 2022, Vol. 42
2022, Vol. 42文章信息
- 赵云峰, 赵彩云, 朱金方, 李飞飞, 杨秀清, 郭朝丹
- ZHAO Yunfeng, ZHAO Caiyun, ZHU Jinfang, LI Fiefie, YANG Xiuqing, GUO Chaodan
- 北京市典型公园外来入侵植物分布格局
- Distribution pattern of alien invasive plants in typical parks in Beijing
- 生态学报. 2022, 42(9): 3656-3665
- Acta Ecologica Sinica. 2022, 42(9): 3656-3665
- http://dx.doi.org/10.5846/stxb202104140971
-
文章历史
- 收稿日期: 2021-04-14
- 网络出版日期: 2022-01-06





2. 山西农业大学林学院, 晋中 030600
2. College of Forestry, Shanxi Agricultural University, Jinzhong 030600, China
生物入侵在生态学、环保、流行病学和粮食安全等许多学科的研究中处于前沿[1-2], 外来植物入侵已成为一个世界性的生态和经济问题, 其对生态环境的破坏已经成为生物多样性丧失的主要原因之一[3-4]。
随着发展中国家城市化进程的加快, 城市地区的建筑和其他硬表面大幅增加, 城市不断扩张, 越来越多的环境问题出现了。城市化改变了原有生境的结构特征, 加剧了栖息地的退化, 而新的栖息地对本地物种造成不同方面的限制[5], 导致了乡土物种的流失。在各类生态系统中, 外来入侵物种对本地物种的生存威胁越来越大[6], 城市绿地生态系统是一个非常脆弱的生态系统, 不同物种经过多年竞争和适应达到一个简单的动态平衡, 这种平衡是非常不稳定的, 而且随着人类对公园和保护区干扰活动的增加, 这种脆弱的平衡很容易被入侵性强的入侵物种打破[7-8]。城市公园不仅是休憩区域, 同时承担着城市生态屏障功能, 而北京市公园的入侵植物种数逐渐增加[9], 大面积的外来物种入侵不仅影响公园的景观格局, 造成管理成本增加, 同时一些入侵种还会危害人类健康[10], 因此保护公园绿地生态系统不被入侵是急需解决的生态问题。
北京是一座历史悠久的城市, 保护当地植物多样性, 增加城市绿化是城市健康发展过程中重要的一部分, 了解外来入侵植物就是首先需要完成的任务, 但是现在我国对城市的外来入侵植物的种类构成等方面的研究还比较少[11]。北京作为中国的心脏, 改革开放以来, 经济发展迅速, 生物入侵的风险也大大增加。入侵植物的扩散与人类活动息息相关[12], 一般情况入侵地的小环境改变, 入侵种入侵的几率就会大大增加, 调査中也发现了类似规律。研究显示人类活动可能是造成城市入侵植物扩散的主要原因[13], 但现有文献较多关注于北京市入侵植物本底资料的调查研究[14-15], 对北京地区入侵植物扩散因素研究相对较少, 同时对入侵植物分布格局的研究也只是集中在大尺度的区县水平分布上[16], 本文主要研究外来入侵植物在公园小尺度的分布格局。
外来植物的不断引入和种植一直是城市景观建设中存在的问题, 开展城市公园的外来植物的种类构成基础调查和入侵植物分布格局研究, 可以为城市小尺度的绿地生态系统的科学发展和外来入侵植物的管理防控提供基础资料。因此, 本文比较了北京市四个不同功能区公园入侵植物分布格局, 分析入侵植物的分布规律, 为小尺度绿地生态系统的外来植物入侵防控提供理论依据。
1 材料和方法 1.1 研究区概况北京市在北纬39°26′—41°03′, 东经115°25′—117°30′之间, 温带季风性气候, 年均温为8.5—12℃, 年均降水量470—640mm, 东西总距离160km, 南北总距离170km, 总面积16410 km2, 经济发展迅速, 人口聚集, 城镇化率达86.6%[17]。
依据北京城市功能发展规划, 以人口密度、经济发展情况和发展方向、距城中心距离等因子, 北京16个区划分为了4个区域(图 1), 分别为首都功能核心区, 包括东城区和西城区;城市功能拓展区, 包括朝阳区、丰台区、石景山区和海淀区;城市发展新区, 包括房山区、顺义区、通州区、昌平区和大兴区;生态涵养发展区, 包括门头沟区、怀柔区、平谷区、密云区和延庆区[18], 分别称为核心区、拓展区、近郊区、远郊区。

|
| 图 1 北京市公园入侵植物生活型分布 Fig. 1 Distribution of invasive plant life forms in parks of Beijing |
2020年7—9月在北京市开展入侵植物的调查, 以北京市4个功能区为研究区域, 19个典型公园为样点(表 1), 全面踏查和样方法结合, 选择干扰活动频繁的公园主道路、湖边和绿化区域设置10—15个1m×1m的样方, 记录样方内的草本植物种类、株数、盖度和高度(cm), 同时记录样方的海拔、经纬度、生境信息。本次调查共设计255个样方。入侵植物的识别和划分参考马金双[3]《中国入侵植物名录》完成。
| 样点 Sampling point |
占地面积/km2 Area |
功能区 Functional zone |
样方数 Number of samples |
分布区域 Distribution area |
| 北海公园 | 0.69 | 核心区 | 10 | 草坪、路边、林下 |
| 天坛公园 | 2.73 | 核心区 | 10 | 路边、林下 |
| 奥林匹克森林公园 | 6.8 | 拓展区 | 15 | 路边绿化带、湖边 |
| 翠湖国家城市湿地公园 | 157.16 | 拓展区 | 15 | 林下、湖边、路边、绿化带 |
| 西山国家森林公园 | 73.64 | 拓展区 | 15 | 路边、林下、苗圃 |
| 北宫国家森林公园 | 9.15 | 拓展区 | 15 | 河道、路边、林下、绿化带 |
| 世纪森林公园 | 1.4 | 拓展区 | 10 | 路边、农田、绿化带 |
| 圆明园公园 | 3.5 | 拓展区 | 10 | 路边、湖边、绿化带、苗圃 |
| 汉石桥湿地公园 | 0.19 | 近郊区 | 15 | 路边、林下、湖边、绿化带 |
| 通州大运河森林公园 | 7.13 | 近郊区 | 15 | 河边、河道、绿化带、林下 |
| 马驹桥湿地公园 | 7.84 | 近郊区 | 10 | 路边 |
| 南海子麋鹿苑公园 | 0.6 | 近郊区 | 10 | 路边 |
| 拒马河水生野生动物自然保护区 | 20.41 | 近郊区 | 15 | 农田、河边、路边、林下 |
| 大杨山国家森林公园 | 21.06 | 近郊区 | 15 | 路边、绿化带、河道、林下 |
| 百花山自然保护区 | 217.43 | 远郊区 | 15 | 林下、路边 |
| 野鸭湖湿地公园 | 68.73 | 远郊区 | 15 | 路边、林下、湖边 |
| 平谷四座楼自然保护区 | 8 | 远郊区 | 15 | 农田、路边 |
| 云蒙山国家森林公园 | 22.08 | 远郊区 | 15 | 路边、苗圃 |
| 北京喇叭沟原始森林公园 | 111.72 | 远郊区 | 15 | 路边、河道 |
(1)α多样性指数计算
计算Shannon-Wiener指数(H′)、Pielou均匀度指数(J′)、Simpson指数(D)和物种丰富度(S)[19]。
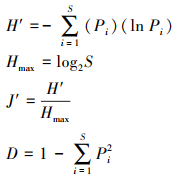
|
式中S为植物种数, 即物种丰富度, Pi为i物种的个体占所有物种个体总数的比例, Hmax为最大的物种多样性指数, N为总个体数量。
(2)重要值计算
采用综合优势度指数(Synthetic dominance index, SDR3i)衡量各草本植物在群落中的优势度[20]。
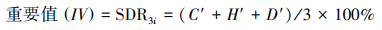
|
式中, SDR3i为物种的优势度, 即为物种的重要值; C′、H′、D′分别为相对盖度、相对高度、相对密度。
(3) 生态位宽度计算
采用Shannon-wiener生态位宽度(Bs)公式计算。
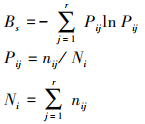
|
式中, Bs为物种i的生态位宽度, Pij为物种i在第j个资源状态下的重要值占该物种所有重要值总和的比例, nij是第i个物种在第j个资源状态下的重要值, Ni是第i物种的总重要值, r为样方数[21]。
(4) 入侵指数计算
计算入侵植物的入侵强度系数(Invasion intensity index, III)和群落可入侵性系数(Community invasibility index, CII) 以评估外来入侵植物的入侵强度以及群落可入侵性的程度。
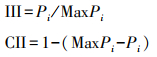
|
式中, Pi是调查样方内入侵植物的相对丰度, MaxPi为所有调查样方内入侵植物相对丰度最大值[22]。
1.4 数据处理方法使用Excel软件整理数据, 并计算物种的重要值;Past软件计算植物丰富度指数、Shannon-Wiener指数、Pielou均匀度指数、Simpson指数;R 3.6中Spaa程序包计算生态位宽度;采用SPSS 25软件Duncan法对入侵植物和本地植物在四个不同区域的多样性指数、入侵植物入侵强度系数和群落可入侵系数进行多重比较;Canoco 5软件进行主坐标分析(Principal coordinate analysis, PCoA)研究本地植物和入侵植物的群落结构;ArcGIS 10.2和GraphPad Prism 8软件作图。
2 结果与分析 2.1 北京市公园入侵植物种类和分布由图 1可知, 外来入侵植物在北京市拓展区的公园分布物种数最多(32种), 近郊区(31种)和远郊区(23种)次之, 核心区最少(8种)。本次调查共记录本地植物193种, 隶属于59科140属, 发现外来入侵植物40种(表 2), 隶属于12科29属, 物种数量最多的是菊科, 有20种, 占所有入侵植物总数的50%, 其次是苋科、大戟科和豆科, 各3种, 锦葵科、旋花科和茄科各有2种, 其他商陆科、大戟科、柳叶菜科、马鞭草科和藜科各包含1种入侵植物。生活型组成主要有一年生草本、二年生草本、多年生草本和灌木, 四个功能区皆是以一年生的草本植物居多, 其次是多年生草本(图 1)。所有样方中外来入侵植物出现频次最高的是牛膝菊(Galinsoga parviflora)为68次, 其次小蓬草(Erigeron canadensis)50次, 反枝苋(Amaranthus retroflexus)37次, 圆叶牵牛(Ipomoea purpurea)37次, 婆婆针(Bidens bipinnata)31次(图 2)。
| 物种Species | 科Family | 属Genus | 原产地Origin | 生活型Life form |
| 斑地锦Euphorbia maculata | 大戟科 | 大戟属 | 北美洲 | 一年生草本 |
| 齿裂大戟Euphorbia dentata | 大戟科 | 大戟属 | 北美洲 | 一年生草本 |
| 通奶草Euphorbia hypericifolia | 大戟科 | 大戟属 | 北美洲 | 一年生草本 |
| 白车轴草Trifolium repens | 豆科 | 车轴草属 | 欧洲、中亚和西亚 | 多年生草本 |
| 野西瓜苗Hibiscus trionum | 锦葵科 | 木槿属 | 非洲 | 一年生草本 |
| 苘麻Abutilon theophrasti | 锦葵科 | 苘麻属 | 印度 | 一年生亚灌木草本 |
| 牛膝菊Galinsoga parviflora | 菊科 | 牛膝菊属 | 南美洲 | 一年生草本 |
| 小蓬草Erigeron canadensis | 菊科 | 白酒草属 | 北美洲 | 一年生草本 |
| 大狼杷草Bidens frondosa | 菊科 | 鬼针草属 | 北美洲 | 一年生草本 |
| 豚草Ambrosia artemisiifolia | 菊科 | 豚草属 | 北美洲 | 一年生草本 |
| 意大利苍耳Xanthium italicum | 菊科 | 苍耳属 | 北美洲和南欧 | 一年生草本 |
| 苏门白酒草Erigeron sumatrensis | 菊科 | 白酒草属 | 南美洲 | 一年或二年草本 |
| 婆婆针Bidens bipinnata | 菊科 | 鬼针草属 | 美洲 | 一年生草本 |
| 加拿大一枝黄花Solidago canadensis | 菊科 | 一枝黄花属 | 北美 | 多年生草本 |
| 钻形紫菀Aster subulatus | 菊科 | 紫菀属 | 北美洲 | 一年生草本 |
| 鬼针草Bidens pilosa | 菊科 | 鬼针草属 | 美洲 | 一年生草本 |
| 三裂叶豚草Ambrosia trifida | 菊科 | 豚草属 | 北美洲 | 一年生草本 |
| 月见草Oenothera biennis | 柳叶菜科 | 月见草属 | 北美洲 | 二年生直立草本 |
| 马缨丹Lantana camara | 马鞭草科 | 马缨丹属 | 热带美洲 | 灌木或蔓性灌木 |
| 曼陀罗Datura stramonium | 茄科 | 曼陀罗属 | 墨西哥 | 草本或亚灌木状 |
| 苦職Physalis angulata | 茄科 | 酢浆属 | 南美洲 | 一年生草本 |
| 垂序商陆Phytolacca americana | 商陆科 | 商陆属 | 北美洲 | 多年生草本 |
| 凹头苋Amaranthus blitum | 苋科 | 苋属 | 热带美洲 | 一年生草本 |
| 反枝苋Amaranthus retroflexus | 苋科 | 苋属 | 美洲 | 一年生草本 |
| 皱果苋Amaranthus viridis | 苋科 | 苋属 | 南美洲 | 一年生草本 |
| 牵牛Ipomoea nil | 旋花科 | 牵牛属 | 南美洲 | 一年生草本 |
| 圆叶牵牛Ipomoea purpurea | 旋花科 | 牵牛属 | 美洲 | 一年生草本 |
| 铺地黍Panicum repens | 禾本科 | 黍属 | 欧洲南部 | 多年生草本 |
| 草木犀Melilotus officinalis | 豆科 | 草木樨属 | 西亚至南欧 | 二年生草本 |
| 紫苜蓿Medicago sativa | 豆科 | 苜蓿属 | 西亚 | 多年生草本 |
| 地毯草Axonopus compressus | 禾本科 | 地毯草属 | 热带美洲 | 多年生草本 |
| 金鸡菊Coreopsis basalis | 菊科 | 金鸡菊属 | 北美洲 | 一年生草本 |
| 剑叶金鸡菊Coreopsis lanceolata | 菊科 | 金鸡菊属 | 美国 | 一年生草本 |
| 秋英Cosmos bipinnatus | 菊科 | 秋英属 | 美国和墨西哥 | 一年生或多年生草本 |
| 鳢肠Eclipta prostrata | 菊科 | 鳢肠属 | 美洲 | 一年生草本 |
| 苦苣菜Sonchus oleraceus | 菊科 | 苦苣菜属 | 欧洲和地中海 | 一年或二年生草本 |
| 黑心金光菊Rudbeckia hirta | 菊科 | 金光菊属 | 北美洲 | 一年或二年生草本 |
| 天人菊Gaillardia pulchella | 菊科 | 天人菊属 | 美洲 | 一年生草本 |
| 金光菊Rudbeckia laciniata | 菊科 | 金光菊属 | 北美洲 | 多年生草本 |
| 小藜Chenopodium ficifolium | 藜科 | 藜属 | 欧洲 | 一年生草本 |

|
| 图 2 外来入侵植物记录频次 Fig. 2 Record frequency of invasive plants |
物种的优势度即物种重要值可以反映物种在群落中的竞争能力、资源利用能力, 选平均重要值最高的前10个物种为群落优势种, 将10种优势物种的优势度和生态位宽度做对比, 优势度比较结果从大到小排序依次为大狼杷草(Bidens frondose) (0.26)、凹头苋(Amaranthus blitum) (0.2)、圆叶牵牛(0.19)、牵牛(Ipomoea nil) (0.19)、牛膝菊(0.18)、反枝苋(0.15)、小蓬草(0.12)、婆婆针(0.11)、通奶草(Euphorbia hypericifolia) (0.11)、小藜(Chenopodium ficifolium) (0.1), 大狼杷草和凹头苋的优势度最大, 对资源的竞争能力强, 在群落中占有优势地位, 相反通奶草和小藜优势度较低, 竞争能力弱, 在群落中相对劣势。生态位宽度比较结果从大到小排序依次为小蓬草(2.77)、牛膝菊(2.51)、反枝苋(2.3)、凹头苋(2.11)、婆婆针(2.0)、圆叶牵牛(1.97)、牵牛(1.94)、大狼杷草(1.74)、通奶草(1.54)、小藜(1.51), 小蓬草的生态位宽度最大, 牛膝菊、反枝苋其次, 这几个物种对资源的利用能力较强, 对环境变化的耐受能力强、分布广泛、生态幅较广, 相反生态位宽度较窄的大狼杷草、通奶草和小藜, 对生境的局限性大、生态幅较窄。结果表明入侵植物优势度排序与生态位宽度排序趋势不同, 优势度最大的是大狼杷草, 生态位宽度却相对较窄, 生态幅最宽的小蓬草, 优势度排在第七位。
2.3 不同功能区入侵植物与本地植物多样性对比由图 3可知北京市不同功能区公园本地植物与外来入侵植物多样性指数表现出不同的分布格局。外来入侵植物丰富度在拓展区和近郊区都显著高于核心区和远郊区(P < 0.05), 而拓展区的本地植物丰富度显著低于核心区和远郊区(P < 0.05), 呈现明显相反的变化趋势;外来入侵植物Shannon-Wiener指数在拓展区和近郊区显著高于核心区和远郊区(P < 0.05), 本地植物Shannon-Wiener指数四个区域差异不显著, 其中拓展区最低;外来入侵植物Simpson指数在远郊区显著高于拓展区和近郊区(P < 0.05), 核心区与其他三区皆没有显著差异, 本地植物Simpson指数在四个区域之间均无显著差异(P>0.05);外来入侵植物Pielou指数在拓展区和近郊区显著高于核心区(P < 0.05), 核心区和远郊区差异不显著, 本地植物Pielou指数在仅近郊区与核心区之间具有显著差异(P < 0.05), 其余区都没有显著差异(图 3)。综合说明入侵植物在北京市拓展区和近郊区丰富度高且分布均匀, 在核心区和远郊区丰富度低且不均匀分布。

|
| 图 3 入侵植物与本地植物多样性对比 Fig. 3 Comparison of invasive and native plant diversity 不同小写字母表示不同功能区在0.05水平差异显著(P < 0.05); 图中数据为平均值±标准误 |
基于Bray-Curtis距离矩阵, 对北京市公园本地草本植物群落和外来入侵植物群落进行主坐标排序分析, 本地植物群落主坐标分析(图 4), 主成分1和主成分2对群落差异性的解释度分别是30.42%、18%。分析可知核心区与拓展区、近郊区、远郊区皆有差异, 与拓展区本地植物群落差异比较大, 但拓展区和近郊区排序比较分散, 与其他区没有明显差异。外来入侵植物群落主坐标分析(图 4), 主成分1和主成分2群落差异性的解释度分别39.62%、18.82%, 累计解释58.44%, 核心区与拓展区、近郊区和远郊区的入侵植物群落均有差异, 与拓展区的差异最大, 拓展区、近郊区和远郊区三者之间的差异都不明显, 特别是近郊区的入侵植物群落比较分散, 说明只有核心区公园植物群落组成与其他区有差异, 其余三个区均较为相似。

|
| 图 4 北京市公园本地植物和入侵植物主坐标分析 Fig. 4 Principal coordinate analysis of native plants and invasive plants in Beijing parks |
北京市各区域公园的群落可入侵性系数没有显著的区别(图 5), 系数最高的是核心区, 依次拓展区、近郊区、远郊区降低, 说明核心区的公园植物群落是最容易被入侵的, 拓展区公园入侵植物入侵强度系数显著高于核心区(P=0.008), 说明在拓展区的公园植物群落中入侵植物占据优势地位而核心区公园还没有入侵种大范围优势的状况, 并且其在各区的变化趋势与入侵植物的丰富度趋于相同。

|
| 图 5 各区公园入侵植物入侵强度系数和群落可入侵系数比较 Fig. 5 Comparison of Regional Parks Invasion intensity index and Community invasibility index 不同小写字母表示不同功能区在0.05水平差异显著(P < 0.05); 图中数据为平均值±标准误 |
本研究主要对北京市小尺度公园绿地的入侵植物分布格局进行研究, 通过实地调查统计北京市公园入侵植物组成和分布的数据。调查发现北京市各功能区公园均有入侵植物分布, 而且分布种类众多, 部分公园的入侵植物已经出现聚集性高密度分布状况, 严重侵占了乡土植物的生存空间, 成为了优势种, 改变了公园小尺度植物群落的组成结构, 从入侵植物入侵强度系数比较结果得出拓展区、近郊区和远郊区均比核心区的高, 原因是这三个功能区公园内入侵植物丰富度较高, 需及时对入侵植物进行防治。
北京市公园外来入侵植物中最多的是菊科, 与孙旭芳等[9]对北京市近郊区郊野公园调查结果一致, 一方面菊科植物以一年的草本居多, 草本植物具有结构简单、生活史短、繁殖方式多、成长消耗少等优势, 降温之后种子进入休眠状态可以避免死亡, 另一方面菊科草本植物的种子数量多个体小, 可以通过多种媒介方式传播, 它还有多种特别结构使其依附于动物的运动将种子带到更远的地方, 极大的提高了其繁殖扩散的效率[11]。美洲是北京市外来入侵植物的主要原产地, 根据研究显示来自美洲的外来植物在我国范围内生存分布具有基因优势[23], 北美与东亚地区存在地质历史关联因素导致其很容易传入我国东南沿海地区[24], 之后在国内传播, 最重要的中国近年来与美洲国家的进出口贸易达到鼎盛时期, 特别是中国与美国的贸易往来非常频繁, 这无疑为美洲植物入侵到中国提供了便利的传播方式和途径。
优势入侵植物的优势度和生态位宽度的排序结果存在差异, 与陈俊华等[25]的研究结果不一致, 与李一凡等对深圳市坝光湿地园银叶树群落优势种生态位特征的研究结果类似[26], 即生态位宽度和优势度并不是相同的变化趋势, 而出现优势度大的植物生态位宽度较窄。优势度是一个综合指标涵盖了物种的数量、频次和植株的高度, 而生态位宽度更能反应物种的分布范围和适应性[27]。本文调查的大狼杷草和凹头苋等物种植株高大, 群落建成初期有关键作用, 群落建成后也有支配作用, 占据绝对优势, 但是繁殖速度慢, 分布不广泛, 生态位宽度达不到最大, 小蓬草和牛膝菊等物种适应范围广, 同时繁殖速度快, 生态位宽度比优势种较大。
城市化背景下, 北京市的入侵植物种数多、入侵严重, 其分布与城市化的梯度有关, 从北京市的拓展区、近郊区再到远郊区公园, 入侵植物丰富度整体上与城市化梯度呈正相关, 越靠近城市中心, 入侵植物种数越多, 王苏铭等[28]研究认为这种结果可能是因为外来入侵植物在北京市频繁的人类干扰状况下逐渐适应并生存, 北京偏远地区影响入侵植物扩散的关键因素是环境因子, 即偏远地区的生境破碎导致入侵植物的扩散, 但是在半人工生态系统和人工生态系统中, 人类的频繁活动加快了外来入侵植物的入侵[29], 并且高干扰环境中广食性天敌对入侵植物的取食和抑制等的作用也降低。海云瑞等[30]研究北京市主要入侵植物的分布特征研究结果显示人口因素是影响北京市入侵植物分布的主要因素之一。王苏铭等[28]研究表明在北京山区入侵植物的分布与海拔有负相关关系, 在高海拔恶劣的环境条件下人类活动比较少[31], 同时大部分入侵植植物不适应高海拔气候的生存, 但是在平原区以及市中心区域发达便利的交通条件为入侵植物的入侵提供了有利的扩散条件[32]。有学者对北京市18个区域的外来归化植物进行了分布格局的研究, 结果表明经济发展迅速, 人口活动频繁, 交通设施完整的区域, 归化种的数目分布也比较多[33]。
入侵植物在人工生态系统和自然生态系统中的影响因素响应不一样, 在人工生态系统中的植物群落生态位不完整, 空位多, 容易被入侵植物占据[34], 同时入侵植物与本地植物相比, 没有天敌的克制, 化感抑制作用降低[35], 越是人工生态系统, 入侵植物更容易入侵成功, 所以核心区公园的群落可入侵系数最高, 但本文核心区和远郊区的比较结果是城市化梯度与公园入侵植物丰富度呈相反的变化趋势, 即核心区公园入侵植物丰富度低于郊区公园的入侵植物丰富度, 并且群落结构比较结果显示核心区的群落组成也与其他三区差异明显。Afons等[36]对开普敦城市和农村的入侵植物分布研究显示入侵植物在城市郊区的丰富度高于城市中心和农村, 与本文的结果相同, 他们解释这种分布的原因是城市郊区既是城市园艺入侵植物的容纳环境也是来自农业入侵植物或者农业遗留的容纳环境, 张楠[37]的研究结果也表明距离市中心越远的区域, 城市化梯度降低, 入侵草本植物多样性增高, 分析原因可能是因为城市中心人工生态系统中, 环境因素的影响非常小, 占据主要地位的因素是人类活动和管理方式和管理强度, 市中心公园重视管理, 所以入侵植物及时被清理, 在郊远区域的绿地公园由于缺乏管理, 所以入侵植物丰富度高。说明在城市的偏远区域环境因素是影响入侵植物分布的主要因素, 但是在高度城市化的区域, 人类活动是影响入侵植物分布的主要因素, 特别是城市绿地的人工生态系统中人类活动和管理强度直接影响入侵植物的扩散和分布。
| [1] |
Giakoumi S, Guilhaumon F, Kark S, Terlizzi A, Claudet J, Felline S, Cerrano C, Coll M, Danovaro R, Fraschetti S, Koutsoubas D, Ledoux J B, Mazor T, Mérigot B, Micheli F, Katsanevakis S. Space invaders; biological invasions in marine conservation planning. Diversity and Distributions, 2016, 22(12): 1220-1231. DOI:10.1111/ddi.12491 |
| [2] |
Katsanevakis S, Wallentinus I, Zenetos A, Leppäkoski E, Çinar M E, Oztürk B, Grabowski M, Golani D, Cardoso A C. Impacts of invasive alien marine species on ecosystem services and biodiversity: a pan-European review. Aquatic Invasions, 2014, 9(4): 391-423. DOI:10.3391/ai.2014.9.4.01 |
| [3] |
马金双. 中国入侵植物名录. 北京: 高等教育出版社, 2013.
|
| [4] |
赵彩云, 赵相健, 柳晓燕, 李俊生. 云南省六个国家级自然保护区外来入侵草本植物与本地植物的关系. 植物保护学报, 2019, 46(1): 122-129. |
| [5] |
Fernando V, Bastias C C, Oscar G, Elena G, Adrián E. Species coexistence in a changing world. Frontiers in Plant Science, 2015, 6: 866. |
| [6] |
Mack R N, Simberloff D, Lonsdale W M, Evans H, Clout M, Bazzaz F A. Biotic invasions: causes, epidemiology, global consequences, and control. Ecological Applications, 2000, 10(3): 689-710. DOI:10.1890/1051-0761(2000)010[0689:BICEGC]2.0.CO;2 |
| [7] |
Lonsdale W M. Global patterns of plant invasions and the concept of invasibility. Ecology, 1999, 80(5): 1522-1536. DOI:10.1890/0012-9658(1999)080[1522:GPOPIA]2.0.CO;2 |
| [8] |
Pyšek P, Jarošík V, Kučera T. Patterns of invasion in temperate nature reserves. Biological Conservation, 2002, 104(1): 13-24. DOI:10.1016/S0006-3207(01)00150-1 |
| [9] |
孙芳旭, 张嘉琦, 赵丹阳, 董丽. 北京市近郊区郊野公园外来植物特征. 西北林学院学报, 2018, 33(5): 278-284, 296-296. DOI:10.3969/j.issn.1001-7461.2018.05.44 |
| [10] |
万方浩, 刘万学, 郭建英, 强胜, 李保平, 王进军, 杨国庆, 牛红榜, 桂富荣, 黄文坤, 蒋智林, 王文琪. 外来植物紫茎泽兰的入侵机理与控制策略研究进展. 中国科学: 生命科学, 2011, 41(1): 13-21. |
| [11] |
严岳鸿, 何祖霞, 龚琴, 陈红锋, 邢福武. 广州的外来植物. 广西植物, 2007, 27(4): 570-575, 554-554. DOI:10.3969/j.issn.1000-3142.2007.04.008 |
| [12] |
张帅, 郭水良, 管铭, 印丽萍, 张若轩. 我国入侵植物多样性的区域分异及其影响因素——以74个地区数据为基础. 生态学报, 2010, 30(16): 4241-4256. |
| [13] |
塞依丁·海米提, 努尔巴依·阿布都沙力克, 阿尔曼·解思斯, 邵华, 维尼拉·伊利哈尔. 人类活动对外来入侵植物黄花刺茄在新疆潜在分布的影响. 生态学报, 2019, 39(2): 629-636. |
| [14] |
杨景成, 王光美, 姜闯道, 赵洪涛, 张志东. 城市化影响下北京市外来入侵植物特征及其分布. 生态环境学报, 2009, 18(5): 1857-1862. DOI:10.3969/j.issn.1674-5906.2009.05.048 |
| [15] |
彭程, 宿敏, 周伟磊, 王苏铭, 李景文, 王玲. 北京地区外来植物组成特征及入侵植物分布. 北京林业大学学报, 2010, 32(S1): 29-35. |
| [16] |
郭仲军, 黄继红, 路兴慧, 王计平, 臧润国. 基于第七次森林资源清查的新疆天然林生态系统服务功能. 生态科学, 2015, 34(4): 118-124. |
| [17] |
杜淼, 邵昱姝, 安帅. 北京市生活垃圾与经济增长——基于面板数据实证研究. 金融理论与教学, 2019(6): 88-93. DOI:10.3969/j.issn.1004-9487.2019.06.022 |
| [18] |
侯冰飞, 贾宝全, 冷平生, 王文和. 北京市城乡交错区绿地和植物种类的构成与分布. 生态学报, 2016, 36(19): 6256-6265. |
| [19] |
姜林, 胡骥, 杨振安, 詹伟, 赵川, 朱单, 何奕忻, 陈槐, 彭长辉. 植物功能群去除对高寒草甸群落结构、多样性及生产力的影响. 生态学报, 2021, 41(4): 1402-1411. |
| [20] |
黄建辉, 韩兴国, 杨亲二, 白永飞. 外来种入侵的生物学与生态学基础的若干问题. 生物多样性, 2003, 11(3): 240-247. DOI:10.3321/j.issn:1005-0094.2003.03.008 |
| [21] |
尼玛曲珍, 张玺, 谭坤, 鲁闯, 王国严, 徐阿生, 罗建. 西藏色季拉山川滇高山栎群落生态位特征. 生态科学, 2018, 37(3): 51-58. |
| [22] |
Wang C Y, Wei M, Wang S, Wu B D, Cheng H Y. Erigeron annuus (L.) Pers. and Solidago canadensis L. antagonistically affect community stability and community invasibility under the co-invasion condition. Science of the Total Environment, 2020, 716: 137128. DOI:10.1016/j.scitotenv.2020.137128 |
| [23] |
胡刚, 张忠华. 南宁的外来入侵植物. 热带亚热带植物学报, 2012, 20(5): 497-505. DOI:10.3969/j.issn.1005-3395.2012.05.011 |
| [24] |
曾文彬. 板块构造对我国东南沿海地区植物区系与东亚-北美间断分布的关系. 厦门大学学报: 自然科学版, 1989, 23(4): 410-413. DOI:10.3321/j.issn:0438-0479.1989.04.017 |
| [25] |
陈俊华, 刘兴良, 何飞, 刘世荣. 卧龙巴朗山川滇高山栎灌丛主要木本植物种群生态位特征. 林业科学, 2010, 46(3): 23-28. |
| [26] |
李一凡, 刘梦芸, 甘先华, 黄芳芳, 张卫强. 深圳市坝光湿地园银叶树群落优势种生态位特征. 生态环境学报, 2020, 29(11): 2171-2178. |
| [27] |
钱逸凡, 伊力塔, 胡军飞, 张超, 余树全, 沈露, 彭东琴. 普陀山主要植物种生态位特征. 生态学杂志, 2012, 31(3): 561-568. |
| [28] |
王苏铭, 张楠, 于琳倩, 赵容慧, 郝鹏, 李景文, 姜英淑, 沙海峰, 刘义, 张志翔. 北京地区外来入侵植物分布特征及其影响因素. 生态学报, 2012, 32(15): 4618-4629. |
| [29] |
黄乔乔, 沈奕德, 李晓霞, 程汉亭, 宋鑫, 范志伟. 外来入侵植物在中国的分布及入侵能力研究进展. 生态环境学报, 2012, 21(5): 977-985. |
| [30] |
海云瑞, 马菁, 姜广辉. 北京主要外来入侵植物分布特征及其影响因素分析. 宁夏农林科技, 2020, 61(8): 54-57. DOI:10.3969/j.issn.1002-204x.2020.08.017 |
| [31] |
冯建孟, 董晓东, 徐成东, 查凤书. 中国外来入侵植物的风险评价及空间分布格局. 西南大学学报: 自然科学版, 2011, 33(2): 57-63. |
| [32] |
任颖, 何萍, 徐杰, 贾娇. 滦河流域河岸带入侵植物分布特征及其与环境的关系. 应用生态学报, 2017, 28(6): 1843-1850. |
| [33] |
Wang H F, López-Pujol J, Meyerson L A, Qiu J X, Wang X K, Ouyang Z Y. Biological invasions in rapidly urbanizing areas: a case study of Beijing, China. Biodiversity and Conservation, 2011, 20(11): 2483-2509. DOI:10.1007/s10531-011-9999-x |
| [34] |
MacDougall A S, Gilbert B, Levine J M. Plant invasions and the niche. Journal of Ecology, 2009, 97(4): 609-615. DOI:10.1111/j.1365-2745.2009.01514.x |
| [35] |
Hou Y P, Peng S L, Chen B M, Ni G Y. Inhibition of an invasive plant (Mikania micrantha H.B.K.) by soils of three different forests in lower subtropical China. Biological Invasions, 2011, 13(2): 381-391. DOI:10.1007/s10530-010-9830-8 |
| [36] |
Afonso L, Esler K J, Gaertner M, Geerts S. Comparing invasive alien plant community composition between urban, peri-urban and rural areas; the city of Cape Town as a case study//Verma P, Singh P, Singh R, Raghubanshi A S, eds. Urban Ecology. Amsterdam: Elsevier, 2020: 221-236.
|
| [37] |
张楠. 北京城市生态廊道植物景观研究[D]. 北京: 北京林业大学, 2014.
|