 2023, Vol. 43
2023, Vol. 43文章信息
- 曹乃欣, 张飞, 李博, 朱帕尔·巴合拜提, 王启果, 雒涛
- CAO Naixin, ZHANG Fei, LI Bo, JUPAR Bahtebay, WANG Qiguo, LUO Tao
- 准噶尔盆地荒漠景观不同地貌类型鼠、蚤多样性及分布特征
- Study on Diversity and Distribution Characteristics of Rodents and Fleas in Different Landforms of Desert Landscape in Junggar Basin
- 生态学报. 2023, 43(10): 4270-4283
- Acta Ecologica Sinica. 2023, 43(10): 4270-4283
- http://dx.doi.org/10.5846/stxb202204050876
-
文章历史
- 收稿日期: 2022-04-05
- 采用日期: 2023-03-11





2. 新疆大学智慧城市与环境建模自治区普通高校重点实验室, 乌鲁木齐 830017;
3. 新疆维吾尔自治区疾病预防控制中心鼠疫防制科, 乌鲁木齐 830002
2. Key Laboratory of Smart City and Environment Modeling, Xinjiang University, Urumqi 830017, China;
3. Xinjiang Center for Disease Control and Prevention, Urumqi 830002, China
鼠疫是一种致命的细菌性疾病, 通过携带鼠疫耶尔森菌的跳蚤在相关动物间传播[1]。不同种类的鼠疫在不同物种(主要是啮齿动物)之间有不同的传播规律。在一定条件下, 鼠疫通过受感染的鼠疫蚤或其他方式传播给人类。临床上, 表现出淋巴和血管系统炎症以及组织出血等症状[2]。因此, 为预防自然疫源地鼠疫的爆发, 有必要对疫源地内鼠、蚤类型和数量进行调查, 即掌握疫源地内鼠、蚤在不同地区的分布特点。
鼠疫耶尔森菌的进化和宿主动物的繁殖需要特定的地理条件。不同的地理条件很大程度上决定了鼠疫宿主动物的种类和分布特征, 这是鼠疫疫源地形成的基本条件[3—4]。根据鼠疫和地理环境条件关系, 国内外学者进行了大量研究。针对不同区域鼠、蚤问题的相关研究各有特点, 采用的方法和研究角度各不相同。其中, 一部分学者利用传统的生态学方法进行研究, 如:采用群落生态研究方法对准噶尔盆地各鼠体蚤类群落进行群落结构因子的相似性分析[5];也有学者在乌苏市古尔图镇鼠疫监测点对长尾黄鼠和灰旱獭进行密度监测, 并计算不同采样点鼠、蚤生态学指标[6]。部分学者研究不同海拔条件下鼠类和蚤类的空间分布特征, 探究鼠类与蚤类集中分布的海拔高度[7]。还有一部分学者利用地理环境要素来研究, 如:探讨生态学指标受气候因素(如气温和降水)影响, 结果表明不同气候条件下宿主的物种丰富度和总蚤指数存在明显变化[8]。另有学者探究地貌类型对鼠类分布的影响[9]。综上所述, 传统的生态学方法已不能满足当前研究的需求, 基于当前的研究进展, 较少的学者研究不同地貌条件对鼠类和蚤类的影响, 加之地貌类型的分类考虑诸多因素(例如海拔、气候等), 一定程度上增加了研究的难度。因此, 本研究聚焦于不同地貌类型条件下的鼠、蚤指标的对比研究, 并分析其变化规律。
除利用生态学研究方法外, 部分学者也使用GIS空间分析方法探究鼠、蚤空间分布特征, 如:杨琳等使用GPS技术测定鼠疫监测点的坐标, 为分析鼠疫时空变化规律创造了条件, 同时借助GIS的最近邻分析和核密度法对血清监测阳性鼠类的分布特点进行分析[10]。杨林生等利用我国人间鼠疫流行资料, 对比我国南方与北方疫源地分布特点, 总结我国不同地区自然疫源地的特点[11];Zhuang等采用空间自相关和冷热点探测方法来探究不同区域鼠蚤指标的空间集聚特征[12];前人在研究准噶尔盆地鼠疫自然疫源地时, 较少的采用GIS空间分析方法。因此, 本文借助GIS空间分析方法探究准噶尔盆地不同地区鼠蚤指标的差异性, 并分析不同鼠蚤指标的空间集聚现象。
综上, 影响鼠疫发生和流行的因素较多。其中, 不同地貌类型之间的气候特征差异较大, 聚焦于不同地貌类型对鼠、蚤的影响具有重要意义。因此, (1)本研究通过计算鼠类和蚤类生态学指标, 对不同地貌类型的鼠、蚤生态学指标进行相关性分析, 探究不同地貌类型群落的相似性;(2)借助空间自相关和标准差椭圆等空间分析方法, 探究不同鼠、蚤及其生态学指标的空间分布特征。本文为研究准噶尔盆地自然疫源地鼠、蚤空间分布特征提供参考依据。
1 材料与方法 1.1 研究区概况如图 1所示, 本研究以准噶尔盆地为主要研究区, 除此之外还包括盆地所在地的县(市)行政区, 共25个行政区, 行政区名称如表 1所示。准噶尔盆地海拔在400 m左右, 地势东高西低。2005年, 中国新疆准噶尔盆地被发现是新型鼠疫自然疫源地[13], 它与中亚荒漠鼠疫自然疫源地的地理生态景观相似, 属于典型的平原荒漠类型鼠疫自然疫源地[14]。

|
| 图 1 研究区地理位置及采样点分布图 Fig. 1 Geographical location of the study area and distribution of sampling points |
| 地貌类型Geomorphic type | 地名Place name |
| 低海拔湖积平原(Ⅰ) Low elevation lacustrine plain | 阿拉山口市, 精河县, 福海县 |
| 低海拔冲积平原(Ⅱ) Low elevation alluvial plain | 精河县, 克拉玛依区, 乌尔禾区, 玛纳斯县, 呼图壁县, 昌吉市, 阜康市, 奇台县 |
| 低海拔冲积洪积平原(Ⅲ) Low elevation alluvial floodplain | 布尔津县, 额敏县, 哈巴河县, 乌苏市, 托里县, 吉木乃县, 石河子市, 白碱滩区, 五家渠市, 沙湾县 |
| 低海拔风积地貌(Ⅳ) Low altitude aeolian landform | 吉木萨尔县, 米东区 |
| 中海拔湖积平原(Ⅴ) Middle elevation lacustrine plain | 巴里坤哈萨克自治县 |
| 中海拔冲积洪积平原(Ⅵ) Middle elevation alluvial floodplain | 伊吾县 |
| 中海拔洪积平原(Ⅶ) Middle elevation flood plain | 木垒哈萨克自治县 |
| 中海拔剥蚀平原(Ⅷ) Middle elevation denudation plain | 富蕴县 |
采样点位置如图 1和表 1所示, 主要分布在低海拔和中海拔地貌类型上。根据[15]的海拔分级指标, 将本研究区按照海拔进行划分, 海拔小于1000m的区域为低海拔地区, [1000, 2000)m之间为中海拔地区, 大于等于2000m的划分为高海拔地区。地貌类型数据来源于青藏高原科学数据中心(http://data.tpdc.ac.cn/zh-hans/)。由于研究区内以低海拔地貌类型为主(Ⅰ、Ⅱ、Ⅲ、Ⅳ), 因此本研究在荒漠景观内设置采样点时, 低海拔地貌类型布置采样点较多。
1.2.2 数据采集① 标本捕获方法
普通鼠捕获:选定鼠类活动区域并寻找鼠洞, 在洞外和相邻有鼠类活动痕迹的鼠洞布弓形夹, 以花生粒为诱饵, 每人负责20个鼠夹, 选择2—4个鼠洞群布夹, 每0.5—1小时观察捕获情况。夜行鼠类捕获:在选定区域太阳快落山时, 采用5m夹线法布小板夹, 每个区域布夹不少于300夹次, 次日太阳升起前查看捕鼠情况。捕获后将鼠类处死, 装入白色布袋。
② 样本采集及运输
为防止感染各种鼠传病原体, 需要做好个人防护。收放鼠夹时应戴一次性帽子、工作服、手套及可防止气溶胶吸入的口罩。鉴定鼠类标本前需要将所有标本在密闭容器中用乙醚或杀虫剂进行熏蒸10min左右, 防止鼠类体表各种寄生虫逃逸及叮咬。
本研究调查地区为准噶尔盆地所涉及的大多数行政县(市), 缺少的县(市)为奎屯市和独山子区。采样时间为2005—2010年, 调查点位于调查区的荒漠景观中, 在荒漠景观中选择不同地貌类型的调查点。调查点以荒漠景观面积约1%的比例设置, 数量范围为1—5。各测点均按均匀分布原则设置在荒漠景观区内[5]。捕获的鼠类名称如表 2所示, 包括3科9属13种;捕获蚤类名称如表 3所示, 包括5科10属19种。
| 科Family | 属Genus | 种Species |
| 仓鼠科Circetidae | 大沙鼠属Rhombomys | 大沙鼠Rhombomys opimus |
| 沙鼠属Meriones | 红尾沙鼠Meriones libycus, 柽柳沙鼠Meriones tamariscinus | |
| 仓鼠属Cricetus | 灰仓鼠Cricetulus migratorius, 短尾仓鼠Cricetulus eversmanni, 长尾仓鼠Cricetulus longicaudatus |
|
| 鼠科Muridae | 沙鼠属Meriones | 子午沙鼠Meriones meridianus Pallas |
| 鼠属Mustella | 小家鼠Mus musculus | |
| 姬鼠属Apodemus | 小林姬鼠Apodemus sylvaticusLinnaeus | |
| 跳鼠科Dipodidae | 三趾跳鼠属Dipus sagitta | 三趾跳鼠Dipus sagitta |
| 五趾跳鼠属Allactaga | 五趾跳鼠Allactaga sibirica, 小五趾跳鼠Allactaga elater | |
| 长耳跳鼠属Euchoreutes | 长耳跳鼠Euchoreutes naso |
| 科Family | 属Genus | 种Species |
| 蚤科Pulicidae | 客蚤属Xenopsylla cheopis | 簇鬃客蚤Xenopsylla skrjabini, 粗鬃客蚤Xenopsylla hirtipes Rothschild, 臀突客蚤Xenopsylla minax jordan, 同形客蚤指名亚种Conformis conformis |
| 角头蚤属Echidnophaga | 长吻角头蚤Oschanini | |
| 角叶蚤科Ceratophyyllidae | 病蚤属Ceratophyllus | 秃病蚤指名亚种laeviceps, 裂病蚤fidus, 四鬃病蚤Tersus, 土库曼病蚤指名亚种Siphonaptera |
| 细蚤科Leptopsyllidae | 怪蚤属Paradoxopsyllus | 无额突怪蚤Paradoxopsyllus teretifrons, 后弯怪蚤Paradoxopsyllus repandus |
| 眼蚤属Ophthalmopsylla | 长突眼蚤Kiritschenkoi | |
| 中蚤属Mesopsylla | 软中蚤Lenis, 真凶中蚤精河亚种Mesopsylla eucta shikho | |
| 多毛蚤科Hystrichopsyllidae | 狭蚤属Stenoponia | 子午狭蚤Meridiana, 重要狭蚤Conspecta |
| 栉眼蚤属Ctenophthalmus | 修长栉眼蚤指名亚种Dolichus | |
| 纤蚤属Rhadinopsylla | 宽臂纤蚤Rhadinopsylla cedestis | |
| 切唇蚤科Coptopsyllidae | 切唇蚤属Coptopsylla | 叶状切唇蚤突高亚种Lamellifer |
物种丰富度:表示物种种类丰富度, 用种类S表示, 即每种鼠蚤类种数之和。
计算染蚤率(rate of infected flea, Rf)和蚤指数(index of fleas, If)的公式为[16]:
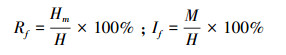
|
(1) |
式中, Hm代表染蚤鼠数, H代表检蚤总鼠数, M代表总蚤数量。
均匀度分析采用Pielon指数, Pielon指数又称物种的相对密度。物种数目越多, 多样性越丰富, 物种数目相同时, 每个物种的个体数越平均, 则多样性越丰富[16]:

|
(2) |
式中, J为均匀度指数, H为实测多样性值, Hmax是理论上群落多样性的最大值, S为种类数。
Simpson生态优势度指数, 生态优势度指数越大, 说明群落内物种数量分布越不均匀, 优势种的地位越突出[17]:

|
(3) |
式中, C′为群落的优势度指数;Ni为物种i的个体数;N为群落中所有物种的总个体数;Pi为物种i的个体数占群落中总个体数的比例;S为群落中的物种数。
分析采用Shannon-Weiner指数, 测定群落内物种多样性[18]:

|
(4) |
式中, H′代表群落的多样性指数, Pi为物种i的个体数占群落中总个体数的比例, S为群落中的物种数量。
Cody指数表明两个梯度群落的异质性, 随着Cody指数的增加, 表明不同梯度之间差异性越大[16]:

|
(5) |
a和b分别为两个梯度所包含物种数, c为两梯度带的共有物种数, g(H)为不同海拔梯度带增加的物种数, l(H)为沿纬度梯度带减少的物种数。群落相似性系数q值[19—20]:

|
(6) |
当q为0—0.25时代表两个群落为极不相似, q为0.25—0.50时代表两个群落为中等不相似, q为0.50—0.75时代表两个群落为中等相似, q为0.75—1.00时代表两个群落为极相似。其数值越大, 代表群落之间的相似性程度越大[21]。
1.4 聚类分析为探究研究区不同县(市)全局和局部的空间关联性, 本研究借助全局Moran′s I和局部Moran′s I, 全局Moran′s I大于0表明存在空间正相关性, 其值越大正相关性越强, 全局Moran′s I小于0表明存在空间负相关性, 其值越小, 负相关性越强。其中, 全局Moran′s I指数计算公式参考[22]。
局部Moran′s I指数计算公式参考[22]。
为表明多个变量之间的空间相关性, 在单变量Moran′s I指数的基础上进一步利用双变量全局自相关和局部自相关, 为探究不同生态学指标空间分布聚集性特征提供了可能。计算公式参考[23]。
1.5 标准差椭圆法与聚类分析不同, 标准差椭圆法(Standard Deviational Ellipse Method)应用于分析空间要素分布方向与趋势的研究中, 能够反映空间要素的主导分布方向, 以及空间要素各个方向的离散程度。本研究通过绘制不同鼠、蚤指标的标准差椭圆, 对鼠类和蚤类的同一生态学指标进行比较分析:
标准差椭圆圆心及其参数计算公式参考[24]。
本研究借助多个生态学指标来反映鼠、蚤群落的特征, 其中染蚤率和蚤指数反映不同类型宿主(鼠类)携带蚤类的概率和数量;物种丰富度和群落内物种多样性能反映群落内鼠、蚤种类的多样性;借助物种均匀度和生态优势度反映区域内物种集中分布程度;借助Cody指数和相似性系数反映不同地貌类型之间群落的相似程度。此外, 借助GIS空间分析方法(空间自相关与标准差椭圆法), 探究不同行政区内的鼠、蚤生态学指标在空间上是否具有聚类现象, 并对比不同区域生态指标的分布特点, 为研究鼠、蚤空间分布提供示范作用。
2 结果与分析依照徐正会等[25]提出的群落构成比, 将构成比大于10%的种类定为优势种, 1%—10%为普通种, 小于1%为稀有种。如表 4所示, 在所有的小型兽类中大沙鼠和子午沙鼠所占比例最大, 分别为42.1%和41%。因此, 大沙鼠和子午沙鼠属于鼠类优势种。本研究所捕获的13种小型兽类中, 11种鼠类染蚤。其中, 大沙鼠感染蚤种最多。在检获不同蚤类中, 簇鬃客蚤的数量最多, 其次是臀突客蚤, 二者属于寄生在大沙鼠(宿主)上的蚤类优势种。在不同鼠种染蚤率对比中, 染蚤率最高的是大沙鼠为85.98%, 其次是柽柳沙鼠为71.43%。总蚤指数最高的是大沙鼠和怪柳沙鼠, 数值分别为9.26和7.86。
| 宿主 Host |
检蚤鼠数 Number of rodent tested for fleas |
染蚤鼠数 Number of rodent infected with fleas |
染蚤率/% The rate of flea infection |
检蚤总数 Total flea count |
总蚤指数 Total flea index |
| 大沙鼠Rhombomys opimus | 1605 | 1380 | 85.98 | 14859 | 9.26 |
| 子午沙鼠Meriones meridianus Pallas | 1564 | 610 | 39.00 | 1822 | 1.16 |
| 红尾沙鼠Meriones libycus | 154 | 95 | 61.69 | 306 | 1.99 |
| 三趾跳鼠Dipus sagitta | 251 | 38 | 15.14 | 90 | 0.36 |
| 柽柳沙鼠Meriones tamariscinus | 28 | 20 | 71.43 | 220 | 7.86 |
| 灰仓鼠Cricetulus migratorius | 46 | 12 | 26.09 | 28 | 0.61 |
| 五趾跳鼠Allactaga sibirica | 67 | 6 | 8.96 | 34 | 0.51 |
| 小林姬鼠Apodemus sylvaticusLinnaeus | 46 | 3 | 6.52 | 5 | 0.11 |
| 小五趾跳鼠Allactaga elater | 35 | 7 | 20.00 | 36 | 1.03 |
| 小家鼠Mus musculus | 13 | 2 | 15.38 | 3 | 0.23 |
| 长耳跳鼠Euchoreutes naso | 3 | 2 | 66.67 | 5 | 1.67 |
| 合计Total | 3812 | 2175 | — | 17408 | — |
如图 2所示, 低海拔地貌类型的鼠类物种多样性和物种丰富度总体上高于中海拔地貌类型, 该生态学指标从低海拔到中海拔地貌类型总体上呈降低趋势, 表明低海拔地区更适宜鼠类的生长和繁殖。中海拔地貌类型生态优势度高于低海拔地貌类型, 表明部分鼠类针对特定的地貌类型表现出较强的适应性。低海拔鼠类均匀度高于中海拔地貌类型, 表明低海拔地貌类型鼠类分布更为均匀, 优势种表现得不明显。

|
| 图 2 不同地貌类型鼠类指标变化趋势 Fig. 2 The change trend of indexes of rodents in different geomorphic types Ⅰ:低海拔湖积平原;Ⅱ:低海拔冲积平原;Ⅲ:低海拔冲积洪积平原;Ⅳ:低海拔风积地貌;Ⅴ:中海拔湖积平原;Ⅵ:中海拔冲积洪积平原;Ⅶ:中海拔洪积平原;Ⅷ:中海拔剥蚀平原;物种多样性、均匀度、生态优势度纵坐标数值参考左边纵轴, 物种丰富度纵坐标数值参考右边纵轴 |
如图 3所示, 低、中海拔冲积平原地貌类型的物种多样性较高, 表明冲积平原地貌类型较适宜蚤类的生存与繁殖;中海拔地貌类型的蚤类均匀度总体上高于低海拔地区, 表明蚤类在中海拔地区分布较地海拔地区分布均匀;蚤类生态优势度与物种多样性呈相反的变化趋势, 中海拔剥蚀平原生态优势度最高(0.792), 其物种丰富度为2。而低海拔湖积平原最低(0.225), 其物种丰富度为17。

|
| 图 3 不同地貌类型蚤类指标变化趋势 Fig. 3 The change trend of indexes of fleas in different geomorphic types 物种多样性、均匀度、生态优势度纵坐标数值参考左边纵轴, 物种丰富度纵坐标数值参考右边纵轴 |
在不同地貌类型中, 对鼠、蚤的生态学指标进行相关性分析。如图 4所示,在鼠类生态学指标的相关性中,物种多样性和生态优势度呈负相关(P小于0.01), 物种多样性和物种丰富度呈正相关(P小于0.01), 生态优势度和物种丰富度呈负相关(P小于0.01), 其他鼠类生态学指标之间为无相关。在蚤类生态学指标相关性分析中, 结果显示物种多样性和生态优势度呈负相关(P小于0.01), 物种多样性和物种丰富度呈正相关(P小于0.05), 其他蚤类生态学指标之间为无相关。在鼠类的生态指标与蚤类的生态指标相关性分析中, 结果显示鼠类物种多样性和蚤类物种多样性呈正相关(P小于0.05), 鼠类物种多样性和蚤类物种丰富度呈正相关(P小于0.01), 鼠类生态优势度和蚤类生态优势度呈正相关(P小于0.05), 鼠类物种丰富度和蚤类物种丰富度呈正相关(P小于0.01), 鼠类生态优势度和蚤类物种丰富度(P小于0.05)、蚤类物种多样性(P小于0.05)、蚤类物种丰富度(P小于0.05)呈负相关。

|
| 图 4 鼠、蚤生态指标相关性热力图 Fig. 4 Heat map of the correlation between ecological indicators of rodents and fleas a1:鼠类物种多样;a2:鼠类均匀度;a3:鼠类生态优势度;a4:鼠类物种丰富度;a5:蚤类物种多样性;a6:蚤类均匀度;a7:蚤类生态优势度;a8:蚤类物种丰富度;*:P<0.05;* *:P<0.01 |
如表 5所示, 表示不同地貌类型鼠类和蚤类的相似性系数和Cody指数, 鼠类在低海拔湖积平原→低海拔冲积平原、低海拔湖积平原→低海拔冲积洪积平原, q值大于0.75为极相似。蚤类在低海拔湖积平原→低海拔冲积平原、低海拔湖积平原→低海拔冲积洪积平原、低海拔湖积平原→低海拔风积地貌、低海拔冲积平原→低海拔风积地貌, q值大于0.75为极相似。综上, 鼠、蚤群落在低海拔地貌类型之间的相似相似性高于中海拔地貌类型。而随相似性系数q值升高, Cody指数下降, 二者变化趋势相反。
| 鼠类指标Rodent index | 蚤类指标Flea index | |||||
| 地貌变化类型 Geomorphic change type |
q值 | Cody指数 | 地貌变化类型 Geomorphic change type |
q值 | Cody指数 | |
| Ⅰ→Ⅱ | 0.90 | 0.5 | Ⅰ→Ⅱ | 0.84 | 1.5 | |
| Ⅰ→Ⅲ | 0.75 | 1.5 | Ⅰ→Ⅲ | 0.76 | 2.0 | |
| Ⅰ→Ⅳ | 0.36 | 3.5 | Ⅰ→Ⅳ | 0.82 | 1.5 | |
| Ⅰ→Ⅴ | 0.33 | 3.0 | Ⅰ→Ⅴ | 0.12 | 7.5 | |
| Ⅰ→Ⅵ | 0.33 | 3.0 | Ⅰ→Ⅵ | 0.24 | 6.5 | |
| Ⅰ→Ⅶ | 0.40 | 3.0 | Ⅰ→Ⅶ | 0.65 | 3.0 | |
| Ⅰ→Ⅷ | 0.22 | 3.5 | Ⅰ→Ⅷ | 0.12 | 7.5 | |
| Ⅱ→Ⅲ | 0.83 | 1.0 | Ⅱ→Ⅲ | 0.63 | 3.5 | |
| Ⅱ→Ⅳ | 0.45 | 3.0 | Ⅱ→Ⅳ | 0.78 | 2.0 | |
| Ⅱ→Ⅴ | 0.30 | 3.5 | Ⅱ→Ⅴ | 0.05 | 9.0 | |
| Ⅱ→Ⅵ | 0.30 | 3.5 | Ⅱ→Ⅵ | 0.16 | 8.0 | |
| Ⅱ→Ⅶ | 0.36 | 3.5 | Ⅱ→Ⅶ | 0.61 | 3.5 | |
| Ⅱ→Ⅷ | 0.20 | 4.0 | Ⅱ→Ⅷ | 0.11 | 8.0 | |
| Ⅲ→Ⅳ | 0.50 | 3.0 | Ⅲ→Ⅳ | 0.69 | 2.5 | |
| Ⅲ→Ⅴ | 0.25 | 4.5 | Ⅲ→Ⅴ | 0.15 | 5.5 | |
| Ⅲ→Ⅵ | 0.25 | 4.5 | Ⅲ→Ⅵ | 0.31 | 4.5 | |
| Ⅲ→Ⅶ | 0.42 | 3.5 | Ⅲ→Ⅶ | 0.50 | 4.0 | |
| Ⅲ→Ⅷ | 0.17 | 5.0 | Ⅲ→Ⅷ | 0.15 | 5.5 | |
| Ⅳ→Ⅴ | 0.29 | 2.5 | Ⅳ→Ⅴ | 0.33 | 4.0 | |
| Ⅳ→Ⅵ | 0.50 | 1.5 | Ⅳ→Ⅵ | 0.29 | 5.0 | |
| Ⅳ→Ⅶ | 0.38 | 2.5 | Ⅳ→Ⅶ | 0.67 | 2.5 | |
| Ⅳ→Ⅷ | 0.33 | 2.0 | Ⅳ→Ⅷ | 0.14 | 6.0 | |
| Ⅴ→Ⅵ | 0.50 | 1.0 | Ⅴ→Ⅵ | 0.50 | 1.0 | |
| Ⅴ→Ⅶ | 0.60 | 1.0 | Ⅴ→Ⅶ | 0.18 | 4.5 | |
| Ⅴ→Ⅷ | 0.25 | 1.5 | Ⅴ→Ⅷ | 0.33 | 1.0 | |
| Ⅵ→Ⅶ | 0.60 | 1.0 | Ⅵ→Ⅶ | 0.36 | 3.5 | |
| Ⅵ→Ⅷ | 0.67 | 0.5 | Ⅵ→Ⅷ | 0.50 | 1.0 | |
| Ⅶ→Ⅷ | 0.40 | 1.5 | Ⅶ→Ⅷ | 0.18 | 4.5 | |
为获取各县(市)鼠、蚤指标的全局Moran′s I指数, 运用Geoda软件对其进行计算, 并进行显著性检验。结果显示:通过显著性检验的指标包括鼠类数量、子午沙鼠数量、蚤类均匀度, 全局Moran′s I指数范围是0.135—0.194, 且不同鼠、蚤指标的全局Moran′s I的P值小于0.05, 即Z值均大于1.65(表 6)。Moran′s I大于0的鼠、蚤指标包括各个县市总的鼠类数量、子午沙鼠数量、蚤类均匀度, 结果表明不同行政区空间分布位置(距离)越近, 行政区之间的相关性也变得越来越显著。
| 类型 Type |
鼠类数量 Number of Rodents |
子午沙鼠数量 Number of Meriones meridianus Pallas |
蚤类均匀度 Flea evenness |
| 全局Moran′s I Global Moran′s I | 0.194 | 0.161 | 0.135 |
| Z值Z scores | 2.65 | 2.83 | 1.74 |
| P值P value | <0.01 | <0.05 | <0.05 |
通过GeoDa空间分析软件, 计算不同鼠、蚤指标之间的双变量全局空间自相关指数。如表 7所示, 鼠类均匀度与蚤类均匀度、蚤类生态优势度表现出正相关。表明鼠类均匀度的聚集过程, 可能导致蚤类均匀度、蚤类生态优势度增加。结果证明, 鼠类均匀度对蚤类均匀度、蚤类生态优势度产生重要影响, 蚤类均匀度对鼠类生态优势度、鼠类物种多样性也具有重要意义。
| 指标名称 Index name |
鼠类均匀度 Rodent evenness |
蚤类均匀度 Flea evenness |
| , 鼠类生态优势度Ecological dominance of rodents | — | 0.224* |
| 鼠类物种多样性Rodent species diversity | — | 0.220* |
| 蚤类均匀度Flea evenness | 0.118* | — |
| 蚤类生态优势度Ecological dominance of fleas | 0.104* | — |
| 其中, 第一行代表第一变量, 第一列代表第二变量;*表示P<0.05, * *表示P<0.01 | ||
为探求不同鼠、蚤指标在空间上是否存在聚集现象, 运用GeoDa软件对25个县(市)级行政区各鼠、蚤指标进行局部空间自相关分析, 结果表明(图 5):本研究涉及到的鼠、蚤指标中, 通过显著性检验(P小于0.05)的指标包括鼠类总量、子午沙鼠数量、蚤类均匀度。如图 5所示, 沙湾县和石河子市属于高-高聚集模式, 表明沙湾县和石河子市鼠类数量较多且呈集聚分布的状态。

|
| 图 5 鼠、蚤指标单变量局部Moran′s I分布图 Fig. 5 Univariate local Moran′s I distribution of rodent and flea indicators |
如图 5所示, 子午沙鼠数量的局部Moran′s I指数存在高-高聚集、低-低聚集、低-高聚集三种聚集模式。其中, 沙湾县、玛纳斯县、石河子市存在高-高聚类模式, 表明该区域子午沙鼠数量较多且呈集聚分布的状态。呼图壁县属于低-高聚类模式, 表明呼图壁县子午沙鼠数量较少, 被周围子午沙鼠数量高的县市包围。布尔津县属于低-低聚类模式, 表明该区域子午沙鼠数量较少且与周围县市存在低值聚集现象。总体来看, 鼠类总量和子午沙鼠数量的空间聚集特点在沙湾县和石河子市表现出相似性。
如图 5所示, 蚤类均匀度的局部Moran′s I指数存在高-高聚集和高-低聚集模式。伊吾县、巴里坤哈萨克自治县属于高-高聚集模式, 表明该区域蚤类均匀度较高且集聚分布。和布塞克蒙古自治县、布尔津县、乌尔禾区属于高-低聚集模式, 表明该区域蚤类均匀度较高, 周围县市蚤类均匀度低。
2.2.4 双变量局部空间自相关为探究不同鼠、蚤指标之间是否存在空间相关性, 本研究运用双变量局部空间自相关分析法, 分析结果均通过显著性检验(P小于0.05), 结果如图 6所示。图 6中, 伊吾县属于高-高聚集模式, 表明伊吾县鼠类均匀度与相邻区域蚤类均匀度表现出高值聚集现象。布尔津县与和布塞克蒙古自治县属于高-低聚集模式, 表明该区域鼠类均匀度较高, 而周围相邻区域蚤类均匀度较低。巴里坤哈萨克自治县属于低-高聚集模式, 表明该区域鼠类均匀度较低, 而相邻区域蚤类均匀度较高。乌尔禾区属于低-低聚集模式, 表明乌尔禾区是鼠类均匀度较低, 且周围相邻区域蚤类均匀度较低;图 6中, 托里县和精河县属于高-低聚集模式, 白碱滩区属于低-低聚集模式, 表明不同蚤类可能存在对不同宿主(鼠种)的选择性, 即特定蚤种只寄生在特定鼠种, 使得蚤类生态优势度较低;图 6中, 具有低-低和高-低聚集模式。其中, 玛纳斯县、沙湾县、额敏县、吉木乃县属于低-低聚集模式, 表明该区域蚤类均匀度与鼠类生态优势度表现出低值的聚集特征。呼图壁县和昌吉市属于高-低聚集模式;图 6中, 克拉玛依区属于高-高聚类模式, 沙湾县、玛纳斯县和石河子市属于低-高聚集模式。综上, 借助双变量局部Moran′s I指数, 研究鼠蚤指标在空间上的聚类模式, 结果表明了鼠、蚤指标之间存在关联性。

|
| 图 6 鼠、蚤指标双变量局部Moran′s I分布图 Fig. 6 Bivariate local Moran′s I distribution of rodent and flea indicators |
总体来看, 研究结果如图 7和表 8所示, 除均匀度指标外, 蚤类生态学指标较鼠类分布更集中, 而鼠类指标则分布较分散。蚤类作为寄生生物, 可能对宿主(鼠类)的类型具有选择性, 致使蚤类较鼠类分布更集中。鼠类与蚤类物种多样性指标的标准差椭圆表现出分布特点的相似性, 鼠类指标的标准差椭圆较蚤类指标标准差椭圆偏北。鼠类与蚤类的均匀度和生态优势度指标的标准差椭圆表现出相似性, 鼠类指标的标准差椭圆较蚤类指标的标准差椭圆偏西。

|
| 图 7 鼠、蚤指数标准差椭圆分布图 Fig. 7 Oval distribution of standard deviation of indices for rodents and fleas |
| 名称 Name |
鼠类物种丰度 Rodent species abundance |
蚤类物种丰度 Flea species abundance |
鼠类物种多样性 Rodent species diversity |
蚤类物种多样性 Flea species diversity |
| 转角/(°) Rotation θ |
102.29 | 100.40 | 104.62 | 100.99 |
| 沿X轴标准差/km X-axis standard deviation |
28.71 | 28.23 | 29.44 | 31.73 |
| 沿Y轴标准差/km Y-axis standard deviation |
16.61 | 13.38 | 17.77 | 15.07 |
| 椭圆面积/km2 Ellipse area |
149813.25 | 118597.54 | 164302.25 | 150246.21 |
| 名称 Name |
鼠类均匀度 Rodent evenness |
蚤类均匀度 Flea evenness |
鼠类生态优势度 Ecological dominance of rodents |
蚤类生态优势度 Ecological dominance of fleas |
| 转角/(°) Rotation θ |
104.68 | 101.64 | 110.73 | 109.88 |
| 沿X轴标准差/km X-axis standard deviation |
30.76 | 33.30 | 29.51 | 26.43 |
| 沿Y轴标准差/km Y-axis standard deviation |
17.49 | 16.96 | 17.14 | 18.36 |
| 椭圆面积/km2 Ellipse area |
169048.82 | 177819.47 | 158890.20 | 152544.26 |
部分学者在研究鼠疫不同疫源地的环境因子变化对宿主影响的相关研究中, 发现环境因子是影响鼠疫发生的主要因素[26]。有研究证明不同景观类型对鼠疫的传播与爆发产生影响, 由于不同植被类型内部的连通性存在差异, 也造成鼠疫在不同区域传播的差异性[27]。也有学者的研究证明, 不同土壤类型中金属元素含量的差异性, 造成了自然疫源地产生鼠疫高发区和静息区现象[28]。考虑到不同海拔、不同地貌类型的水热条件、土壤因子、气候特点存在较大差异。一些学者通过研究鼠类与蚤类的空间分布特征与不同景观和海拔梯度之间的关系, 发现中低海拔地区适宜鼠疫传播[29—30]。因此, 本文通过对比不同海拔鼠、蚤指标的变化规律, 探究鼠类和蚤类在不同海拔、不同地貌类型上的分布规律, 分析不同地貌类型鼠类和蚤类生态学指标的相关性, 探究不同地貌类型鼠类和蚤类群落结构相似性。基于不同地貌类型, 研究鼠、蚤及其生态指标在空间上的差异性, 成为本研究的一大亮点。
关于不同地貌类型采样点布置原则问题, 不同学者遵循的采样点布设密度存在差异。有学者在不同地貌类型采用了不同的采样点布设密度, 即在不同地貌或生境下布置的采样点密度不相同[31—32];也有学者在不同地区鼠类分布特征研究中, 遵循相同的采样点布设密度[33—34]。综上, 在不同地貌、地区鼠、蚤分布特点的研究中, 不同地貌类型设置采样点密度的原则不尽相同。在本研究中, 由于研究区以低、中海拔地貌类型为主, 因此选取的采样点多分布于低海拔地貌类型。导致不同地貌类型的采样未按照均匀分布的原则进行, 这成为后续研究需要着重探究的问题。在今后的研究中, 将进一步探究低中海拔类型与高海拔地貌类型的鼠、蚤分布的差异。
部分学者在研究荒漠与半荒漠景观鼠、蚤群落特点及分布特征时, 研究结果存在一定差异性。有学者在研究锡林郭勒地区鼠类群落特征的研究中发现[35], 不同采样点的优势度指数和均匀性指数偏低。也有学者研究准噶尔盆地不同地区鼠类分布特征时发现[36], 在不同区域的鼠类群落中, 鼠类优势种各不相同, 这与本研究的结果具有相似性。有学者研究表明[37], 在新疆天山地区捕获的灰土拨鼠, 体外携带寄生虫在种群内持续传播, 并发现灰土拨鼠是鼠疫菌株的主要宿主, 且鼠疫菌株可通过灰土拨鼠传播到次级宿主(其它鼠类)。前人的研究成果表明荒漠与半荒漠景观的鼠、蚤相关问题的研究亟待进一步加强。
另外一些学者利用GIS的空间分析功能, 对鼠疫疫源县的分布、发病情况和地理环境因素制作专题地图并进行叠加分析, 按照时间和空间将动物鼠疫和人间鼠疫流行病进行分类汇总和统计[38]。本文借助地理学空间分析方法, 研究典型区域鼠、蚤分布特征, 进行鼠类和蚤类指标的对比研究。本研究存在两个问题需要在后续研究中解决:首先, 由于原始数据的缺乏, 在采样过程中未在同一地点采样多年数据, 致使本研究无法考虑自然因素和人文因素变化对鼠、蚤指标产生的影响, 也无法进行鼠、蚤生态学指标时序变化研究及其驱动力分析。其次, 在奎屯市和独山子区未进行采样, 导致研究区缺乏完整性。
鼠疫生物群落的稳定性受地理环境因子时空周期变化的制约。除自然因素外, 随着旅游业的发展, 人类生产生活方式不断变化, 进入鼠疫自然疫源地的人口不断增加, 致使宿主动物的栖息地受到影响。当区域的生态环境发生重大变化, 自然种群的进化和生存受到影响时, 鼠疫暴发和流行的风险将急剧增加。因此, 深入研究鼠疫与地理要素的关系, 对于揭示鼠疫自然疫源地的结构特征、时空分布格局和鼠疫流行规律具有重要意义。
3.2 结论本研究探讨准噶尔盆地荒漠景观不同地貌类型鼠、蚤生态学指标的变化特征, 借助空间自相关、标准差椭圆及重心迁移方法研究不同鼠、蚤生态学指标的空间变化特征, 主要得出以下结论:
(1) 在鼠、蚤生态指标的相关性分析中, 物种多样性与物种丰富度整体上呈正相关, 物种多样性与生态优势度整体上呈现负相关。鼠类与蚤类在低海拔地貌类型的群落相似度较高, 在中海拔地貌类型的群落相似度较低。
(2) 在不同县市通过显著性检验的鼠、蚤指标单变量全局Moran′s I指数均大于0, 空间聚集态势表现为鼠类数量大于蚤类均匀度, 蚤类均匀度大于子午沙鼠数量。通过显著性检验的双变量全局Moran′s I指数均为正相关。单变量局部空间自相关结果表明, 鼠、蚤指标存在四种聚集模式, 且以高-高聚集模式最为典型。双变量Moran′s I指数表明, 蚤类生态指标可能受鼠类生态指标的影响。
(3) 鼠类物种多样性和蚤类物种多样性的标准差椭圆显示了二者分布特征的相似性, 鼠类指标的标准差椭圆较蚤类指标标准差椭圆偏北。均匀度和生态优势度指标的标准差椭圆表现出相似性, 鼠类指标的标准差椭圆较蚤类指标的标准差椭圆偏西。综上, 蚤类作为寄生生物, 不同蚤类可能会分布在特定宿主体内, 因此蚤类指标比鼠类指标分布更集中。
综上, 现有研究缺乏对鼠疫流行与蚤类数量、自然疫源地的地理环境关系的相关性探究, 这些因素在空间上不是独立分布的, 其具有复杂的时空拓扑。而数理统计、生态学、流行病学等方法无法分析这些地理因素之间复杂的时空关系, 本研究在考虑统计学相关性的基础上, 引入空间分析方法, 探究不同地貌、区域鼠、蚤生态学指标的空间差异。因此, 利用GIS空间分析方法研究鼠疫有关问题势在必行。
| [1] |
Eisen R J, Atiku L A, Mpanga J T, Enscore R E, Acayo S, Kaggwa J, Yockey B M, Apangu T, Kugeler K J, Mead P S. An evaluation of the flea index as a predictor of plague epizootics in the West Nile region of Uganda. Journal of Medical Entomology, 2020, 57(3): 893-900. DOI:10.1093/jme/tjz248 |
| [2] |
Galy A, Loubet P, Peiffer-Smadja N, Yazdanpanah Y. The plague: an overview and hot topics. La Revue De Medecine Interne, 2018, 39(11): 863-868. DOI:10.1016/j.revmed.2018.03.019 |
| [3] |
方喜业, 许磊, 刘起勇, 张荣祖. 中国鼠疫自然疫源地分型研究Ⅰ.生态地理景观特征. 中华流行病学杂志, 2011, 32(12): 1232-1236. DOI:10.3760/cma.j.issn.0254-6450.2011.12.012 |
| [4] |
方喜业, 杨瑞馥, 刘起勇, 董兴齐, 张荣祖, 许磊. 中国鼠疫自然疫源地分型研究Ⅱ.鼠疫自然疫源地分型方法研究. 中华流行病学杂志, 2012, 33(2): 234-238. DOI:10.3760/cma.j.issn.0254-6450.2012.02.024 |
| [5] |
王启果, 曹汉礼, 孟卫卫, 雒涛, 阿不力克木, 阿不力米提, 戴翔, 阿扎提, 王信惠, 李冰, 蒋卫, 张晓兵, 雷刚, 郭荣, 张渝疆. 准噶尔盆地鼠疫自然疫源地鼠体蚤类群落及其在动物鼠疫流行病学中的作用. 中国媒介生物学及控制杂志, 2013, 24(1): 11-16. |
| [6] |
贵有军, 王启果, 雒涛, 阿不力米提·买托呼提, 阿布力克木·阿布都热西提, 王信惠, 王诚, 古丽阿依·包开西, 张晓兵, 李冰, 黎唯. 新疆乌苏市古尔图镇2006—2010年动物间鼠疫监测分析. 中国媒介生物学及控制杂志, 2019, 30(3): 331-333. |
| [7] |
Sariyeva G, Bazarkanova G, Maimulov R, Abdikarimov S, Kurmanov B, Abdirassilova A, Shabunin A, Sagiyev Z, Dzhaparova A, Abdel Z, Mussagaliyeva R, Morand S, Motin V, Kosoy M. Marmots and Yersinia pestis strains in two plague endemic areas of Tien Shan Mountains. Frontiers in Veterinary Science, 2019, 6: 207. DOI:10.3389/fvets.2019.00207 |
| [8] |
张爱萍, 魏荣杰, 熊浩明, 王祖郧. 气候因素对鼠疫分布影响的研究进展. 中华预防医学杂志, 2016, 50(5): 459-462. DOI:10.3760/cma.j.issn.0253-9624.2016.05.015 |
| [9] |
黄钦妮, 董治宝, 肖南, 冯淼彦, 石寰宇, 陈国祥, 白子怡, 张欣, 高冲, 王晓枝. 海南岛景心角海滩老鼠艻(Spinifex littoreus)灌丛沙堆沉积及形态特征. 中国沙漠, 2021, 41(6): 157-168. |
| [10] |
杨琳, 赵金, 王新华, 牛毅, 尹文江, 解卫刚, 艾尼瓦尔, 陈文亮, 文国新. 空间分析方法在准噶尔盆地大沙鼠血清学监测中的应用. 医学动物防制, 2019, 35(8): 726-729. |
| [11] |
杨林生, 陈如桂, 王五一, 谭见安. 1840年以来我国鼠疫的时空分布规律. 地理研究, 2000, 19(3): 243-248. DOI:10.3321/j.issn:1000-0585.2000.03.003 |
| [12] |
Zhuang D F, Du H W, Wang Y, Jiang X S, Shi X M, Yan D. Probing the spatial cluster of Meriones unguiculatus using the nest flea index based on GIS technology. Acta Tropica, 2016, 163: 157-166. DOI:10.1016/j.actatropica.2016.08.007 |
| [13] |
张渝疆, 戴翔, 阿不力米提, 蒋卫, 阿不力克木, 王信惠, 布仁明德, 热娜, 李冰, 雷刚, 孟卫卫, 木合塔尔, 张晓兵, 王启国, 雒涛, 郭荣, 王忠, 唐建国, 刘成全, 阿扎提, 叶瑞玉, 于心, 曹汉礼. 准噶尔盆地鼠疫调查. 中华流行病学杂志, 2008, 29(2): 136-144. DOI:10.3321/j.issn:0254-6450.2008.02.008 |
| [14] |
张鸿猷, 盛广吉, 汤国厚. 新疆鼠疫. 地方病通报编辑部, 1993, 31-40. |
| [15] |
李炳元, 潘保田, 韩嘉福. 中国陆地基本地貌类型及其划分指标探讨. 第四纪研究, 2008, 28(4): 535-543. DOI:10.3321/j.issn:1001-7410.2008.04.004 |
| [16] |
詹银珠, 郭宪国, 左小华, 王乔花, 吴滇. 云南省部分地区地里纤恙螨分布调查. 中华流行病学杂志, 2011, 32(1): 13-16. |
| [17] |
刘维暐, 王杰, 王勇, 杨帆. 三峡水库消落区不同海拔高度的植物群落多样性差异. 生态学报, 2012, 32(17): 5454-5466. |
| [18] |
李明娟, 赵娟娟, 江南, 潘妮, 张曼琳, 束承继. 深圳近海公园植物群落结构特征及其与景观格局的相关性. 生态学报, 2021, 41(22): 8732-8745. |
| [19] |
赵志模, 郭依泉. 群落生态学原理与方法(第三版). 重庆: 科学技术文献出版社重庆分社, 1990, 1-288. |
| [20] |
洪梅, 刘正祥, 杨桂荣, 吴爱国, 王国良, 宋志忠, 高子厚. 香格里拉县小型兽类寄生蚤群落多样性特征. 现代预防医学, 2015, 42(8): 1483-1488. |
| [21] |
尹锴, 崔胜辉, 石龙宇, 吝涛, 郭青海, 吕静. 人为干扰对城市森林灌草层植物多样性的影响——以厦门市为例. 生态学报, 2009, 29(2): 563-572. |
| [22] |
杨晴青, 刘倩, 尹莎, 张戬, 杨新军, 高岩辉. 秦巴山区乡村交通环境脆弱性及影响因素——以陕西省洛南县为例. 地理学报, 2019, 74(6): 1236-1251. |
| [23] |
姚小薇, 曾杰, 李旺君. 武汉城市圈城镇化与土地生态系统服务价值空间相关特征. 农业工程学报, 2015, 31(9): 249-256. |
| [24] |
朱磊, 杨爱民, 夏鑫鑫, 武红旗. 基于空间自相关的1975—2015年玛纳斯河流域耕地时空特征变化分析. 中国生态农业学报: 中英文, 2020, 28(6): 887-899. |
| [25] |
徐正会, 杨比伦, 胡刚. 西双版纳片断山地雨林蚁科昆虫群落研究. 动物学研究, 1999, 20(4): 288-293. |
| [26] |
王丽君, 周立志, 马勇. 中国鼠疫宿主鼠类丰富度格局及疫区环境因子分析. 兽类学报, 2010, 30(4): 411-417. |
| [27] |
袁星, 杨林生, 李海蓉, 王利, 韦炳干. 2000—2018年内蒙古长爪沙鼠疫源地地表景观特征及变化. 地理研究, 2022, 41(3): 867-879. |
| [28] |
魏有文, 陈洪舰, 孟献亚, 王雪, 喇翠玲, 周奎章, 米宝玉, 李千, 马英. 青海青南地区鼠疫自然疫源地鼠疫高发区与静息区土壤中12种金属元素含量分析. 中华地方病学杂志, 2021, 40(12): 947-952. |
| [29] |
刘霖. 滇南家鼠鼠疫疫原地鼠、蚤多样性空间分布格局的研究[D]. 大理: 大理学院, 2014.
|
| [30] |
张长国. 玉龙鼠疫自然疫源地宿主和媒介的群落[D]. 大理: 大理大学, 2016.
|
| [31] |
王庭柱. 丹江库区鼠类种群分布及携带寄生虫调查[D]. 郑州: 郑州大学, 2010.
|
| [32] |
冯蕾, 赵运林, 张美文, 王勇, 徐正刚, 彭姣, 王双业. 洞庭湖洲滩及滨湖区鼠类分布格局及其多样性. 生态学报, 2017, 37(17): 5771-5779. |
| [33] |
阿不都热合曼·吐尔逊余亮, 艾尼瓦尔·吐米尔, 张大铭. 新疆北部干旱地区鼠类群落空间结构差异的比较研究. 新疆大学学报(自然科学版), 25(2): 8. |
| [34] |
侯祥, 韩宁, 封托, 张博, 陈晓宁, 王京, 常罡. 气候因素对黑线姬鼠种群动态影响的非线性效应. 生态学报, 2020, 40(14): 4836-4841. |
| [35] |
王利清, 杨玉平, 张福顺, 解永凤, 董维惠. 锡林郭勒盟荒漠草原区鼠类群落特征. 中国草地学报, 2015, 37(4): 114-118. |
| [36] |
王启果, 古丽阿依·包开西, 雒涛, 李博, 戴翔, 曹汉礼, 张渝疆. 簇鬃客蚤对宿主侵袭能力的实验观察. 疾病预防控制通报, 2013, 28(5): 1-3. |
| [37] |
Sariyeva G, Bazarkanova G, Maimulov R, Abdikarimov S, Kurmanov B, Abdirassilova A, Shabunin A, Sagiyev Z, Dzhaparova A, Abdel Z, Mussagaliyeva R, Morand S, Motin V, Kosoy M. Marmots and Yersinia pestis strains in two plague endemic areas of Tien Shan Mountains. Frontiers in Veterinary Science, 2019, 6: 207. |
| [38] |
唐新元, 王梅, 陈洪舰, 李红英, 蔡佳作, 何多龙, 崔百忠. 基于地理信息系统的青海省60年鼠疫流行病学特征. 中国媒介生物学及控制杂志, 2018, 29(6): 60-64. |