 2023, Vol. 43
2023, Vol. 43文章信息
- 王志韬, 任珩, 辛存林
- WANG Zhitao, REN Heng, XIN Cunlin
- 不同降水条件下沙鞭种群非生殖株丛空间格局分析
- Spatial pattern of Psammochloa villosa asexual cluster populations under different precipitation conditions
- 生态学报. 2023, 43(10): 4121-4129
- Acta Ecologica Sinica. 2023, 43(10): 4121-4129
- http://dx.doi.org/10.5846/stxb202204291199
-
文章历史
- 收稿日期: 2022-04-29
- 采用日期: 2023-03-07





2. 中国科学院西北生态环境资源研究院, 兰州 730000;
3. 中国科学院临泽内陆河流域研究站, 兰州 730000;
4. 甘肃省绿洲资源环境与可持续发展重点实验室, 兰州 730070
2. Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China;
3. Linze Inland River Basin Research Station, Chinese Academy of Sciences, Lanzhou 730000, China;
4. Key Laboratory of Resource Environment and Sustainable Development of Oasis, Gansu Province, Lanzhou 730070, China
种群空间格局指种群个体在水平空间的分布状态, 是受自身、外界多种因素影响的综合结果, 一直是生态学研究的热点和基本问题之一[1—4]。对于植物种群而言, 形成种群分布格局最常见原因之一是物种生活史差异导致的生长权衡策略的不同[5]。在长期的自然选择下, 植物必须权衡资源分配, 不同功能或器官获得的资源和空间都是权衡分配的综合结果, 进而表现出生活史对策差异性[6—7]。克隆植物兼具有性与无性两种繁殖方式[8], 其无性繁殖方式是极端条件下种群空间拓展的关键, 是确保植物种群资源保存、种群延续的重要繁殖方式[9]。降水作为干旱半干旱地区限制性资源因子, 对于植物种群结构、功能及空间分布有着决定性作用[10—11]。不同降水梯度下, 植物种群非生殖株丛的物质积累和对应器官的资源获取必然受到降水的极大影响, 从而通过改变资源获取方式和策略来协调无性繁殖过程、生存与生长间的协同进化关系以适应生境条件[12]。因此, 开展植物种群非生殖株丛空间分布格局研究能够揭示物种无性繁殖过程在异质生境中资源配置的更新机理, 对于揭示植物种群应对干旱环境的生活史特征和响应机制具有重要意义。
沙鞭(Psammochloa villosa)是典型的旱生、沙生根茎型克隆植物, 对流动沙丘具很强的适应性, 为沙地植物群聚的优势种, 主要分布在内蒙古库布齐沙漠、浑善达克沙地西部、巴丹吉林沙漠和甘肃河西走廊沙地等地区[13]。沙鞭植株能够通过地下根茎无性繁殖方式, 拓展风蚀形成的裸沙斑块[14], 开启裸沙斑块上的植被群落演替, 对沙区景观具有明显的改造作用[15—16]。目前, 关于沙鞭的研究主要包括种子休眠和萌发策略及对沙埋和种子大小的响应[17—18]与克隆繁殖[15, 19]、等位酶变异[20]、形态变异[21]、遗传多样性[22]、固沙能力[23]、AM真菌群落结构和多样性[24], 极大地丰富和发展了沙鞭的相关研究, 但是关于不同降水梯度沙鞭的自适应策略和内在机制的研究尚未涉及。基于此, 本研究在实地调查我国主要沙鞭生长生境和群落学特征基础上, 选择不同降水条件下的沙鞭种群, 通过基于完全随机、泊松聚块和嵌套双聚块模型的点格局方法开展种群非生殖株丛空间分布格局研究, 旨在研究沙鞭种群非生殖株丛空间点位特征和无性繁殖过程及其影响因素, 为防沙治沙提供科学理论支持。
1 材料与方法 1.1 研究区概况根据中国气象数据网数据划分不同降水区, 在生长有沙鞭且地形地貌和植被分布稀疏程度相对一致的流动沙丘, 依据沙地群落学调查和降水梯度情况, 选择内蒙古自治区阿拉善右旗巴丹镇、内蒙古自治区阿拉善左旗吉兰泰镇、甘肃省临泽县平川镇、内蒙古自治区杭锦旗独贵塔拉镇、宁夏回族自治区沙坡头区迎水桥镇、内蒙古自治区准格尔旗大路镇等6个地区, 开展不同降水条件下沙鞭种群非生殖株丛空间分布格局研究(图 1)。研究区各样地相关信息如表 1所示。

|
| 图 1 研究区位置图 Fig. 1 The location of the study area |
| 样地 Plot |
位置 Location |
海拔 Altitude/m |
降水量 Precipitation/(mm/a) |
经纬度 Latitude and longitude |
自然景 观类型 Landscape type |
丰富度 Richness |
群落Community | |
| 优势种 Dominant species |
伴生种 Companion species |
|||||||
| A | 内蒙古自治区阿拉善右旗巴丹镇 | 1229 | 89 | 102°24′00″E, 39°32′31.92″N |
沙漠 | 3 | 沙鞭 | 沙米, 沙蒿 |
| B | 内蒙古自治区阿拉善左旗吉兰泰镇 | 979 | 107.8 | 105°42′55.44″E, 39°40′36.12″N |
荒漠戈壁 | 6 | 沙鞭 | 沙米, 沙蒿, 泡泡刺, 沙拐枣, 梭梭 |
| C | 甘肃省临泽县平川镇 | 1229 | 117.4 | 100°39′00″E, 39°13′19.92″N |
荒漠绿洲 | 3 | 沙鞭 | 沙米, 沙蒿 |
| D | 内蒙古自治区杭锦旗独贵塔拉镇 | 1076 | 186 | 108°39′1.79″E, 40°25′58.44″N |
荒漠绿洲 | 4 | 沙鞭 | 沙米, 沙蒿, 鹅绒藤 |
| E | 宁夏回族自治区沙坡头区迎水桥镇 | 1265 | 191.1 | 105°32′7.79″E, 37°31′17.04″N |
沙漠 | 5 | 沙鞭 | 沙米, 沙蒿, 虫实, 雾冰藜 |
| F | 内蒙古自治区准格尔旗大路镇 | 985 | 363 | 111°12′15.48″E, 40°10′41.87″N |
荒漠草原 | 5 | 沙鞭 | 沙米, 沙蒿, 虫实, 苦菜 |
本研究于2021年5月开展沙鞭主要生长区域降水、温度等气候条件调研, 2021年6月植物生长盛期在沙鞭主要生长区开展野外调查, 包含沙鞭群落、种群空间分布格局调查等。其中, 设置6个空间格局调查样地, 分别在每个调查样地内随机设置3个4 m×4 m的样方, 共计18个样方, 实测并记录每个样方内的情况, 以样方框左下角顶点为基点, 用1 m×1 m的样方框按从左到右, 从上到下的顺序依次放置16次进行沙鞭非生殖株点格局调查。记录样方中每个沙鞭非生殖株丛的地面投影和相对位置, 用坐标值直接表示距离。室内将每一个样方按照取样时样方框放置顺序进行拼接、合并, 统计每个样方内沙鞭非生殖株丛数、高度、茎高。
1.3 研究方法 1.3.1 点格局分析处于研究区域中, 在二维空间的点构成点事件, 以k(r)函数揭示点事件空间布局的依赖程度[25], 定义为:
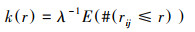
|
(1) |
式中, λ代表着研究区域中每个单位面积点的事件密度, E代表着在一定的距离尺度中点事件的数量期望, #代表数量, i、j代表为研究区域中两个拥有相同特征的点事件, rij代表了某个点与其它点之间的距离, r则为距离尺度。如果假定在一定的距离尺度中, 点事件是随机分布的, 则有:
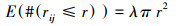
|
(2) |
那么此时, k(r)=πr2。在点事件处于聚集分布时, 如果距离尺度比较小, 则k(r)的观测值就会大于πr2。估计式为:

|
(3) |
式中, A代表着研究区域的面积, 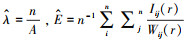
一般情况下在一定尺度中点事件的分布类型是以L(r)函数的估计值来确定的。在距离尺度r中, 如果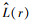
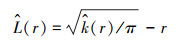
|
(4) |
选择合适的零模型是顺利使用Ripley′s K函数运算所需解决的生态学问题的关键, 同时可以完善地解释实测数据和零模型的偏离。为了揭示降水梯度下沙鞭种群非生殖株丛空间分布格局的分布特征, 本研究选择了完全空间随机模型、泊松聚块模型和嵌套双聚块模型。
(1) 完全空间随机模型
完全空间随机模型(CSR)在点格局中是最简单、最常用的一种, 其实质是均质泊松过程(homogeneous Poissonprocess, HP)。在此模型所选择的研究范围中, 任意一点(或个体)出现(或发生)在任意一个位置上的机会是均等的;同时, 点(或个体)与点间相互独立, 即任意两点间均不发生相互作用。物种的空间分布格局常以其作为零假设(nulltest)来检验[26], CSR模型K函数同上文定义。
(2) 泊松聚块模型
泊松聚块模型(NS), 也可称为均质托马斯过程(homogeneous Thomas process, TH), 是对一种聚块机制的描述。该模型假设母体事件以完全空间随机过程发生, 而每一个母体周围以一定的概率分布产生随机数量的子代个体, 同时这些子代个体的空间分布遵循某双变量概率密度函数。如果子代个体的数量遵循泊松分布并且其在空间的位置相对于母体符合双变量高斯分布, 那么子代个体的发生就符合泊松聚块过程[25, 27—28]。泊松聚块模型的K函数表达式如下:
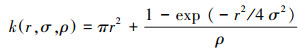
|
(5) |
式中, r为尺度, ρ为该过程中母体的密度, σ2为高斯分布的方差。
(3) 嵌套双聚块模型
嵌套双聚块模型(Nested double-cluster process, DC)是泊松聚块模型的多代扩展, 泊松聚块过程的子代产生自己的后代个体, 若未偏离泊松聚块模型则嵌套双聚块模型不用再做。详细描述参考以下文献[26, 29—30], 嵌套双聚块模型的K函数表达式如下:

|
(6) |
且, rsum2=σ1+σ2
而参数r、ρ和σ2的意义同泊松聚块模型, 其中, 下角标1代表第1代, 下脚标2代表第2代[31—33]。
1.4 数据处理扫描野外调查实际观测图, 利用GetData Graph Digitizer 2.22软件提取植物株丛实际坐标值, 将坐标值录入Excel软件, 应用Thorsten Wiegand的ProgramitaEnero2019软件中进行种间空间格局及其关联分析。本文中采用的步长为1 cm, Monte Carlo随机模拟的数目为199次, 得到上下两条包迹线围成的95%置信区间。
2 结果与分析 2.1 沙鞭种群非生殖株丛生物学特征沙鞭种群非生殖株丛生物学特征在不同降水条件下表现出不同趋势(表 2)。降水相对充足区域(样地D、E、F), 沙鞭非生殖株丛数表现出随降水量增加而增加的趋势, 其他样地(样地A、B、C)则未表现出明显规律。高度和盖度表现出与降水量不一致的趋势。
| 样地 Plot |
总株丛数 Total number/株 |
总盖度 Total Coverage/% |
总高度 Total height/cm |
非生殖株丛Asexual cluster | |
| 株丛数Number/株 | 高度Height/cm | ||||
| A | 255±33d | 38.73±5.01a | 86.35±29.13bc | 216±24c | 76.14±14.17bc |
| B | 248±30e | 35.4±4.22b | 90.44±28.06b | 194±21d | 78.21±13.19b |
| C | 468±61a | 32.87±4.28b | 83.07±29.25cd | 415±47a | 74.77±16.64c |
| D | 288±35c | 38.6±5.01a | 101.52±38.85a | 214±25c | 81.21±14.20a |
| E | 251±32de | 26.15±3.39c | 80.02±17.61d | 218±23c | 75.00±11.11c |
| F | 324±40b | 34.50±4.48ab | 91.02±27.39b | 272±28b | 81.46±15.65a |
| *总株丛数、总盖度、总高度, 非生殖株丛株丛数、高度显著性差异(P<0.05) | |||||
从图 2可见, 不同降水量地区中, 沙鞭种群非生殖株丛在不同尺度上空间点格局特征存在差异。A样地平均年降水量89 mm/a, 在3—126 cm尺度上呈聚集分布;B样地平均年降水量107.8 mm/a, 在9—200 cm尺度上呈聚集分布;C样地平均年降水量117.4 mm/a, 在9—53 cm、59—73 cm、83—129 cm尺度上呈聚集分布;D样地平均年降水量186 mm/a, 在6—77 cm尺度上呈聚集分布;E样地平均年降水量191.1 mm/a, 在2—95 cm尺度上呈聚集分布;F样地平均年降水量363 mm/a, 在2—96 cm尺度上呈聚集分布。

|
| 图 2 沙鞭种群非生殖株丛基于完全空间随机模型的点格局分析 Fig. 2 Point pattern analysis of Psammochloa villosa asexual cluster based on completel spatial randomness 图中A—F代表样地; A. 内蒙古自治区阿拉善右旗巴丹镇;B. 内蒙古自治区阿拉善左旗吉兰泰镇;C. 甘肃省临泽县平川镇;D. 内蒙古自治区杭锦旗独贵塔拉镇;E. 宁夏回族自治区沙坡头区迎水桥镇;F. 内蒙古自治区准格尔旗大路镇 |
从图 3可见, 6个样地均有偏离完全随机模型的现象, 对6个样地进行泊松聚块模型点格局分析发现:AD样地在0—200 cm尺度上符合泊松聚块模型;B样地在15—19 cm尺度上正向偏离泊松聚块模型, 在0—14 cm与20—200 cm尺度上符合泊松聚块模型;C样地在2—6 cm尺度上正向偏离泊松聚块模型, 在0—1 cm与7—200 cm尺度上符合泊松聚块模型;E样地在2—4 cm尺度上正向偏离泊松聚块模型, 在0—1 cm与5—200 cm尺度上符合泊松聚块模型;F样地在9—25 cm尺度上正向偏离泊松聚块模型, 在0—8 cm与26—200 cm尺度上符合泊松聚块模型(图 3)。

|
| 图 3 沙鞭种群非生殖株丛基于泊松聚块模型的点格局分析 Fig. 3 Point pattern analysis of Psammochloa villosa asexual cluster based on poisson cluster process |
对BCEF两样地进一步进行嵌套双聚块模型模拟, 结果显示, 在BCEF四样地, 沙鞭种群非生殖株丛均在0—200 cm整个尺度上符合嵌套双聚块模型(图 4)。

|
| 图 4 沙鞭种群非生殖株丛基于嵌套双聚块模型的点格局分析 Fig. 4 Point pattern analysis of Psammochloa villosa asexual cluster based on nested double cluster process |
植物种群空间分布格局研究在过去几十年不断加强和深入, 因其与种群的生态过程紧密相连, 能够从当前的空间格局推断种群个体在空间上的形成过程[4]。目前使用的各种方法模型中, 最重要且常见的为完全空间随机模型, 可以将种群空间分布格局分为聚集分布、均匀分布和随机分布等3个类型, 聚集分布(正向偏离)代表株丛抱团获取相关资源, 均匀分布(负向偏离)代表株丛间存在竞争关系, 偏离尺度越大代表株丛间关系在更大范围内表现出对应关系, 对于认识植被空间分布的机理具有一定的价值和作用。
植物种群空间分布格局能够显示出某些环境因子对于植物个体行为、生长以及生存的影响[34]。根据降水尺度以及分析结果, 分为AC与DEF两组进行讨论, 从基于完全空间随机模型的沙鞭种群非生殖株丛点格局分析结果来看, DEF样地在不同降水环境下沙鞭非生殖株丛呈现随降水量增加聚集尺度逐渐增大的规律, 在6—77 cm至2—96 cm尺度范围内表现为聚集分布, 与沙鞭生殖株丛随降水量变化特征相吻合[35]。这与对沙鞭的群落学调查结果表现一致(表 2), 即随降水量增加, 株丛数由214株上升至272株。造成这种现象的原因可能是随降水量增加, 群落丰富度达到4—5, 种间竞争加剧, 沙鞭非生殖株丛间通过相互协作增强种间竞争能力, 实现水资源争夺与群落空间占据目的。同时, 相较于生殖株丛, 非生殖株丛能依靠母体的供养更容易安全地度过幼龄期, 成功定居率相对较高[36], 这种较大尺度聚集格局可能是种内老、幼株丛间相互协作、共同分配资源的结果。AC样地表现出不一致的规律, 即AC样地虽然降水量小于DEF样地, 但空间分布聚集尺度却大于DEF样地。造成这个现象的原因可能是由于AC样地丰富度仅为3, 且伴生物种为沙蒿、沙米等草本植物, 种间竞争相对较弱, 株丛间的大尺度协作有助于种群争夺群落内水分和空间, 进而确保沙鞭株丛能够在极端干旱地区存活。沙鞭非生殖株丛种群采用株丛间协作的策略, 即“抱团”来共同抵御其他植物种群给予的竞争压力, 及克服极端干旱地区水资源匮乏的生存环境, 从而达到种内个体资源与风险互相分摊, 有利于种群延续和拓展。
B样地降水量107.8mm的地区, 聚集尺度达到9—200 cm, 表现出不一致的格局特征, 这可能是由于气候、生物等多种因素影响造成的。一是, B样地, 伴生物种最多, 丰富度达到6, 是所有样地中的最大值, 且伴生物种包括泡泡刺、沙拐枣、梭梭等灌木物种, 群落内种间竞争较其他样地大, 加剧了沙鞭非生殖株丛在大尺度范围争夺资源的压力;二是, B样地自然景观类型为荒漠戈壁, 样地内分布大量的碎石块, 不利于沙鞭种群株丛的生长, 株丛间的协作有利于个体存活和种群延续。野外调研过程中, 也发现沙鞭株丛存在死亡现象, 株丛分布较为分散, 沙鞭种内关系主要为个体间协作。因此, B样地出现降水量相对较低, 但聚集尺度较大的现象。除了降水因素外, 植物种群空间格局差异可能与不同样地的海拔、温度、地形等因素密切相关, 尽管样地选择过程中尽可能减少相关因素影响, 但仍不容忽视, 需要进一步开展其他因素对沙鞭生长的影响研究。
基于完全随机模型的点格局分析, 6个样地均存在不同尺度的聚集分布格局, 需要进一步采取基于泊松聚块模型的沙鞭种群非生殖株丛点格局分析。分析结果显示, BCEF四个样地中, 沙鞭非生殖株丛分别在15—19 cm、2—6 cm、2—4 cm、9—25 cm尺度上正向偏离泊松聚块模型而符合基于嵌套双聚块模型的空间分布格局, 表明沙鞭非生殖株丛在四个样地中的大聚块中存在密度较高的小聚块。造成这种原因可能是由于沙鞭种群有别于其它物种的无性拓展方式有关。沙鞭种群地下根茎沿延伸方向每间隔一定距离形成节(间隔子), 在节(间隔子)处会生长芽(根茎芽或分株芽), 芽横向生长则形成根茎芽, 向上生长突破沙层则形成分株芽。这种生长模式与羊草等物种不一致[37], 根茎芽会沿着新的拓展方向向外拓展, 实现占据空间和争夺资源的目的, 但分株芽则向上生长, 逐渐生成新的地上分株以及分株系统。野外调查时观测到, 沙鞭拓展过程中, 沙鞭根茎节的间隔长度会随着资源丰富程度表现出不一致的现象, 当资源丰富区域, 间隔尺度明显缩小, 最小达到3—6cm, 而资源不丰富区域, 间隔尺度则可能达到10—15cm, 甚至距离更大, 因此, 这种独特的无性拓展特征可能是造成小尺度上形成小聚块的原因之一。此外, 沙鞭极易出现簇状或密集的生成地上分株的现象, 呈现出“爆发式”地上分株生成模式[38], 这也可能是小尺度上形成小聚块的关键因素。但是, 沙鞭“爆发式”地上分株生成机制尚不明晰, 需要进一步开展相关研究。
4 结论本研究利用群落学调查和基于完全随机、泊松聚块和嵌套双聚块模型的点格局方法, 分析了沙鞭种群非生殖株丛空间分布格局特征, 探讨了空间格局对降水梯度的响应和适应机制。主要结论为:
(1) 随降水量上升, 沙鞭非生殖株丛种群采取扩张策略, 大量抢占群落空斑为植物种群谋取更多资源和空间, 在空间格局上表现出聚集尺度逐渐增大的趋势;
(2) 沙鞭种群非生殖株丛可能会在较小尺度上形成小聚块的空间分布特征, 这可能与其繁殖方式有关。
| [1] |
任珩, 赵成章, 高福元, 石福习, 张茜. 退化草地阿尔泰针茅生殖株丛与非生殖株丛的空间格局. 生态学报, 2012, 32(21): 6909-6916. |
| [2] |
李源, 李海兵, 姜超, 邰阳, 刘丹辉, 张维华, 王鑫厅. 典型草原不同放牧强度下羊草种群点格局分析. 内蒙古大学学报: 自然科学版, 2021, 52(4): 417-424. |
| [3] |
顾荣, 张彩彩, 和正华, 杨荣, 陈瑶, 冯萍, 斯那取宗, 赵冬莲, 益西央初, 吴俊华, 林露湘. 香格里拉普达措国家公园长苞冷杉种群空间分布格局及关联性. 生态学杂志, 2021, 40(12): 3860-3869. |
| [4] |
王鑫厅, 王殿杰, 李海兵, 邰阳, 姜超, 刘芳, 李素英, 苗百岭. 点格局研究过程中K-函数的累积效应. 应用生态学报, 2022, 33(5): 1275-1282. |
| [5] |
Kneitel J M, Chase J M. Trade-offs in community ecology: linking spatial scales and species coexistence. Ecology Letters, 2004, 7(1): 69-80. DOI:10.1046/j.1461-0248.2003.00551.x |
| [6] |
张景光, 王新平, 李新荣, 张志山, 王刚, 王桑, 马风云. 荒漠植物生活史对策研究进展与展望. 中国沙漠, 2005, 25(3): 306-314. DOI:10.3321/j.issn:1000-694X.2005.03.002 |
| [7] |
陈磊, 叶其刚, 潘丽珠, 徐立铭, 黄宏文. 长江中下游湖泊两种混生苦草属植物生活史特征与共存分布格局. 植物生态学报, 2008, 32(1): 106-113. DOI:10.3773/j.issn.1005-264x.2008.01.012 |
| [8] |
李卓琳, 穆春生, 吴春英. 气候变化对草原多年生克隆植物无性繁殖的影响. 草业科学, 2017, 34(8): 1694-1704. |
| [9] |
Sutherland W J. Growth and foraging behaviour. Nature, 1987, 330(6143): 18-19. DOI:10.1038/330018a0 |
| [10] |
吴旭东, 季波, 何建龙, 任小玢, 俞鸿千, 王占军. 控制降水梯度对荒漠草原优势植物叶功能性状及土壤养分的影响. 生态学报, 2021, 41(7): 2719-2727. |
| [11] |
郭亚飞, 张丽华, 赵锐锋, 谢忠奎, 王军锋, 蒿廉伊, 高江平, 王梦梦, 高晗. 月尺度视角下荒漠草原植物群落特征对降水变化的响应. 生态学杂志, 2021, 40(7): 1895-1906. |
| [12] |
王颖, 蔺吉祥, 丁雪梅, 张娜, 韩相奎. 降水格局变化对植物繁殖策略影响的研究进展. 中国农学通报, 2014, 30(36): 253-256. DOI:10.11924/j.issn.1000-6850.2014-1623 |
| [13] |
中国科学院兰州沙漠研究所. 中国沙漠植物志(第一卷). 北京: 科学出版社, 1985.
|
| [14] |
Yu F H, Dong M, Krüsi B. Clonal integration helps Psammochloa villosa survive sand burial in an inland dune. New Phytologist, 2004, 162(3): 697-704. DOI:10.1111/j.1469-8137.2004.01073.x |
| [15] |
董鸣, 阿拉腾宝, 邢雪荣, 王其兵. 根茎禾草沙鞭的克隆基株及分株种群特征. 植物生态学报, 1999, 23(4): 302-310. DOI:10.3321/j.issn:1005-264X.1999.04.002 |
| [16] |
董鸣. 克隆植物生态学. 北京: 科学出版社, 2011.
|
| [17] |
朱雅娟, 董鸣, 黄振英. 沙埋和种子大小对固沙禾草沙鞭的种子萌发与幼苗出土的影响. 植物生态学报, 2005, 29(5): 730-739. |
| [18] |
黄振英. 鄂尔多斯高原固沙禾草沙鞭种子休眠和萌发与环境的关系. 西北植物学报, 2003, 23(7): 72-77. |
| [19] |
董鸣. 切断根茎对根茎禾草沙鞭和赖草克隆生长的影响. 植物学报, 1999, 41(2): 194-198. |
| [20] |
王可青, 葛颂, 董鸣. 根茎禾草沙鞭的等位酶变异及克隆多样性. 植物学报, 1999, 41(5): 88-91. |
| [21] |
吕婷, 刘涛, 梁瑞芳, 苏旭, 刘玉萍. 沙生植物沙鞭不同居群形态变异研究. 植物研究, 2021, 41(1): 60-66. |
| [22] |
吕婷, 刘玉萍, 周勇辉, 刘涛, 张晓宇, 苏旭. 荒漠植物沙鞭(Psammochloa villosa)种质资源收集及谱系遗传分化初探. 草地学报, 2018, 26(3): 733-740. |
| [23] |
陈玉福, 于飞海, 张称意, 董鸣. 根茎禾草沙鞭的克隆生长在毛乌素沙地斑块动态中的作用. 生态学报, 2001, 21(11): 1745-1750. |
| [24] |
贺学礼, 李英鹏, 赵丽莉, 刘雪伟. 毛乌素沙地克隆植物沙鞭生长对AM真菌生态分布的影响. 生态学报, 2010, 30(3): 751-758. |
| [25] |
Diggle P. Statistical analysis of spatial point patterns. London: Academic Press, 1983.
|
| [26] |
Symanzik J. Statistical analysis of spatial point patterns. 2nd ed.. London: Hodder Education Publishers, 2003.
|
| [27] |
Wiegand T, Moloney K A. Rings, circles, and null-models for point pattern analysis in ecology. Oikos, 2004, 104(2): 209-229. |
| [28] |
Batista J L F, A Maguire D. Modeling the spatial structure of topical forests. Forest Ecology and Management, 1998, 110(1/2/3): 293-314. |
| [29] |
Stoyan D, Stoyan H. Estimating pair correlation functions of planar cluster processes. Biometrical Journal, 1996, 38(3): 259-271. |
| [30] |
Watson D M, Roshier D A, Wiegand T. Spatial ecology of a root parasite-from pattern to process. Austral Ecology, 2007, 32(4): 359-369. |
| [31] |
高福元, 赵成章. 甘肃臭草型退化草地优势种群空间格局及其关联性. 生态学报, 2012, 32(21): 6661-6669. |
| [32] |
高福元, 赵成章. 高寒退化草地狼毒种群株丛间格局控制机理. 生态学报, 2013, 33(10): 3114-3121. |
| [33] |
高福元, 石福习. 基于不同零模型的三江平原沼泽湿地主要物种小尺度点格局分析. 生态学报, 2015, 35(7): 2029-2037. |
| [34] |
何志斌, 赵文智. 黑河盆地荒漠过渡带两种主要灌木种群的空间格局. 应用生态学报, 2004, 15(6): 947-952. |
| [35] |
王志韬, 任珩, 辛存林, 张东梅. 不同降水条件下沙鞭种群生殖株丛空间格局. 生态学杂志, 2023, 42(4). DOI:10.13292/j.1000-4890.202303.014 |
| [36] |
张晓娜, 哈达朝鲁, 潘庆民. 刈割干扰下内蒙古草原两种丛生禾草繁殖策略的适应性调节. 植物生态学报, 2010, 34(3): 253-262. |
| [37] |
白乌云. 羊草根茎克隆生长特性种内分化及与原生境地理和气候因素的关联研究[D]. 北京: 中国农业科学院, 2021.
|
| [38] |
黄浩, 温国胜. 毛竹爆发式生长机理的探究. 科技资讯, 2009, 7(31): 218-219. |