 2022, Vol. 42
2022, Vol. 42文章信息
- 王悟敏, 匡雪韶, 胡佳瑶, 刘文胜
- WANG Wumin, KUANG Xueshao, HU Jiayao, LIU Wensheng
- 锰胁迫对盐肤木种子萌发、幼苗生长及理化特性的影响
- Effects of manganese stress on seed germination, seeding growth and physiological and biochemical characteristic of Rhus chinensis
- 生态学报. 2022, 42(9): 3706-3715
- Acta Ecologica Sinica. 2022, 42(9): 3706-3715
- http://dx.doi.org/10.5846/stxb202104261101
-
文章历史
- 收稿日期: 2021-04-26
- 网络出版日期: 2022-01-06





锰(Mn)是植物体所必需的微量元素。适量的锰在进行植物光合作用以及酶反应等新陈代谢中发挥重要作用[1]。然而, 环境中过量的锰会对植物产生毒害[2], 甚至通过食物链危害人类健康。锰矿开采与利用在带来经济利益的同时, 长期不合理的开采也导致矿区生态环境遭到严重破坏, 锰矿区污染土壤的治理与修复成了亟待解决的问题。植物修复技术是矿区重要的治理技术, 与传统技术相比, 植物修复具有成本低、效果久、环境友好等特点[3]。其中, 木本植物具有生物量较高、修复体系稳定的优势。筛选出适合的植物并揭示其抗胁迫机理是进行植物修复的关键。
种子萌发与幼苗生长是植物对环境最敏感的阶段[4]。逆境胁迫下, 种子的萌发与幼苗生长受到极大的影响, 植物的种子萌发情况与幼苗生理响应在一定程度上反映其抗逆性。研究表明, 低浓度的必需元素对植物的种子萌发有促进作用, 而高浓度胁迫对种子萌发产生抑制甚至毒害作用[5]。胁迫条件下植物的生长受到抑制, 生物量减少, 幼苗存活率降低[6]。同时, 幼苗会产生一系列生理响应机制来适应该逆境环境, 减轻毒害作用。例如, 锰胁迫下大豆(Glycine max)的超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性升高[7], 盐胁迫下小麦(Triticum aestivum)中渗透调节物质大量积累[8], 从而减轻胁迫产生的危害。当然, 随着胁迫强度的增加与胁迫时间的延长, 植物抗胁迫体系将受到破坏, 其生长受到抑制。如长时间高锰胁迫下青葙(Celosia argentea)[9]、美洲商陆(Phytolacca Americana)[10]幼苗叶片的光合色素合成受到抑制, SOD、POD、CAT活性降低, 渗透调节物质含量下降, 植物细胞受到损伤, 丙二醛(malondialdehyde, MDA)含量增加。因而, 进行植物种子萌发与幼苗生理的研究可揭示植物抗逆境胁迫的相关机理, 这是开展植物修复的关键环节。
盐肤木(Rhus chinensis)为漆树科(Anacardiaceae)盐肤木属(Rhus)落叶小乔木, 在我国大部分省区均有分布, 资源丰富。该植物能适应各种环境, 耐干旱、耐贫瘠, 抗逆性强, 在重金属铜、铬、铅胁迫下均表现出良好的耐受性, 是废弃地恢复的先锋植物[11-13]。盐肤木具有良好的观赏价值, 其虫瘿称“五倍子”, 是一种可供医药、墨水、鞣革和塑料等工业用的化工原料, 因而, 该植物也是一种重要的经济树种[14-15]。在湖南湘潭锰矿废弃地现场调查显示, 盐肤木在该区已自然恢复成林, 生长良好, 开花结实正常, 说明该植物对高锰污染土壤具有较好的适应能力。因而, 盐肤木具有较高的生态及经济价值, 在锰污染土壤修复中具有较高的应用潜力。然而, 目前对盐肤木在锰胁迫下的种子萌发、幼苗生长及生理生化响应机制尚未系统研究, 这限制了该植物在矿区修复中的推广应用。
本文以盐肤木作为研究对象, 设置锰浓度梯度开展盐肤木种子萌发、幼苗生长试验, 测定胁迫后不同时间盐肤木幼苗生理生化特征。其目的是揭示盐肤木抗胁迫机理, 为利用盐肤木进行重金属锰污染土壤的植物修复提供理论依据。
1 材料与方法 1.1 试验材料盐肤木种子在2019年11月采集于湖南省湘潭市锰矿废弃地(27°58′N, 112°51′E, 海拔26 m)。该区土壤锰含量达52319.25 mg/kg, 盐肤木种群平均高度约5.0 m。采集时, 随机选取50株盐肤木采摘其果实, 采回的果实储藏于实验室阴凉通风处。果实呈褐色时去除果皮及其他杂质, 将得到的种子装入种子袋中。种子千粒重为9.785 g;种子长为3 mm, 宽为4 mm。
1.2 试验方法 1.2.1 萌发试验根据王琼和宋桂龙[16]的研究, 盐肤木种子硬实率高达90.2%, 其种皮对于种子吸水有明显的机械阻碍作用。因而, 本研究采用徐莉清等[17]方法对种子进行酸蚀处理以提高种子萌发率, 即用98%浓硫酸浸泡90 min, 然后在0.5%的KMnO4溶液中浸泡30 min, 接着用去离子水彻底洗净。
萌发实验参照潘高等[18]方法进行, 即用MnCl2·4H2O设置1、5、10、15、20 mmol/L共5个Mn2+浓度, 以等量蒸馏水设置为对照组(CK)。每个培养皿25粒种子, 每个浓度6个重复, 将培养皿放入人工气候箱(温度25 ℃, 12 h光照+12 h黑暗)中进行萌发试验。每24 h换一次锰溶液与滤纸, 以保持胁迫浓度的恒定。每天定时记录种子萌发情况, 以胚根达到种子长度的1/2时为种子萌发的标志, 连续3天内没有新种子萌发时萌发结束。测定幼苗生物量, 计算种子发芽率、发芽势、发芽指数、活力指数与根冠比。
1.2.2 幼苗生长将预处理后的种子播种到装有150 mL干净纯沙的杯中, 放入人工气候箱中, 适量浇水, 伸出子叶后用1/2 Hoagland营养液培养盐肤木幼苗。待幼苗伸出5—6片真叶时, 设置与萌发试验相同的Mn2+浓度梯度, 对照组(CK)施以等量Hoagland营养液。幼苗培养在培养箱中进行, 期间每3 d浇一次营养液, 分别在胁迫后的第7、15、30天时测定幼苗叶片生理生化指标。
1.3 测定项目与方法 1.3.1 种子萌发指标测定

|
式中, Gt为在t日的发芽种子数;Dt为发芽天数;S为幼苗高度。
1.3.2 幼苗生物量测定萌发实验结束后, 每个Mn2+浓度随机选取20株幼苗, 用游标卡尺(0.1 mm)测量最长的根长、芽长。将所有幼苗(每个浓度梯度100株, 共计600株)洗净, 分成地上和地下两个部分装袋;于105 ℃杀青5 min, 再于烘箱中70 ℃烘干24 h直至恒重, 用万分之一天平测定生物量。
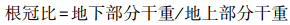
|
在胁迫第7、15、30天时取不同处理下盐肤木幼苗新鲜叶片进行测定, 各个生理指标做3个重复。
(1) 叶绿素含量
取新鲜叶片0.2 g用80%丙酮研磨提取, 上清液在紫外分光光度计下测646 nm、663 nm和470 nm处的吸光值, 按Chavoushi等[19]的方法计算叶绿素含量(mg/g FW(鲜重))。
(2) 抗氧化酶活性
参照Gao等[20]的方法, 取新鲜叶片0.1 g于预冷的研钵中, 用5 mL的磷酸盐缓冲液(0.05 mol/L, pH 7.8)将叶子研磨至匀浆, 于4 ℃下8000 r/min离心15 min, 取上清液待测。SOD活性采用氮蓝四唑(NBT)法测定, 以抑制NBT光还原的50%为一个SOD酶活性单位(U/g FW);POD活性采用愈创木酚法测定, 以每分钟内470 nm处吸光值升高0.01为一个POD酶活性单位(U g-1 min-1 FW);CAT活性采用紫外吸收法测定, 以每分钟内240 nm处吸光值减少0.01为一个CAT酶活性单位(U g-1 min-1 FW)。
(3) 渗透调节物质含量
参照等Li[21]的方法, 取新鲜叶片0.2 g。可溶性糖含量用蒽酮比色法测定, 在紫外分光光度计下测620 nm处的吸光值;可溶性蛋白含量用考马斯亮蓝G—250法测定, 以牛血清白蛋白为标准, 在紫外分光光度计下测595 nm处的吸光值;脯氨酸含量用酸性茚三酮显色法测定, 使用3%磺基水杨酸溶液5 mL, 在沸水浴中提取10 min, 冷却后过滤至干净管子。将2 mL滤液与2 mL酸性茚三酮和2 mL冰醋酸混合, 混合物在沸水浴中加热30 min。冷却后加入4 mL甲苯, 于3000 r/min离心5 min。以甲苯作为空白, 上层液在紫外分光光度计下测520 nm处的吸光值。
(4) MDA含量
参照Gao等[20]的方法, 取新鲜叶片0.2 g, 加入石英砂和10%三氯乙酸磨成匀浆状, 于3000 r/min离心10 min。取2 mL提取液, 以等量蒸馏水作对照, 加入0.67% TBA溶液2 mL混匀, 沸水浴15 min, 冷却后再离心。使用紫外分光光度计在450 nm、532 nm和600 nm处测量上清液。
1.3.4 数据处理利用Excel 2016统计数据, 计算平均值与标准差。采用SPSS 22.0进行ANOVA单因素方差分析, 运用LSD多重比较法(显著性水平设为0.05)对不同浓度处理下的各项生理生化指标进行差异性分析。利用Sigma Plot 12.5进行绘图。
2 结果与分析 2.1 不同浓度锰胁迫对盐肤木种子萌发与幼苗生长的影响萌发试验开始后第2天发现有种子开始萌发, 第13天后种子大部分种子已萌发完毕。锰胁迫下盐肤木种子发芽率在80.0%—81.6%之间, 不同Mn2+浓度下种子发芽率差异不显著。种子发芽势、发芽指数与活力指数则呈先升后降的趋势。Mn2+浓度为1 mmol/L时种子发芽势、发芽指数与活力指数最高;20 mmol/L时最低, 分别比CK下降19.05%、9.65%、85.70%(表 1)。
| Mn2+浓度 Mn2+ Concentration/(mmol/L) |
发芽率 Germination/% |
发芽势 Germination energy/% |
发芽指数 Germination index |
活力指数 Vigor index |
| 0 | 81.3%±0.08a | 53.3%±0.09ab | 26.3±2.06ab | 1562.6±385.40a |
| 1 | 80.0%±0.05a | 62.7%±0.14a | 28.2±3.21a | 1791.2±291.92a |
| 5 | 80.0%±0.06a | 52.0%±0.11ab | 26.2±2.64ab | 1037.2±533.02b |
| 10 | 81.3%±0.08a | 48.7%±0.09b | 24.6±3.52b | 883.4±98.41bc |
| 15 | 81.6%±0.04a | 47.2%±0.13b | 24.6±2.44b | 573.2±179.50c |
| 20 | 81.5%±0.08a | 43.2%±0.09b | 23.8±2.13b | 223.5±28.58c |
| 同列数据中不同的小写字母代表不同浓度之间差异显著(P < 0.05) | ||||
根长、幼根干重、幼苗干重、总干重等呈先升后降的趋势。总干重在Mn2+浓度为1 mmol/L时最大, 10—20 mmol/L处理下比CK显著下降(P < 0.05), 20 mmol/L时比CK下降23.98%, 说明在高浓度时盐肤木幼苗生长受到抑制。根长、芽长、幼根干重、幼苗干重表现出相同的趋势(表 2)。
| Mn2+浓度 Mn2+Concentration/ (mmol/L) |
根长 Root length/mm |
芽长 Shoot length/mm |
幼根干重 Root dry weight/ (mg/株) |
幼苗干重 Shoot dry weight/ (mg/株) |
总干重 Total dry weight/ (mg/株) |
根冠比 Root-shoot ratio |
| 0 | 59.3±13.4a | 15.5±1.4a | 1.01±0.20a | 3.40±0.33ab | 4.42±0.40a | 0.30±0.06a |
| 1 | 63.7±7.8a | 16.7±0.8a | 1.07±0.13a | 3.36±0.16ab | 4.43±0.24a | 0.32±0.04a |
| 5 | 40.2±16.7b | 17.1±1.7a | 0.80±0.33b | 3.60±0.39a | 4.41±0.44a | 0.23±0.10b |
| 10 | 36.5±5. 8b | 16.5±1.4a | 0.65±0.11bc | 3.23±0.27ab | 3.88±0.33b | 0.20±0.03bc |
| 15 | 22.6±7.1c | 16.6±2.4a | 0.46±0.15c | 3.21±0.46ab | 3.68±0.40bc | 0.15±0.06c |
| 20 | 9.2±0.6d | 16.3±2.1a | 0.26±0.02c | 3.10±0.39b | 3.36±0.41c | 0.09±0.01c |
胁迫7 d时, 盐肤木叶片中叶绿素a、叶绿素b含量、叶绿素a/b呈现先增后降的趋势。叶绿素a、叶绿素b含量在Mn2+浓度为10 mmol/L时最高, 比CK增加6.84%、4.92%, 叶绿素a/b基本保持不变;类胡萝卜素含量呈现出增加的趋势, Mn2+浓度为15、20 mmol/L时均显著高于CK(P < 0.05), 20 mmol/L时比CK增加17.50%(表 3)。
| 处理后天数 Days after treatment/d |
Mn2+浓度 Mn2+ Concentration/ (mmol/L) |
叶绿素a Chlorophyll a/ (mg/g FW) |
叶绿素b Chlorophyll b/ (mg/g FW) |
类胡萝卜素 Carotenoid/ (mg/g FW) |
叶绿素a/b Chlorophyll a/b |
| 7 | 0 | 1.90±0.04b | 0.61±0.02b | 0.40±0.03b | 3.13±0.11a |
| 1 | 1.96±0.10ab | 0.61±0.02ab | 0.40±0.05b | 3.20±0.12a | |
| 5 | 1.92±0.08ab | 0.60±0.01b | 0.38±0.05b | 3.23±0.13a | |
| 10 | 2.03±0.02a | 0.64±0.01a | 0.45±0.01ab | 3.18±0.05a | |
| 15 | 1.88±0.09b | 0.61±0.02b | 0.46±0.01a | 3.07±0.08a | |
| 20 | 1.83±0.04b | 0.59±0.02b | 0.47±0.03a | 3.04±0.04b | |
| 15 | 0 | 2.02±0.10b | 0.60±0.01ab | 0.42±0.05ab | 3.35±0.08ab |
| 1 | 2.17±0.06a | 0.62±0.04a | 0.43±0.03ab | 3.49±0.15a | |
| 5 | 2.09±0.17ab | 0.62±0.05a | 0.45±0.02a | 3.37±0.29ab | |
| 10 | 1.94±0.02b | 0.63±0.10a | 0.45±0.04a | 3.10±0.42b | |
| 15 | 1.71±0.02c | 0.54±0.03b | 0.47±0.02a | 3.16±0.10b | |
| 20 | 1.65±0.06c | 0.52±0.01b | 0.39±0.02b | 3.15±0.11b | |
| 30 | 0 | 2.00±0.04b | 0.58±0.06a | 0.49±0.01a | 3.46±0.31ab |
| 1 | 2.22±0.04a | 0.62±0.01a | 0.50±0.02a | 3.58±0.02a | |
| 5 | 1.89±0.06c | 0.60±0.09a | 0.46±0.05a | 3.18±0.37ab | |
| 10 | 1.46±0.02d | 0.47±0.05b | 0.40±0.01b | 3.14±0.33ab | |
| 15 | 1.42±0.04d | 0.46±0.03b | 0.39±0.03b | 3.06±0.13b | |
| 20 | 1.41±0.01d | 0.46±0.06b | 0.38±0.01b | 3.13±0.39b | |
| FW: 鲜重Fresh weight | |||||
胁迫15 d时, 盐肤木叶片中叶绿素a、叶绿素b含量呈先增后降的趋势, Mn2+浓度为1 mmol/L时最高, 15、20 mmol/L时显著降低(P < 0.05), 20 mmol/L时分别比CK下降18.32%、13.33%, 叶绿素a/b变化不显著。类胡萝卜素含量呈现先增后降的趋势, Mn2+浓度为15 mmol/L时最高, 比CK增加11.90%, 20 mmol/L时最低, 比CK下降7.14%。
胁迫30 d时, 盐肤木叶片中叶绿素a、叶绿素b含量呈先增后降的趋势, Mn2+浓度为20 mmol/L时最低, 分别比CK下降29.50%、20.68%, 叶绿素a/b变化不显著。类胡萝卜素含量呈现下降的趋势, Mn2+浓度为10—20 mmol/L时比CK显著降低(P < 0.05), 20 mmol/L时比CK下降22.45%。
2.3 锰胁迫对盐肤木叶片中抗氧化酶活性的影响胁迫7、15 d时, 盐肤木叶片中SOD活性均显示出随Mn2+浓度的升高而升高的趋势, Mn2+浓度为20 mmol/L时达到最高, 分别为CK的1.40、1.72倍。胁迫30 d时, 盐肤木叶片中SOD活性在Mn2+浓度小于为10 mmol/L时随浓度的升高而升高(P < 0.05), 20 mmol/L时SOD活性为CK的1.76倍(图 1)。

|
| 图 1 锰胁迫下盐肤木叶片SOD、POD、CAT活性 Fig. 1 SOD, POD and CAT activities of R. chinensis leaves under manganese stress SOD: 超氧化物歧化酶Superoxide dismutase; POD: 过氧化物酶Peroxidase; CAT: 过氧化氢酶Catalase; FW: 鲜重Fresh weight; 柱状图上不同的小写字母代表不同浓度之间差异显著(P < 0.05) |
胁迫7 d时, 盐肤木叶片中POD活性随Mn2+浓度的升高而升高, Mn2+浓度为20 mmol/L时达到最高, 为CK的11.21倍。胁迫15、30 d时, POD活性随Mn2+浓度升高呈现出先升高再下降的趋势。胁迫15 d时, Mn2+浓度为10 mmol/L时达到最高, 为CK的4.96倍;胁迫30 d时, Mn2+浓度为5 mmol/L时达到最高, 为CK的3.64倍, 均显著高于20 mmol/L时的POD活性(P < 0.05)(图 1)。
胁迫7 d时, 盐肤木叶片中CAT活性随Mn2+浓度的升高而升高, Mn2+浓度为20 mmol/L时达到最高, 为CK的1.95倍。胁迫15、30 d时, CAT活性随Mn2+浓度升高呈现出先升高再下降的趋势。胁迫15 d时, Mn2+浓度为15 mmol/L时达到最高, 为CK的3.28倍;胁迫30 d时, Mn2+浓度为10 mmol/L时达到最高, 为CK的4.39倍, 均显著高于20 mmol/L时的CAT活性(P < 0.05)(图 1)。
2.4 锰胁迫对盐肤木叶片中可溶性糖、可溶性蛋白、游离脯氨酸含量的影响胁迫7、15 d时, 盐肤木叶片中可溶性糖含量随Mn2+浓度的升高而升高, Mn2+浓度达到20 mmol/L时均显著高于CK(P < 0.05), 分别为CK的1.46、1.50倍。胁迫30 d时, 可溶性糖含量呈先升后降的趋势, Mn2+浓度为15 mmol/L时达到峰值, 为CK的1.55倍(图 2)。

|
| 图 2 锰胁迫下盐肤木叶片可溶性糖、可溶性蛋白、游离脯氨酸含量 Fig. 2 Contents of soluble sugar, soluble protein and free proline in leaves of R. chinensis under manganese stress |
胁迫7、15 d时, 盐肤木叶片中可溶性蛋白含量呈先升高后降趋势, Mn2+浓度为15 mmol/L时达到峰值, 分别为CK的2.40、2.39倍。胁迫30 d时, 与胁迫7、15 d时的趋势相同, 10 mmol/L时达到峰值, 为CK的2.14倍(图 2)。
胁迫7 d时, 盐肤木叶片中游离脯氨酸含量呈增加的趋势, 在Mn2+浓度为10—20 mmol/L时显著升高(P < 0.05), Mn2+浓度达到20 mmol/L时为CK的5.80倍。在胁迫15、30 d时, 游离脯氨酸含量呈先升后降的趋势, 15 mmol/L时达到峰值, 分别为CK的7.96、3.69倍(图 2)。
2.5 锰胁迫对盐肤木叶片中MDA含量的影响胁迫7 d时, 盐肤木叶片中MDA含量在不同Mn2+浓度处理之间没有显著差异。胁迫15 d时, MDA含量在Mn2+浓度为1、5 mmol/L处理下与CK之间没有显著差异, 10—20 mmol/L时MDA含量显著升高(P < 0.05)。胁迫30 d时, MDA含量在Mn2+浓度为5—20 mmol/L处理下均显著升高(P < 0.05), 分别为CK的2.69、4.31、5.73、7.03倍(图 3)。

|
| 图 3 锰胁迫下盐肤木叶片MDA含量 Fig. 3 MDA content of R. chinensis leaves under manganese stress MDA: 丙二醛Malondialdehyde |
植物种子的萌发情况可反映出植物在胁迫情况下的生命力。种子发芽势反映胁迫下植物种子的出芽速度与整齐度。发芽指数与活力指数反映污染物对种子发芽的胁迫情况。本文研究结果显示, 锰胁迫下盐肤木种子发芽率均达80%以上, 且不同Mn2+浓度下发芽率无显著差异。这与镉胁迫对不同品种糜子(Panicum miliaceum)的种子发芽率的影响[22]结果类似。这说明盐肤木种子具有一定的锰耐受性。而种子发芽势、发芽指数与活力指数则存在显著差异, 呈先升后降的趋势。这与锰胁迫下苍耳(Xanthium sibiricum)种子萌发[18]规律一致。Mn2+浓度为1 mmol/L时, 种子发芽势、发芽指数与活力指数均升高, 这是因为必需元素积累是植物种子发芽的先决条件。锰作为必需元素, 其在种子萌发过程中在种子胚根中积累, 可促进种子萌发[23]。
锰胁迫下盐肤木幼苗生长表现出随着锰浓度的升高而先升后降的现象。这与铅锌胁迫下毛竹(Phyllostachys pubescens)[5]、铅胁迫下金花茶(Camellia petelotii)[24]幼苗生长影响结果一致, 说明重金属对植物生长的低促高抑作用是一种普遍现象。本文研究结果显示, 锰胁迫对盐肤木幼苗地上部分影响较小, 而对根的抑制显著, 说明锰胁迫对盐肤木根的抑制大于芽。这与锰胁迫对紫花苜蓿(Medicago sativa)和地三叶草(Trifolium repens)[6]幼苗的影响相同。锰胁迫对盐肤木根部生长的影响显著高于对芽生长的影响, 这与铅胁迫下绿豆(Vigna radiata)种子及幼苗生长规律相似[25]。其原因是幼根作为直接吸收与转运土壤中重金属的器官, 与胁迫环境直接接触, 这使得其受到的毒害作用大于芽。
3.2 光合色素光合色素是植物进行光合作用时吸收、传递光能, 引起光化学反应的色素, 它能够较好反映植物光合作用的能力[26]。本文研究结果显示, 盐肤木叶片的叶绿素a、叶绿素b含量均呈现随着Mn2+浓度升高而先升后降的趋势, 叶绿素a/b则变化不显著。胁迫7 d时, 在Mn2+浓度为10 mmol/L时出现明显的促进作用;胁迫15、30 d时, 1 mmol/L时促进, 15—20 mmol/L时出现抑制现象。这与锰胁迫下青葙[9]、镉胁迫下梭鱼草(Pontederia cordata)[27]叶片的叶绿素含量变化趋势一致。这是由于锰元素是光合色素形成的重要元素, 还直接参与光合作用中的光合放氧过程。适量的锰可以促进盐肤木叶片中叶绿素的合成, 但在高浓度锰胁迫下植物叶绿体正常结构遭到破坏, 叶绿素合成减少[28-29]。其次, Mn2+的结合能力较强, 过量的锰会抑制Mg2+、Fe2+的吸收, Mg2+和Fe2+是叶绿素的重要组成, Mg2+、Fe2+的吸收的减少使得叶绿素的合成减少, 光合能力减弱[30]。另外, 叶绿素a主要作为光合中心色素等参与光系统反应, 叶绿素b主要作为天线色素参与能量传递, 二者具有不同的功能。本文研究中, 盐肤木幼苗在锰胁迫下较好地保持了叶绿素a/b值, 说明其在高锰胁迫下仍能有效地利用叶绿体进行光合作用。
3.3 抗氧化酶活性SOD、POD、CAT是植物抗氧化系统的重要组成部分, 是植物逆境环境下氧化应激的关键酶, 它们可缓解或降低活性氧自由基对植物造成的伤害[31]。其中SOD能够将O2-转化为H2O2和O2, 而POD和CAT则可将H2O2分解为H2O和O2, 从而降低活性氧的含量。SOD、POD与CAT三种酶之间具有协同作用, SOD、POD、CAT的表达量高低、活性的强弱直接决定了植物的抗逆性高低。本文研究结果表明, 胁迫7 d时, 盐肤木叶片中SOD、POD、CAT活性随Mn2+浓度的升高而升高。这与低锰胁迫下大豆[7]、镉胁迫下小麦[32]叶片中SOD、POD、CAT活性在胁迫强度最高时活性较高规律相同。其原因是在植物受到胁迫时, 可通过提高SOD、POD、CAT等抗氧化酶活性和抗氧化物质含量的方式清除叶片细胞受胁迫产生的过量活性氧, 从而保护植物细胞膜结构, 维持植株正常代谢功能[33]。较高的SOD、POD与CAT活性表明盐肤木具有较高抗逆性能力。
胁迫15、30 d时, 高Mn2+浓度下SOD活性仍然较高;POD、CAT活性则明显下降。这与铬胁迫下盐肤木[14]、锰胁迫下野大豆(Glycine soja)[34]、镉与锌胁迫下烟草(Nicotiana tabacum)[35]抗氧化酶活性变化规律一致。这是由于SOD、POD、CAT为植物体内重要的抗氧化酶, 在一定程度胁迫时可提高其含量来应对胁迫环境。但随着胁迫时间的延长、强度的增加, SOD、POD、CAT活性受到一定程度的抑制, 说明过高浓度、过长时间的锰胁迫使盐肤木抗氧化体系遭到破坏, 从而削弱了其保护能力, 造成对植物体的损伤, 最终导致其生物量的降低。因而, 抗氧化酶系统通过调节酶活性来缓解胁迫环境, 是植物体响应逆境胁迫的一种适应机制。
3.4 可溶性糖、可溶性蛋白、游离脯氨酸含量在逆境环境中, 植物都会产生逆境胁迫导致的直接或者间接水分缺失, 其生理生化过程可以反映植物的应对机制, 此时植物体内会大量的积累可溶性糖、可溶性蛋白、脯氨酸等渗透调节物质来保持细胞内水分平衡, 以维持细胞正常生命代谢活动, 减轻胁迫对植物带来的伤害[36]。本文研究结果表明, 盐肤木叶片中可溶性糖、可溶性蛋白、游离脯氨酸含量在植物受到锰胁迫时随Mn2+浓度升高而升高, 但在长期、高浓度锰胁迫下其含量降低。这与锰胁迫下黄花草(Cleome viscosa)[37]、小飞蓬(Comnyza canadensis)、杠板归(Polygonum perfoliatum)、美洲商陆[38]的渗透调节物质含量变化趋势相同。脯氨酸和可溶性糖是2种重要的渗透调节物质, 脯氨酸具有很高的水溶性, 可溶性糖是合成有机溶质的碳架和能量来源, 它们可使水分的跨膜运输朝着有利于植物生长的方向发展。植物体内的可溶性蛋白大多是参与代谢的酶类, 其含量高低反映植物代谢强度;同时, 可溶性蛋白具较强的亲水性, 可提高细胞保水性能, 有效防止其脱水。在本研究中, 一定时间内随Mn2+浓度的升高, 盐肤木幼苗能够通过提高体内渗透调节物质含量来缓解锰毒害, 使其具有一定抗逆性。但随着胁迫强度的增强与胁迫时间的延长, 渗透调节物质合成受阻, 其含量减少, 这种渗透调节机制被破坏, 导致盐肤木幼苗受到一定伤害[39]。因而, 渗透调节物质的调节是植物应对锰胁迫的一种重要机制。
3.5 MDA含量MDA是植物细胞膜脂过氧化的最终产物。在逆境环境下, 植物细胞中的活性氧产生与清除平衡会被破坏, 自由基大量积累造成细胞膜脂过氧化, 植物体内MDA含量增加。因而, MDA含量高低可反映植物遭受毒害的程度, 同时也能体现植物抗逆性高低[10]。本文研究结果显示, 胁迫7 d或Mn2+浓度低于10 mmol/L时盐肤木幼苗叶片中MDA含量无明显变化;胁迫30 d或Mn2+浓度高于10 mmol/L时盐肤木幼苗叶片中MDA含量增加。这与中等盐水平下蓖麻(Ricinus communis)幼苗叶片中MDA含量没有显著增加[40], 锰胁迫下苍耳[41]、盐胁迫下越橘(Vaccinum corymbosum)[42]、马尾松(Pinus massoniana)[43]MDA含量随胁迫浓度增加均增加的趋势一致。这说明在短时间或较低锰浓度的胁迫时细胞未受到明显伤害, 盐肤木具有一定的抗逆能力。而当胁迫浓度过高或胁迫时间延长时, 盐肤木受到毒害作用, 叶片细胞遭受破坏, 细胞膜脂过氧化程度随Mn2+浓度的升高和胁迫时间的延长而加重, 从而在体内积累MDA。
本文研究结果表明, 盐肤木在锰胁迫下发芽率较高, 在Mn2+浓度为10 mmol/L时SOD、POD、CAT活性、渗透调节物种含量仍然较高, 说明该植物适应锰胁迫能力较强, 为修复锰污染土壤的候选植物。
4 结论锰胁迫下盐肤木种子发芽率较高, Mn2+浓度对发芽率影响较小;幼苗生长随着Mn2+浓度的增加而呈先升后降的趋势。盐肤木幼苗叶片中叶绿素a、叶绿素b含量在低Mn2+浓度下上升, 但随着胁迫程度的增加和胁迫时间的延长, 叶绿素a、叶绿素b含量均降低。短期或低Mn2+浓度下幼苗SOD、POD、CAT活性随Mn2+浓度的升高而增强;长期的高锰胁迫下POD、CAT活性下降。短期Mn2+浓度处理下可溶性糖、可溶性蛋白、游离脯氨酸含量增加, 长期高锰胁迫下则下降。随着胁迫程度的增加和胁迫时间的延长, MDA含量增加。本研究说明盐肤木具有较强的耐锰能力, 是锰矿区修复的理想候选植物, 它能通过产生渗透调节物质以及提高抗氧化酶活性等方式来适应锰胁迫环境。
| [1] |
Nagajyoti P C, Lee K D, Sreekanth T V M. Heavy metals, occurrence and toxicity for plants: a review. Environmental Chemistry Letters, 2010, 8(3): 199-216. DOI:10.1007/s10311-010-0297-8 |
| [2] |
Xie Q Q, Li Z J, Yang L M, Lv J, Jobe T O, Wang Q Q. A newly identified passive hyperaccumulator Eucalyptus grandis×E. urophylla under manganese stress. PLoS One, 2015, 10(9): e0136606. DOI:10.1371/journal.pone.0136606 |
| [3] |
Bolan N S, Park J H, Robinson B, Naidu R, Huh K Y. Phytostabilization: a green approach to contaminant containment. Advances in Agronomy, 2011, 112: 145-204. |
| [4] |
Nasr S M H, Savadkoohi S K, Ahmadi E. Effect of different seed treatments on dormancy breaking and germination in three species in arid and semi-arid lands. Forest Science and Practice, 2013, 15(2): 130-136. DOI:10.1007/s11632-013-0209-7 |
| [5] |
陈俊任, 柳丹, 吴家森, 李松, 晏闻博, 彭丹莉, 叶正钱, 王海龙. 重金属胁迫对毛竹种子萌发及其富集效应的影响. 生态学报, 2014, 34(22): 6501-6509. |
| [6] |
Hayes R C, Conyers M K, Li G D, Poile G J, Price A, McVittie B J, Gardner M J, Sandral G A, McCormick J I. Spatial and temporal variation in soil Mn2+ concentrations and the impact of manganese toxicity on lucerne and subterranean clover seedlings. Crop and Pasture Science, 2012, 63(9): 875-885. DOI:10.1071/CP12138 |
| [7] |
Santos E F, Santini J M K, Paixão A P, Júnior E F, Lavres J, Campos M, dos Reis A R. Physiological highlights of manganese toxicity symptoms in soybean plants: Mn toxicity responses. Plant Physiology and Biochemistry, 2017, 113: 6-19. DOI:10.1016/j.plaphy.2017.01.022 |
| [8] |
Poustini K, Siosemardeh A, Ranjbar M. Proline accumulation as a response to salt stress in 30 wheat (Triticum aestivum L.) cultivars differing in salt tolerance. Genetic Resources and Crop Evolution, 2007, 54(5): 925-934. DOI:10.1007/s10722-006-9165-6 |
| [9] |
尚伟伟, 刘杰, 张学洪, 余轲. 锰胁迫对青葙生长及叶绿素荧光特性的影响. 生态环境学报, 2013, 22(8): 1353-1357. DOI:10.3969/j.issn.1674-5906.2013.08.013 |
| [10] |
王佳, 罗学刚, 石岩, 焦阳, 杨圣. 锰胁迫对美洲商陆种子发芽及幼苗生长的影响. 环境科学与技术, 2014, 37(11): 47-52. |
| [11] |
李庆亭, 杨锋杰, 张兵, 张霞, 周广柱. 重金属污染胁迫下盐肤木的生化效应及波谱特征. 遥感学报, 2008, 12(2): 284-290. |
| [12] |
陈晶, 穆燕, 陈明, 李明浩, 赵薇薇, 汤才国, 黄胜威, 詹福如. 盐肤木对重金属铬胁迫的生理生化反应研究. 生态科学, 2017, 36(2): 26-31. |
| [13] |
Shi X, Wang S F, Wang D X, Sun H J, Chen Y T, Liu J F, Jiang Z P. Woody species Rhus chinensis Mill. seedlings tolerance to Pb: physiological and biochemical response. Journal of Environmental Sciences, 2019, 78(4): 63-73. |
| [14] |
高洁莹, 龚力民, 刘平安, 李斌, 李顺祥. 盐肤木属植物研究进展. 中国实验方剂学杂志, 2015, 21(8): 215-218. |
| [15] |
刘玉兰, 王小磊, 刘海兰, 田原. 盐肤木果及其果油与籽油品质比较. 食品科学, 2018, 39(20): 197-201. DOI:10.7506/spkx1002-6630-201820029 |
| [16] |
王琼, 宋桂龙. 盐肤木种子硬实与萌发特性研究. 种子, 2008, 27(4): 59-61. |
| [17] |
徐莉清, 舒常庆. 酸蚀处理促进盐肤木种子萌发的研究. 华中农业大学学报, 2007, 26(2): 243-245. DOI:10.3321/j.issn:1000-2421.2007.02.024 |
| [18] |
潘高, 张合平, 刘鹏, 刘文胜. 锰胁迫对苍耳种子萌发及幼苗生理生化特性的影响. 草业学报, 2017, 26(11): 157-166. DOI:10.11686/cyxb2017033 |
| [19] |
Chavoushi M, Najafi F, Salimi A, Angaji S A. Effect of salicylic acid and sodium nitroprusside on growth parameters, photosynthetic pigments and secondary metabolites of safflower under drought stress. Scientia Horticulturae, 2020, 259: 108823. DOI:10.1016/j.scienta.2019.108823 |
| [20] |
Gao S S, Wang Y L, Yu S, Huang Y Q, Liu H C, Chen W, He X Y. Effects of drought stress on growth, physiology and secondary metabolites of Two Adonis species in Northeast China. Scientia Horticulturae, 2020, 259: 108795. DOI:10.1016/j.scienta.2019.108795 |
| [21] |
Li B Y, Feng Y N, Zong Y Z, Zhang D S, Hao X Y, Li P. Elevated CO2-induced changes in photosynthesis, antioxidant enzymes and signal transduction enzyme of soybean under drought stress. Plant Physiology and Biochemistry, 2020, 154: 105-114. DOI:10.1016/j.plaphy.2020.05.039 |
| [22] |
张盼盼, 王小林, 郭亚宁, 边茂华, 薛灵聪, 张雄. 40个不同糜子品种种子萌发期耐镉性评价. 西北农业学报, 2020, 29(12): 1803-1813. DOI:10.7606/j.issn.1004-1389.2020.12.005 |
| [23] |
Eroglu S, Giehl R F H, Meier B, Takahashi M, Terada Y, Ignatyev K, Andresen E, Küpper H, Peiter E, von Wirén N. Metal tolerance protein 8 mediates manganese homeostasis and iron reallocation during seed development and germination. Plant physiology, 2017, 174(3): 1633-1647. DOI:10.1104/pp.16.01646 |
| [24] |
粟春青, 蒋霞, 亢亚超, 赵苹羽, 王凌晖, 滕维超. 金花茶幼苗对铅胁迫的生长生理响应. 森林与环境学报, 2019, 39(5): 467-474. |
| [25] |
段代祥, 刘俊华. 重金属铅胁迫对绿豆种子萌发及幼苗生长的抑制效应. 种子, 2021, 40(1): 84-87, 98-98. |
| [26] |
Chaves M M, Flexas J, Pinheiro C. Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Annals of Botany, 2009, 103(4): 551-560. DOI:10.1093/aob/mcn125 |
| [27] |
辛建攀, 李文明, 祁茜, 田如男. 镉对梭鱼草叶片保护酶活性、光合及荧光特性的影响. 草业学报, 2018, 27(10): 23-34. DOI:10.11686/cyxb2017463 |
| [28] |
Xue S G, Wang J, Wu C, Li S, Hartley W, Wu H, Zhu F, Cui M Q. Physiological response of Polygonum perfoliatum L. following exposure to elevated manganese concentrations. Environmental Science and Pollution Research, 2018, 25(1): 132-140. DOI:10.1007/s11356-016-8312-7 |
| [29] |
Chen M, Tang Y L, Ao J, Wang D. Effects of strontium on photosynthetic characteristics of oil seed rape seedlings. Russian Journal of Plant Physiology, 2012, 59(6): 772-780. DOI:10.1134/S1021443712060052 |
| [30] |
Pittman J K. Managing the manganese: molecular mechanisms of manganese transport and homeostasis. New Phytologist, 2005, 167(3): 733-742. DOI:10.1111/j.1469-8137.2005.01453.x |
| [31] |
Verma S, Dubey R S. Lead toxicity induces lipid peroxidation and alters the activities of antioxidant enzymes in growing rice plants. Plant Science, 2003, 164(4): 645-655. DOI:10.1016/S0168-9452(03)00022-0 |
| [32] |
Qin S Y, Liu H E, Nie Z J, Gao W, Li C, Lin Y H, Zhao P. AsA-GSH cycle and antioxidant enzymes play important roles in Cd tolerance of wheat. Bulletin of Environmental Contamination and Toxicology, 2018, 101(5): 684-690. DOI:10.1007/s00128-018-2471-9 |
| [33] |
Bhaduri A M, Fulekar M H. Antioxidant enzyme responses of plants to heavy metal stress. Reviews in Environmental Science and Bio/Technology, 2012, 11(1): 55-69. DOI:10.1007/s11157-011-9251-x |
| [34] |
文珂, 郭晓玉, 谭娜娜, 刘雅妮, 刘文胜. 锰胁迫对野大豆种子萌发及幼苗生理生化特征的影响. 种子, 2018, 37(3): 40-45. |
| [35] |
Lyubenova L, Nehnevajova E, Herzig R, Schröder P. Response of antioxidant enzymes in Nicotiana tabacum clones during phytoextraction of heavy metals. Environmental Science and Pollution Research, 2009, 16(5): 573-581. DOI:10.1007/s11356-009-0175-8 |
| [36] |
王芳, 常盼盼, 陈永平, 彭云玲, 方永丰, 王汉宁. 外源NO对镉胁迫下玉米幼苗生长和生理特性的影响. 草业学报, 2013, 22(2): 178-186. |
| [37] |
肖泽华, 李欣航, 潘高, 吴耀文, 杨灿鑫, 匡雪韶, 刘文胜. 锰胁迫对黄花草种子萌发及幼苗生理生化特征的影响. 草业学报, 2019, 28(12): 75-84. |
| [38] |
任立民, 刘鹏, 蔡妙珍, 徐根娣, 方旭燕, 程正新. 水蓼、小飞蓬、杠板归和美洲商陆对锰毒的生理响应. 水土保持学报, 2007, 21(3): 81-85. |
| [39] |
Hajlaoui H, El Ayeb N, Garrec J P, Denden M. Differential effects of salt stress on osmotic adjustment and solutes allocation on the basis of root and leaf tissue senescence of two silage maize (Zea mays L.) varieties. Industrial Crops and Products, 2010, 31(1): 122-130. |
| [40] |
Li G, Wan S W, Zhou J, Yang Z Y, Qin P. Leaf chlorophyll fluorescence, hyperspectral reflectance, pigments content, malondialdehyde and proline accumulation responses of castor bean (Ricinus communis L.) seedlings to salt stress levels. Industrial Crops and Products, 2010, 31(1): 13-19. |
| [41] |
薛晨阳, 高英美, 曲波. 镉胁迫对瘤突苍耳、苍耳及其杂合体种子萌发与幼苗生长的影响. 生态学报, 2020, 40(1): 345-355. |
| [42] |
乌凤章. 三个越橘品种对盐胁迫的生长和生理响应及耐盐性差异. 植物生理学报, 2019, 55(11): 1638-1646. |
| [43] |
宫杰芳, 周运超, 李小永, 冉欣. 马尾松幼苗生理指标对高锰胁迫的响应. 生态学杂志, 2012, 31(3): 520-525. |