA | 1.17±0.22cA | 1.18±0.22bA | |
| P | 3.07±0.08aB | 2.82±0.08aB | 3.14±0.09aA | 2.03±0.02Bb | 2.14±0.21bA | 2.40±0.45aA | |
| NP | 3.45±0.42aA | 3.22±0.16aA | 3.50±0.04aA | 1.58±0.15cA | 1.57±0.06cBA | 1.56±0.09bA | |
0—10 cm土壤层中, 0.25—2 mm和0.053—0.25 mm粒级团聚体AN含量与AGB均显著正相关(图 5)。由直线斜率可知, 随着AGB的增加, >2 mm团聚体的AN含量增量最大。然而0—10 cm土壤层各粒级团聚体AP含量与AGB没有显著的相关性。

|
| 图 5 0—10 cm土层不同粒级土壤团聚体速效氮、速效磷含量与地上生物量的相关关系 Fig. 5 Correlation between aggregate-associated available nitrogen, available phosphorus in the soil layer of 0—10 cm and aboveground biomass NS:不显著;*:显著性水平P < 0.05;**:显著性水平P < 0.01;***:显著性水平P < 0.001 |
通过图 6可知, 连续4年氮磷添加背景下两土层团聚体内养分与N、P有效率间有显著的相关性。0—10 cm土壤层中团聚体SOC含量与AN含量、N有效率显著正相关, 与AP含量、P有效率显著负相关;AN含量与AP含量、P有效率显著负相关;AN含量与N有效率显著正相关, 相关系数为0.526;AP含量与P有效率显著正相关, 相关系数为0.963;AP含量与N有效率显著负相关(P < 0.05)。10—20 cm土壤层中, SOC与P有效率显著正相关;TN含量与N有效率显著负相关;TP含量、AN含量、AP含量、N有效率、P有效率之间分别显著正相关(P < 0.05)。

|
| 图 6 土壤团聚体养分及其N、P有效率间的相关分析 Fig. 6 Correlation analysis between aggregate-associated nutrients and percentage of available N and P in total N and P SOC:土壤有机碳;TN:全氮;TP:全磷;AN:速效氮;AP:速效磷 |
作为土壤结构的基本单元, 土壤团聚体的粒级组成可以直接影响土壤的孔隙结构, 改变土壤的通气性和渗透性, 决定土壤对水分和养分的吸收, 并影响土壤生产力[8]。而团聚体的稳定性代表着土壤的固碳和抗侵蚀能力[3]。MWD是评估土壤团聚体稳定性的关键指标, MWD值越大, 土壤平均粒径的团聚度越高, 稳定性越好, 土壤抗侵蚀能力越强[8]。
巴音布鲁克高寒草原0—10 cm和10—20 cm土壤层均以>2 mm团聚体占主导, 该结果与马盼盼等[34]的研究结果相类似。土壤中>2 mm团聚体比例越高, 越利于维持土壤结构的稳定性[8]。但是, N添加却显著降低了0—10 cm土层中>2 mm团聚体比例及团聚体MWD(图 3), 这些结果与郭虎波等[35]、Huang等[23]的研究结果一致。之所以出现这种结果, 主要与土壤中SOC的动态平衡有关[15]。SOC被视为土壤团聚体的主要胶结物质, 可以通过吸附在有机或无机粘土表面形成土壤团聚体[12]。高N添加会加速SOC矿化分解, 不利于SOC的积累, 进而阻碍土壤团聚体的形成和稳定[17]。而N添加处理下AGB的提高会增加凋落物质量, 进而提高土壤SOC含量, 可以在一定程度上抵消高N添加引起的SOC加速矿化分解[15], 所以全土和各粒级团聚体SOC含量未发生显著变化。N添加对>2 mm团聚体比例和团聚体稳定性的影响主要集中在0—10 cm土壤层, 对10—20 cm土壤层没有显著影响。
本研究发现, 10—20 cm土层中, 相比于对照, P添加显著降低了>2 mm团聚体比例以及团聚体MWD(图 3)。这与Huang等[23]和Du等[36]的研究结果一致, 即P添加会促进大团聚体分解为微团聚体。Huang等[23]的研究表明P添加会改变植物的养分获取策略, 减少植物对地下养分获取的投资, 导致根系生物量较少, 从而降低土壤碳含量。本研究中, 0—10 cm土壤层P添加处理下各粒级团聚体SOC浓度低于对照, 可能是P添加导致根系生物量与根系分泌物的减少, 从而降低了表土中有机质含量向深处输入, 不利于土壤颗粒的团聚。另外, 根据微生物N挖掘理论[37], 在N有限的情况下, 有些微生物通过分解土壤中难降解有机质来满足其对N的需求。基于这一理论, P添加会刺激微生物生长, 增加微生物对N的需求, 增加微生物的采N量, 这可能会造成对>0.25 mm团聚体的破坏[38]。本研究中, 0—10 cm土壤层P添加处理下土壤MBC含量显著高于对照(图 2), 说明P添加增加了土壤微生物生物量, 刺激了微生物生长, 进而有可能增加微生物对N的需求, 导致大粒级团聚体比例的下降。然而具体的解释还需要量化氮磷添加下土壤团聚体形成、稳定和分解的转化过程, 并深入研究微生物活性的变化。
3.2 氮磷添加对不同土壤层团聚体养分分布的影响作为土壤结构的基本单元, 土壤团聚体主导着土壤养分积累的变化[8]。土壤团聚体与SOC存在反馈调节机制:一方面, SOC可以作为胶结物质促进土壤团聚体的形成和稳定;另一方面, 团聚体可以作为SOC的物理屏障, 减少微生物降解和干扰[12]。
本研究结果表明0—10 cm与10—20 cm土壤层团聚体SOC含量存在显著差异(图 4)。0—10 cm土壤层水热条件好, 有较高的凋落物输入与植物根系, 且微生物生物量高[12]。因此, 0—10 cm土壤层各粒级团聚体SOC含量明显高于10—20 cm土壤层, 这与凌小莉等[12]的研究结果相同。本研究发现, 与对照相比, N添加处理下0—10 cm土层各粒级团聚体SOC含量有一定程度提高, 但在统计学上并不显著(图 4)。我们推测N添加处理下土壤团聚体SOC含量取决于初级生产力对SOC的输入和微生物对SOC的分解矿化作用之间的相对平衡[15]。本研究中, 0—10 cm土壤层N处理下各粒级团聚体AN含量及N有效率显著高于对照, 说明N添加提高了土壤N有效性。而团聚体AN含量与AGB呈显著正相关关系(图 5), 说明N添加通过提高AGB增加了团聚体AN含量[39]。植物的生长和凋落物的积累会导致土壤团聚体SOC输入的增加[40]。然而过高浓度的N添加会加速SOC的矿化, 导致土壤中积累的SOC总量减少, 抵消了植物生产力增加带来的SOC含量的提高, 最终使土壤中SOC含量没有显著变化[17]。
0—10 cm土壤层P添加处理和NP交互添加下各粒级团聚体SOC含量低于对照(图 4)。Li等[41]将P添加下SOC含量的降低归因于植物细根生物量的减少, 其研究认为, 在施肥条件下植物会减少对地下养分获取的投入[42], 导致根系生物量及根系分泌物的减少。0—10 cm土层P添加处理下MBC显著低于对照, 说明P添加改变了土壤微环境, 降低了土壤微生物量。P元素添加导致的0—10 cm土壤层团聚体SOC和全土MBC含量的下降都表明P添加不利于土壤碳含量的提高[43]。本研究还发现, 0—10 cm土壤层团聚体SOC含量对氮磷添加的响应相比于10—20 cm土壤层更强烈。
氮磷添加未显著改变0—10 cm土壤层各粒级团聚体TN含量(图 4), 这可能是因为氮磷添加增加了植物和微生物对无机N的吸收利用, 使得土壤团聚体TN没有显著变化[44]。但在10—20 cm土壤层, 相比于对照, N添加和P添加显著提高了各粒级团聚体TN含量(图 4)。这可能是因为, 相比于0—10 cm土壤层, 10—20 cm土壤层植物根系少且土壤微生物量较低, 植物和土壤微生物对无机N的吸收利用程度更小, 使氮素在10—20 cm土壤层得以积累[44]。团聚体中TP含量比较稳定(图 4), 对氮磷添加的响应很小, 这与Yang等[45]的研究结果一致。这是因为土壤TP含量主要受土壤母质的影响, 而受外界影响较小[46]。
本研究还发现, 相比于对照, N添加显著降低了各粒级团聚体AP含量与P有效率(图 4, 表 1), 这一作用也表现在图 6中0—10 cm土壤层团聚体AN与AP, AN与P有效率, AP与N有效率, N有效率与P有效率的负相关关系中;且由于团聚体SOC含量与AN、N有效率呈显著正相关, 所以SOC与P有效性也呈显著负相关关系。这是因为高浓度N添加增加了土壤团聚体AN的含量, 促进了植物生产力的提高, 土壤所提供的P不足以平衡增加的N, 植物需要吸收更多的P才能达到养分平衡[47]。N添加对团聚体P有效性的削弱还可能与土壤酶活性、土壤微生物、植物根系分泌物等因素有关, 具体原因有待进一步研究。
4 结论本研究依托全球营养网络中国科学院巴音布鲁克草原生态系统研究站, 开展短期(4年)氮磷添加对高寒草原土壤团聚体结构及其养分含量的影响研究, 得出以下结论:巴音布鲁克高寒草原土壤水稳定性团聚体整体以>2 mm粒级占主导, 有利于维持土壤结构的稳定;氮磷添加对土壤团聚体粒级组成和稳定性影响不同, N添加主要作用于0—10 cm土壤层, 而P添加则主要作用于10—20 cm土壤层;0—10 cm土壤层各粒级团聚体SOC含量显著高于10—20 cm土壤层;N添加加速了SOC矿化分解, 降低了0—10 cm土壤层>2 mm团聚体比例和团聚体稳定性, 但增加了植物生物量, 导致SOC含量没有显著变化;N添加驱动了土壤N元素和P元素的再分配, 分别显著提高和降低了土壤团聚体AN和AP的含量, P添加总体上对团聚体养分影响不显著。
| [1] |
Leff J W, Jones S E, Prober S M, Barberán A, Borer E T, Firn J L, Harpole W S, Hobbie S E, Hofmockel K S, Knops J M H, McCulley R L, La Pierre K, Risch A C, Seabloom E W, Schütz M, Steenbock C, Stevens C J, Fierer N. Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(35): 10967-10972. DOI:10.1073/pnas.1508382112 |
| [2] |
Orwin K H, Ostle N, Wilby A, Bardgett R D. Effects of species evenness and dominant species identity on multiple ecosystem functions in model grassland communities. Oecologia, 2014, 174(3): 979-992. DOI:10.1007/s00442-013-2814-5 |
| [3] |
Zhang X R, Zhang W Q, Sai X, Chun F, Li X J, Lu X X, Wang H R. Grazing altered soil aggregates, nutrients and enzyme activities in a Stipa kirschnii steppe of Inner Mongolia. Soil and Tillage Research, 2022, 219: 105327. DOI:10.1016/j.still.2022.105327 |
| [4] |
Kemper W D. Aggregate stability and size distributions. Methods of Soil Analysis, Part Ⅰ. Physical and Mineralogical Methods-Agronomy Monograph no 9(2nd Edition), 1986: 425-442.
|
| [5] |
Peng X H, Zhu Q H, Zhang Z B, Hallett P D. Combined turnover of carbon and soil aggregates using rare earth oxides and isotopically labelled carbon as tracers. Soil Biology and Biochemistry, 2017, 109: 81-94. DOI:10.1016/j.soilbio.2017.02.002 |
| [6] |
陆琪, 马红彬, 俞鸿千, 王丽, 沈艳, 许冬梅, 谢应忠. 轮牧方式对荒漠草原土壤团聚体及有机碳特征的影响. 应用生态学报, 2019, 30(9): 3028-3038. |
| [7] |
王志康, 祝乐, 许晨阳, 李艳, 耿增超, 王强, 刘莉丽, 秦一郎, 杜旭光. 秦岭天然林凋落物去除对土壤团聚体稳定性及细根分布的影响. 生态学报, 2022, 42(13): 5493-5503. |
| [8] |
Liu D D, Ju W L, Jin X L, Li M D, Shen G T, Duan C J, Guo L, Liu Y Y, Zhao W, Fang L C. Associated soil aggregate nutrients and controlling factors on aggregate stability in semiarid grassland under different grazing prohibition timeframes. Science of the Total Environment, 2021, 777: 146104. DOI:10.1016/j.scitotenv.2021.146104 |
| [9] |
Zhu J X, Wang Q F, He N P, Smith M D, Elser J J, Du J Q, Yuan G F, Yu G R, Yu Q. Imbalanced atmospheric nitrogen and phosphorus depositions in China: implications for nutrient limitation. Journal of Geophysical Research: Biogeosciences, 2016, 121(6): 1605-1616. DOI:10.1002/2016JG003393 |
| [10] |
郭润泉, 熊德成, 宋涛涛, 蔡瑛莹, 陈廷廷, 陈望远, 郑欣, 陈光水. 模拟氮沉降对杉木幼苗细根化学计量学特征的影响. 生态学报, 2018, 38(17): 6101-6110. |
| [11] |
Smil V. Phosphorus in the environment: natural flows and human interferences. Annual Review of Energy and the Environment, 2000, 25: 53-88. DOI:10.1146/annurev.energy.25.1.53 |
| [12] |
凌小莉, 史宝库, 崔海莹, 宋文政, 孙伟. 氮磷添加对松嫩草地土壤团聚体结构及其碳含量的影响. 中国草地学报, 2021, 43(2): 54-63. |
| [13] |
Zhong X L, Li J T, Li X J, Ye Y C, Liu S S, Hallett P D, Ogden M R, Naveed M. Physical protection by soil aggregates stabilizes soil organic carbon under simulated N deposition in a subtropical forest of China. Geoderma, 2017, 285: 323-332. DOI:10.1016/j.geoderma.2016.09.026 |
| [14] |
Yu G R, Jia Y L, He N P, Zhu J X, Chen Z, Wang Q F, Piao S L, Liu X J, He H L, Guo X B, Wen Z, Li P, Ding G A, Goulding K. Stabilization of atmospheric nitrogen deposition in China over the past decade. Nature Geoscience, 2019, 12(6): 424-429. DOI:10.1038/s41561-019-0352-4 |
| [15] |
Yuan X, Qin W K, Xu H, Zhang Z H, Zhou H K, Zhu B. Sensitivity of soil carbon dynamics to nitrogen and phosphorus enrichment in an alpine meadow. Soil Biology and Biochemistry, 2020, 150: 107984. DOI:10.1016/j.soilbio.2020.107984 |
| [16] |
Wilson G W T, Rice C W, Rillig M C, Springer A, Hartnett D C. Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: results from long-term field experiments. Ecology Letters, 2009, 12(5): 452-461. DOI:10.1111/j.1461-0248.2009.01303.x |
| [17] |
李明, 秦洁, 红雨, 杨殿林, 周广帆, 王宇, 王丽娟. 氮素添加对贝加尔针茅草原土壤团聚体碳、氮和磷生态化学计量学特征的影响. 草业学报, 2019, 28(12): 29-40. DOI:10.11686/cyxb2019297 |
| [18] |
Egan G, Crawley M J, Fornara D A. Effects of long-term grassland management on the carbon and nitrogen pools of different soil aggregate fractions. Science of the Total Environment, 2018, 613-614: 810-819. DOI:10.1016/j.scitotenv.2017.09.165 |
| [19] |
Zeng D H, Li L J, Fahey T J, Yu Z Y, Fan Z P, Chen F S. Effects of nitrogen addition on vegetation and ecosystem carbon in a semi-arid grassland. Biogeochemistry, 2010, 98(1-3): 185-193. DOI:10.1007/s10533-009-9385-x |
| [20] |
Zhang L Q, Wei X R, Hao M D, Zhang M. Changes in aggregate-associated organic carbon and nitrogen after 27 years of fertilization in a dryland alfalfa grassland on the Loess Plateau of China. Journal of Arid Land, 2015, 7(4): 429-437. DOI:10.1007/s40333-015-0003-6 |
| [21] |
Zhang H Y, Niu L A, Hu K L, Hao J M, Li F, Wang X, Chen H. Long-term effects of nitrogen and phosphorus fertilization on soil aggregate stability and aggregate-associated carbon and nitrogen in the North China Plain. Soil Science Society of America Journal, 2020, 85(3): 732-745. |
| [22] |
李春越, 常顺, 钟凡心, 薛英龙, 苗雨, 王益, 党廷辉. 种植模式和施肥对黄土旱塬农田土壤团聚体及其碳分布的影响. 应用生态学报, 2021, 32(1): 191-200. |
| [23] |
Huang J S, Liu L L, Qi K B, Yang T H, Yang B, Bao W K, Pang X Y. Differential mechanisms drive changes in soil C pools under N and P enrichment in a subalpine spruce plantation. Geoderma, 2019, 340: 213-223. DOI:10.1016/j.geoderma.2019.01.027 |
| [24] |
张欣, 任海燕, 康静, 朱毅, 李冬慧, 韩国栋. 增温和施氮对内蒙古荒漠草原土壤理化性质的影响. 中国草地学报, 2021, 43(6): 17-24. |
| [25] |
吴建国, 李璇, 吾买尔·吾守. 新疆巴音布鲁克高寒草原毒害草马先蒿的危害与防除效果评价. 新疆畜牧业, 2019, 34(6): 44-46. |
| [26] |
阿依敏·波拉提, 安沙舟, 董乙强, 杨娇, 张爱宁. 巴音布鲁克高寒草原不同退化程度土壤化学计量比特征. 新疆农业科学, 2018, 55(5): 957-965. |
| [27] |
陈丽, 田新民, 任正炜, 董六文, 谢晨笛, 周小龙. 养分添加对天山高寒草地植物多样性和地上生物量的影响. 植物生态学报, 2022, 46(3): 280-289. |
| [28] |
Elliott E T. Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils. Soil Science Society of America Journal, 1986, 50(3): 627-633. |
| [29] |
鲍士旦. 土壤农化分析 (3版). 北京: 中国农业出版社, 2000.
|
| [30] |
刘放, 吴明辉, 魏培洁, 贾映兰, 陈生云. 疏勒河源高寒草甸土壤微生物生物量碳氮变化特征. 生态学报, 2020, 40(18): 6416-6426. |
| [31] |
Beare M H, Hendrix P F, Coleman D C. Water-stable aggregates and organic matter fractions in conventional- and no-tillage soils. Soil Science Society of America Journal, 1994, 58(3): 777-786. |
| [32] |
朱孟涛, 刘秀霞, 王佳盟, 刘志伟, 郑聚锋, 卞荣军, 王艮梅, 张旭辉, 李恋卿, 潘根兴. 生物质炭对水稻土团聚体微生物多样性的影响. 生态学报, 2020, 40(5): 1505-1516. |
| [33] |
Ding Y X, Chen J L. Effect of continuous plantation of Chinese fir on soil fertility. Pedosphere, 1995, 5(1): 57-66. |
| [34] |
马盼盼. 退化高寒草地土壤团聚体稳定性及其养分特征[D]. 兰州: 兰州大学, 2019.
|
| [35] |
郭虎波, 袁颖红, 吴建平, 樊后保, 刘文飞, 黄荣珍, 房焕英. 氮沉降对杉木人工林土壤团聚体及其有机碳分布的影响. 水土保持学报, 2013, 27(4): 268-272. |
| [36] |
Du J X, Liu K L, Huang J, Han T F, Zhang L, Anthonio C K, Shah A, Khan M N, Qaswar M, Abbas M, Huang Q H, Xu Y M, Zhang H M. Organic carbon distribution and soil aggregate stability in response to long-term phosphorus addition in different land-use types. Soil and Tillage Research, 2022, 215: 105195. |
| [37] |
Fisk M, Santangelo S, Minick K. Carbon mineralization is promoted by phosphorus and reduced by nitrogen addition in the organic horizon of northern hardwood forests. Soil Biology and Biochemistry, 2015, 81: 212-218. |
| [38] |
Wang Q K, Wang Y P, Wang S L, He T X, Liu L. Fresh carbon and nitrogen inputs alter organic carbon mineralization and microbial community in forest deep soil layers. Soil Biology and Biochemistry, 2014, 72: 145-151. |
| [39] |
Su Y, Ma X F, Le J J, Li K H, Han W X, Liu X J. Decoupling of nitrogen and phosphorus in dominant grass species in response to long-term nitrogen addition in an Alpine Grassland in Central Asia. Plant Ecology, 2021, 222(2): 261-274. DOI:10.1007/s11258-020-01103-3 |
| [40] |
Ye C L, Chen D M, Hall S J, Pan S, Yan X B, Bai T S, Guo H, Zhang Y, Bai Y F, Hu S J. Reconciling multiple impacts of nitrogen enrichment on soil carbon: plant, microbial and geochemical controls. Ecology Letters, 2018, 21(8): 1162-1173. |
| [41] |
Li J H, Hou Y L, Zhang S X, Li W J, Xu D H, Knops J M H, Shi X M. Fertilization with nitrogen and/or phosphorus lowers soil organic carbon sequestration in alpine meadows. Land Degradation and Development, 2017, 29(6): 1634-1641. |
| [42] |
Albaugh T J, Lee A H, Dougherty P M, Kress L W, King J S. Leaf area and above- and belowground growth responses of loblolly pine to nutrient and water additions. Forest Science, 1998, 44(2): 317-328. |
| [43] |
杨佳, 柯立, 崔珺, 徐小牛. 亚热带常绿阔叶林土壤溶解性有机碳和微生物量碳对氮磷添加的响应. 土壤通报, 2014, 45(4): 876-883. |
| [44] |
岳泽伟, 李向义, 李磊, 林丽莎, 刘波, 曾凡江. 氮添加对昆仑山高山草地土壤、微生物和植物生态化学计量特征的影响. 生态科学, 2020, 39(3): 1-8. |
| [45] |
Yang K, Zhu J J, Gu J C, Yu L Z, Wang Z Q. Changes in soil phosphorus fractions after 9 years of continuous nitrogen addition in a Larix gmelinii plantation. Annals of Forest Science, 2015, 72(4): 435-442. |
| [46] |
刘红梅, 李洁, 王丽丽, 赵建宁, 王慧, 杨殿林. 氮添加对贝加尔针茅草原植物和土壤化学计量特征的影响. 草业学报, 2018, 27(7): 25-35. |
| [47] |
李焕茹, 朱莹, 田纪辉, 魏锴, 陈振华, 陈利军. 碳氮添加对草地土壤有机碳氮磷含量及相关酶活性的影响. 应用生态学报, 2018, 29(8): 2470-2476. |