 2022, Vol. 42
2022, Vol. 42文章信息
- 张亚芬, 郑子洪, 陈旭波, 骆争荣
- ZHANG Yafen, ZHENG Zihong, CHEN Xubo, LUO Zhengrong
- 入侵植物藿香蓟与常见伴生杂草的生态位特征
- Niche characteristics of the invasive plant Ageratum conyzoides and its commonly associated weeds
- 生态学报. 2022, 42(9): 3727-3737
- Acta Ecologica Sinica. 2022, 42(9): 3727-3737
- http://dx.doi.org/10.5846/stxb202104060881
-
文章历史
- 收稿日期: 2021-04-06
- 网络出版日期: 2022-01-06





2. 浙江九龙山国家级自然保护区管理中心, 遂昌 323300
2. Administration Center of Jiulongshan National Nature Reserve, Suichang 323300, China
生态位反映了物种在一定层次和范围内生存和发展所需要的条件及其在群落中的地位和作用[1]。在解释生物入侵的众多假说中, 很多都明确或隐含地认为外来种和土著种生态位的差异是外来种能否成功入侵的关键[2]。例如, 空余生态位假说认为成功入侵的原因在于外来种恰好占据了生态系统中的空余生态位[3];干扰假说和资源机遇假说认为通过不同的机制产生了“生态位机会”, 从而有利于外来种的入侵[4-5];物种多样性阻抗假说则默认了物种多样性高的群落中空余的生态位更少, 因此更难以被入侵[3];而达尔文归化假说则认为和本土物种亲缘关系远的外来种更容易归化, 因为它们和本土物种差异大, 生态位重叠小, 种间竞争弱[6]。另一方面, 生态位重叠是物种间利用性竞争排斥的必要条件[7]。外来种和本地种的生态位关系不仅与外来种能否成功入侵有关, 而且关系到外来种入侵后造成危害的严重程度[2]。因此, 在群落中研究外来种与本地种的生态位关系对于理解外来种的入侵机制以及它们造成的危害具有重要意义。
藿香蓟(Ageratum conyzoides L.)是菊科(Asteraceae)的一年生草本植物, 原产于热带美洲, 于19世纪传入我国华南, 现已广泛分布于我国长江流域及以南地区[8-9]。该种喜温暖、阳光充足的环境, 对土壤要求不严, 分枝力强。由于其具有较强的化感效应[10-12]、光竞争能力[13]、种子繁殖能力[14-15]和表型可塑性[16], 已对包括我国南方在内的旧大陆热带和亚热带生态系统和生物多样性造成了严重影响[9, 17-18]。在我国亚热带地区, 藿香蓟常见于农村荒地、疏林、乡村路边、垃圾堆或溪边等地, 常和其他外来入侵植物或本地杂草共生形成杂草群落[19]。
为了了解藿香蓟与其他杂草在群落中的生态关系, 探究造成藿香蓟成功入侵的可能机制及危害, 为制定相应防治对策和措施提供决策依据, 本研究在浙西南对杂草群落开展了广泛地调查, 分析了藿香蓟和其他常见杂草的生态位宽度和生态位重叠情况。生物成功入侵的一个重要特性是具有较高的生态位宽度[20]。许微楠等的实验研究表明, 藿香蓟对不同的光照、土壤碳氮比和水分条件表现出较强的表型可塑性, 其生态适应性很强[16]。因此, 本研究预测藿香蓟在群落中会有较宽的生态位。根据传统生态位理论的预期, 受藿香蓟入侵的杂草群落应该很少出现与藿香蓟具有较高生态位重叠的本地物种。一方面, 资源利用效率、对抗天敌取食和生长繁殖能力(即竞争能力[2])较强的本地种将阻止与它们有较高生态位重叠的外来种入侵;另一方面, 藿香蓟实现入侵后, 与藿香蓟有较高生态位重叠的竞争能力较弱的本地种将首先被藿香蓟挤出原生境。
1 材料与方法 1.1 研究区域概况本研究的野外群落调查在浙江省西南部的丽水市莲都区(28°06′— 28°44′N, 119°32′—120°08′E;面积约1502.1 km2)开展。莲都区处在括苍山、洞宫山、仙霞岭3山脉之间, 地形属浙南中山区, 以丘陵山地为主(面积占45.18%), 间有小块河谷平原(面积占26.3%)。基岩主要为沉积岩、花岗岩和白垩纪凝灰岩, 土壤类型主要是黄红壤、红壤和黄壤, 农耕区以各种类型的水稻土为主[21]。莲都区气候属中亚热带季风气候类型, 温暖湿润, 雨量充沛, 四季分明, 年降水量1395.5 mm, 年平均气温18.5 ℃, 具有明显的山地立体气候特征[22]。全区目前森林覆盖率约为79%, 动植物资源丰富。
1.2 群落调查2014—2016年(每年的5—12月), 在莲都区84个行政村的乡村路边、弃耕地、溪边、果园等处调查了109个1 m × 1 m的植物群落样方, 其中81个样方被藿香蓟入侵, 其余28个样方未被藿香蓟入侵。每村设置样方1—3个, 同村的样方间距大于200 m。在野外收割每个样方内的所有植物的地上部分, 带回实验室后将所有植物进行分类(按种)、称量其新鲜质量(鲜重)、计数并鉴定。
1.3 土壤取样和理化性质分析群落调查的同时, 本研究现场调查了每个样方的土壤干湿度(分为5级)、坡度、坡向、海拔和受光率, 并记录调查日期。受光率为样方正上方的光照强度和附近无任何遮阴处的光照强度的比值。在样方的4个顶点和中心处取10 cm深的土壤, 剔除石块和较大的植物残体后合并为该样方的土样。土壤样本带回实验室, 自然风干、研磨过筛后分析土壤的pH值、全氮含量、水解氮含量、有效磷含量、有效钾含量和有机质含量。生境调查和土壤理化性质分析更多细节详见郑珊珊等[23]。
1.4 数据处理和分析 1.4.1 生态位宽度和重叠度本研究分别用两种方法计测生态位宽度和生态位重叠, 以期更准确地了解丽水农村杂草群落的生态位特征。为了更简洁地呈现丽水农村杂草群落的生态位特征, 本研究共调查到286种植物(含种下分类单位),但为了更简洁地呈现丽水农村杂草群落的生态位特征,本研究仅分析藿香蓟及其它16种常见植物(均在25个以上样方出现)。17种常见杂草的基本信息见表 1。
| 编号 No. |
物种名 Species |
出现频次 Frequency |
平均相对多度* Average relative abundance |
生长型 Growth form |
| 1 | 藿香蓟Ageratum conyzoides | 81 | 0.236 | 一年生 |
| 2 | 升马唐Digitaria ciliaris | 64 | 0.064 | 一年生 |
| 3 | 铁苋菜Acalypha australis | 52 | 0.010 | 一年生 |
| 4 | 牛筋Eleusine indica | 48 | 0.038 | 一年生 |
| 5 | 碎米莎草Cyperus iria | 41 | 0.012 | 一年生 |
| 6 | 爵床Justicia procumbens | 39 | 0.019 | 一年生 |
| 7 | 酢浆草Oxalis corniculata | 39 | 0.001 | 多年生 |
| 8 | 叶下珠Phyllanthus urinaria | 38 | 0.004 | 一年生 |
| 9 | 柔枝莠竹Microstegium vimineum | 34 | 0.083 | 一年生 |
| 10 | 细风轮Clinopodium gracile | 30 | 0.014 | 多年生 |
| 11 | 鸭跖草Commelina communis | 30 | 0.046 | 一年生 |
| 12 | 短叶水蜈蚣Kyllinga brevifolia | 30 | 0.004 | 多年生 |
| 13 | 糠稷Panicum bisulcatum | 29 | 0.103 | 一年生 |
| 14 | 狗尾草Setaria viridis | 27 | 0.041 | 一年生 |
| 15 | 匍茎通泉草Mazus miquelii | 25 | 0.003 | 多年生 |
| 16 | 无辣蓼Polygonum pubescens | 25 | 0.147 | 一年生 |
| 17 | 龙葵Solanum nigrum | 25 | 0.042 | 一年生 |
| *相对多度=某物种在某样方中的鲜重/该样方中所有植物的总鲜重, 平均相对多度为该物种在出现该种的样方中相对多度的均值 | ||||
首先将1m × 1 m的样方作为一个综合资源位, 以各物种在各资源位(样方)的地上鲜重作为多度指标来计算物种的生态位宽度和种间的生态位重叠度。生态位宽度反映物种或种群对环境适应的状况或对资源利用的程度[24]。生态位宽度计测采用Levins公式中的Shannon-Weiner指数[25]:
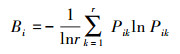
|
式中, Bi物种i的生态位宽度, Pik=nik/Ni, 即种i对第k个资源的利用占其对全部资源利用的比例, njk是第i个物种利用资源状态k的鲜重, Ni是该种所有资源状态中的总鲜重, r为资源状态总数, 即样方数。该指数的值域在0—1之间, 数值越大表示生态位越宽。
生态位重叠是2个或多个物种或种群在适应环境和利用资源的实际幅度或潜在能力方面所表现出的共同性或相似性[24]。在传统的生态位计测方法中, 选用对称α法, 即Pianka指数计算常见种间之间的生态位重叠[24-25]。

|
式中, Oij为物种i和物种j的重叠指数; Pik和Pjk分别为种i和种j在资源k上的利用比例, 计算时用种i和种j的鲜重占样方k的植物总鲜重的比例来表示。该指数的值域也在0—1之间, 数值越大表示生态位重叠度越高, 便于对不同种群的生态位重叠进行客观比较[25]。
上述生态位宽度和生态位重叠计测公式虽然都有明确的几何学解释, 生物意义明确, 并且充分利用了物种在不同综合资源位上的多度信息, 但是都没有考虑到资源的可利用性[24]。为考虑资源含量的实际情况, 充分利用本研究调查的生境数据(含调查日期, 即生长季节), 使用基于概率的方法来计测杂草群落常见种的n-维超体积生态位重叠。生态位重叠度(OBA)被定义为在物种B的n-维超体积生态位范围内物种A的个体出现的概率[26]。
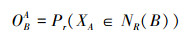
|
物种的生态位范围(NR)被定义为一个特定的n维空间, 在此空间内找到满足物种生存所需条件的概率为α(如95%)[26], 即
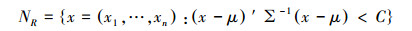
|
上述两式中n为生态位维数(或生态因子数), X为出现某物种的生境中各生境因子(n维)的观测值, 服从均值为μ, 方差为∑的多元正态分布, C为Pr(X∈NR)=95%时的常数。在具体应用中, 由于均值μ和方差∑都是未知的, 因此需要基于调查数据进行参数估计。Swanson等建议利用贝叶斯法来进行参数估计并计算估计值的不确定性[26]。考虑到季节变化具有周期性, 本研究以2月3日(立春)作为一年中的首日, 各调查日期先转化为相应日序(值域为1—365), 再将日序乘以2π/365后进行余弦转换。坡向也是环形变量, 原始数据同样进行了余弦转换。
Shannon-Weiner生态位宽度指数和Pianka生态位重叠度指数的计算使用R语言的spaa软件包。基于概率方法的n维超体积生态位宽度和生态位重叠计算采用R语言的nicheROVER软件包。
1.4.2 典范对应分析近年来, 多元统计在生态位分析中不断得到应用[24-25]。该方法可以把多个与种分离高度相关的生态参数进行合并或剔除, 使维数降低且互相独立, 能将物种间的生态关系更直观的呈现出来。如郭水良和曹同应用典范对应分析(CCA)探讨了长白山金发藓科植物的生态位分化, 取得了很好的效果[27]。除了直接计测常见种的生态位宽度和重叠, 本研究也利用典范对应分析研究杂草群落中主要物种与藿香蓟的生态关系。本研究用R语言软件包vegan进行典范对应分析。
1.4.3 种间相关分析为了回答与藿香蓟具有较高生态位重叠的本地物种是否较少出现在受藿香蓟入侵且其优势度较高的杂草群落的问题, 本研究分析了藿香蓟及16个常见本地种的种间关系, 并进一步分析种间关系的亲密程度和种间生态位重叠程度的关系。种间相关采用Pearson相关系数来衡量, 并用t-检验来检测种间相关的显著程度。
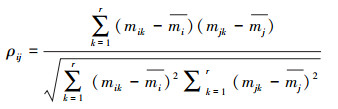
|
式中, ρij代表种i和j之间的相关系数, r为样方数, mik和mjk分别是种i和j在样方k中的鲜重值, 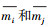
对丽水农村杂草群落中17种常见杂草的生态位宽度分析表明, 藿香蓟的生态位宽度最宽(0.804), 其他杂草的生态位宽度适中, Shannon-Weiner生态位宽度指数均介于0.40—0.65之间(表 2)。除藿香蓟以外, 生态位宽度相对较宽的还有升马唐(Digitaria ciliaris, 0.64)、碎米莎草(Cyperus iria, 0.60)和铁苋菜(Acalypha australis, 0.59)。生态位宽度最小的是细风轮(Clinopodium gracile), Shannon-Weiner生态位宽度指数为0.41。
| 物种编号 Species no. |
生态位宽度 Niche width |
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 |
| 1 | 0.80 | — | 0.05 | -0.05 | -0.04 | 0.01 | -0.11 | -0.08 | 0.00 | -0.11 | -0.06 | -0.10 | -0.06 | -0.13 | -0.04 | 0.10 | -0.11 | 0.20 |
| 2 | 0.64 | [0.24] | — | 0.16 | [0.33] | 0.09 | -0.08 | 0.03 | -0.08 | -0.09 | -0.02 | -0.05 | -0.04 | -0.09 | 0.10 | 0.03 | 0.06 | -0.06 |
| 3 | 0.59 | [0.16] | [0.26] | — | -0.05 | -0.08 | 0.17 | -0.09 | 0.07 | -0.06 | -0.05 | -0.03 | -0.10 | 0.06 | 0.04 | 0.12 | [0.31] | 0.16 |
| 4 | 0.58 | [0.16] | [0.41] | [0.07] | — | 0.20 | -0.11 | 0.15 | -0.07 | -0.09 | 0.03 | 0.11 | 0.00 | -0.09 | [0.49] | -0.01 | -0.07 | -0.04 |
| 5 | 0.60 | [0.21] | [0.20] | [0.05] | [0.29] | — | -0.08 | [0.46] | 0.03 | -0.10 | 0.19 | [0.23] | 0.11 | -0.10 | -0.02 | 0.19 | -0.08 | -0.08 |
| 6 | 0.58 | [0.11] | [0.05] | [0.26] | [0.01] | [0.04] | — | -0.03 | 0.02 | -0.05 | 0.02 | -0.04 | 0.04 | 0.15 | 0.04 | -0.11 | -0.05 | -0.05 |
| 7 | 0.58 | [0.12] | [0.14] | [0.03] | [0.23] | [0.52] | [0.07] | — | -0.07 | 0.01 | [0.23] | -0.03 | 0.11 | -0.09 | -0.01 | 0.05 | -0.06 | -0.02 |
| 8 | 0.52 | [0.16] | [0.02] | [0.15] | [0.02] | [0.12] | [0.10] | [0.02] | — | -0.08 | -0.04 | -0.04 | -0.05 | 0.04 | -0.06 | -0.05 | -0.04 | -0.07 |
| 9 | 0.52 | [0.08] | [0.02] | [0.04] | 0.00 | 0.00 | [0.05] | [0.10] | 0.00 | — | -0.05 | -0.04 | 0.03 | 0.01 | -0.07 | -0.10 | -0.05 | 0.15 |
| 10 | 0.41 | [0.08] | [0.06] | [0.03] | [0.10] | [0.25] | [0.09] | [0.28] | 0.02 | 0.01 | — | 0.01 | 0.18 | -0.06 | 0.05 | 0.05 | -0.05 | -0.05 |
| 11 | 0.43 | [0.04] | [0.03] | [0.04] | [0.17] | [0.28] | [0.03] | [0.04] | 0.01 | [0.02] | 0.05 | — | -0.06 | -0.05 | -0.04 | 0.06 | 0.03 | -0.04 |
| 12 | 0.55 | [0.14] | [0.08] | [0.02] | [0.11] | [0.21] | [0.14] | [0.20] | [0.04] | [0.12] | [0.24] | 0.01 | — | -0.09 | -0.09 | [0.29] | -0.05 | -0.08 |
| 13 | 0.52 | [0.07] | [0.02] | [0.15] | 0.01 | 0.00 | [0.23] | 0.01 | [0.11] | 0.09 | 0.00 | [0.01] | 0.01 | — | -0.07 | -0.10 | -0.06 | -0.04 |
| 14 | 0.51 | [0.12] | [0.19] | [0.13] | [0.54] | [0.08] | [0.12] | [0.07] | 0.01 | 0.01 | [0.11] | 0.02 | 0.00 | 0.01 | — | -0.03 | -0.01 | -0.05 |
| 15 | 0.55 | [0.27] | [0.15] | [0.22] | [0.10] | [0.28] | 0.01 | [0.15] | [0.04] | 0.00 | [0.11] | [0.12] | [0.36] | [0.01] | 0.06 | — | -0.08 | -0.05 |
| 16 | 0.44 | [0.05] | [0.14] | [0.37] | [0.01] | 0.01 | [0.03] | [0.02] | 0.03 | [0.02] | 0.01 | [0.07] | [0.03] | 0.01 | [0.06] | 0.00 | — | -0.02 |
| 17 | 0.42 | [0.30] | [0.03] | [0.23] | [0.04] | 0.01 | [0.03] | [0.06] | 0.00 | [0.20] | 0.00 | 0.01 | 0.00 | [0.03] | 0.02 | 0.03 | 0.04 | — |
| []内数字表示其与0存在显著差异 | ||||||||||||||||||
以Pianka指数计测的生态位重叠度分析表明, 常见杂草间的生态位重叠度在0.5以上的种对仅2对, 分别为狗尾草(Setaria viridis)和牛筋(Eleusine indica, 0.54)、酢浆草(Oxalis corniculata)和碎米莎草(0.52)。生态位重叠度在0.3—0.5之间的种对有升马唐和牛筋(0.41)、匍茎通泉草(Mazus miquelii)和短叶水蜈蚣(Kyllinga brevifolia, 0.36)。其余种对的生态位重叠全在0.3以下(表 2)。外来杂草藿香蓟与其他常见本地杂草的生态位重叠度介于0.04—0.296之间, 显著高于本地杂草之间的生态位重叠(P=0.04)。直线相关分析表明, 某种杂草与其他杂草的平均Pianka生态位重叠值与其Shannon-Weiner生态位宽度指数呈显著正相关(r=0.72, P<0.001)。
n-维超体积生态位重叠分析表明不同种对之间生态位重叠程度差异较大, OBA值介于0.00—0.94之间。短叶水蜈蚣对于藿香蓟的n-维超体积生态位重叠值最高, 达到了0.94;另外无辣蓼(Polygonum pubescens)、叶下珠(Phyllanthus urinaria)、铁苋菜和升马唐对于藿香蓟的n-维超体积生态位重叠值都很高, 分别为0.86、0.83、0.82和0.80。这说明这些物种的个体生长所需的环境大部分与藿香蓟的生态位空间重叠。同时, 各常见杂草对于细风轮、无辣蓼和龙葵(Solanum nigrum)的n-维超体积生态位重叠值几乎为0(表 3)。进一步分析表明本地杂草重叠于藿香蓟的程度(表 3第1列)显著高于藿香蓟重叠于本地杂草(表 3第1行, 成对t检验P<0.001)以及本地杂草间的相互重叠(成组t检验P<0.001)。直线相关分析表明, 其他物种(作为物种A)对于某物种(作为物种B)的平均重叠度与该物种(物种B)自身的Shannon-Weiner生态位宽度呈显著正相关(r=0.84, P<0.001);而某物种(作为物种A)对于其他物种(作为物种B)的平均重叠度与该物种(物种A)自身的Shannon-Weiner生态位宽度无关(r =-0.27, P=0.28)。直线回归分析表明每个种对的n-维超体积生态位重叠的均值与Pianka生态位重叠指数呈显著正相关(β=0.58, P<0.001), 但解释度R2仅0.23。这说明上述两种生态位重叠的计测方法反映的物种间生态位重叠具有一致的变化趋势, 但也存在较大的差异。
| 物种 Species |
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 |
| 1 | — | [0.18] | [0.03] | 0.04 | [0.08] | [0.13] | [0.24] | 0.01 | [0.11] | 0.00 | 0.01 | 0.00 | 0.00 | 0.02 | 0.01 | 0.00 | 0.00 |
| 2 | [0.80] | — | [0.04] | [0.09] | [0.10] | [0.12] | [0.29] | 0.00 | [0.14] | 0.00 | 0.04 | 0.01 | 0.00 | 0.03 | 0.01 | 0.00 | 0.00 |
| 3 | [0.82] | [0.43] | — | [0.10] | [0.14] | [0.61] | [0.45] | [0.13] | [0.30] | 0.00 | [0.07] | 0.04 | 0.04 | [0.13] | 0.05 | 0.00 | 0.00 |
| 4 | [0.52] | [0.33] | 0.03 | — | [0.43] | [0.07] | [0.45] | 0.01 | [0.06] | 0.00 | [0.11] | 0.01 | 0.00 | 0.04 | 0.04 | 0.00 | 0.00 |
| 5 | [0.48] | [0.17] | 0.02 | [0.29] | — | [0.07] | [0.51] | 0.02 | [0.08] | 0.00 | [0.07] | 0.01 | 0.00 | 0.01 | 0.05 | 0.00 | 0.00 |
| 6 | [0.77] | [0.18] | [0.13] | [0.05] | [0.08] | — | [0.35] | [0.04] | [0.14] | 0.00 | 0.03 | 0.01 | 0.01 | [0.08] | 0.03 | 0.00 | 0.00 |
| 7 | [0.52] | [0.16] | 0.02 | [0.14] | [0.30] | [0.12] | — | 0.01 | [0.12] | 0.00 | [0.07] | 0.00 | 0.00 | 0.03 | 0.03 | 0.00 | 0.00 |
| 8 | [0.83] | [0.20] | [0.17] | [0.06] | [0.15] | [0.28] | [0.38] | — | [0.14] | 0.00 | 0.02 | 0.03 | 0.02 | [0.08] | 0.06 | 0.00 | 0.00 |
| 9 | [0.59] | [0.13] | 0.03 | 0.02 | 0.05 | [0.08] | [0.21] | 0.00 | — | 0.00 | 0.04 | 0.00 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 |
| 10 | [0.65] | [0.12] | 0.03 | [0.13] | [0.38] | [0.21] | [0.46] | 0.01 | 0.05 | — | 0.03 | 0.01 | 0.00 | [0.09] | 0.02 | 0.00 | 0.00 |
| 11 | [0.19] | [0.12] | 0.01 | [0.18] | [0.29] | 0.04 | [0.30] | 0.01 | [0.10] | 0.00 | — | 0.00 | 0.00 | 0.01 | 0.04 | 0.00 | 0.00 |
| 12 | [0.94] | [0.58] | [0.24] | [0.29] | [0.45] | [0.41] | [0.64] | [0.12] | [0.24] | 0.01 | 0.03 | — | 0.02 | [0.11] | [0.12] | 0.00 | 0.00 |
| 13 | [0.61] | [0.14] | [0.13] | 0.04 | [0.06] | [0.23] | [0.19] | 0.03 | [0.19] | 0.00 | 0.01 | 0.01 | — | 0.02 | 0.02 | 0.00 | 0.00 |
| 14 | [0.71] | [0.19] | [0.06] | [0.16] | [0.14] | [0.23] | [0.41] | 0.02 | [0.11] | 0.00 | 0.03 | 0.01 | 0.00 | — | 0.01 | 0.00 | 0.00 |
| 15 | [0.73] | [0.32] | [0.05] | [0.19] | [0.26] | [0.16] | [0.49] | 0.04 | [0.07] | 0.00 | 0.05 | 0.02 | 0.01 | 0.01 | — | 0.00 | 0.00 |
| 16 | [0.86] | [0.60] | [0.37] | 0.03 | [0.06] | [0.39] | [0.41] | 0.02 | 0.50 | 0.00 | [0.20] | 0.02 | 0.00 | [0.12] | 0.01 | — | 0.00 |
| 17 | [0.79] | [0.10] | [0.07] | [0.07] | 0.08 | [0.18] | [0.28] | 0.00 | [0.27] | 0.00 | 0.03 | 0.00 | 0.00 | 0.05 | 0.01 | 0.00 | — |
| 生态位重叠显示为行物种的个体在列物种的超体积生态位范围内出现的概率。[]内数字表示其与0存在显著差异 | |||||||||||||||||
对83种出现频度较高(至少在5个样方中出现)的植物进行典范对应分析, 从前4个排序轴来看, 外来杂草和本地杂草充分混杂, 并没有出现外来杂草或本地杂草各自聚成种团的现象(图 1)。这说明外来杂草和本地杂草具有一定的生态相似度。相对于本地杂草, 外来杂草并无明显的生态特化。特别是本研究重点关注的藿香蓟位于两个排序图的中间位置。这说明藿香蓟具有较高的中生性。在影响杂草群落物种组成的各生境因子中土壤有机质和湿度对第一排序轴有重要影响;土壤pH值、样地坡度和调查日期对第二排序轴有重要影响;光照、土壤有机质、湿度和速效氮对第三排序轴影响较大;坡度对第四排序轴影响较大(图 1)。

|
| 图 1 丽水农村杂草群落环境因子和常见植物的典范对应分析(CCA) 排序图 Fig. 1 Canonical correspondence analysis (CCA) ordination showing the distribution of environmental factors and 83 common weed species in the weed community in the rural areas of southwest Zhejiang Province 空心圆点表示73种至少在5个样方中出现本地植物, 实心圆点表示9种至少在5个样方中出现外来植物(除藿香蓟外), *点表示藿香蓟 |
种间相关分析结果表明, 136个种对中仅有7个种对呈显著正相关, 包括狗尾草与牛筋(0.49)、酢浆草与碎米莎草(0.46)、升马唐与牛筋(0.33)、无辣蓼与铁苋菜(0.31)、匍茎通泉草与短叶水蜈蚣(0.29)、碎米莎草和鸭跖草(0.23)以及细风轮和酢浆草(0.23)。藿香蓟与本地杂草间未见显著的种间相关性。所有17种常见杂草间无显著种间负相关(表 2)。
3 讨论与结论生态位宽度主要反映种群对资源利用的程度或对环境的适应状况[24]。国内外很多已有的研究都表明, 成功入侵的外来生物的一个重要特性是具有较高的生态位宽度[20, 27-30]。本研究的结果表明, 在调查的几类生境类型中, 藿香蓟具有最大的现实生态位宽度。在已往对丽水市莲都区开展的入侵植物种类和分布调查也发现, 在调查的100多个行政村中, 藿香蓟的出现频率达到90.32%, 且多度级大多为极多(最高级别)。调查发现藿香蓟是丽水市莲都区所有入侵植物中占据生境类型多样性最高的入侵种之一, 分布于24种生境。虽然总体上藿香蓟喜全光照和较湿润的土壤环境, 但控制性实验研究也表明, 藿香蓟具有很强的耐荫和耐旱能力[15-16]。在透光率21.2%的遮荫网遮盖下, 藿香蓟的开花数量甚至有所提高。这说明藿香蓟在丽水地区具有很强的适应性, 对环境资源的利用程度较高, 特化程度较小。藿香蓟的这种宽生态幅特性为其成功入侵我国南方地区奠定了坚实的基础。
种间生态位重叠反映了物种或种群在适应环境和利用资源的实际幅度或潜在能力方面所表现出的共同性或相似性[24]。本研究的结果表明外来杂草藿香蓟与其他常见杂草的Pianka生态位重叠指数介于0.04—0.296之间。有学者认为Pianka生态位重叠值相对于其他生态位重叠指数偏小[31]。由于其具有较为直观的几何解释, 而且生态位重叠不超过1, 便于对不同种群的生态位重叠进行客观比较, 本研究综合分析了国内已报道的入侵植物与本地植物间的Pianka生态位重叠值的频度分布 [20, 29-30, 32-36], 发现其中位数为0.315(图 2)。对比来看, 藿香蓟与丽水本地常见杂草的生态位重叠值处于中低水平。这说明即使在资源供应不足时, 藿香蓟与丽水本地常见杂草的利用性竞争也不会很激烈。况且藿香蓟主要出现在受干扰的生境中, 光照和主要土壤养分因子的供应量处于中高水平[19], 群落内部各物种间(包括外来种和本地种)的竞争可能很弱。来自本地植物微弱的利用性竞争阻力给藿香蓟成功入侵受干扰生境中的本地植物群落创造良好的“生态位机会”。

|
| 图 2 国内文献已报道的外来-本地植物间Pianka生态位重叠指数的分布状况 Fig. 2 The distribution of Pianka′s niche overlap index between exotic and native plants reported in domestic literatures |
由于Pianka指数具有对称性质(即物种A重叠于物种B等于物种B重叠于物种A), 无法分析种对内相互重叠的差异[24]。为了进一步解析种对内部两物种相互重叠的差异, 同时充分考虑资源的实际状态, 本研究进一步利用n-维超体积生态位重叠分析了丽水农村常见杂草间的生态位重叠关系。结果发现藿香蓟与本地杂草间的生态位重叠存在明显不对称性——本地杂草重叠于藿香蓟的程度显著高于藿香蓟重叠于本地杂草。藿香蓟对于本地杂草的n-维超体积生态位重叠仅0.00—0.18。这进一步说明了很多本地常见杂草对于藿香蓟的入侵阻抗很弱, 部分本地物种对藿香蓟几乎没有阻抗。这让藿香蓟几乎可以畅行无阻地入侵本地群落。同时, 本研究也发现本地杂草对于藿香蓟的n-维生态位重叠值都很高, 短叶水蜈蚣对于藿香蓟的n-维生态位重叠值甚至高达0.94。在受藿香蓟入侵群落中, 藿香蓟可以凭借其强大的种子繁殖能力和资源利用能力[13-15], 使种群快速增殖。本研究的结果说明当资源被不断扩增的藿香蓟消耗到较低水平时, 这些常见本地植物将受到来自藿香蓟的激烈竞争排挤。这可能是人们在野外常看到藿香蓟单优群落的原因所在。藿香蓟入侵后会对本地其他物种的生长造成严重影响。
不管是从Pianka指数的角度还是n-维超体积生态位重叠概率的角度, 本研究的结果都显示外来杂草藿香蓟与其他常见杂草的生态位重叠度都显著高于本地杂草之间的生态位重叠。有学者认为, 这是因为外来植物入侵导致了本土植物间的生态位重叠明显下降, 同时大大增强本地植物与外来植物之间的竞争[33]。但曲波等人的研究发现三裂叶豚草(Ambrosia trifida)入侵会显著降低本地早春植物的生态位宽度, 提高本地植物间生态位重叠系数, 干扰植物对资源的利用, 增强本地植物间的竞争[37]。这种现象很大程度上应该是藿香蓟本身具有较大的生态位宽度造成的。本研究发现种间生态位重叠值与物种(主要是物种B)的生态位宽度显著正相关。在前人的很多研究中也发现类似的关联[20, 30, 32]。这是由生态位重叠的数学和生态学意义决定的。
典范对应分析结果表明藿香蓟位于两个CCA排序图的较中间位置, 体现了藿香蓟对中生性环境的偏好。在自然界, 中生性环境是分布最广、出现频率最高的生境类型。对中生环境的高度适应促进了藿香蓟在浙西南地区的广泛传播。另一方面, 这一结果也说明藿香蓟的入侵并不依赖于对于环境中特殊的未利用资源的开发, 其生态位并未发生明显特化。除此之外, 本研究结果显示调查到的所有外来杂草和本地杂草在排序图上充分混合, 所有外来种都未见明显的生态位特化。这充分说明经过较长时期的适应或筛选, 成功入侵的外来植物在重要生态因子的需求上已经与本地种基本相似。这与郑珊珊等从谱系距离角度的研究结论基本一致。他们发现本研究调查的群落中, 入侵种-本地种谱系距离接近本地植物间现实的谱系距离[23], 入侵植物没有明显的特别之处。
总的来说, 丽水农村杂草群落的植物种间生态位重叠并不高, 资源利用性竞争不足以产生种间排斥。因此, 本研究的结果显示所有17种常见杂草间无显著种间负相关。由于杂草群落普遍受到不同程度的干扰, 群落各物种尚未形成稳定的协调搭配和紧密的耦合关系(本研究仅有7个种对呈现了显著的正相关);群落结构和功能不稳定, 物种组成以一年生草本植物为主, 多年生植物稀少。这种状况十分有利于包括藿香蓟在内的具有较高繁殖力和适合度的外来植物入侵丽水农村。
本研究用植物鲜重替代传统的盖度、多度或者重要值来表征植物的资源获取情况, 以此计算物种的生态位宽度、生态位重叠和种间相关, 并进行排序分析。理论上来说, 植物的干物质质量(干重)是植物生产力或资源获取情况(竞争力)的最合适表征。考虑到工作量的问题, 本研究在调查时没有收集植物的干重数据。由于处于不同生育阶段的不同物种组织含水量差异较大, 鲜重差异仅近似表征植物生产力/竞争力种间差异。本研究虽然在方法上相对于前人做了改进, 但这种改进仍不够彻底, 即没有用干重来表征植物资源获取情况。因此, 本研究的结果可能仍存在一定的误差。在今后的研究中, 需要用干重作为基础数据来研究植物的生态位特征, 并验证本研究的结论。
| [1] |
张光明, 谢寿昌. 生态位概念演变与展望. 生态学杂志, 1997, 16(6): 46-51. DOI:10.3321/j.issn:1000-4890.1997.06.010 |
| [2] |
于文波, 黎绍鹏. 基于现代物种共存理论的入侵生态学概念框架. 生物多样性, 2020, 28(11): 1362-1375. DOI:10.17520/biods.2020243 |
| [3] |
Elton C S. The Ecology of Invasions by Animals and Plants. London: Methuen, 1958, 54-54. |
| [4] |
Mack R N, Simberloff D, Lonsdale W M, Evans H, Clout M, Bazzaz F A. Biotic invasions: causes, epidemiology, global consequences, and control. Ecological Applications, 2000, 10(3): 689-710. DOI:10.1890/1051-0761(2000)010[0689:BICEGC]2.0.CO;2 |
| [5] |
Davis M A, Grime J P, Thompson K. Fluctuating resources in plant communities: a general theory of invasibility. Journal of Ecology, 2000, 88(3): 528-534. DOI:10.1046/j.1365-2745.2000.00473.x |
| [6] |
Daehler C C. Darwin's naturalization hypothesis revisited. The American Naturalist, 2001, 158(3): 324-330. DOI:10.1086/321316 |
| [7] |
May R, McLean A. 理论生态学——原理及应用(第三版). 陶毅, 王百桦, 译. 北京: 高等教育出版社, 2010: 103-106.
|
| [8] |
万方浩, 刘全儒, 谢明. 生物入侵: 中国外来入侵植物图鉴. 北京: 科学出版社, 2012: 28-28.
|
| [9] |
马金双, 李惠茹. 中国外来入侵植物名录. 北京: 高等教育出版社, 2018: 113-115.
|
| [10] |
孔垂华, 黄寿山, 胡飞. 胜红蓟化感作用研究Ⅴ. 挥发油对真菌、昆虫和植物的生物活性及其化学成份. 生态学报, 2001, 21(4): 584-587. DOI:10.3321/j.issn:1000-0933.2001.04.011 |
| [11] |
Kong C H, Hu F, Xu X H. Allelopathic potential and chemical constituents of volatiles from Ageratum conyzoides under stress. Journal of Chemical Ecology, 2002, 28(6): 1173-1182. DOI:10.1023/A:1016229616845 |
| [12] |
Kong C H. Ecological pest management and control by using allelopathic weeds (Ageratum conyzoides, Ambrosia trifida, and Lantana camara) and their allelochemicals in China. Weed Biology and Management, 2010, 10(2): 73-80. DOI:10.1111/j.1445-6664.2010.00373.x |
| [13] |
Banyikwa F F, Rulangaranga Z K. Growth analysis of groundnuts (Arachis hypogea) in competition with Ageratum conyzoides. Turrialba, 1985, 35(3): 215-219. |
| [14] |
郝建华, 吴海荣, 强胜. 部分菊科入侵种种子(瘦果)的萌发能力和幼苗建群特性. 生态环境学报, 2009, 18(5): 1851-1856. DOI:10.3969/j.issn.1674-5906.2009.05.047 |
| [15] |
陈翠琴. 逆境胁迫对胜红蓟(Ageratum conyzoides Linn. )生长和种子萌发影响的研究. 金华: 浙江师范大学, 2011.
|
| [16] |
许微楠, 郑珊珊, 余其娇, 骆争荣. 环境因子对藿香蓟形态、存活和繁殖的影响. 丽水学院学报, 2019, 41(5): 34-40. DOI:10.3969/j.issn.2095-3801.2019.05.007 |
| [17] |
Dogra K S, Kohli R K, Sood S K, Dobhal P K. Impact of Ageratum conyzoides L. on the diversity and composition of vegetation in the Shivalik hills of Himachal Pradesh (Northwester Himalaya), India. International Journal of Biodiversity and Conservation, 2009, 1(4): 135-145. |
| [18] |
Kashina B D, Mabagala R S, Mpunni A A. First report of Ageratum conyzoides L. and Sida acuta Burm F. as new weed hosts of Tomato yellow leaf curl Tanzania virus. Plant Protection Science, 2003, 39(1): 18-22. |
| [19] |
Luo Z R, Chen X R, Xia G S, Chen X B. Extrinsic environmental factors, not resident diversity itself, lead to invasion of Ageratum conyzoides L. in diverse communities. Ecological Research, 2018, 33(6): 1245-1253. DOI:10.1007/s11284-018-1637-6 |
| [20] |
李俊娇, 谷艳芳, 张丽霞, 丁圣彦. 开封市郊区主要生境外来杂草生态位. 生态学杂志, 2008, 27(9): 1543-1548. |
| [21] |
浙江省丽水地区农业局. 丽水土壤. 上海: 上海科学技术出版社, 1989: 18-22.
|
| [22] |
强玉华, 李敏. 近60年丽水市气候变化特征分析. 现代农业科技, 2010(12): 238-240. DOI:10.3969/j.issn.1007-5739.2010.12.156 |
| [23] |
郑珊珊, 陈旭波, 许微楠, 骆争荣, 夏更寿. 外来种-本地种亲缘关系对外来植物归化和入侵的影响. 植物生态学报, 2018, 42(10): 990-999. DOI:10.17521/cjpe.2018.0101 |
| [24] |
李德志, 石强, 臧润国, 王绪平, 盛丽娟, 朱志玲, 王长爱. 物种或种群生态位宽度与生态位重叠的计测模型. 林业科学, 2006, 42(7): 95-103. |
| [25] |
李, 朱金兆, 朱清科. 生态位理论及其测度研究进展. 北京林业大学学报, 2003, 25(1): 100-107. DOI:10.3321/j.issn:1000-1522.2003.01.022 |
| [26] |
Swanson H K, Lysy M, Power M, Stasko A D, Johnson J D, Reist J D. A new probabilistic method for quantifying n-dimensional ecological niches and niche overlap. Ecology, 2015, 96(2): 318-324. DOI:10.1890/14-0235.1 |
| [27] |
郭水良, 曹同. 应用典范对应分析探讨长白山金发藓科植物的生态位分化. 木本植物研究, 2000, 20(3): 286-293. |
| [28] |
Hertling U M, Lubke R A. Assessing the potential for biological invasion-the case of Ammophila arenaria in South Africa. South African Journal of Science, 2000, 96(9/10): 520-527. |
| [29] |
吴海荣, 强胜, 林金成. 南京市春季外来杂草调查及生态位研究. 西北植物学报, 2004, 24(11): 2061-2068. DOI:10.3321/j.issn:1000-4025.2004.11.015 |
| [30] |
陈秀芝, 朱莉莉, 郭水良. 长江口九段沙典型潮沟植物群落的物种多样性和种群生态位. 长江流域资源与环境, 2010, 19(S2): 20-27. |
| [31] |
郭水良, 李扬汉, 赵铁桥. 浙江金华地区小麦-杂草群落中杂草生态位的研究. 植物生态学报, 1998, 22(1): 76-84. DOI:10.3321/j.issn:1005-264X.1998.01.009 |
| [32] |
陈伟, 兰国玉, 安锋, 蒋菊生, 王真辉, 陈秋波, 吴新民. 海南外来杂草——假臭草群落生态位特征研究. 西北林学院学报, 2007, 22(2): 24-27. DOI:10.3969/j.issn.1001-7461.2007.02.006 |
| [33] |
江焕, 张辉, 龙文兴, 方燕山, 符明期, 朱孔新. 金钟藤入侵群落的种间联结及生态位特征. 生物多样性, 2019, 27(4): 388-399. |
| [34] |
曹培健, 于明坚, 金孝锋, 丁炳扬. 水盾草入侵沉水植物群落主要种群生态位和种间联结研究. 浙江大学学报: 农业与生命科学版, 2006, 32(3): 334-340. DOI:10.3321/j.issn:1008-9209.2006.03.018 |
| [35] |
洪思思, 缪崇崇, 方本基, 胡仁勇, 丁炳扬. 浙江省阔叶丰花草入侵群落物种多样性、生态位及种间联结研究. 武汉植物学研究, 2008, 26(5): 501-508. DOI:10.3969/j.issn.2095-0837.2008.05.013 |
| [36] |
岳茂峰, 冯莉, 杨彩宏, 田兴山, 辛国荣. 珠三角地区四季草坪杂草群落组成及其生态位. 生态学杂志, 2009, 28(12): 2483-2488. |
| [37] |
曲波, 薛晨阳, 许玉凤, 高英美, 陈鲜, 王文筠. 三裂叶豚草入侵对撂荒农田早春植物群落的影响. 沈阳农业大学学报, 2019, 50(3): 358-364. |