 2023, Vol. 43
2023, Vol. 43文章信息
- 魏杰, 王晶苑, 温学发
- WEI Jie, WANG Jingyuan, WEN Xuefa
- 植物光合与呼吸过程CO2及其δ13C的变异特征与影响因素研究进展
- Research progress on variations and influencing factors of CO2 and its δ13C during photosynthesis and respiration in plants
- 生态学报. 2023, 43(10): 4319-4331
- Acta Ecologica Sinica. 2023, 43(10): 4319-4331
- http://dx.doi.org/10.5846/stxb202202180392
-
文章历史
- 收稿日期: 2022-02-18
- 采用日期: 2023-02-13

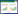




植物光合与呼吸过程是陆地生态系统碳循环的重要组成部分, 其与大气之间的CO2交换量是人为释放CO2量的15—20倍[1]。光合过程是植物碳过程的起始环节, 由于受到叶片气孔、叶肉和羧化作用等影响, 大气中的CO2通过气孔进入叶片合成初级光合产物的过程存在同位素效应[2—5]。呼吸过程的底物供应来自于叶片光合产物的转化和迁移, 由于底物和分馏驱动的同位素效应, 叶片、茎杆和根系等呼吸释放CO2 δ13C具有明显时空分布特征[6—7]。植物光合与呼吸过程CO2及其δ13C的变异特征与驱动机制可以为植物与生态系统碳循环过程解析以及模型模拟研究提供理论基础和数据支撑[8—10]。
植物光合与呼吸过程CO2及其δ13C具有明显的日变化和季节变化特征, 主要受控于气孔导度、叶肉导度、羧化作用和光合产物供应等生物因素以及光照、温度和湿度等气象因素[6, 11—15]。叶片扩散分馏和羧化分馏导致光合产物同位素组成的差异[16—18];而葡萄糖合成和淀粉累积过程、韧皮部蔗糖释放/回收过程以及新老碳的混合等导致茎杆和根系呼吸CO2 δ13C变异特征与叶片存在一定的时间滞后效应[7, 19]。
到目前为止, 关于不同时间尺度上光合与呼吸CO2 δ13C的变异特征及其影响因素仍然缺少清晰认识。弥补这一知识空白对于植物碳分配模式[9, 20]、碳代谢关键过程[6, 8, 14, 21]以及生态系统呼吸组分拆分等研究至关重要[5, 22—23]。本文系统梳理了植物光合与呼吸碳过程的同位素效应及其时间变化特征与影响因素, 总结了存在的问题, 并展望了未来的研究方向, 以期为今后该领域的研究提供理论依据和有效支撑。
1 植物碳过程及其同位素效应概述植物碳过程主要包括光合过程和后羧化过程两部分, 光合过程是植物碳过程的起始环节, 吸收大气CO2合成初级光合产物;后羧化过程是光合产物转化和迁移环节, 用于维持植物生长和抵御环境胁迫(图 1)。

|
| 图 1 植物光合与呼吸碳过程及其同位素效应示意图[6—7] Fig. 1 Schematic diagram of carbon processes and their isotopic effects during plant photosynthesis and respiration [6—7] 1表示光合产物合成过程, 其同位素效应主要为光合同位素分馏;2, 6和9表示后羧化过程中与淀粉代谢相关的碳过程, 其同位素效应主要为蔗糖与淀粉之间相互转化过程中的分馏效应;3, 7和10表示后羧化过程中与不同化合物之间碳分配的相关过程, 其同位素效应主要为NSC与SC相互转化过程中的分馏效应;4, 8和11表示呼吸过程, 其同位素效应主要为呼吸分馏;5表示韧皮部运输过程, 其同位素效应主要为蔗糖装载、运输和卸载中的分馏效应;12表示通过菌根真菌网络进行的地下碳交换过程, 其同位素效应主要为具有不同同位素信号的碳组分交换过程中的平衡分馏效应。1为植物叶片光合过程CO2同化过程, 2—12为后羧化过程中碳的储存、转化、迁移和释放过程 |
光合过程主要发生在叶片的叶绿体内, 包括原初反应、光反应和暗反应(图 2)。其中, 原初反应主要发生在叶绿体的类囊体, 是光合过程的初始环节;光反应主要发生在叶绿体类囊体膜, 包括光反应1和光反应2两个过程, 提供必需的酶和ATP[16, 24—25];根据光合途径的不同, 暗反应主要分为C3途径、C4途径和景天科酸代谢途径(Crassulacean acid metabolism, CAM)[6, 17, 26]。以C3途径为例, 光合过程包括:大气CO2通过边界层到达气孔并进入叶肉细胞, 在核酮糖-1, 5-二磷酸羧化酶(Rubisco)作用下, CO2与核酮糖-1, 5-二磷酸(RuBP)合成甘油酸-3-磷酸(PGA), 在甘油醛磷酸脱氢酶作用下被还原型辅酶Ⅱ(NADPH)和H+还原形成甘油醛-3-磷酸(PGAld), 即初级光合产物。C3途径, 即卡尔文循环, 是所有植物光合作用碳同化的基本途径, 也是植物合成葡萄糖和淀粉的光合产物的唯一途径;C4和CAM途径并不普遍存在, 且只能固定和转运CO2, 而不能形成葡萄糖和淀粉等光合产物。光照、温度、水分等条件适宜时, 光合作用形成的葡萄糖转化为蔗糖, 部分蔗糖进一步合成淀粉并暂时储存在叶片中;在环境胁迫条件下, 光合作用受限, 光合产物供应不足, 淀粉则转化为蔗糖, 供叶片生长、呼吸等代谢作用[6—7, 11]。

|
| 图 2 叶片结构、叶绿体结构以及光合碳合成过程 Fig. 2 Structure of leaf and chloroplast and the photosynthetic carbon synthesis |
在光合碳合成过程中, 由于扩散分馏和羧化分馏等同位素效应的存在, 植物优先利用较轻的12CO2, 导致植物体内有机物13C贫化[2—3, 27]。扩散分馏主要发生在大气CO2通过边界层和气孔进入叶绿体的过程中, CO2通过边界层、气孔和细胞内转移过程的分馏系数分别为2.9‰、4.4‰和1.8‰。其中, CO2在细胞内转移过程中的分馏包括气态CO2溶于水的扩散分馏(0.7‰)和CO2在溶液中的平衡分馏(1.1‰, 25 ℃)[2—3]。羧化分馏主要发生在CO2与核酮糖-1, 5-双二磷酸/磷酸烯醇式丙酮酸在羧化酶催化作用下的羧化过程, 其分馏系数为29‰[2—3]。呼吸分馏主要包括光呼吸分馏和暗呼吸分馏, 其中, 光呼吸分馏系数为8‰;不同研究者计算得到的暗呼吸分馏系数不同, 目前应用较多的分馏系数为0‰。
1.2 后羧化过程及其同位素效应叶片后羧化过程主要发生在叶绿体基质和细胞质中, 光合过程形成的甘油醛-3磷酸等主要通过两种途径进入下一步反应, 其中一部分在叶绿体基质内形成果糖-6磷酸, 进一步转化为ADP-葡萄糖, 最后形成淀粉, 暂时储存在叶绿体中;一部分则通过叶绿体被膜上的转运器运出叶绿体, 在细胞质中形成蔗糖[19, 28]。其次, 叶绿体中的淀粉在适当条件下转化成蔗糖;细胞质中的蔗糖主要用于叶片呼吸, 合成纤维素和木质素, 通过筛管向茎杆和根系运输, 并在茎杆和根系中进行NSC和SC的转化、并通过呼吸代谢释放CO2等[6, 23]。蔗糖由叶片向茎杆和根系的运输过程主要是在韧皮部筛管中完成的, 包括叶片韧皮部装载、维管束的运输以及在茎杆和根系韧皮部中卸载, 其中, 蔗糖转运蛋白、转化酶和单糖转运蛋白对于蔗糖等运输至关重要[29]。根系中的细根通常与丛枝菌根或外生菌根共生, 形成菌根真菌网络[8, 30], 并基于此网络, 进行地下碳交换[8—9]。
后羧化过程中的同位素效应与代谢途径以及特定有机质输出等有关[6, 28, 31], 主要包括叶片后羧化分馏过程、叶片-茎杆-根系运输过程以及茎杆和根系中物质转化和混合等过程的同位素效应(表 1)。叶片后羧化过程的同位素效应主要包括葡萄糖合成过程中的同位素效应(分馏驱动, 包括碎分裂分馏以及和酶相关的分馏等)、淀粉累积过程中的同位素效应(底物驱动, 主要为淀粉与蔗糖相互转化等)、呼吸(苹果酸脱羧)过程中的同位素效应(底物驱动, 主要为葡萄糖分解)、以及光促进暗呼吸过程中的同位素效应(底物驱动, 包括丙酮酸脱羧等)[6]。叶片-茎杆茎干-根系运输过程的同位素效应主要包括蔗糖在韧皮部蔗糖中的释放/回收过程中的同位素效应(底物驱动, 主要为不同来源蔗糖的混合)、以及不同代谢过程和滞留时间的蔗糖混合过程中的同位素效应(底物驱动, 主要为蔗糖混合过程的平衡分馏)[6, 28]。茎干和根系后羧化过程的同位素效应主要包括物质转换过程中的同位素效应(底物驱动, 主要为可溶性糖、淀粉等相互转化)、PEPC重新固定CO2过程中的同位素效应(分馏驱动, 包括PEPC排斥13C、溶解和水合过程等)、碳交换过程中的同位素效应(底物驱动, 主要为不同δ13C可溶性碳的交换过程)和呼吸过程中的同位素效应等[6, 31]。
| 植物碳过程 Carbon processes in plant |
不同器官碳过程 Carbon processes in different plant organ |
碳同位素效应 Carbon isotope effect |
分馏系数 Fractionation factor |
| 光合过程 | 扩散过程(叶片) | 大气CO2通过边界层的扩散分馏 | 2.9‰ |
| Photosynthetic process | 大气CO2通过气孔的扩散分馏 | 4.4‰ | |
| 气态CO2溶于水过程中的扩散分馏 | 0.7‰ (25℃) | ||
| CO2在溶液平衡过程中的平衡分馏 | 1.1‰ (25℃) | ||
| 羧化过程(叶片) | 核酮糖-1, 5-二磷酸羧化过程中的羧化分馏 | 30.0‰ | |
| 光呼吸分馏 | 8.0‰ | ||
| 暗呼吸分馏 | 0.0‰或-6‰ | ||
| 后羧化过程 | 合成和转化过程(叶片) | 葡萄糖合成过程中的分裂分馏 | — |
| Post-carboxylated processes | 淀粉累积过程中的酶促分馏 | — | |
| 呼吸(苹果酸脱羧)过程中的脱羧分馏 | 5.7‰ (磷酸戊糖途径) | ||
| 光促进暗呼吸过程中的脱羧分馏 | — | ||
| 运输过程(叶片-茎杆-根系) | 蔗糖运输过程的动力学分馏和平衡分馏 | — | |
| 转化过程(茎杆和根系) | 物质转换过程中的同位素效应 | 8.9‰ | |
| PEPc重新固定CO2过程中的同位素效应 | 5.7‰ | ||
| 碳交换过程中的同位素效应 | — |
植物光合过程的关键参数叶肉导度是光合作用的重要影响因素, 也是叶片尺度模型以及区域甚至全球尺度的碳循环模型的输入参数, 其测定方法通常包括叶绿素荧光法、曲线拟合法和碳同位素分馏在线联测法[32—34]。叶绿素荧光法通常基于光呼吸补偿点(Γ*)和日间线粒体呼吸速率(RL)计算获得, 而Γ*和RL的经典测定方法Laisk法的理论基础和准确性受到了广泛质疑, 且二者的测定误差对光合速率计算结果影响显著[25, 35]。此外, 叶片对光的吸收能力及其在光系统中的分配比例等经验系数均存在不确定性。虽然Γ*也是碳同位素分馏在线联测法的输入参数, 但其对该方法中叶肉导度的计算影响很小[36]。曲线拟合法易于理解、便于操作, 但其拟合模型因光合作用的发生状态不同而不同, 不利于推广[33]。越来越多的研究者认为, 碳同位素分馏在线联测法更适合多种环境条件下叶肉导度的准确计算[26, 36], 而叶绿素荧光法中的Laisk法在低辐射条件下可能导致叶片呼吸速率的低估[35]。
植物呼吸过程CO2及其δ13C的观测方法主要包括微气象学方法(涡度协方差法和通量梯度法)和箱式法。微气象学方法多用于冠层、生态系统或区域尺度上CO2通量观测, 其中, 通量梯度法直接观测两个高度处CO2浓度, 结合湍流扩散系数, 即可计算呼吸通量;或者直接观测两个高度处δ13C即可得到同位素通量比值, 适用于湍流较弱和有效风浪区较小的情况。涡度协方差法已成为生态系统尺度CO2通量观测的标准化、规范化方法;但易受复杂地形和复杂气象条件的制约与影响。箱式法是单株植物或叶片尺度上的CO2通量观测方法。箱式法则从早期的静态箱-碱液吸收滴定[37]、静态箱-气相色谱测定[38]以及静态箱(放置离体叶片、茎杆或根系)-稳定同位素质谱仪[7, 35]等人工离线测定模式, 逐渐发展到自动箱-红外光谱测定[39—42]等自动在线测定模式, 已成为目前通用的方法。
以上观测技术与方法通常在自然条件(同位素自然丰度)或人工条件(同位素脉冲标记/连续标记)下进行。基于自然条件或人工条件下的CO2及其δ13C的连续观测以及植物体不同器官和组分碳含量及其δ13C组成, 与生物和气象要素等相结合, 前人开展了植物体内关键碳过程、碳分配模式以及生态系统光合与呼吸组分拆分等研究[7, 19, 41, 43]。
3 光合过程CO2及其δ13C变异特征与影响因素光合过程CO2及其δ13C日变化特征通常表现为单峰曲线或双峰曲线, 气孔导度、叶肉导度和羧化作用是光合过程的关键限制因素[5, 25, 44—45]。
3.1 光合过程CO2及其δ13C变异特征叶片光合过程CO2日变化特征通常表现为单峰曲线或双峰曲线, 即光合CO2同化速率在上午或下午达到峰值, 而中午由于气孔开度降低或关闭, 光合CO2同化速率降至最低, 出现“午休”现象[44—45]。其季节变异普遍表现为“单峰型”, 即光合CO2同化速率在生长季中期达到最高, 而在生长季初期和末期较低。此外, 少量研究发现了植物光合CO2同化速率的“双峰型”变化趋势, 即光合CO2同化速率在生长季初期逐渐升高并达到峰值, 随后逐渐回落至低谷, 在生长季中期前后再次逐渐升高并达到峰值, 并在末期快速回落。与植物光合CO2同化速率变化趋势相似, 光合过程CO2 δ13C通常表现为“单峰型”或“双峰型”两种类型, 即上午或下午分馏较大, 而中午前后分馏较小的变异特征[3, 27]。
3.2 光合过程的影响因素光合过程的影响因素通常包括生物因素和非生物因素。早期一些研究认为CO2从气孔下腔到叶绿体羧化位点的扩散阻力趋近于无穷小, 从而简单地把生物影响因素划分为气孔限制和非气孔限制两个因素[35]。随着叶肉导度研究的深入, 发现不同植物叶肉导度存在差异, 并且随着光照、温度、湿度等环境条件发生变化。因此, 叶肉导度是光合过程的重要限制因素, 进而将光合过程的影响因素进一步细分为气孔限制、叶肉限制和羧化限制三个限制因素[35—36]。
光合过程的非生物影响因素主要包括光照、温度和湿度的环境因素。其中, 强光、高温和低湿等环境下, 植物气孔开度降低或关闭, 导致光合作用原料供给不足和CO2同化速率降低[2—3]。低光强下发育的叶片, 叶肉细胞壁厚度增加, 碳酸酐酶和水孔蛋白等活性降低, 叶绿体的位置改变, 导致叶肉限制显著偏高[46—47]。羧化过程中的核酮糖-1, 5-二磷酸羧化酶和甘油醛磷酸脱氢酶等酶活性降低, 产生光抑制并阻止光合产物合成[25, 45, 48]。气孔限制和叶肉限制条件下, 扩散分馏是光合过程同位素效应的主要控制因素, 此时, 12CO2优先通过边界层和气孔进入叶片, 溶于水后进一步输送到羧化位点。由于气孔限制和叶肉限制导致的分馏效应远低于羧化分馏, 因此, 其分馏系数低于气孔和叶肉不受限制时的分馏系数, 导致初级光合产物δ13C偏正[6—7, 36, 49]。羧化限制条件下, 由于酶活性降低和光抑制效应, 用于酶促羧化反应的12CO2明显减少, 导致初级光合产物δ13C偏正[3, 34]。光合过程的分馏效应使得植物体有机质δ13C比大气偏负20‰左右[2—3]。
4 呼吸过程CO2及其δ13C变异特征与影响因素叶片[27, 50]、茎杆[15, 51]和根系[13, 52]呼吸过程CO2及其δ13C日变化通常表现为单峰曲线或双峰曲线, 达到峰值的时间一般在中午12点到晚上22点之间[27, 50], 主要受控于光合产物供应[52]、物候[53—54]等生物因素以及土壤温度[55]、湿度[56—57]、氮含量[58]等非生物因素。呼吸过程CO2 δ13C变异的驱动机制通常包括呼吸底物合成和转化过程的同位素效应、PEPc重新固定CO2过程中的同位素效应、呼吸脱羧过程的同位素效应以及碳交换过程中的同位素效应等[6—7]。
4.1 呼吸过程CO2及其δ13C变异特征植物呼吸主要包括叶片呼吸、茎杆呼吸和根系呼吸过程, 其CO2及其δ13C日变化通常表现为单峰曲线或双峰曲线。叶片呼吸包括光呼吸和暗呼吸两部分, 受到温度、湿度、光照、O2和CO2等环境条件的共同影响[32, 59—61]。光呼吸需要叶绿体、过氧化物体和线粒体的协同作用, 代谢途径中的几种主要物质(乙醇酸, 乙醛酸和甘氨酸)都是二碳化合物(也称为C2循环), 主要受控于叶绿体内O2和CO2浓度, 表现为高O2和低CO2浓度促进光呼吸[62—63]。暗呼吸也称作细胞呼吸或线粒体呼吸, 在有光和无光条件下均可进行, 包括白天的非光呼吸和夜间呼吸, 白天暗呼吸通常低于夜间, 这可能是光照抑制暗呼吸作用引起的, 即kok效应[32, 35]。茎杆呼吸和根系呼吸过程底物来源主要包括光合产物由地上向地下的运输过程以及地下碳向上运输过程, 具体过程请见4.2和4.3节。
叶片呼吸、茎杆呼吸和根系呼吸过程的同位素效应主要与呼吸底物合成和转化过程、脱羧过程和碳交换过程等有关。光呼吸分馏主要与丙酮酸或谷氨酸脱羧过程有关, 释放的CO2 13C贫化, 其δ13C通常表现为早晚偏正、中午偏负的趋势, 总的分馏系数为7‰—8‰[62, 64]。叶片暗呼吸、茎杆呼吸和根系呼吸顾城的同位素效应主要与呼吸底物合成和转化过程、PEPc重新固定CO2过程以及呼吸脱羧过程有关[6—7]。呼吸底物合成和转化过程的同位素效应首先发生在葡萄糖合成过程的醛缩酶缩合作用, 使13C在己糖的C-3和C-4位置相对富集, 而12C在磷酸丙糖中相对富集, 导致呼吸释放的CO2相对于底物更加富集13C[6, 65—66]。PEPc重新固定CO2主要包括三个分馏过程, 即PEPc排斥13C, 分馏系数约为2.2‰;CO2进入水中溶解平衡, 使得气态中富集13C, 分馏系数为1.1‰;水合作用平衡分馏富集13C, 分馏系数为9‰;PEPc重新固定CO2总的分馏作用达到5.7‰[3, 67]。呼吸脱羧过程的同位素效应在糖酵解、TCA和PPP途径中均有发生, 且与葡萄糖合成过程相反。糖酵解途径及其后的丙酮酸脱氢酶复合物(PDH)的脱羧反应利用葡萄糖C-3和C-4位置的碳, 释放出13C相对富集的CO2[7, 12]。PPP过程产生的CO2来自于13C相对贫化的葡萄糖C-1位置, 其对13C和12C的分馏作用分别为9.6‰和4‰, 使得CO2的13C相对呼吸底物贫化[68]。
4.2 光合产物向下运输过程对呼吸CO2及其δ13C的影响机制蔗糖作为茎杆呼吸和根系呼吸底物的主要来源, 主要通过韧皮部筛管由叶片向下迁移(图 3), 其运输方式包括通过蔗糖转运蛋白的质外体装载、沿着共质体的运输以及扩散过程[15, 41, 51]。蔗糖运输的驱动机制目前仍存在争议, 包括基于质量流量模型的源库膨压差驱动机制[69—70]和基于均匀膨压以及膨压和溶质浓度扰动的长距离快速分布机制[71]两种观点。氧化还原电位控制的蔗糖转运以及蔗糖与植物激素间的信号传达是碳水化合物分配的重要控制因素[72]。茎杆呼吸过程不仅受控于蔗糖供应, 还取决于茎杆利用现有蔗糖的能力[6, 73]。

|
| 图 3 茎杆结构与可溶性糖在茎杆中的运输过程及其同位素效应 Fig. 3 Stem structure and the transport process and isotope effect of soluble sugar in stem |
蔗糖从筛管渗出后的代谢反应中发生排斥13C的分馏, 导致呼吸释放的CO2 13C贫化;根据质量守恒, 重新回收到韧皮部中的蔗糖中13C更加富集(图 3)[6, 28]。白天韧皮部蔗糖主要由醛缩酶/转酮醇酶的反应产物丙糖转化而来, 其13C相对贫化;而夜间韧皮部蔗糖由暂时淀粉降解而来, 其13C相对富集[66, 74]。不同代谢过程和滞留时间蔗糖具有不同的δ13C值, 其混合过程明显减弱了茎杆中淀粉δ13C值的昼夜变异, 并且随着向基部运输距离增加而增强[7]。茎杆基部和根韧皮部蔗糖的δ13C值通常没有明显的昼夜变化[70, 75], 表明运输过程中蔗糖混合过程可能不是呼吸释放CO2的δ13C值昼夜变异的主要控制因素, 而呼吸过程所利用底物(可溶性糖、淀粉、脂类/氨基酸、或储存的和新鲜的同化物等)的昼夜变化可能是这一现象的主要控制因素[7, 76—77]。
4.3 根系CO2向上运输过程对呼吸CO2及其δ13C的影响机制根系呼吸通常被认为是土壤呼吸的重要组成部分, 但最近的研究表明, 很大一部分根系呼吸释放的CO2继续储存在根系中, 并通过木质部导管, 与水和矿物质等同步向地上部分输送, 并通过茎干呼吸或叶片呼吸释放到大气中, 进而影响CO2排放速率的日变化和季节变异特征[78]。除根系自身呼吸产生的CO2外, 根系还可以从土壤中吸收溶解无机碳, 但在大多数情况下, 由于根系-土扩散梯度限制, 植物从土壤中吸收的CO2很少, 表明根木质部中的大部分CO2来自根系呼吸。Aubrey和Teskey研究发现, 通过木质部蒸腾流运输到地上部分的CO2是根系呼吸释放到土壤中CO2的2倍[79]。已有的研究表明, 根系来源的CO2对茎杆呼吸的贡献比例最高可达12%[79—81]。定量分析根系木质部CO2通量, 对于了解根系呼吸作用具有重要意义, 并将重新定义根系代谢的能量成本以及地下碳分配的估算, 提高对于根呼吸和森林碳循环的理解[78]。
不同植物根系通过菌根真菌网络进行地下碳交换, 其交换的量级及其同位素组成对根系呼吸以及从根系运输至地上部分的CO2浓度及其δ13C具有显著影响(图 4)。几乎所有陆地植物都存在地下真菌菌根网络[82], 前人的研究多关注于对养分吸收的贡献, 但是对于碳交换的研究相对较少[8, 82—83]。通过对挪威云杉大树进行长期连续原位标记研究发现, 地下碳交换在细根总碳量中占比达到40%[8], 而对花旗松幼苗开展控制实验研究的结果表明, 地下碳交换只占到整个标记期间总碳量的0.1%[9]。根系碳交换通常双向进行, 而不是根据供需梯度决定交换方向, 因此, 其对参与碳交换的植物根系呼吸底物均有显著影响[8]。

|
| 图 4 植物间根系与菌根真菌网络共生关系、根系与丛植菌根共生关系以及根系与外生菌根共生关系示意图 Fig. 4 Symbiotic relationship between roots and mycorrhizal fungi network, roots and arbuscular mycorrhizal, and roots and ectomycorrhizal among plants |
基于自然条件下CO2及其δ13C的连续观测以及植物体不同器官和组分碳含量及其δ13C组成, 与生物和气象要素等相结合, 前人开展了植物碳在植物体内分配模式和迁移速率及其内在驱动机制研究[7, 41]。将箱式法与涡度协方差法相结合, Wingate等分析了光合碳同位素分馏与植物茎杆、土壤和生态系统呼吸同位素信号的相关关系, 发现呼吸释放CO2 δ13C值变异幅度低于光合分馏, 并且具有2—10d的延迟, 可能是新合成碳与植物体内碳混合过程引起的分馏效应导致的[11]。将涡度协方差法与手动和自动箱式法相结合, Brændholt等分析了日尺度、季节尺度和年际尺度上温带山毛榉林树干、根系和土壤呼吸CO2的相对贡献, 结果发现, 冬季茎杆呼吸和土壤呼吸对生态系统呼吸的贡献分别为6%和52%, 夏季茎杆呼吸、根系呼吸和土壤呼吸对生态系统呼吸的贡献分别为16%、17%和49%[41]。Brændholt等利用自动箱-红外光谱技术对森林土壤、茎杆和根系CO2及其δ13C通量开展了两个月的连续观测, 发现不同生态系统单元(完整土壤、沟状土壤、树干和粗根)呼吸CO2 δ13C值差异较小[41]。
基于人工条件下的同位素脉冲/连续标记方法研究了标记CO2进入树木及其释放到土壤和大气中的情况[19, 43]。脉冲标记在量化碳分配、评估碳分配在植物生长过程中的作用、资源获取和碳储存等应用广泛。根据47项相关研究的数据, Epron等发现阔叶和针叶树物种之间的转移速率不同, 并且随着温度和土壤含水量的降低以及干旱的增强而降低[84]。标记的碳在韧皮部溶液(转移库)和成熟叶(源器官)中的半衰期较短, 而在库器官(生长组织、季节性贮藏)中的半衰期较长。在高时间分辨率下, 植物呼吸CO2 δ13C观测提供了新合成的碳在呼吸底物库中平均停留时间的最佳估测值, 并为不同器官分别建模提供了理论和数据基础。Studer等利用脉冲标记和连续标记方法研究了植物-土壤系统中碳的迁移、分配和储存时间, 结果发现当第8d达到13C输入和输出平衡时, 两种方法得到的结果才达到一致[19]。
与脉冲标记方法更适用于从叶片到茎杆和根系等的最小迁移时间研究不同, 连续标记方法则更适用于平均迁移时间研究。针对全球升温对植物呼吸的影响, Drake等开展了大树冠层脉冲标记实验, 并对叶片、冠层、根系和土壤呼吸CO2及其δ13C值进行了连续观测, 结果发现实验升温3℃并没有导致叶片和冠层呼吸速率的增加, 也没有改变整株植物不同器官呼吸贡献的相对比例以及碳的平均储存时间, 呼吸对变暖的完全适应可能会抑制正的气候变暖反馈[43]。总的来看, 标记的碳中10%用于地上部分呼吸, 40%用于地下部分呼吸, 其余50%储存在植物体内。
5.2 生态系统光合与呼吸组分拆分生态系统光合与呼吸是构成净生态系统碳交换量(NEE)的重要组分, 基于生态系统光合和呼吸过程, 理论上NEE可细分为总生态系统光合通量、生态系统叶片光呼吸通量、生态系统叶片暗呼吸通量以及生态系统非叶片呼吸通量[85]。叶片光合过程中, 边界层、气孔、叶肉、羧化限制以及光呼吸和暗呼吸等产生的同位素效应导致光合产物13C贫化而大气CO2的13C富集的过程称为光合碳同位素判别[2—3]。呼吸过程中, 后羧化过程的同位素效应导致呼吸释放的CO2的13C贫化, 进而使得大气CO2的13C贫化[86]。基于生态系统光合产物和呼吸CO2的同位素组成(δ13C)的差异可以实现NEE组分光合和呼吸通量的拆分, 该技术称为同位素通量拆分方法。生态系统非叶片呼吸δ13C和冠层尺度光合判别(13Δcanopy)是同位素通量拆分方法中两个关键参数。传统的基于涡度协方差系统观测的生态系统光合和呼吸通量拆分通常采用温度/光照响应函数等统计拆分方法[87—88], 但存在温度敏感性变化、自相关以及忽略光对叶片呼吸的抑制作用(即Kok效应)造成光照条件下呼吸通量被高估等问题[23, 88]。而高时间分辨率大气CO2和δ13C数据与涡度协方差通量观测相结合的同位素通量拆分方法, 不借助与温度或光照相关的功能函数关系, 通过独立计算可实现昼夜和日尺度光合和呼吸通量的拆分, 有效避免了涡度协方差技术通常采用的温度/光照响应函数等统计拆分方法存在的自相关及高估白天呼吸通量等问题[5, 22]。
生态系统光合和呼吸通量同位素组成的差异是生态系统NEE光合和呼吸通量拆分的理论基础。Wehr和Saleska利用完整生长季的同位素通量数据分析发现, NEE大的日变化可能是由通量塔采样足迹在高通量和低通量区域之间的变化引起的, 而这是涡度协方差技术无法发现的[22]。结果表明光合作用的同位素特征随季节变化在-24‰和-28‰之间, 在生长季的大部分时间里, 生态系统碳输入和输出之间的同位素差异保持在-0.5‰左右。Wehr等基于温带落叶林3年以上的同位素通量观测研究, 发现基于同位素通量拆分理论计算的生态系统呼吸在6—7月仅为夜间NEE的一半左右, 首次在生态系统尺度上发现了Kok效应[23]。Oikawa等研究表明, 基于Reichstein方法和人工神经网络方法利用夜间NEE来推断日间生态系统呼吸高估了日间GPP和生态系统呼吸的量级, 可能是由于植物夜间呼吸速率高于白天导致的, 进一步证实了Kok效应的存在[89]。Chen等选择植物生长旺季高温干旱和低温湿润两个时期作为对比研究时段, 基于大气CO2 δ13C比值和通量廓线观测系统7个高度的大气CO2和δ13C数据以及涡度协方差通量数据, 利用同位素通量拆分和统计拆分两种理论实现了小时尺度的NEE拆分, 同位素通量拆分理论在高温干旱的中午时段估算的呼吸通量低于夜间, 而统计拆分方法没有捕捉到这种白天呼吸通量的降低, 推测与Kok效应和研究区季节性干旱有关[5]。
6 结论与展望植物光合与呼吸过程CO2及其δ13C的时间变异特征能够反映植物碳分配方式、碳代谢关键过程以及植物与环境相互作用等重要生理生态过程, 可以为植物和生态系统碳循环过程和模型模拟研究提供理论依据和数据支撑。随着观测技术和方法的进步, 逐步实现了非破坏性的野外原位高频观测, 有效获取了小时尺度或更小时间尺度上植物光合与呼吸CO2及其δ13C的变异特征, 揭示了植物体内碳的分配模式和迁移时间以及生态系统光合与呼吸拆分等问题[5, 22]。
由于光合与呼吸涉及碳合成、转化和迁移等多种代谢过程, 其内在机理和分馏机制仍需进一步研究。光合过程的关键限制因素(比如:叶肉导度)并不能直接观测, 需要多种手段相结合以有效提升计算的准确性[34]。由于白天的呼吸代谢是由光合作用、光呼吸和氮同化等相互作用的结果, 这些相互作用在细胞代谢水平如何整合及其对脱羧过程的影响仍不清楚, 导致无法预测光下叶片呼吸CO2通量及其对环境条件的响应机制[32]。此外, Laisk方法和Kok方法通常低估了白天叶片呼吸, 为了更好地理解叶片呼吸代谢的机制, 叶片对辐射和CO2摩尔分数的响应还有待进一步研究[35]。虽然前人已经对非结构性碳水化合物与结构性碳水化合物相互转化的条件及其对碳同位素组成的影响开展了大量研究, 但仍需将实验方法与对广泛植物种类和环境条件的实地研究相结合, 以进一步确定非结构性碳水化合物在近期和未来的双重作用及其对呼吸CO2及其δ13C变异特征的影响机制。在不同植物种类中, 根系CO2通过木质部导管运移地上部分的比例及其对植物和生态系统碳动态和同位素特征的影响等研究很少[79]。植物间地下碳交换的过程、方向和量级仍不清楚, 植被本身和环境条件对上述过程的影响仍需进一步研究[8—9]。
| [1] |
Landman W. Climate change 2007: The physical science basis. South African Geographical Journal, 2010, 92: 86-87. DOI:10.1080/03736245.2010.480842 |
| [2] |
Farquhar G D, O'Leary M H, Berry J A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Functional Plant Biology, 1982, 9(2): 121. DOI:10.1071/PP9820121 |
| [3] |
Farquhar G D, Ehleringer J R, Hubick K T. Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology, 1989, 40: 503-537. DOI:10.1146/annurev.pp.40.060189.002443 |
| [4] |
Hikosaka K, Niinemets V, Anten N P R. Canopy Photosynthesis: From Basics to Applications. Dordrecht: Springer Netherlands, 2016.
|
| [5] |
Chen C H, Wei J, Wen X F, Sun X M, Guo Q J. Photosynthetic carbon isotope discrimination and effects on daytime NEE partitioning in a subtropical mixed conifer plantation. Agricultural and Forest Meteorology, 2019, 272/273: 143-155. DOI:10.1016/j.agrformet.2019.03.027 |
| [6] |
Brüggemann N, Gessler A, Kayler Z, Keel S G, Badeck F, Barthel M, Boeckx P, Buchmann N, Brugnoli E, Esperschütz J, Gavrichkova O, Ghashghaie J, Gomez-Casanovas N, Keitel C, Knohl A, Kuptz D, Palacio S, Salmon Y, Uchida Y, Bahn M. Carbon allocation and carbon isotope fluxes in the plant-soil-atmosphere continuum: a review. Biogeosciences, 2011, 8: 3457-3489. DOI:10.5194/bg-8-3457-2011 |
| [7] |
Werner C, Gessler A. Diel variations in the carbon isotope composition of respired CO2 and associated carbon sources: a review of dynamics and mechanisms. Biogeosciences, 2011, 8(9): 2437-2459. DOI:10.5194/bg-8-2437-2011 |
| [8] |
Klein T, Siegwolf R T W, Körner C. Belowground carbon trade among tall trees in a temperate forest. Science, 2016, 352(6283): 342-344. DOI:10.1126/science.aad6188 |
| [9] |
Pickles B J, Wilhelm R, Asay A K, Hahn A S, Simard S W, Mohn W W. Transfer of 13C between paired Douglas-fir seedlings reveals plant kinship effects and uptake of exudates by ectomycorrhizas. New Phytologist, 2017, 214: 400-411. DOI:10.1111/nph.14325 |
| [10] |
Ivanov Y V, Kartashov A V, Zlobin I E, Sarvin B, Stavrianidi A N, Kuznetsov V V. Water deficit-dependent changes in non-structural carbohydrate profiles, growth and mortality of pine and spruce seedlings in hydroculture. Environmental and Experimental Botany, 2019, 157: 151-160. DOI:10.1016/j.envexpbot.2018.10.016 |
| [11] |
Wingate L, Ogée J, Burlett R, Bosc A, Devaux M, Grace J, Loustau D, Gessler A. Photosynthetic carbon isotope discrimination and its relationship to the carbon isotope signals of stem, soil and ecosystem respiration. New Phytologist, 2010, 188(2): 576-589. DOI:10.1111/j.1469-8137.2010.03384.x |
| [12] |
Werner R A, Buchmann N, Siegwolf R T W, Kornexl B E, Gessler A. Metabolic fluxes, carbon isotope fractionation and respiration—lessons to be learned from plant biochemistry. New Phytologist, 2011, 191(1): 10-15. DOI:10.1111/j.1469-8137.2011.03741.x |
| [13] |
Lai Z R, Lu S, Zhang Y Q, Wu B, Qin S G, Feng W, Liu J B, Fa K Y. Diel patterns of fine root respiration in a dryland shrub, measured in situ over different phenological stages. Journal of Forest Research, 2016, 21(1): 31-42. DOI:10.1007/s10310-015-0511-4 |
| [14] |
Gong J N, Wang B, Jia X, Feng W, Zha T S, Kellomäki S, Peltola H. Modelling the diurnal and seasonal dynamics of soil CO2 exchange in a semiarid ecosystem with high plant-interspace heterogeneity. Biogeosciences, 2018, 15(1): 115-136. DOI:10.5194/bg-15-115-2018 |
| [15] |
Salomón R L, Steppe K, Crous K Y, Noh N J, Ellsworth D S. Elevated CO2 does not affect stem CO2 efflux nor stem respiration in a dry Eucalyptus woodland, but it shifts the vertical gradient in xylem[CO2]. Plant, Cell & Environment, 2019, 42(7): 2151-2164. |
| [16] |
Tholen D, Zhu X G. The Mechanistic Basis of Internal Conductance: A Theoretical Analysis of Mesophyll Cell Photosynthesis and CO2 Diffusion. Plant Physiology, 2011, 156(1): 90-105. DOI:10.1104/pp.111.172346 |
| [17] |
Tholen D, Ethier G, Genty B, Pepin S, Zhu X G. Variable mesophyll conductance revisited: theoretical background and experimental implications. Plant, Cell & Environment, 2012, 35(12): 2087-2103. |
| [18] |
Busch F A, Holloway-Phillips M, Stuart-Williams H, Farquhar G D. Revisiting carbon isotope discrimination in C3 plants shows respiration rules when photosynthesis is low. Nature Plants, 2020, 6: 245-258. DOI:10.1038/s41477-020-0606-6 |
| [19] |
Studer M, Siegwolf R, Abiven S. Carbon transfer, partitioning and residence time in the plant-soil system: a comparison of two 13CO2 labelling techniques. Biogeosciences, 2014, 11: 1637-1648. DOI:10.5194/bg-11-1637-2014 |
| [20] |
Galiano L, Timofeeva G, Saurer M, Siegwolf R, Martínez-Vilalta J, Hommel R, Gessler A. The fate of recently fixed carbon after drought release: towards unravelling C storage regulation in Tilia platyphyllos and Pinus sylvestris. Plant, Cell & Environment, 2017, 40(9): 1711-1724. |
| [21] |
Martínez-Vilalta J, Sala A N, Asensio D, Galiano L, Hoch G, Palacio S, Piper F I, Lloret F. Dynamics of non-structural carbohydrates in terrestrial plants: a global synthesis. Ecological Monographs, 2016, 86(4): 495-516. DOI:10.1002/ecm.1231 |
| [22] |
Wehr R, Saleska S R. An improved isotopic method for partitioning net ecosystem-atmosphere CO2 exchange. Agricultural and Forest Meteorology, 2015, 214/215: 515-531. DOI:10.1016/j.agrformet.2015.09.009 |
| [23] |
Wehr R, Munger J W, McManus J B, Nelson D D, Zahniser M S, Davidson E A, Wofsy S C, Saleska S R. Seasonality of temperate forest photosynthesis and daytime respiration. Nature, 2016, 534(7609): 680-683. DOI:10.1038/nature17966 |
| [24] |
Tomás M, Flexas J, Copolovici L, Galmés J, Hallik L, Medrano H, Ribas-Carbó M, Tosens T, Vislap V, Niinemets V. Importance of leaf anatomy in determining mesophyll diffusion conductance to CO2 across species: quantitative limitations and scaling up by models. Journal of Experimental Botany, 2013, 64(8): 2269-2281. DOI:10.1093/jxb/ert086 |
| [25] |
Xiao Y, Zhu X G. Components of mesophyll resistance and their environmental responses: A theoretical modelling analysis. Plant, Cell & Environment, 2017, 40: 2729-2742. |
| [26] |
Ma W T, Tcherkez G, Wang X M, Schäufele R, Schnyder H, Yang Y S, Gong X Y. Accounting for mesophyll conductance substantially improves 13C-based estimates of intrinsic water-use efficiency. New Phytologist, 2021, 229(3): 1326-1338. DOI:10.1111/nph.16958 |
| [27] |
Wingate L, Seibt U, Moncrieff J B, Jarvis P G, Lloyd J. Variations in 13C discrimination during CO2 exchange by Picea sitchensis branches in the field. Plant, Cell & Environment, 2007, 30(5): 600-616. |
| [28] |
Hobbie E A, Werner R A. Intramolecular, compound-specific, and bulk carbon isotope patterns in C3 and C4 plants: a review and synthesis. New Phytologist, 2004, 161(2): 371-385. DOI:10.1111/j.1469-8137.2004.00970.x |
| [29] |
张懿, 张大兵, 刘曼. 植物体内糖分子的长距离运输及其分子机制. 植物学报, 2015, 50(1): 107-121. |
| [30] |
何新华, 段英华, 陈应龙, 徐明岗. 中国菌根研究60年: 过去、现在和将来. 中国科学: 生命科学, 2012, 42(6): 431-454. DOI:10.3969/j.issn.0253-2778.2012.06.001 |
| [31] |
Badeck F W, Tcherkez G, Nogués S, Piel C, Ghashghaie J. Post-photosynthetic fractionation of stable carbon isotopes between plant organs—a widespread phenomenon. Rapid Communications in Mass Spectrometry, 2005, 19(11): 1381-1391. DOI:10.1002/rcm.1912 |
| [32] |
Tcherkez G, Gauthier P, Buckley T N, Busch F A, Barbour M M, Bruhn D, Heskel M A, Gong X Y, Crous K Y, Griffin K, Way D, Turnbull M, Adams M A, Atkin O K, Farquhar G D, Cornic G. Leaf day respiration: low CO2 flux but high significance for metabolism and carbon balance. New Phytologist, 2017, 216(4): 986-1001. DOI:10.1111/nph.14816 |
| [33] |
朱凯, 袁凤辉, 关德新, 吴家兵, 王安志. 植物叶肉导度的测定及计算方法综述. 应用生态学报, 2019, 30(5): 1772-1782. DOI:10.13287/j.1001-9332.201905.002 |
| [34] |
巩晓颖, 马薇婷, 余咏枝, 李蕾. 植物叶肉导度的测定方法及其研究进展. 应用生态学报, 2020, 31(6): 1882-1888. DOI:10.13287/j.1001-9332.202006.010 |
| [35] |
Gong X Y, Tcherkez G, Wenig J, Schäufele R, Schnyder H. Determination of leaf respiration in the light: comparison between an isotopic disequilibrium method and the Laisk method. New Phytologist, 2018, 218(4): 1371-1382. DOI:10.1111/nph.15126 |
| [36] |
Gu L H, Sun Y. Artefactual responses of mesophyll conductance to CO2 and irradiance estimated with the variable J and online isotope discrimination methods. Plant, Cell & Environment, 2014, 37(5): 1231-1249. |
| [37] |
Lundegårdh H. Carbon dioxide evolution of soil and crop growth. Soil Science, 1927, 23(6): 417-453. DOI:10.1097/00010694-192706000-00001 |
| [38] |
Midwood A J, Millard P. Challenges in measuring the δ13C of the soil surface CO2 efflux. Rapid Communications in Mass Spectrometry, 2011, 25(1): 232-242. DOI:10.1002/rcm.4857 |
| [39] |
Plain C, Gerant D, Maillard P, Dannoura M, Dong Y W, Zeller B, Priault P, Parent F, Epron D. Tracing of recently assimilated carbon in respiration at high temporal resolution in the field with a tuneable diode laser absorption spectrometer after in situ 13CO2 pulse labelling of 20-year-old beech trees. Tree Physiology, 2009, 29(11): 1433-1445. DOI:10.1093/treephys/tpp072 |
| [40] |
Savage K, Phillips R, Davidson E. High temporal frequency measurements of greenhouse gas emissions from soils. Biogeosciences, 2014, 11(10): 2709-2720. DOI:10.5194/bg-11-2709-2014 |
| [41] |
Brændholt A, Ibrom A, Ambus P, Larsen K S, Pilegaard K. Combining a quantum cascade laser spectrometer with an automated closed-chamber system for δ13C measurements of forest soil, tree stem and tree root CO2 fluxes. Forests, 2019, 10(5): 432. DOI:10.3390/f10050432 |
| [42] |
魏杰, 陈昌华, 王晶苑, 温学发. 箱式通量观测技术和方法的理论假设及其应用进展. 植物生态学报, 2020, 44(4): 318-329. |
| [43] |
Drake J E, Furze M E, Tjoelker M G, Carrillo Y, Barton C V M, Pendall E. Climate warming and tree carbon use efficiency in a whole-tree 13CO2 tracer study. New Phytologist, 2019, 222(3): 1313-1324. DOI:10.1111/nph.15721 |
| [44] |
Flexas J, Barbour M M, Brendel O, Cabrera H M, Carriquí M, Díaz-Espejo A, Douthe C, Dreyer E, Ferrio J P, Gago J, Gallé A, Galmés J, Kodama N, Medrano H, Niinemets V, Peguero-Pina J J, Pou A, Ribas-Carbó M, Tomás M, Tosens T, Warren C R. Mesophyll diffusion conductance to CO2: An unappreciated central player in photosynthesis. Plant Science, 2012, 193/194: 70-84. DOI:10.1016/j.plantsci.2012.05.009 |
| [45] |
韩吉梅, 张旺锋, 熊栋梁, Jaume FLEXAS, 张亚黎. 植物光合作用叶肉导度及主要限制因素研究进展. 植物生态学报, 2017, 41(8): 914-924. |
| [46] |
Flexas J, Díaz-Espejo A, Conesa M A, Coopman R E, Douthe C, Gago J, Gallé A, Galmés J, Medrano H, Ribas-Carbo M, Tomàs M, Niinemets V. Mesophyll conductance to CO2 and Rubisco as targets for improving intrinsic water use efficiency in C3 plants. Plant, Cell & Environment, 2016, 39(5): 965-982. |
| [47] |
Galmés J, Molins A, Flexas J, Conesa M À. Coordination between leaf CO2 diffusion and Rubisco properties allows maximizing photosynthetic efficiency in Limonium species. Plant, Cell & Environment, 2017, 40(10): 2081-2094. |
| [48] |
Atkin O K, Evans J R, Ball M C, Lambers H, Pons T L. Leaf respiration of snow gum in the light and dark. interactions between temperature and irradiance. Plant Physiology, 2000, 122(3): 915-923. DOI:10.1104/pp.122.3.915 |
| [49] |
Kohn M J. Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(46): 19691-19695. DOI:10.1073/pnas.1004933107 |
| [50] |
Sun W, Resco V, Williams D G. Diurnal and seasonal variation in the carbon isotope composition of leaf dark-respired CO2 in velvet mesquite (Prosopis velutina). Plant, Cell & Environment, 2009, 32(10): 1390-1400. |
| [51] |
Salomón R L, Valbuena-Carabaña M, Gil L, McGuire M A, Teskey R O, Aubrey D P, González-Doncel I, Rodríguez-Calcerrada J. Temporal and spatial patterns of internal and external stem CO2 fluxes in a sub-Mediterranean oak. Tree Physiology, 2016, 36(11): 1409-1421. |
| [52] |
Makita N, Kosugi Y, Kamakura M. Linkages between diurnal patterns of root respiration and leaf photosynthesis in Quercus crispula and Fagus crenata seedlings. Journal of Agricultural Meteorology, 2014, 70(3): 151-162. DOI:10.2480/agrmet.D-14-00006 |
| [53] |
Savage K, Davidson E A, Tang J. Diel patterns of autotrophic and heterotrophic respiration among phenological stages. Global Change Biology, 2013, 19(4): 1151-1159. DOI:10.1111/gcb.12108 |
| [54] |
Lak Z A, Sandén H, Mayer M, Rewald B. Specific root respiration of three plant species as influenced by storage time and conditions. Plant and Soil, 2020, 453: 615-626. DOI:10.1007/s11104-020-04619-9 |
| [55] |
Chen T T, Lin C F, Song T T, Guo R Q, Cai Y Y, Chen W Y, Xiong D C, Jiang Q, Chen G S. Does root respiration of subtropical Chinese fir seedlings acclimate to seasonal temperature variation or experimental soil warming?. Agricultural and Forest Meteorology, 2021, 308/309: 108612. DOI:10.1016/j.agrformet.2021.108612 |
| [56] |
Hasibeder R, Fuchslueger L, Richter A, Bahn M. Summer drought alters carbon allocation to roots and root respiration in mountain grassland. New Phytologist, 2015, 205(3): 1117-1127. DOI:10.1111/nph.13146 |
| [57] |
Paudel I, Bar-Tal A, Rotbart N, Ephrath J, Cohen S. Water quality changes seasonal variations in root respiration, xylem CO2, and sap pH in citrus orchards. Agricultural Water Management, 2018, 197: 147-157. DOI:10.1016/j.agwat.2017.11.007 |
| [58] |
Jing H, Liu Y, Wang G L, Liu G B. Effects of nitrogen addition on root respiration of trees and understory herbs at different temperatures in Pinus tabulaeformis forest. Plant and Soil, 2021, 463: 447-459. DOI:10.1007/s11104-021-04925-w |
| [59] |
Ayub G, Smith R A, Tissue D T, Atkin O K. Impacts of drought on leaf respiration in darkness and light in Eucalyptus saligna exposed to industrial-age atmospheric CO2 and growth temperature. New Phytologist, 2011, 190(4): 1003-1018. DOI:10.1111/j.1469-8137.2011.03673.x |
| [60] |
王兆国, 王传宽. 三种温带树种叶片呼吸的时间动态及其影响因子. 生态学报, 2013, 33: 1456-1464. |
| [61] |
谢双泽. 水分亏缺对不同抗旱性小麦穗部光呼吸特性及其关键酶基因表达的影响[D]. 西安: 西北农林科技大学, 2017.
|
| [62] |
Ehlers I, Augusti A, Betson T R, Nilsson M B, Marshall J D, Schleucher J. Detecting long-term metabolic shifts using isotopomers: CO2-driven suppression of photorespiration in C3 plants over the 20th century. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(51): 15585-15590. DOI:10.1073/pnas.1504493112 |
| [63] |
Walker X J, Mack M C, Johnstone J F. Stable carbon isotope analysis reveals widespread drought stress in boreal black spruce forests. Global Change Biology, 2015, 21(8): 3102-3113. DOI:10.1111/gcb.12893 |
| [64] |
Ghashghaie J, Badeck F W, Lanigan G, Nogués S, Tcherkez G, Deléens E, Cornic G, Griffiths H. Carbon isotope fractionation during dark respiration and photorespiration in C3 plants. Phytochemistry Reviews, 2003, 2(1): 145-161. |
| [65] |
Tcherkez G, Nogués S, Bleton J, Cornic G, Badeck F, Ghashghaie J. Metabolic origin of carbon isotope composition of leaf dark-respired CO2 in French bean. Plant Physiology, 2003, 131(1): 237-244. DOI:10.1104/pp.013078 |
| [66] |
Tcherkez G, Farquhar G, Badeck F, Ghashghaie J. Theoretical considerations about carbon isotope distribution in glucose of C3 plants. Functional Plant Biology, 2004, 31(9): 857-877. DOI:10.1071/FP04053 |
| [67] |
Gessler A, Tcherkez G, Karyanto O, Keitel C, Ferrio J P, Ghashghaie J, Kreuzwieser J, Farquhar G D. On the metabolic origin of the carbon isotope composition of CO2 evolved from darkened light-acclimated leaves in Ricinus communis. New Phytologist, 2009, 181(2): 374-386. DOI:10.1111/j.1469-8137.2008.02672.x |
| [68] |
Bathellier C, Tcherkez G, Bligny R, Gout E, Cornic G, Ghashghaie J. Metabolic origin of the δ13C of respired CO2 in roots of Phaseolus vulgaris. New Phytologist, 2009, 181(2): 387-399. DOI:10.1111/j.1469-8137.2008.02679.x |
| [69] |
Münch E. Die Stoffbewegungen in der Pflanze. Gustav Fischer, Jena, Germany.
|
| [70] |
Gessler A, Keitel C, Kodama N, Weston C, Winters A J, Keith H, Grice K, Leuning R, Farquhar G D. δ13C of organic matter transported from the leaves to the roots in Eucalyptus delegatensis: short-term variations and relation to respired CO2. Functional Plant Biology, 2007, 34(8): 692-706. DOI:10.1071/FP07064 |
| [71] |
Thompson M V. Scaling phloem transport: elasticity and pressure-concentration waves. Journal of Theoretical Biology, 2005, 236(3): 229-241. DOI:10.1016/j.jtbi.2005.03.009 |
| [72] |
Rolland F, Baena-Gonzalez E, Sheen J. Sugar sensing and signaling in plants: conserved and novel mechanisms. Annual Review of Plant Biology, 2006, 57: 675-709. DOI:10.1146/annurev.arplant.57.032905.105441 |
| [73] |
Dietze M C, Sala A, Carbone M S, Czimczik C I, Mantooth J A, Richardson A D, Vargas R. Nonstructural Carbon in Woody Plants. Annual Review of Plant Biology, 2014, 65: 667-687. DOI:10.1146/annurev-arplant-050213-040054 |
| [74] |
Gessler A, Tcherkez G, Peuke A D, Ghashghaie J, Farquhar G D. Experimental evidence for diel variations of the carbon isotope composition in leaf, stem and phloem sap organic matter in Ricinus communis. Plant, Cell & Environment, 2008, 31(7): 941-953. |
| [75] |
Rascher K G, Máguas C, Werner C. On the use of phloem sap δ13C as an indicator of canopy carbon discrimination. Tree Physiology, 2010, 30(12): 1499-1514. DOI:10.1093/treephys/tpq092 |
| [76] |
Werner C, Wegener F, Unger S, Nogués S, Priault P. Short-term dynamics of isotopic composition of leaf-respired CO2 upon darkening: measurements and implications. Rapid Communications in Mass Spectrometry, 2009, 23(16): 2428-2438. DOI:10.1002/rcm.4036 |
| [77] |
Lehmeier C A, Lattanzi F A, Gamnitzer U, Schäufele R, Schnyder H. Day-length effects on carbon stores for respiration of perennial ryegrass. New Phytologist, 2010, 188(3): 719-725. DOI:10.1111/j.1469-8137.2010.03457.x |
| [78] |
Bloemen J, Teskey R O, McGuire M A, Aubrey D P, Steppe K. Root xylem CO2 flux: an important but unaccounted-for component of root respiration. Trees, 2016, 30: 343-352. DOI:10.1007/s00468-015-1185-4 |
| [79] |
Aubrey D P, Teskey R O. Root-derived CO2 efflux via xylem stream rivals soil CO2 efflux. New Phytologist, 2009, 184(1): 35-40. DOI:10.1111/j.1469-8137.2009.02971.x |
| [80] |
Levy P E, Meir P, Allen S J, Jarvis P G. The effect of aqueous transport of CO2 in xylem sap on gas exchange in woody plants. Tree Physiology, 1999, 19(1): 53-58. DOI:10.1093/treephys/19.1.53 |
| [81] |
Teskey R O, McGuire M A. CO2 transported in xylem sap affects CO2 efflux from Liquidambar styraciflua and Platanus occidentalis stems, and contributes to observed wound respiration phenomena. Trees, 2005, 19(4): 357-362. DOI:10.1007/s00468-004-0386-z |
| [82] |
van der Heijden M G A, Martin F M, Selosse M A, Sanders I R. Mycorrhizal ecology and evolution: the past, the present, and the future. The New Phytologist, 2015, 205(4): 1406-1423. DOI:10.1111/nph.13288 |
| [83] |
Heijden M G A. Underground networking. Science, 2016, 352(6283): 290-291. DOI:10.1126/science.aaf4694 |
| [84] |
Epron D, Bahn M, Derrien D, Lattanzi F A, Pumpanen J, Gessler A, Högberg P, Maillard P, Dannoura M, Gérant D, Buchmann N. Pulse-labelling trees to study carbon allocation dynamics: a review of methods, current knowledge and future prospects. Tree Physiology, 2012, 32(6): 776-798. DOI:10.1093/treephys/tps057 |
| [85] |
陈昌华, 王晶苑, 魏杰, 温学发. 生态系统光合与呼吸拆分的同位素理论、方法和应用进展. 应用生态学报, 2022, 33(6): 1441-1450. |
| [86] |
Bowling D R, Ballantyne A P, Miller J B, Burns S P, Conway T J, Menzer O, Stephens B B, Vaughn B H. Ecological processes dominate the 13C land disequilibrium in a Rocky Mountain subalpine forest. Global Biogeochemical Cycles, 2014, 28: 352-370. DOI:10.1002/2013GB004686 |
| [87] |
Griffis T J, Black T A, Gaumont-Guay D, Drewitt G B, Nesic Z, Barr A G, Morgenstern K, Kljun N. Seasonal variation and partitioning of ecosystem respiration in a southern boreal aspen forest. Agricultural and Forest Meteorology, 2004, 125: 207-223. DOI:10.1016/j.agrformet.2004.04.006 |
| [88] |
Vickers D, Thomas C K, Martin J G, Law B. Self-correlation between assimilation and respiration resulting from flux partitioning of eddy-covariance CO2 fluxes. Agricultural and Forest Meteorology, 2009, 149(9): 1552-1555. |
| [89] |
Oikawa P Y, Sturtevant C, Knox S H, Verfaillie J, Huang Y W, Baldocchi D D. Revisiting the partitioning of net ecosystem exchange of CO2 into photosynthesis and respiration with simultaneous flux measurements of 13CO2 and CO2, soil respiration and a biophysical model, CANVEG. Agricultural and Forest Meteorology, 2017, 234/235: 149-163. |