 2023, Vol. 43
2023, Vol. 43文章信息
- 阎弘, 孙滢洁, 周婉莹, 刘滨辉
- YAN Hong, SUN Yingjie, ZHOU Wanying, LIU Binhui
- 大兴安岭不同纬度兴安落叶松生长对干旱适应性及生长衰退的差异
- Distinctions in drought adaptability and growth decline of Larix gmelinii at different latitudes in Greater Khingan Mountains
- 生态学报. 2023, 43(10): 3958-3970
- Acta Ecologica Sinica. 2023, 43(10): 3958-3970
- http://dx.doi.org/10.5846/stxb202112283696
-
文章历史
- 收稿日期: 2021-12-28
- 采用日期: 2023-01-13

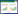




IPCC第六次报告指出由于人类活动导致全球未来气温将持续上升, 随之而来的极端事件频发将严重影响人类社会和自然生态系统[1]。据估计45%的陆地碳储存在森林中[2], 保持森林生态系统的健康和稳定有助于减少二氧化碳浓度的增加从而减缓全球气候变暖进程, 以往的研究证明极端干旱对森林生态系统破坏极为严重[3—4], 是引发森林大面积衰退甚至死亡的重要因素之一[5—6]。森林对干旱响应特征受林分结构、空间位置、树种特性影响[7—9], 了解不同地区的森林和树木如何抵御干旱并在干旱事件后恢复的能力, 是确定干旱将如何影响森林生态系统稳定性及碳储存能力的关键问题。
森林有一定承受、适应气候变化的能力, 但极端气候事件会导致树木恢复能力减弱甚至丧失[10—11], 造成树木生长下降并增加森林死亡率[12]。目前通过树木年轮学方法建立的林分或单木径向生长时间序列, 可有效评估树木对干旱事件的适应能力[11], 辨析森林生长衰退的特征和过程[13]。研究表明树木对干旱的适应能力受多种因素及其相互作用的影响, 包括干旱特征、树种、径级、年龄、竞争、海拔以及纬度等生物与非生物因素[7, 14—17], 尤其是在纬度梯度上的空间变异性最为明显[18]。研究发现裸子植物和被子植物对干旱的抵抗力和恢复弹力方面表现出不同的空间格局[8]; 北半球树木对干旱的抵抗力随纬度、土壤湿度和坡度的增加而增强, 对干旱的恢复力随纬度、土壤湿度的增加而降低[7], 来自干燥地区的树木对干旱的抵抗力较弱, 但比来自潮湿地区的树木恢复得更快[9]。目前多数报道都基于国际年轮数据库从林分尺度进行分析, 比较不同地区单木个体对干旱适应能力以及出现生长衰退程度的研究较少, 单木尺度的研究可以更加深入了解环境因素对生长—气候响应调节作用[19]。
大兴安岭是我国气候变暖最为剧烈的地区, 气候变暖导致干旱事件频率显著增加[20—21]。兴安落叶松(Larix gmelinii)是大兴安岭地区森林主要的建群种, 是欧亚大陆北方针叶林的一部分。通过模型预测发现, 气候变暖将引起大兴安岭地区兴安落叶松林生长下降并导致分布区逐渐北移, 甚至可能北移出我国境内[22—24]。目前对兴安落叶松的研究还集中在生长对气候因子的响应, 包括生长对气候响应空间规律以及生长对气候响应稳定性等方面。研究发现纬度、坡向、小地形对兴安落叶松生长与气候响应有影响[25—27], 如韩艳刚等人发现气候变化改变了兴安落叶松树木生长与气候的关系, 而且具有明显的空间差异特征[28], 兴安落叶松生长明显受区域水热条件共同控制[29], 当年6月和7月温度是影响兴安落叶松生长的重要因素[20]。兴安落叶松如何适应干旱以及生长衰退发生规律的认识还较少, 特别是在纬度梯度上, 阻碍了我们对未来森林动态的预测以及采取有效应对措施以减少干旱事件对这一地区森林生态系统功能和陆地碳平衡的影响。我们对不同纬度大兴安岭兴安落叶松林进行树木年轮学分析, 希望能够揭示以下科学问题: (1)兴安落叶松对干旱的适应能力在纬度梯度上有何差异; (2)树木对干旱的适应性与兴安落叶松出现生长衰退是否有联系及生长衰退在纬度梯度上有何差异; (3)不同纬度兴安落叶松对干旱的适应性特征是否受生物因素影响。
1 材料与方法 1.1 研究区概况研究区位于中国东北部大兴安岭地区, 在兴安落叶松分布范围内沿纬度梯度(47°17′—51°17′N)自南向北进行采样, 依次是阿尔山(AES)、库都尔(KDE)、大白山(DBS)(图 1; 表 1)。研究区属于寒温带大陆性季风气候, 冬季寒冷漫长, 夏季短暂而温暖, 阿尔山降水最少, 库都尔和大白山降水差异不明显(图 2)。三处采样点均为兴安落叶松天然林, 组成林分的主要树种为兴安落叶松, 主要伴生树种为白桦(Betula platyphylla)。

|
| 图 1 大兴安岭三个纬度兴安落叶松树木年轮采样点信息 Fig. 1 Sampling site information of treering at three latitudes in Greater Khingan Mountains PDSI: 帕尔默干旱指数 |
| 类型Type | 大白山 | 库都尔 | 阿尔山 |
| 树种species | 兴安落叶松Larix gmelinii | ||
| 纬度Latitude | 51°10′ N | 50°7′ N | 47°17′ N |
| 经度Longitude | 123°7′ E | 121°32′ E | 120°28′ E |
| 海拔Altitude/m | 1230 | 959 | 1112 |
| 时段(子样本信号强度>0.85) Time length(SSS>0.85) | 1943—2019 | 1941—2020 | 1920—2020 |
| 平均年龄Mean age/a | 74 | 68 | 58 |
| 平均胸径Mean DBH/cm | 19 | 27 | 31 |
| 平均树高Mean height/m | 12 | 12 | 16 |
| 平均冠幅Mean crown/m | 3.8 | 2.6 | 3 |
| 平均竞争指数Average competition index | 2.4 | 1.6 | 1.7 |
| 样本量(树/样芯)Sample size(tree/cores) | 28/42 | 52/93 | 45/90 |
| 样本总体代表性Express population signal(EPS) | 0.974 | 0.973 | 0.979 |
| 标准差Standard deviation (SD) | 0.3685 | 0.3388 | 0.2468 |
| 信噪比Signal to noise ratio (SNR) | 37.317 | 35.833 | 46.940 |
| 平均敏感度Mean sensitivity (MS) | 0.2192 | 0.1580 | 0.1982 |
| 一阶自相关Autocorrelation order 1 (AC1) | 0.7654 | 0.8514 | 0.5774 |
| 样芯间平均相关系数Mean interseries correlation(MC) | 0.483 | 0.521 | 0.494 |

|
| 图 2 大兴安岭不同纬度兴安落叶松采样点气象资料变化比较 Fig. 2 Comparison of meteorological data changes of Larix gmelinii sampling sites at different latitudes in Greater Khingan Mountains 经方差分析, 小写字母差异具有统计学意义(P < 0.05) |
在阿尔山、库都尔、大白山的兴安落叶松天然林中分别建立一处70m×70m远离人为干扰的样地, 对样地内所有乔木统一编号并测量记录其胸径、树高、冠幅(按树冠的正东、南、西、北), 对样地中心50m×50m区域内胸径超过10cm的兴安落叶松用生长锥在胸高1.3m处采集树芯, 每棵树采集两根树芯, 阿尔山共钻取45棵树90根样芯, 库都尔共钻取52棵树104根样芯, 大白山共钻取28棵树56根样芯, 样地基本情况见表 1。
分别对3个采样点的样芯在实验室中进行固定后打磨抛光, 直至可以在LINTAB 6.0年轮宽度分析仪(精度0.001mm)辨识, 在显微镜下目测交叉定年后利用年轮分析仪测得树轮原始宽度。利用COFFCHA程序[30]对定年和测量结果进行检验, 剔除不符合主序列的样本数据, 最终保留阿尔山90根样芯, 库都尔93根样芯, 大白山42根样芯。经过ARSTAN程序[31]中的负指数函数去除树木生长趋势, 用双权重平均法最终得到3个地区的林分标准年表(STD)。利用未标准化的年轮原始宽度计算每棵树的树轮断面积增量(Basal Area Increment, BAI)公式如下:
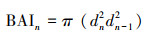
|
式中, d是树木半径, n是年份[32]。森林衰退是对林分的整体评价易采用林分年表, 而树木对干旱的适应性需从单木角度了解, 因此本研究计算适应性指标采用单木的树轮断面积增量(BAI)。
1.3 气象数据我国国家基准气象站是在1950左右开始建立的, 由于建站初期观测数据缺失较多, 一般大部分站点1960年以后数据质量开始稳定(降水数据1965年以后), 无法提供较长时间的与树木年轮相匹配的气象数据用以研究。本研究气象数据(1901—2020年平均气温、最高气温、最低气温、降水量和衡量干旱程度的帕尔默干旱指数(Palmer Drought Severity Index, PDSI), 均来源于荷兰皇家气象研究所数据共享网站(http://climexp.knmi.nl)的格点气象数据(CRUTS4.05格点数据集, 分辨率为0.5°×0.5°), 数据经过检验与研究区气候吻合[33—34], 说明两套数据的反映气候变化过程比较一致。年轮宽度指数与气候因子的相关分析, 由SAS软件中的皮尔逊(Pearson)相关分析完成, 阿尔山、库都尔和大白山的相关计算均在年表可靠区间内(SSS>0.85), 分别为1920—2020年、1941—2020年和1943—2019年。制图由Origin和ArcGis软件完成。
1.4 竞争分析为了量化每棵树的竞争压力, 我们使用了最常用的与距离、径级相关的竞争指数(CI)[35], 其公式为:
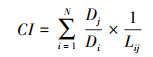
|
式中, CI是对象木i的竞争指数, Di是对象木i的胸径, Dj为竞争木j的胸径, Lij代表对象木i和竞争木j间的距离, N表示竞争木的数量。将取样树芯的树木作为对象木, 以对象木为圆心半径8m内, 径级大于10cm的树木作为竞争木, 测量对象木与竞争木间的距离与方位角。为避免对象木样圆超出样地范围, 本研究仅在样地中心50m×50m的区域中选取对象木, 其余空间作为缓冲区。
1.5 树木生长对干旱的适应性和生长衰退分析树木生长对极端干旱的抵抗力(Resistance, Rt)、恢复力(Recovery, Rc)和恢复弹力(Resilience, Rs)可以反应树木对干旱的适应能力。根据Lloret[11]等的方法进行计算其公式为:
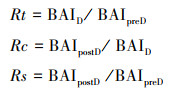
|
式中, Rt为干旱期间(低生长期间)与干旱发生之前树木生长的比值, Rt>1表示干旱期间树木生长没有发生下降, 树木对干旱的抵抗能力较强。Rc为干旱发生之后与干旱期间树木生长的比值, Rc>1表示相较于干旱期间, 干旱后树木生长水平恢复; Rs为干旱发生之后与之前树木生长的比值, Rs>1表示与干旱前相比, 干旱后树木完全恢复并加速生长。如果比值小于1结果则相反[7, 36]。式中, BAID表示干旱事件发生时的BAI, BAIpreD和BAIpostD表示干旱事件发生前3年和后3年的平均BAI(比较了3年、4年、5年的时间尺度发现了相似的结果, 本研究采用3年)[33]。
森林冠层绿度与径向生长在北美和欧洲都检测到存在广泛的干旱遗留影响, 前一个生长季后期或非生长季的缺水引发的树木形态或生理调整, 会影响树木在随后的干旱胁迫期间的响应特征[37—38]。前一年气候要素对兴安落叶松树木径向生长的“滞后效应”已经被证实存在, 例如高海拔兴安落叶松径向生长与前年生长季中后期的温度关系密切[39]。为分析树木生长对干旱事件的适应性, 参考国家《气象干旱等级》(GB/T20481—2006)的标准, 将上年6月至当年8月平均PDSI < -2的年份定义为干旱年, 若干旱持续一年以上则认定为一个干旱事件[33, 40]。
利用树木径向生长变化率判定兴安落叶松是否出现生长衰退[41—42], 计算公式如下:
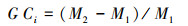
|
式中, Ci代表树木在第i年的前后各5年的径向生长变化率, 为避免个别年份生长特异性对计算结果的干扰, 以5年为一个窗口进行滑动平均, M1和M2各表示前5年(包括当年)与后5年(不包括当年)树轮宽度指数的平均值。径向生长变化率低于-25%表示出现了生长衰退, 超过75% 表示树木发生生长释放。
2 结果与分析 2.1 不同纬度的兴安落叶松树轮年表统计特征首先我们对年表统计特征进行检验, 如表 1所示, 三个地区的样本总体代表性(EPS)为0.973—0.979, 均高于树木年轮总体代表性阙值0.85, 样本所含信息能够代表总体特征。标准差(SD)和信噪比(SNR)均较高, 说明样本中包含了较多的气候和环境信息。一阶自相关(AC1)为0.5774—0.8514, 说明树木上一年的生长水平对当年有明显的影响。样芯间平均相关系数(MC)为0.483—0.512代表样芯间树轮宽度变化相对一致。总体而言, 三个纬度的年表统计特征值表明年表质量较高, 符合树木年轮学研究要求。
2.2 不同纬度兴安落叶松径向生长与气象因子的关系不同纬度的兴安落叶松标准年表与月降水量、帕尔默干旱指数(PDSI)、平均气温、平均最高气温和平均最低气温的相关关系如图 3所示, 阿尔山地区年表与当年7月降水, 当年7月和8月PDSI均呈显著正相关(P < 0.05), 与上年8月和当年7月平均最高气温呈显著负相关(P < 0.05)。库都尔地区年表与当年6月和7月降水, 当年7月和8月PDSI均呈显著正相关(P < 0.05), 与上年12月、当年4月、5月平均气温和平均最低气温, 当年5月平均最高气温呈显著正相关(P < 0.05), 与上年10月和当年7月平均最高气温呈显著负相关(P < 0.05)。大白山地区年表与当年3月和8月降水, 上年6月至当年8月的PDSI均呈显著正相关(P < 0.05), 与当年8月平均最高气温呈显著负相关(P < 0.05), 与上年9月和当年6月平均最低气温呈显著正相关(P < 0.05)。

|
| 图 3 不同纬度的兴安落叶松标准年表与气候因子的相关关系 Fig. 3 Correlation between the standard chronology of Larix gmelinii at different latitudes and climatic factors *号表示显著(P < 0.05), p表示上一年(Previous), c表示当年(current) |
由此可见, 研究区不同纬度的兴安落叶松生长均与当年树木生长旺盛的7月或8月最高气温呈显著负相关(P < 0.05)、与当年7月和8月PDSI显著正相关(P < 0.05), 生长季的高温和干旱是影响3个纬度地区兴安落叶松径向生长的主要因素。
2.3 不同纬度的兴安落叶松对干旱事件的适应性根据树轮宽度指数和单木BAI, 结合前一年6月至当年8月平均PDSI < -2, 阿尔山(AES)选出1951年、1968—1969年、1973年、2000—2011年和2017—2018年5个干旱事件, 库都尔(KDE)选出1945年、1951年、2004—2008年和2017—2018年4个干旱事件, 大白山(DBS)选出1975—1976年、1980年、2002—2003年和2017—2018年4个干旱事件(图 4)。为从单木角度比较不同纬度间适应性的差异, 我们将每棵树在不同干旱事件获得的值取平均, 获取每个地点每棵树的独特测量值(Rt、Rc、Rs), 并分析对比三个地区这些测量值的总体差异[7]。

|
| 图 4 不同纬度的兴安落叶松树轮宽度标准年表、BAI(树轮断面积增量)和前一年6月至当年8月平均PDSI Fig. 4 Standard chronology of ring width of Larix gmelinii at different latitudes, BAI(Basal Area Increment) and average PDSI from June of previous Year to August of current Year (1910—2020年) (1910—2020) |
图 5显示, 处于较低纬度地区的阿尔山对干旱的抵抗力(Rt)最强, 处于中间纬度地区的库都尔次之, 处于较高纬度地区的大白山最弱。恢复力(Rc)与抵抗力的结果相反, 大白山最强, 库都尔次之, 阿尔山最弱。恢复弹力(Rs)与抵抗力结果相似, 较低纬度地区的阿尔山最强, 中间纬度地区的库都尔次之, 较高纬度地区的大白山最弱。从树木样本的分布情况发现, 阿尔山和库都尔地区仅有少数树木样本Rt、Rc和Rs < 1, 说明研究区中、低纬度地区的兴安落叶松对干旱的适应能力较强。大白山地区的多数树木样本Rt和Rs < 1, 说明研究区高纬度地区的兴安落叶松在干旱期间生长下降明显, 干旱后生长依然没有达到干旱前的生长水平。

|
| 图 5 不同纬度的兴安落叶松对干旱事件的抵抗力、恢复力和恢复弹力 Fig. 5 Resistance, recovery and resilience to drought events in Larix gmelinii at different latitudes 经方差分析, 小写字母差异具有统计学意义(P < 0.05); 箱线图显示了中位数(水平线), 第25, 50和75四分位数 |
三个纬度地区的兴安落叶松适应性指标与主要生物因素的相关性结果(表 2)具有相似性。树木对干旱的抵抗力均与径级和冠幅呈显著负相关(P < 0.05), 恢复力与各生物因素间的相关性均不强, 恢复弹力与径级均表现为显著负相关(P < 0.05)。
| 采样点 Sampling site |
指标 Index |
抵抗力 Rt |
恢复力 Rc |
恢复弹力 Rs |
竞争 Competition |
径级 DBH |
树高 Hight |
冠幅 Crown |
年龄 Age |
| Rt | 1 | 0.01 | 0.89*** | 0.25 | -0.49*** | -0.31 | -0.51*** | -0.57 | |
| 阿尔山 | Rc | 1 | 0.35** | -0.09 | 0.12 | 0.18 | 0.06 | 0.12 | |
| Rs | 1 | 0.18 | -0.35** | -0.13 | -0.39** | -0.39** | |||
| Rt | 1 | -0.19 | 0.8*** | 0.06 | -0.4** | -0.32** | -0.33** | -0.52*** | |
| 库都尔 | Rc | 1 | 0.34** | 0.06 | 0.09 | 0.11 | 0.04 | 0.17 | |
| Rs | 1 | 0.02 | -0.27** | -0.19 | -0.19 | -0.35** | |||
| Rt | 1 | -0.19 | 0.84*** | 0.09 | -0.53** | -0.31 | -0.51** | -0.09 | |
| 大白山 | Rc | 1 | 0.05 | -0.02 | -0.19 | -0.27 | -0.25 | 0.16 | |
| Rs | 1 | 0.37 | -0.69*** | -0.55** | -0.52** | -0.09 | |||
| Rt: 抵抗力resistance; Rc: 恢复力recovery; Rs: 恢复弹力resilience; DBH: 胸径Diameter at breast height. Pearson相关, N表示样本量; **代表P<0.05, ***代表P<0.001 | |||||||||
综上所述, 兴安落叶松对干旱的适应能力存在纬度差异, 抵抗力和恢复弹力随纬度升高而降低, 恢复力随纬度升高而增加。大径级树木对干旱的抵抗力和恢复弹力更低, 对干旱的适应能力更弱。
2.4 不同纬度的兴安落叶松生长衰退差异兴安落叶松树木径向生长变化率(图 6)反映不同纬度地区树木出现了不同程度的生长衰退。阿尔山地区(AES)在1947—1948、1986—1989年期间出现生长衰退, 树木径向生长变化率最低达-37%。库都尔地区(KDE)仅在1941年出现生长衰退, 树木径向生长变化率达-27%, 1960、1985—1986、2002—2003和2010—2011年径向生长变化率接近-25%, 接近出现生长衰退。大白山地区(DBS)在1951、1987—1991、1996—2005、2006—2016和2018年期间出现生长衰退, 树木径向生长变化率最低达-48%, 特别在1980年以后大白山地区出现生长衰退的频率明显增加。

|
| 图 6 不同纬度的兴安落叶松径向生长变化率 Fig. 6 Change rates of radial growth of Larix gmelinii at different latitudes 虚线是75%和25%控制线 |
综上所述, 研究区范围内处在最高纬度的大白山地区出现生长衰退的频率和强度最高, 处于中间纬度和最低纬度的库都尔和阿尔山地区出现生长衰退较少。
3 讨论 3.1 不同纬度兴安落叶松径向生长对气象响应的差异研究区不同纬度的兴安落叶松生长与当年7月和8月PDSI显著正相关(P < 0.05), 与当年7月或8月最高气温显著负相关(P < 0.05)。7月份是树木径向生长最旺盛的月份, 树木蒸腾需要从土壤中汲取大量的水分。PDSI反映了降水和温度对土壤水分的综合影响, 生长季充足的水分保证了树木的各项生理活动及光合产物的积累, 有利于树木生长[43], 生长季的高温加剧了水分胁迫, 温度会加速土壤水分蒸发散失, 造成干旱而影响兴安落叶松的生长[44], 这都说明3个纬度地区的树木径向生长都受到了水分胁迫的抑制。大白山地区树木生长与上年6月至当年8月的PDSI均呈显著正相关(P < 0.05), AES和KDE的选取的采样点地形较为平坦, DBS选择在大兴安岭北部的最高峰-大白山的东南坡设置采样点。已有研究证实坡向对树木径向生长有显著影响[45], 北半球的南坡较北坡更加干燥, 而且越往北差异越明显[26], 因此DBS采样点的兴安落叶松径向生长与PDSI关系体现较强“遗留效应”特征。库都尔地区树木径向生长不仅受生长季降水和温度的影响, 还与当年4月、5月温度呈显著正相关(P < 0.05), 这一时期正值树木的生长季前, 大兴安岭地区4月和5月温度较低, 气温的回升有利于树木开始恢复各项生理活动, 高温可能有利于春季积雪融化, 从而加快形成层的活动促进树木生长[29]。阿尔山地区位于兴安落叶松分布区南界, 虽然降水较少但温度条件较好, 仅受生长季水分和高温影响。“分异现象”已经被广泛报道, 在全球持续变暖的背景下, 北方森林和温带森林对温度响应的敏感性逐渐减弱, 分布区北端或上限的树木生长由受低温限制逐渐向由水分限制转化[46]。关注是否发生干旱胁迫以及胁迫程度对于兴安落叶松生长和适应性影响, 对于研究气候变暖对北方林森林结构和群落演替影响具有重要意义。
3.2 不同纬度的兴安落叶松对干旱事件的适应性差异兴安落叶松对干旱的适应能力存在纬度差异, 抵抗力和恢复弹力随纬度升高而降低, 恢复力随纬度升高而增加。我们的研究结果表明, 干旱对兴安落叶松树木生产力的影响随气候梯度的变化而变化, 这印证了之前对欧洲温带森林和智利中南部地区的研究结果[47—48]。一般情况下纬度越高气候变暖越严重[49], 对水分胁迫越敏感, 处于较高纬度的大白山地区在干旱期间受到更强的干旱胁迫可能导致树木光合作用减弱造成碳亏缺出现生长大幅度下降, 抵抗力减弱[50]。在欧洲的类似研究也发现了这种模式, 干旱压力较大的森林对极端干旱的抵抗力更低, 更容易导致树木因水分胁迫而死亡[9]。然而, 对北半球森林的研究发现对干旱的抵抗能力随纬度增加而增加, 恢复弹力随海拔的升高而增加[7]。这些相异的结果证实了不同地区森林存在着不同的应对干旱的策略[7], 表明需要加强区域性尺度干旱适应性研究。抵抗力与恢复弹力呈正相关关系, 恢复力与抵抗力呈负相关关系, 这与其他研究结果一致[7, 11], 从年表与气候因子的响应结果看, 相较于研究区高纬度地区, 中、低纬度地区受到的干旱胁迫相对较低, 阿尔山地区位于兴安落叶松分布区的最南界, 降水较少但对干旱的适应能力强, 树木长期适应水分胁迫提高了水分利用效率。德国西南部森林的研究也发现, 易干旱地区的树木具有更高的抵抗能力[51]。研究发现来自较干燥地区的树木由于形成更耐旱的木质部所以应对干旱的能力更强[9]。
研究结果表明, 三个纬度地区的树木对干旱的抵抗力和恢复弹力与径级均表现为显著负相关(P < 0.05)由此推断, 大径级的树木对干旱的抵抗力和恢复弹力更低, 对干旱的适应能力更弱。相关研究报告同样发现, 树木径级的大小与对干旱的易损性呈正相关[52—54], 这可能与样木所处地区环境条件和其固有的生理特性有关。研究表明大径级树木对水分需求更多, 对干旱更加敏感[55], 大树更强的蒸腾作用和更长的水分输送路径, 加剧了干旱胁迫[56—57]。
3.3 不同纬度的兴安落叶松生长衰退差异研究结果表明研究区范围内高纬度地区更容易出现生长衰退, 中间纬度和最低纬度的库都尔和阿尔山地区出现生长衰退较少。以往的研究已经证明温度和降水是影响兴安落叶松纬度梯度径向生长差异性的重要因素[18], 帕尔默干旱指数(PDSI)反映了降水和温度对土壤水分的综合影响。通过三个纬度地区的年表与PDSI的关系表明最北的大白山地区的兴安落叶松对干旱更加敏感, 容易受干旱胁迫, 特别是在1980年快速升温以后出现生长衰退的频率明显增加。气候变暖以后干旱导致森林死亡事件已被广泛报道, 尤其在北纬地区树木生长会受干旱事件抑制, 并因水分供应不足或碳饥饿而导致死亡[6—8]。最近干旱引发的极端事件[33]增加了欧洲中部森林的脆弱性, 降低了森林树木的活力和生产力[58]。抵抗力低的树木更容易出现生长衰退[17], 我们的研究也发现研究区高纬度地区的树木对干旱的抵抗力和恢复弹力最弱, 这可能造成大白山地区的树木生长下降甚至死亡。中间、较低纬度的库都尔和阿尔山地区受干旱胁迫影响相对较小, 对干旱的抵抗力、恢复力和恢复弹力均较高, 干旱适应力强, 可能降低了出现生长衰退的频率。大白山地区后期出现生长衰退的时间与干旱事件时间基本吻合, 库都尔和阿尔山出现衰退时间与干旱时间不对应, 说明出现森林出现生长衰退不仅与树木对干旱适应能力有关还受当地的水热条件等因素综合影响。库都尔地区径向生长变化率多个年份接近-25%, 未来可能存在出现生长衰退的趋势, 尤其是在1980年气候快速变暖以后。有报道称1980年后智利中部树木的生长呈下降趋势, 这与该地区普遍的干旱和变暖趋势有关[47, 59]。
4 结论本文运用树木年轮学的方法, 通过对不同纬度的树木生长变化率和干旱适应性进行分析, 结果表明不同纬度地区的兴安落叶松对干旱具有不同的适应性特征。研究区不同纬度的兴安落叶松生长均与生长季中生长旺盛期的7, 8月的PDSI显著正相关(P < 0.05), 表明都受到了生长季水分胁迫的抑制。兴安落叶松对干旱的适应能力存在纬度差异, 抵抗力和恢复弹力随纬度升高而降低, 恢复力随纬度升高而增加。较高纬度地区的树木对干旱的适应能力较弱导致出现生长衰退的频率和强度最高, 处于中间、较低纬度地区兴安落叶松对干旱的适应能力较强出现生长衰退较少。三个纬度的兴安落叶松均表现出, 大径级的树木对干旱的抵抗力和恢复弹力更低, 对干旱的适应能力更弱。在气候变暖逐渐改变区域水分平衡特征背景下, 未来我们将结合水分梯度和温度梯度设置采样点, 增加采样密度, 以期更全面揭示不同生态梯度兴安落叶松树木径向生长对干旱适应性变化规律, 同时开展主要伴生树种对干旱的适应性和生态弹性研究, 为制定有针对性的应对气候变化森林管理与资源保护措施提供科学依据。
致谢: 感谢呼中、库都尔、阿尔山林场工作人员在树轮样芯采集中提供的支持与帮助。| [1] |
Masson-Delmotte V, Zhai P, Prtner H, Roberts D, Skea J, Shukla P R, Pirani A, Moufouma-Okia W, Péan C, Pidcock R. Global warming of 1.5 C. Global warming of 1.5 C. An IPCC Special Report on the impacts of global warming of, 2018, 1(5): 32. |
| [2] |
Bonan G B. Forests and climate change: forcings, feedbacks, and the climate benefits of forests. Science, 2008, 320(5882): 1444-1449. DOI:10.1126/science.1155121 |
| [3] |
Hartmann H, Moura C F, Anderegg W R L, Ruehr N K, Salmon Y, Allen C D, Arndt S K, Breshears D D, Davi H, Galbraith D, Ruthrof K X, Wunder J, Adams H D, Bloemen J, Cailleret M, Cobb R, Gessler A, Grams T E E, Jansen S, Kautz M, Lloret F, O'Brien M. Research frontiers for improving our understanding of drought‐induced tree and forest mortality. New Phytologist, 2018, 218(1): 15-28. DOI:10.1111/nph.15048 |
| [4] |
Yang Y, Saatchi S S, Xu L, Yu Y F, Choi S, Phillips N, Kennedy R, Keller M, Knyazikhin Y, Myneni R B. Post-drought decline of the Amazon carbon sink. Nature Communications, 2018, 9: 3172. DOI:10.1038/s41467-018-05668-6 |
| [5] |
Allen C D, Breshears D D, McDowell N G. On underestimation of global vulnerability to tree mortality and forest Die-off from hotter drought in the Anthropocene. Ecosphere, 2015, 6(8): art129. DOI:10.1890/ES15-00203.1 |
| [6] |
Allen C D, Macalady A K, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears D D, Hogg E H T, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim J H, Allard G, Running S W, Semerci A, Cobb N. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management, 2010, 259(4): 660-684. DOI:10.1016/j.foreco.2009.09.001 |
| [7] |
Gazol A, Camarero J J, Anderegg W R L, Vicente-Serrano S M. Impacts of droughts on the growth resilience of Northern Hemisphere forests. Global Ecology and Biogeography, 2017, 26(2): 166-176. DOI:10.1111/geb.12526 |
| [8] |
Li X Y, Piao S L, Wang K, Wang X H, Wang T, Ciais P, Chen A P, Lian X, Peng S S, Peñuelas J. Temporal trade-off between gymnosperm resistance and resilience increases forest sensitivity to extreme drought. Nature Ecology & Evolution, 2020, 4(8): 1075-1083. |
| [9] |
Sánchez-Salguero R, Camarero J J, Rozas V, Génova M, Olano J M, Arzac A, Gazol A, Caminero L, Tejedor E, de Luis M, Linares J C. Resist, recover or both? Growth plasticity in response to drought is geographically structured and linked to intraspecific variability in Pinus pinaster. Journal of Biogeography, 2018, 45(5): 1126-1139. DOI:10.1111/jbi.13202 |
| [10] |
Bréda N, Badeau V. Forest tree responses to extreme drought and some biotic events: towards a selection according to hazard tolerance?. Comptes Rendus Geoscience, 2008, 340(9/10): 651-662. |
| [11] |
Lloret F, Keeling E G, Sala A N. Components of tree resilience: effects of successive low-growth episodes in old ponderosa pine forests. Oikos, 2011, 120(12): 1909-1920. DOI:10.1111/j.1600-0706.2011.19372.x |
| [12] |
Guada G, Camarero J J, Sánchez-Salguero R, Cerrillo R M. Limited growth recovery after drought-induced forest dieback in very defoliated trees of two pine species. Frontiers in Plant Science, 2016, 7: 418. |
| [13] |
Fritts H C, Swetnam T W. Dendroecology: a tool for evaluating variations in past and present forest environments. Advances in Ecological Research, 1989, 19: 111-188. |
| [14] |
Anderegg W R L, Schwalm C, Biondi F, Camarero J J, Koch G, Litvak M, Ogle K, Shaw J D, Shevliakova E, Williams A P, Wolf A, Ziaco E, Pacala S. FOREST ECOLOGY. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science, 2015, 349(6247): 528-532. DOI:10.1126/science.aab1833 |
| [15] |
Gomes Marques I, Campelo F, Rivaes R, Albuquerque A, Ferreira M T, Rodríguez-González P M. Tree rings reveal long-term changes in growth resilience in Southern European riparian forests. Dendrochronologia, 2018, 52: 167-176. DOI:10.1016/j.dendro.2018.10.009 |
| [16] |
Merlin M, Perot T, Perret S, Korboulewsky N, Vallet P. Effects of stand composition and tree size on resistance and resilience to drought in sessile oak and Scots pine. Forest Ecology and Management, 2015, 339: 22-33. DOI:10.1016/j.foreco.2014.11.032 |
| [17] |
Sun S J, Lei S, Jia H S, Li C Y, Zhang J S, Meng P. Tree-ring analysis reveals density-dependent vulnerability to drought in planted Mongolian pines. Forests, 2020, 11(1): 98. DOI:10.3390/f11010098 |
| [18] |
Yasmeen S, Wang X, Zhao H, Zhu L, Yuan D, Li Z, Zhang Y, Ahmad S, Han S. Contrasting climate-growth relationship between Larix gmelinii and Pinus sylvestris var. mongolica along a latitudinal gradient in Daxing'an Mountains, China. Dendrochronologia, 2019, 58: 125645. DOI:10.1016/j.dendro.2019.125645 |
| [19] |
Rozas V. Individual-based approach as a useful tool to disentangle the relative importance of tree age, size and inter-tree competition in dendroclimatic studies. IForest-Biogeosciences and Forestry, 2015, 8(2): 187-194. DOI:10.3832/ifor1249-007 |
| [20] |
Jiang Y G, Zhang J H, Han S J, Chen Z J, Setälä H, Yu J H, Zheng X B, Guo Y T, Gu Y. Radial growth response of Larix gmelinii to climate along a latitudinal gradient in the greater khingan mountains, northeastern China. Forests, 2016, 7(12): 295. DOI:10.3390/f7120295 |
| [21] |
Chen H P, Sun J Q. Changes in drought characteristics over China using the standardized precipitation evapotranspiration index. Journal of Climate, 2015, 28(13): 5430-5447. DOI:10.1175/JCLI-D-14-00707.1 |
| [22] |
Xu D, Guo Q, Yan H. A study on the impacts of climate change on forests in China. Beijing: China Scientific and Technological Press, 1997.
|
| [23] |
张新时. 研究全球变化的植被-气候分类系统. 第四纪研究, 1993, 13(2): 157-169, 193. DOI:10.3321/j.issn:1001-7410.1993.02.006 |
| [24] |
程肖侠, 延晓冬. 气候变化对中国大兴安岭森林演替动态的影响. 生态学杂志, 2007, 26(8): 1277-1284. |
| [25] |
白学平, 常永兴, 张先亮, 马艳军, 吴涛, 李俊霞, 陈振举. 近30年快速升温对两种典型小地形上兴安落叶松径向生长的影响. 应用生态学报, 2016, 27(12): 3853-3861. |
| [26] |
刘欣, 刘滨辉. 大兴安岭不同坡向兴安落叶松径向生长对气候变化的响应. 东北林业大学学报, 2014, 42(12): 13-17, 21. DOI:10.3969/j.issn.1000-5382.2014.12.003 |
| [27] |
张朋磊. 兴安落叶松对气候变化响应的时间稳定性分析[D]. 哈尔滨: 东北林业大学, 2015.
|
| [28] |
韩艳刚, 盖学瑞, 邱思玉, 张月, 王守乐, 周莉, 于大炮. 大兴安岭兴安落叶松径向生长对气候响应的时空变化. 应用生态学报, 2021, 32(10): 3397-3404. |
| [29] |
张先亮, 崔明星, 马艳军, 吴涛, 陈振举, 丁玮航. 大兴安岭库都尔地区兴安落叶松年轮宽度年表及其与气候变化的关系. 应用生态学报, 2010, 21(10): 2501-2507. |
| [30] |
Holmes R L. Computer-assisted quality control in tree-ring dating and measurement, 1983.
|
| [31] |
Cook E. A time series analysis approach to tree ring standardization (dendrochronology, forestry, dendroclimatology, autoregressive process). Ph. D. Thesis. Tuscon, AZ, USA: The University of Arizona, 1985, 171.
|
| [32] |
Biondi F, Qeadan F. A theory-driven approach to tree-ring standardization: defining the biological trend from expected basal area increment. Tree-Ring Research, 2008, 64(2): 81-96. DOI:10.3959/2008-6.1 |
| [33] |
申佳艳, 李帅锋, 黄小波, 王绍武, 苏建荣. 金沙江流域不同海拔处云南松生态弹性及生长衰退过程. 林业科学, 2020, 56(6): 1-11. |
| [34] |
Zhong Z Q, He B, Guo L L, Zhang Y F. Performance of various forms of the palmer drought severity index in China from 1961 to 2013. Journal of Hydrometeorology, 2019, 20(9): 1867-1885. DOI:10.1175/JHM-D-18-0247.1 |
| [35] |
Hegyi F. A simulation model for managing jack-pine standssimulation. RoyalColl. For, Res. Notes, 1974, 30: 74-90. |
| [36] |
李晓琴, 张凌楠, 曾小敏, 王可逸, 王雅波, 路强强, 刘晓宏. 黄土高原中部针叶树与灌木径向生长对气候的响应差异. 生态学报, 2020, 40(16): 5685-5697. |
| [37] |
Kannenberg S A, Schwalm C R, Anderegg W R L. Ghosts of the past: how drought legacy effects shape forest functioning and carbon cycling. Ecology Letters, 2020, 23(5): 891-901. DOI:10.1111/ele.13485 |
| [38] |
Gazol A, Camarero J J, Sánchez-Salguero R, Vicente-Serrano S M, Serra-Maluquer X, Gutiérrez E, Luis M, Sangüesa-Barreda G, Novak K, Rozas V, Tíscar P A, Linares J C, Martínez del Castillo E, Ribas M, García-González I, Silla F, Camisón Á, Génova M, Olano J M, Hereş A M, Curiel Yuste J, Longares L A, Hevia A, Tomas-Burguera M, Galván J D. Drought legacies are short, prevail in dry conifer forests and depend on growth variability. Journal of Ecology, 2020, 108(6): 2473-2484. DOI:10.1111/1365-2745.13435 |
| [39] |
台秉洋. 大兴安岭北部高山兴安落叶松树木生长与气候变化的关系[D]. 哈尔滨: 东北林业大学, 2012.
|
| [40] |
中国气象局. 气象干旱等级: GB/T 20481-2006: 北京: 中国标准出版社, 2006.
|
| [41] |
Payette S, Filion L, Delwaide A. Disturbance regime of a cold temperate forest as deduced from tree-ring patterns: the Tantaré Ecological Reserve, Quebec. Canadian Journal of Forest Research, 1990, 20(8): 1228-1241. DOI:10.1139/x90-162 |
| [42] |
Nowacki G J, Abrams M D. Radial-growth averaging criteria for reconstructing disturbance histories from presettlement-origin oaks. Ecological Monographs, 1997, 67(2): 225-249. |
| [43] |
Bonan G B, Sirois L. Air temperature, tree growth, and the northern and southern range limits toPicea mariana. Journal of Vegetation Science, 1992, 3(4): 495-506. DOI:10.2307/3235806 |
| [44] |
Lévesque M, Siegwolf R, Saurer M, Eilmann B, Rigling A. Increased water-use efficiency does not lead to enhanced tree growth under xeric and mesic conditions. The New Phytologist, 2014, 203(1): 94-109. DOI:10.1111/nph.12772 |
| [45] |
张鹏, 张美淑, 吴玉德, 刘继生, 金春德. 不同坡向人工林赤松材性变异. 东北林业大学学报, 2003, 31(6): 13-15. DOI:10.3969/j.issn.1000-5382.2003.06.004 |
| [46] |
Babst F, Bouriaud O, Poulter B, Trouet V, Girardin M P, Frank D C. Twentieth century redistribution in climatic drivers of global tree growth. Science Advances, 2019, 5(1): eaat4313. DOI:10.1126/sciadv.aat4313 |
| [47] |
Urrutia-Jalabert R, Barichivich J, Rozas V, Lara A, Rojas Y, Bahamóndez C, Rojas-Badilla M, Gipoulou-Zuiga T, Cuq E. Climate response and drought resilience of Nothofagus obliqua secondary forests across a latitudinal gradient in south-central Chile. Forest Ecology and Management, 2021, 485: 118962. DOI:10.1016/j.foreco.2021.118962 |
| [48] |
Zang C, Hartl-Meier C, Dittmar C, Rothe A, Menzel A. Patterns of drought tolerance in major European temperate forest trees: climatic drivers and levels of variability. Global Change Biology, 2014, 20(12): 3767-3779. DOI:10.1111/gcb.12637 |
| [49] |
卢爱刚, 庞德谦, 何元庆, 庞洪喜, 院玲玲. 全球升温对中国区域温度纬向梯度的影响. 地理科学, 2006, 26(3): 345-350. |
| [50] |
孙振静, 赵慧颖, 朱良军, 李宗善, 张远东, 王晓春. 大兴安岭北部不同降水梯度下兴安落叶松生长对升温的响应差异. 北京林业大学学报, 2019, 41(6): 1-14. |
| [51] |
Kunz J, Lffler G, Bauhus J. Minor European broadleaved tree species are more drought-tolerant than Fagus sylvatica but not more tolerant than Quercus petraea. Forest Ecology and Management, 2018, 414: 15-27. |
| [52] |
Bennett A C, McDowell N G, Allen C D, Anderson-Teixeira K J. Larger trees suffer most during drought in forests worldwide. Nature Plants, 2015, 1: 15139. |
| [53] |
Rowland L, da Costa A C L, Galbraith D R, Oliveira R S, Binks O J, Oliveira A A R, Pullen A M, Doughty C E, Metcalfe D B, Vasconcelos S S, Ferreira L V, Malhi Y, Grace J, Mencuccini M, Meir P. Death from drought in tropical forests is triggered by hydraulics not carbon starvation. Nature, 2015, 528(7580): 119-122. |
| [54] |
Matusick G, Ruthrof K X, Fontaine J B, Hardy G E St J. Eucalyptusforest shows low structural resistance and resilience to climate change-type drought. Journal of Vegetation Science, 2016, 27(3): 493-503. |
| [55] |
Fang K Y, Chen D, Gou X, D'Arrigo R, Davi N. Influence of non-climatic factors on the relationships between tree growth and climate over the Chinese Loess Plateau. Global and Planetary Change, 2015, 132: 54-63. |
| [56] |
Mencuccini M, Martínez-Vilalta J, Vanderklein D, Hamid H A, Korakaki E, Lee S, Michiels B. Size-mediated ageing reduces vigour in trees. Ecology Letters, 2005, 8(11): 1183-1190. |
| [57] |
Ryan M G, Phillips N, Bond B J. The hydraulic limitation hypothesis revisited. Plant, Cell and Environment, 2006, 29(3): 367-381. |
| [58] |
Stojanović D B, Levanič T, Matović B, Orlović S. Growth decrease and mortality of oak floodplain forests as a response to change of water regime and climate. European Journal of Forest Research, 2015, 134(3): 555-567. |
| [59] |
Venegas-González A, Roig F A, Peña-Rojas K, Hadad M A, Aguilera-Betti I, Muñoz A A. Recent consequences of climate change have affected tree growth in distinct Nothofagus macrocarpa (DC.) FM vaz & rodr age classes in central Chile. Forests, 2019, 10(8): 653. |