 2022, Vol. 42
2022, Vol. 42文章信息
- 史娜娜, 郭宁宁, 刘高慧, 王琦, 韩煜, 肖能文
- SHI Nana, GUO Ningning, LIU Gaohui, WANG Qi, HAN Yu, XIAO Nengwen
- 北京市两栖爬行动物空间分布格局及影响因素
- Spatial distribution pattern and influencing factors of amphibians and reptiles in Beijing
- 生态学报. 2022, 42(9): 3806-3821
- Acta Ecologica Sinica. 2022, 42(9): 3806-3821
- http://dx.doi.org/10.5846/stxb202102260534
-
文章历史
- 收稿日期: 2021-02-26
- 网络出版日期: 2022-01-06





2. 天津市滨海新区环境创新研究院, 天津 300450
2. Research Institute for Environmental Innovation (Tianjin Binhai), Tianjin 300450, China
生物多样性是人类赖以生存和发展的物质基础。我国生物多样性保护工作在生物多样性维持机制、生物多样性与生态系统功能关系、生物多样性致危因素、生物安全等方面取得了长足进步[1-2], 对促进生物多样性和生态学研究具有重要意义。两栖爬行动物是研究脊椎动物由水生向陆生演化的典型对象, 是食物链和生态系统中的重要中间类群[3];与其它脊椎动物相比, 具有体型较小、种类较少、分布范围对环境敏感等特征, 但却与人类生活息息相关, 其种类和种群数量常常被用于表征生态系统稳定性, 在维持生态系统完整和保障生态系统健康中举足轻重[4-5]。因此, 两栖爬行动物可作为生态系统健康的重要指示类群, 能够尽早预警环境变化[3, 6]。
近几十年来, 全球两栖爬行动物面临着诸多威胁, 其种群数量快速下降[7-9]。根据IUCN最新结果[9], 受威胁物种包括41%的两栖动物, 这一比例远高于鸟类(14%)和哺乳动物(26%);我国最新评估结果显示两栖类和爬行类受威胁物种分别占43.1%和29.7%[10], 显而易见, 两栖爬行动物的处境非常严峻。土地利用变化是导致生物多样性丧失的最主要因子, 而两栖爬行动物是全球最易受威胁脊椎动物的直接典型代表[10-13]。然而, 对土地利用变化如何影响两栖爬行动物几乎没有共识[11]。国外部分学者评估了最常见的土地利用, 如农业、畜牧业、城市化、毁林、造林和选择性采伐等对两栖爬行动物群落的影响[14-15];但国内仅有少数研究, 是以IUCN红色名录为基础[10-11], 近年来, 国内关于城市化对生物多样性影响的研究逐渐增多, 如人类活动对上海当地生物多样性造成显著影响, 但湿地的增加对两栖和爬行动物的丰富度具有显著促进作用[16];内蒙古城市化造成物种空间分布格局发生变化[17]等。
城市生物多样性是保障城市永续发展的物质基础, 是实现人与动物和谐共生的基础。2019年, 北京城镇化率为86.6%, 如何在高度城市化的地区实现生物资源可持续利用是研究的重点和难点。因此, 本文以北京市两栖爬行动物调查数据为基础, 结合土地利用数据, 分析土地利用变化下两栖爬行动物的分布格局, 以期为生物资源可持续利用提供支撑。
1 材料与方法 1.1 研究区域北京市地处华北平原西北部, 辖16个市辖区。境内有太行山脉和燕山山脉, 地势由西北向东南倾斜, 地形以山地、丘陵为主, 东南部为平原;山地海拔一般1000-1500 m, 东灵山海拔较高, 为2303 m。地表水资源量8.61亿m3, 地下水资源量为15.95亿m3;18座大、中型水库可利用来水量为7.06亿m3;境内水系可划分为潮白河、永定河、北运河、大清河和蓟运河五大水系。多年平均降水量约为644 mm;典型的温带大陆性季风气候, 四季分明。自然地理条件、气候植被类型以及人类活动等因素决定了北京境内物种的分布、组成与多样性空间格局。
1.2 数据来源及分析数据包括土地利用数据、水质数据、两栖爬行动物野外调查数据。其中, 土地利用数据包含1980、1990、2000、2010、2015年和2018年6期, 来源于中国科学院资源环境数据中心, 数据分辨率为1 km;水质数据采用北京市水务局网站1980-2019年北京市水资源状况公报(http://swj.beijing.gov.cn/zwgk/szygb/);两栖爬行动物数据来源于野外调查, 其调查方法和鉴定依据如下。
1.2.1 野外调查方法北京两栖爬行动物调查分为试点和全面调查两个时间段。试点调查区域为门头沟区和延庆区, 调查时间为2017-2018年4-9月;全面调查区域涵盖其余14区, 调查时间为2020年4-9月。首先, 按照《县域两栖类和爬行类多样性调查与评估技术规定》(原环境保护部2017年第84号公告)将北京市划分为212个10 km×10 km的网格;然后, 秉承区域全覆盖、重点突出、生境代表性、道路可达性等原则, 选取调查网格94个;再次, 根据植被类型、湿地分布、湖滨灌草丛、水流方向、人类干扰等生境因子, 结合文献记载、事先居民访谈等获取的两栖爬行动物分布情况, 合理设置调查样线。样线长度视地形进行调整, 生境复杂时设置多条短样线;样线宽度依视野范围而定。每条样线进行徒步调查, 2人一组, 行进时注意观察搜寻各自周边5-10 m的范围, 记录物种名称、数量、经纬度、生境、干扰等数据;重点调查溪流、湖库、池塘及林间小路, 并特别留意两栖爬行动物易隐藏的小生境, 如枯枝落叶堆、雨后小水潭、石洞等。
两栖动物以夜行性为主, 爬行动物昼行性和夜行性兼而有之, 因此, 为力求覆盖各类群活动时间, 每条样线分白天和夜间2个时段调查。每条样线调查两次, 共计进行4次重复调查, 涉及样线1378条。调查过程中, 采用生物多样性调查移动APP进行相关数据录入。
1.2.2 物种鉴定物种鉴定依据《北京鱼类和两栖、爬行动物志》(1993)[18]、《中国两栖动物及其分布彩色图鉴》(2012)[19]、《中国动物志》[20-22]等。物种红色名录等级依据《中国脊椎动物红色名录》[23], 物种保护等级依据《国家重点保护野生动物名录》(2021年)、北京市一级保护野生动物名录(北京市园林绿化局公告2007年第1号)、北京市二级保护野生动物名录(北京市园林绿化局公告2007年第2号)。对于野外难以识别或鉴定存疑物种采集个体标本, 用于实验室测定形态数据和分类鉴定。测量工具采用电子数显游标卡尺, 精确到0.1 mm。标本以8%-10%福尔马林溶液固定, 回到室内经清水冲洗, 最终以75%酒精保存。
1.2.3 分析方法(1) 物种丰富度
物种丰富度采用物种种类数计算。
(2) 优势度
采用Berger-Parker优势度指数进行计算[24],
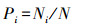
|
Pi表示优势度指数;Ni表示第i个物种的个体数;N表示总个体数。
(3) Shanoon多样性指数

|
Pielou均匀度指数

|
Margalef丰富度指数
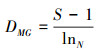
|
式中, Pi是频度;S是类群数目;Ni是第i个类群的个体数;N是所有类群总个体数。
(4) 多样性空间格局
利用AcrGIS的Extract功能, 获得每个物种点的海拔、生境, 然后进行分类统计;在每个10 km×10 km的栅格上, 统计物种种类, 制作物种密度图;分析全域生态系统类型变化与物种信息的位置关系;参考张利等的研究[25], 将1500 m作为两栖类的最大迁徙距离, 研究迁徙距离内生态类型变化对两栖动物生境的影响。此处仅分析了两栖动物生境影响, 由于爬行动物迁移距离不确定, 因此未展开分析。具体分析过程如下:
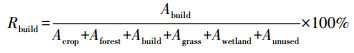
第一步, 以物种点为中心, 将其四周1500 m的缓冲区作为最大迁徙距离;第二步, 将获取的多个物种点的缓冲区进行dissolve处理, 获取唯一的迁徙距离边界;第三步, 以该边界为掩膜, 分别提取1980、1990、2000、2010、2015年和2018年6期生态系统数据, 包含农田、森林灌丛、草地、水域、建设用地、未利用地6种类型;第四步, 计算每个栅格上建设用地的百分比, 并利用ArcGIS的柱状图功能进行展示。每个柱状图由6种颜色构成, 分别代表 6个年份的建设用地百分比, 以此表征建设用地的影响程度。其中, 建设用地百分比计算过程如下:
|
式中, Rbuild表示建设用地百分比;Acrop、Aforest、Abuild、Agrass、Awetland、Aunused分别表示农田、森林灌丛、建设用地、草地、湿地和未利用地的面积。
2 结果分析 2.1 物种组成北京市两栖爬行动物多样性调查共获取有效物种信息1290条, 记录2纲3目10科18属24种(表 1), 其中两栖类的牛蛙(Rana catesbeiana)和爬行类的红耳龟(Trachemys scripta)为外来物种, 实际记录本地两栖爬行动物2纲3目10科17属22种, 后续分析仅考虑本地物种。记录两栖类1目3科4属5种, 包含蟾蜍科(Bufonidae)1属1种, 蛙科(Ranidae)2属3种, 姬蛙科(Microhylidae)1属1种;单型科3个, 单型属3个;以蛙科种类和数量最多, 优势种为林蛙属(Rana)的中国林蛙(R. chensinensis), 占两栖类物种总数的59.28%;其次为侧褶蛙属(Pelophylax)的黑斑侧褶蛙(P. nigromaculatus), 占24.54%;蟾蜍属(Bufo)中华蟾蜍(B. gargarizans)占15.02%。爬行类2目7科13属17种, 包含鳖科(Trionychidae)1属1种, 地龟科(Geoemydidae)1属1种, 壁虎科(Gekkonidae)1属1种, 石龙子科(Scincidae)1属1种, 蜥蜴科(Lacertidae)1属2种, 游蛇科(Colubridae)7属9种, 蝰科(Viperidae)1属2种;单型科6个, 单型属10个;游蛇科种类最多;优势种为麻蜥属(Eremias)的山地麻蜥(E. brenchleyi), 占爬行类物种数量的66.79%;壁虎属(Gekko)的无蹼壁虎(G. swinhonis)占17.45%。
| 纲、目、科、属 Class, Order, Family, Genus |
物种 Species |
中国生物多样性红色名录等级 Red list of biodiversity in China |
特有种 Endemic species |
保护等级 Protection level |
三有物种 List of important ecological, scientific and social value |
| 爬行纲Reptilia | |||||
| 龟鳖目Testudoformes | |||||
| 鳖科Trionychidae | |||||
| 鳖属Pelodiscus | 中华鳖P. sinensis | EN | 否 | 是 | |
| 地龟科Geoemydidae | |||||
| 拟水龟属Mauremy | 乌龟M. reevesii | EN | 否 | 国家Ⅱ级 | 是 |
| 彩龟属Trachemys | 红耳龟T. scripta | ||||
| 有鳞目Squamata | |||||
| 蜥蜴亚目Lacertilia | |||||
| 壁虎科Gekkonidae | |||||
| 壁虎属Gekko | 无蹼壁虎G. swinhonis | VU | 是 | 是 | |
| 石龙子科Scincidae | |||||
| 石龙子属Plestiodon | 黄纹石龙子P. capito | LC | 是 | 北京一级 | 是 |
| 蜥蜴科Lacertidae | |||||
| 麻蜥属Eremias | 丽斑麻蜥E. argus | LC | 否 | 是 | |
| 山地麻蜥E. brenchleyi | LC | 否 | 是 | ||
| 蛇亚目Ophidia | |||||
| 游蛇科Colubridae | |||||
| 玉斑蛇属Euprepiophis | 玉斑锦蛇E. mandarinus | VU | 否 | 北京一级 | 是 |
| 锦蛇属Elaphe | 赤峰锦蛇E. anomala | VU | 是 | 北京二级 | 是 |
| 王锦蛇E. carinata | EN | 否 | 北京一级 | 是 | |
| 白条锦蛇E. dione | LC | 否 | 北京二级 | 是 | |
| 晨蛇属Orthriophis | 黑眉锦蛇O. taeniurus | EN | 否 | 北京二级 | 是 |
| 颈槽蛇属Rhabdophis | 虎斑颈槽蛇R. tigrinus | LC | 否 | 北京二级 | 是 |
| 乌梢蛇属Zaocys | 乌梢蛇P. dhumnades | VU | 否 | 北京二级 | 是 |
| 游蛇属Coluber | 黄脊游蛇C. spinalis | LC | 北京二级 | 是 | |
| 链蛇属Dinodon | 赤链蛇D.rufozonatum | LC | 否 | 北京二级 | 是 |
| 蝰科Viperidae | |||||
| 亚洲蝮属Gloydius | 短尾蝮G. brevicaudus | NT | 否 | 北京二级 | 是 |
| 华北蝮G. stejnegeri | 未收录 | ||||
| 两栖纲Amphibia | |||||
| 无尾目Anura | |||||
| 蟾蜍科Bufonidae | |||||
| 蟾蜍属Bufo | 中华蟾蜍B. gargarizans | LC | 否 | / | 是 |
| 蛙科Ranidae | |||||
| 侧褶蛙属Pelophylax | 黑斑侧褶蛙P. nigromaculatus | NT | 否 | 北京二级 | 是 |
| 金线侧褶蛙P. plancyi | LC | 是 | 北京一级 | 是 | |
| 林蛙属Rana | 中国林蛙R. chensinensis | LC | 是 | 北京二级 | 是 |
| 牛蛙R. catesbeiana | |||||
| 姬蛙科Microhylidae | |||||
| 狭口蛙属Kaloula | 北方狭口蛙K. borealis | LC | 否 | 是 | |
| EN: 濒危Endangered; VU: 易危Vulnerable; LC: 无危Least concern; NT: 近危Near threatened | |||||
根据《中国生物多样性红色名录--脊椎动物卷》, 濒危4种, 易危4种, 近危2种;中国特有种5种, 占总种数的23.81%;三有动物21种;国家Ⅱ级保护物种1种;北京市一级保护物种4种, 二级保护物种10种, 保护物种占总种数的68.19%, 表明北京市两栖爬行动物保护的重要性及紧迫性。
2.2 不同海拔梯度物种分布将北京市域划分为 < 50 m、50-100 m、100-200 m、200-500 m和>500 m 5个海拔梯度, 统计不同梯度上的物种丰富度和物种数量, 各海拔梯度上的样线数分别为58、40、45、95和102条(图 1和表 2)。在物种丰富度上, 200-500 m丰富度最高, 记录物种18种, 包括5种两栖动物和13种爬行动物, 分别占两栖和爬行动物物种总数的100%和76.47%;Shannon多样性指数和Margalef丰富度指数均为最高, Pielou均匀度指数排第2位;其次为>500 m的海拔范围, 分布着4种两栖动物和12种爬行动物, Margalef丰富度指数排在第2位, Pielou均匀度指数偏低;50 m以下分布着4种两栖动物和7种爬行动物, Shannon多样性指数最低。在物种数量上, >500 m的海拔范围物种数量最多, 达到16种, 其数量占物种总数的46.42%, 几乎一半的物种都分布在这一范围内;200-500 m内物种数量占32.39%;由此可见, 北京市两栖爬行动物主要分布在海拔200 m以上的地段, 从空间上来看, 主要分布在北京西北部和东北部, 涉及延庆、门头沟、怀柔、密云、平谷等。

|
| 图 1 北京市不同海拔梯度两栖爬行动物丰富度和多度比较 Fig. 1 Comparison of richness and numbers of amphibians and reptiles at different altitudes in Beijing |
| 物种Species | < 50 m | 50-100 m | 100-200 m | 200-500 m | >500 m |
| 中华鳖P. sinensis | 2 | 5 | |||
| 乌龟M. reevesii | 1 | ||||
| 无蹼壁虎G. swinhonis | 4 | 13 | 9 | 49 | 70 |
| 黄纹石龙子P. capito | 6 | ||||
| 丽斑麻蜥E. argus | 25 | 3 | |||
| 山地麻蜥E. brenchleyi | 4 | 5 | 12 | 117 | 417 |
| 玉斑锦蛇E. mandarinus | 3 | ||||
| 赤峰锦蛇E. anomala | 1 | 8 | 14 | ||
| 王锦蛇E. carinata | 1 | ||||
| 白条锦蛇E. dione | 1 | 6 | |||
| 黑眉锦蛇O. taeniurus | |||||
| 虎斑颈槽蛇R. tigrinus | 1 | 1 | |||
| 乌梢蛇P. dhumnades | 4 | 1 | |||
| 黄脊游蛇C. spinalis | 1 | 1 | 3 | 8 | |
| 赤链蛇D.rufozonatum | 1 | 3 | 9 | 8 | |
| 短尾蝮G. brevicaudus | 1 | 1 | |||
| 华北蝮G. stejnegeri | 7 | ||||
| 中华蟾蜍B. gargarizans | 74 | 51 | 21 | 215 | 79 |
| 黑斑侧褶蛙P. nigromaculatus | 229 | 78 | 185 | 205 | 22 |
| 金线侧褶蛙P. plancyi | 1 | 7 | 5 | 1 | |
| 中国林蛙R. chensinensis | 15 | 1 | 68 | 552 | 1101 |
| 北方狭口蛙K. borealis | 6 | 2 | 17 | 1 | |
| 物种种类数 Number of species |
11 | 8 | 8 | 18 | 16 |
| Shannon多样性指数 Shannon′s diversity index |
1.010 | 1.286 | 1.186 | 1.628 | 1.134 |
| Pielou均匀度指数 Pielou′s evenness index |
0.421 | 0.618 | 0.570 | 0.563 | 0.409 |
| Margalef丰富度指数 Margalef′s richness index |
1.717 | 1.383 | 1.224 | 2.393 | 2.010 |
从表 2可以看出, < 50 m、50-100 m、100-200 m这3个海拔梯度的优势种均为黑斑侧褶蛙, 优势度分别为0.6735、0.5132、0.6086;200-500 m和>500 m海拔梯度的优势种为中国林蛙, 优势度分别为0.4525、0.6299。整体来看, 爬行动物数量较少, 优势种为山地麻蜥, 主要分布在200-500 m和>500 m的海拔段, 优势度分别为0.0959、0.2386。中国林蛙、黑斑侧褶蛙、中华蟾蜍、山地麻蜥、无蹼壁虎在各个海拔段均有分布, 相对而言可划分为广布种;综合上述, 两栖类分布相对广泛, 爬行类主要活动范围在200 m以上, 且丰富度和优势度均较低。
2.3 不同生境类型物种分布两栖爬行动物生境可划分为森林灌丛、草地、农田和湿地4种类型, 统计不同生境类型的物种丰富度和物种相对多度(图 2和表 3), 各生境类型的样线数分别为49、61、50和134条。在物种丰富度上, 森林灌丛最高, 记录物种19种, 包括5种两栖动物和14种爬行动物, 分别占两栖和爬行动物物种总数的100%和87.5%;Margalef丰富度指数最大, Shannon多样性指数排在第2位, Pielou均匀度指数偏低;草地分布着两栖动物5种和爬行动物7种;Margalef丰富度指数和Shannon多样性指数均最低;农田分布两栖动物5种和爬行动物6种;Margalef丰富度指数和Shannon多样性指数排在第3位;湿地分布两栖动物4种和爬行动物10种;Margalef丰富度指数仅次于森林灌丛, Shannon多样性指数排在第2位, Pielou均匀度指数最高。在物种相对多度上, 森林和灌丛最多, 占物种数量的41.49%;其次为草地, 占30.36%;再次为湿地, 占16.54%。从空间上来看, 主要分布在北京西北部, 该地带以森林灌丛为主, 植被覆盖度较高, 是两栖爬行动物的主要栖息地。

|
| 图 2 北京市不同生境类型两栖爬行动物丰富度和数量比较 Fig. 2 Comparison of richness and numbers of amphibians and reptiles in different habitats in Beijing |
| 物种 Species |
森林灌丛 Forest and shrubbery |
草地 Grassland |
农田 Farmland |
湿地 Wetland |
| 中华鳖P. sinensis | 7 | |||
| 乌龟M. reevesii | 1 | |||
| 无蹼壁虎G. swinhonis | 56 | 1 | 23 | 65 |
| 黄纹石龙子P. capito | 6 | |||
| 丽斑麻蜥E. argus | 23 | 5 | ||
| 山地麻蜥E. brenchleyi | 418 | 71 | 7 | 59 |
| 玉斑锦蛇E. mandarinus | 3 | 3 | ||
| 赤峰锦蛇E. anomala | 13 | 7 | ||
| 王锦蛇E. carinata | 1 | |||
| 白条锦蛇E. dione | 2 | 3 | 2 | |
| 黑眉锦蛇O. taeniurus | 1 | |||
| 虎斑颈槽蛇R. tigrinus | 2 | 1 | 1 | 1 |
| 乌梢蛇P. dhumnades | 1 | |||
| 黄脊游蛇C. spinalis | 8 | 3 | 2 | |
| 赤链蛇D.rufozonatum | 11 | 6 | 3 | 1 |
| 短尾蝮G. brevicaudus | 1 | 1 | ||
| 华北蝮G. stejnegeri | 7 | |||
| 中华蟾蜍B. gargarizans | 192 | 69 | 65 | 114 |
| 黑斑侧褶蛙P. nigromaculatus | 198 | 137 | 195 | 189 |
| 金线侧褶蛙P. plancyi | 6 | 1 | 1 | |
| 中国林蛙R. chensinensis | 604 | 840 | 114 | 179 |
| 北方狭口蛙K. borealis | 4 | 1 | 19 | 2 |
| Shannon多样性指数Shannon′s diversity index | 1.651 | 0.945 | 1.467 | 1.660 |
| Pielou均匀度指数Pielou′s evenness index | 0.561 | 0.380 | 0.612 | 0.629 |
| Margalef丰富度指数Margalef′s richness index | 2.449 | 1.562 | 1.648 | 2.019 |
从表 3可以看出, 黑斑侧褶蛙、中国林蛙、中华蟾蜍、山地麻蜥、无蹼壁虎在不同生境中的优势度较大。具体来说, 森林灌丛的优势种为中国林蛙, 优势度为0.3877;其次为山地麻蜥, 优势度为0.2683;然后为黑斑侧褶蛙和中华蟾蜍, 优势度分别为0.1271、0.1232。草地的优势种为中国林蛙, 优势度为0.7356, 其次为黑斑侧褶蛙, 优势度为0.1199, 二者占据了绝对优势;农田和湿地的优势种为黑斑侧褶蛙, 优势度分别为0.4462、0.3038。中国林蛙、黑斑侧褶蛙、中华蟾蜍、北方狭口蛙、山地麻蜥、无蹼壁虎、赤链蛇、虎斑颈槽蛇8个物种在4种生境类型均有分布。总体而言, 爬行动物数量较少, 优势种为山地麻蜥, 主要分布在森林灌丛、草地, 优势度分别为0.2683、0.0622;两栖动物数量多且分布相对广泛, 这一结论与不同海拔梯度两栖类分布广泛的结果一致。
2.4 空间分布格局 2.4.1 空间分布两栖爬行动物空间分布呈现由西北向东南递减的趋势(图 3):以西北部山区最为丰富, 包括门头沟、延庆、怀柔、房山、昌平、密云, 在功能区划上隶属于生态涵养区。这6区的主要植被类型涵盖落叶落叶林、常绿针叶林、落叶针叶林、针阔混交林、常绿阔叶灌丛、落叶阔叶灌丛、常绿针叶灌丛, 约占市域总面积的57.45%;区内植被覆盖度较高, 人为破坏较小, 动植物生境保护较完整, 集中分布了所调查到的全部两栖动物和15种爬行动物。北京东南部的城六区和平原区由于城市化和农业化程度较高, 调查到的物种种类及数量则相对较少。

|
| 图 3 北京市两栖爬行动物空间分布 Fig. 3 Spatial distribution of amphibians and reptiles in Beijing |
北京市一级保护野生动物金线侧褶蛙仅调查到8只, 密云5只, 房山、通州、延庆各1只;黄纹石龙子仅在门头沟百花山发现;王锦蛇仅在房山拒马河市级水生野生动物自然保护区发现1条。北京市二级保护野生动物中两栖纲的中国林蛙和黑斑侧褶蛙数量较多, 分布范围也较广;爬行纲的动物数量少, 且主要分布于门头沟、怀柔、延庆、房山。
2.4.2 不同市辖区物种分布由表 4可以看出, 门头沟和延庆物种种类最多, 均为14种, 占调查总种数的66.67%;房山12种, 占57.14%;怀柔10种, 占47.62%;密云和顺义均为9种, 占42.86%;昌平7种, 占33.33%;在保护物种方面, 延庆10种, 门头沟9种, 怀柔7种, 房山和密云均为6种。在物种数量上, 门头沟最多, 占物种数量的39.13%, 其次为房山(19.71%), 然后为怀柔(12.70%), 这3区集中分布了北京市域71.54%的物种数量。不同地区物种分布态势再次印证了北京市两栖爬行动物总体空间分布格局。
| 区名 Administrative Region |
物种数 Species number |
占比/% Proportion |
保护物种数 Number of protected species |
数量/只 Number |
比例/% Proportion |
| 昌平区 | 7 | 33.33 | 4 | 184 | 4.90 |
| 朝阳区 | 5 | 23.81 | 3 | 91 | 2.42 |
| 大兴区 | 3 | 14.29 | 1 | 6 | 0.16 |
| 房山区 | 12 | 57.14 | 6 | 740 | 19.70 |
| 丰台区 | 3 | 14.29 | 2 | 4 | 0.11 |
| 海淀区 | 4 | 19.05 | 2 | 41 | 1.09 |
| 东城区 | 1 | 4.76 | 1 | 1 | 0.03 |
| 西城区 | 1 | 4.76 | 1 | 1 | 0.03 |
| 怀柔区 | 10 | 47.62 | 7 | 477 | 12.70 |
| 门头沟区 | 14 | 66.67 | 9 | 1470 | 39.13 |
| 密云区 | 9 | 42.86 | 6 | 193 | 5.14 |
| 平谷区 | 5 | 23.81 | 2 | 59 | 1.57 |
| 石景山区 | 2 | 9.52 | / | 2 | 0.05 |
| 顺义区 | 9 | 42.86 | 5 | 249 | 6.63 |
| 通州区 | 4 | 19.05 | 3 | 27 | 0.72 |
| 延庆区 | 14 | 66.67 | 10 | 212 | 5.64 |
两栖爬行动物共计调查网格94个, 占总网格的44.34%。每个网格的物种种类在1-8之间;将其划分为1-2、2-4、4-6、6-8 4类, 所占比例分别为44.68%、26.60%、17.02%和11.70%, 可见, 包含4种以上物种的网格较少, 占比为28.72%。
从图 4可以看出, 物种数达到8种的网格有2个, 分别位于延庆大滩次生林保护区和怀柔宝山镇菜食河附近;此外, 包含6种以上的网格有9个, 主要位于延庆官厅水库、怀柔怀九河上游、门头沟百花山和永定河山峡段、房山拒马河、密云黄花岭等地;优势种为中国林蛙、黑斑侧褶蛙、山地麻蜥, 这些区域水源丰富, 水生植物长势良好, 自然生境保存较完整, 是两栖爬行动物生存与繁衍的良好栖息地。物种数为4-6种的网格主要分布在怀柔西银沟、延庆白河堡水库和莲花山、密云雾灵山和密云水库、门头沟百花山、房山石花洞和蒲洼、朝阳亮马河、顺义汉石桥湿地和唐指山等地;优势种为中国林蛙、黑斑侧褶蛙、中华蟾蜍, 这些区域大部分都已经建立了自然保护区、湿地保护区等多种类型的保护地, 在一定程度上表明保护地效果较好。物种数为2-4的网格主要在延庆玉渡山、怀柔怀九河和喇叭沟门、密云云蒙山、丰台永定河平原段、大兴大龙河和永定河平原段等地;优势种为中国林蛙、黑斑侧褶蛙、山地麻蜥。

|
| 图 4 不同网格的物种种类空间分布 Fig. 4 Spatial distribution of species in different survey grid |
两栖爬行动物的主要生境为森林灌丛、草地、农田和湿地, 因此, 统计1980、1990、2000、2010、2015年和2018年生境变化情况(图 5)。由图 5可见, 1980-2018年, 湿地、森林灌丛整体略有增加, 增幅分别为0.09%和1.57%;草地稍有减少, 减幅0.28%;农田减少最多, 减幅为13.11%;建设用地增幅11.62%, 且主要来自于农田转换(图 6)。

|
| 图 5 1980-2018年两栖爬行动物生境比例变化 Fig. 5 Changes in habitat proportion of amphibians and reptiles during 1980-2018 |

|
| 图 6 1980-2018年北京两栖爬行动物生境变化及其空间转换 Fig. 6 Habitat change and spatial transformation of amphibians and reptiles in Beijing during 1980-2018 |
由于物种数据只有1期, 因此统计2018年各行政区划内4种主要生境类型中两栖爬行动物的丰富度, 并利用SPSS软件进行显著性分析(表 5)。结果表明, 生境与物种丰富度之间存在显著相关性, 相关系数为0.722, 在0.01水平上显著相关。
| 相关性Significance analysis | VAR00001 | VAR00002 |
| VAR00001 Pearson相关性显著性(双侧) | 1 | 0.722** |
| Pearson correlation significance (bilateral)N | 0.000 | |
| 56 | 56 | |
| VAR00002 Pearson相关性显著性(双侧) | 0.722** | 1 |
| Pearson correlation significance (bilateral)N | 0.000 | |
| 56 | 56 | |
| ** 在0.01水平(双侧)上显著相关 | ||
根据调查结果, 农田中的两栖爬行物种最少, 原因可能是, 农田减少最大, 而它又是非常重要的栖居地, 生境的破坏造成物种种类和数量骤减;两栖爬行动物对环境变化极其敏感, 生境一旦破坏, 种群将很难修复[26]。
以两栖动物为例, 分析其1500 m迁徙距离内生境变化趋势(图 7)。可以看出, 1980-2018年, 迁徙距离内生境类型以森林灌丛为主;其次为农田, 其占比由1980年的26.92%下降至2018年的16.17%, 下降10.75%;草地面积略有减少, 由1980年的11.90%下降至2018年的11.64%, 下降0.26%;湿地面积整体增加0.36%;建设用地增加8.14%。从空间分布来看, 森林灌丛主要分布在西北部, 农田主要分布在城六区, 湿地在各区均有分布, 建设用地主要集中在建成区。

|
| 图 7 1980-2018年北京市两栖动物迁徙距离内生境空间分布 Fig. 7 Spatial distribution of amphibian habitats within migration distance during 1980-2018 in Beijing |
为了更直观的展示生境变化情况, 制作完成调查网格内建设用地百分比空间分布图(图 8)。可以看出, 建设用地变化较大的区域主要分布在北京东南部, 包括城六区、平原区(顺义、大兴、通州), 特别是城六区, 城镇化增速显著。1980年, 城六区还是以农田为主, 而城镇化扩张侵占了大量农田, 产生的大量污染物排至河流中, 导致两栖动物繁殖受到一定影响, 进而造成其数量不断减少。例如, 中国林蛙对水质要求较高, 因此, 城市河流中很难发现其身影, 本次调查仅在颐和园发现2只, 东坝郊野公园发现5只。中华蟾蜍和黑斑侧褶蛙是城六区的优势物种, 优势度0.4706和0.4314, 分别为主要分布在海淀颐和园、朝阳奥林匹克森林公园和东坝郊野公园。此外, 在颐和园发现2只北方狭口蛙, 其数量较少, 而在1989年的调查中有13个点位发现其身影, 但数量一般都少于3只, 可见, 北方狭口蛙的生存状态不容乐观, 需要加强保护和监管。在平原区, 优势种依然是黑斑侧褶蛙和中华蟾蜍。近年来, 水体水质好转, 水岸生态环境改善, 改善了两栖动物的生存环境。

|
| 图 8 北京市两栖动物迁徙距离内不同栅格建设用地百分比 Fig. 8 Percentage of construction land within the migration distance of amphibians in Beijing |
北京市保护地类型有自然保护区、森林公园、风景名胜区、地质公园、自然遗产地等(图 9), 面积约5038 km2, 占北京总面积的30.69%。各类保护地共覆盖两栖爬行类20种(占北京总种数的90.91%), 覆盖物种数量的60.45%;其中, 包含国家和北京一级和二级野生保护动物14种。保护地覆盖了绝大多数的两栖爬行动物, 且多数分布在北京西北部, 以保护区为例, 截至目前, 北京共有自然保护区21个, 其中, 国家级保护区2个, 市级保护区12个, 区级保护区7个;在行政区划上, 延庆10个, 密云3个, 平谷1个, 门头沟1个, 怀柔2个, 顺义1个, 房山3个;这些保护区以保护森林生态为主, 如百花山和松山国家级自然保护区;此外, 国家级森林公园17个, 湿地保护区45个。百花山国家级自然保护区、大滩区级自然保护区、霞云岭国家森林公园、喇叭沟门国家森林公园、白河堡区级自然保护区、拒马河市级水生野生动物自然保护区、银河谷森林公园、十三陵国家森林公园等两栖爬行物种丰富度较高。整体而言, 各类自然保护地虽覆盖了较大面积的物种, 但仍有部分物种未得到有效保护, 且种群数量较大, 如怀柔宝山镇白河上段、密云水库来水的潮河上段辛庄桥、平谷洳河大兴庄镇-峪口镇等, 目前存在较大的保护空缺(图 9)。

|
| 图 9 北京市自然保护地空间分布 Fig. 9 Spatial distribution of nature reserves in Beijing |
北京市虽地形较复杂, 但由于其纬度变化不大的自然地理环境特征, 因而两栖爬行动物种类并不丰富[28]。累计2017-2020年调查结果, 北京市共记录两栖类1目3科4属5种, 爬行类2目7科13属17种。在两栖动物方面, 与高武等1989年[27]和2004年[28]调查结果相比, 本次调查占历史记录的62.5%, 未发现花背蟾蜍(S. raddei)、东方铃蟾(B. orientalis)、大鲵(A. davidianus)。究其原因, 花背蟾蜍目前数量较少, 1989年, 高武[27]在延庆康庄、下营, 大兴庞各庄、黄村、岳家务等地虽有发现其身影, 但数量多数在10只以下;东方铃蟾只分布于香山、卧佛寺樱桃沟等局地范围, 种群规模相对固定, 本次调查未在此设置调查点位;大鲵虽之前被捕获到, 但并未发现其繁殖地, 近年来的相关调查也未见报道。在爬行动物方面, 由于高武等[28]的研究未列出爬行动物名录, 北京松山国家级自然保护区调查结果较少, 因此, 将本次调查结果与北京陆生野生动物名录(2021)、北京市一级保护野生动物名录(2007年)和北京市二级保护野生动物名录(2007年)进行对比, 未发现中国石龙子(一级)、双斑锦蛇(二级)、团花锦蛇(国家二级)、红纹滞卵蛇(二级)4种;调查发现保护动物11种, 已调查到的保护动物占比为64.71%。
在海拔的垂直分布上, 中国林蛙、黑斑侧褶蛙、中华蟾蜍、山地麻蜥、无蹼壁虎在各个海拔段均有分布, 可见, 它们可分布到海拔1500 m以上的山间溪流、山林、岩壁上, 如东灵山、百花山、松山等, 也可分布于海拔50 m的平原区, 这点与高武等[27]的研究不同;海拔200 m以下, 黑斑侧褶蛙分布较广, 海拔200 m以上, 中国林蛙分布较广。在生境类型分布上, 优势物种主要有黑斑侧褶蛙、中国林蛙、中华蟾蜍、山地麻蜥、无蹼壁虎等, 在森林灌丛、草地、农田和湿地等4种生境类型均有分布。
根据分析结果, 北京市两栖爬行动物总体呈现由西北向东南递减的趋势。究其原因, 生态涵养区人为干扰相对较小, 生境多样, 物种丰富度和多度较高;城六区虽城市化水平较高, 但由于湿地公园、城市公园、森林公园等保护地类型较多, 调查到的种类大于平原区;平原区由于以农田为主, 生境较为单一, 调查到的物种种类及数量则相对较少。综合而言, 随着生态环境提高, 生境逐渐转好, 有利于两栖爬行动物栖息地改善。
环境因素也影响两栖爬行动物分布。根据《北京市水资源公报》, 1986-2000年, 北运河水系污染较为严重, 主要污染物为氨氮和CODMn等;据文献记载[27], 中国林蛙只生活在氨氮含量0.5 mg/L以下的水环境, 此次调查中国林蛙主要分布在怀柔、门头沟、密云和房山, 调查结果也证实了这一点;黑斑侧褶蛙、中华蟾蜍也要求氨氮在10 mg/L以下, 高武等[28]在凉水河中段、通惠河下游未发现两栖动物。随着北京市水生态修复工程的实施, 水质逐渐改善, 2010、2015、2018、2020年, 达标河长分别占总评价河长的48%、53.4%、71.6%和87.1%, 黑斑侧褶蛙、中华蟾蜍在凉水河中下段亦庄湿地、大运河森林公园均有发现, 表明水质直接影响两栖动物的生存繁衍。
一般而言, 地形地貌、植被结构、生境等自然环境越复杂多变, 即空间异质性越高, 动物群落多样性也越高[3, 29]。此次调查结果凸显了影响两栖爬行动物种群生存的关键地形和生境, 为后续制定两栖爬行资源保护规划提供了数据支撑。
3.2 开展长期定位监测, 构建生物多样性保护网络在生物多样性监测领域, 研究的热点之一便是将两栖爬行动物作为监测物种[30]。两栖爬行动物为变温动物, 其分布受越冬地、食物来源等影响, 因此, 监测对象遴选、监测方法标准化、监测点位布设规范等, 逐渐成为保护生物学领域持续关注的议题[31]。监测中要充分考虑两栖爬行动物的繁殖季节、繁殖习性和栖息生境, 根据本次调查结果, 可在重要栖息地、重要生态节点, 如怀柔宝山镇白河上游、密云辛庄桥和密云水库、平谷海子水库, 朝阳奥林匹克森林公园、门头沟百花山、延庆大滩和官厅水库等地设置长期监测样地和样线, 加大监测力度;同时, 在显著位置设立提示标牌, 开展科普及宣传教育, 减少旅游活动对两栖爬行动物及其生境的影响。
此外, 在调查过程中发现, 部分宽阔道路的车流量较大, 路杀可能是影响两栖爬行动物生存的潜在威胁, 因此, 生物廊道构建既能解决这一问题又能提高道路附近动物群落多样性[32]。同时, 积极提倡整合多源数据, 建立大数据共享平台, 充分利用多方数据提升两栖爬行动物多样性监测能力, 逐步完善监测网络, 为北京生物资源保护与可持续利用提供支撑。
3.3 开展栖息地外来物种监测, 强化外来物种防控本次调查发现外来物种红耳龟和牛蛙, 数量较少, 并未形成种群规模。建议在东坝郊野公园等设置监测点, 开展外来物种专项调查, 监测其种群数量、生态习性、潜在风险等, 评估其对当地环境及物种的影响并采取相应防范措施;建立监测预警体系, 形成常态化监测。同时, 加强外来物种防控工作, 加大公众宣传, 增强公众对外来有害生物危害的认识, 在市内开展宣教活动, 在交通要道设立警示牌, 严禁人为放生外来物种, 并对已经引入的外来入侵物种应进行有效清除, 避免因肆意放养、放生造成外来物种入侵。
3.4 探索建立高强度土地利用下的多样化保护机制城市生物多样性能够满足人们追求诗意栖居的需求, 既有利于人与自然和谐相处, 又实现了城市生物资源的可持续利用。作为城镇化率极高的超大城市, 2019年, 北京城镇人口高达86.6%, 并且自2011年以来均保持在86%以上, 面对如此之高的城镇化率, 人与动物如何“共享”城市空间和资源, 实现生物多样性保护与城市发展协同增效, 开拓一种全新的多样化保护机制, 是当前迫切需要探讨的重要问题[33]。如, 近危物种黑斑侧褶蛙分布区很宽, 白天喜欢隐匿在农作物、水生植物或草丛中, 但由于过度捕捉和栖息地被侵占等原因, 其种群数量不断减少。因此, 要摒弃传统上将人类用地与生物多样性保护区域分离的传统发展模式, 探索将城镇用地、农业用地和物种栖息地有机融合, 构成一个互惠互利的生命共同体, 依托顶层设计, 以农业生产、道路建设、城市开发等为因, 以生态完整性、物种保护成效、遗传多样性、经济产出等为果, 建立全面有效的生物资源多样化保护与可持续利用机制。
| [1] |
李海东, 高吉喜. 生物多样性保护适应气候变化的管理策略. 生态学报, 2020, 40(11): 3844-3850. |
| [2] |
Mi X C, Feng G, Hu Y B, Zhang J, Chen L, Corlett R T, Hughes A C, Pimm S, Schmid B, Shi S H, Svenning J-C, Ma K P. The global significance of biodiversity science in China: an overview. National Science Review, 2021, 8(7): 1-25. |
| [3] |
李成, 谢锋, 车静, 江建平. 中国关键地区两栖爬行动物多样性监测与研究. 生物多样性, 2017, 25(3): 246-254. |
| [4] |
张勇, 龚大洁, 黄帅, 贺俊生, 李隆. 民勤连古城国家级自然保护区两栖爬行物种资源调查及分析. 干旱区资源与环境, 2020, 34(4): 148-153. |
| [5] |
Wake D B. Climate change implicated in amphibian and lizard declines. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(20): 8201-8202. DOI:10.1073/pnas.0702506104 |
| [6] |
Nori J, Lemes P, Urbina-Cardona N, Baldo D, Lescano J, Loyola R. Amphibian conservation, land-use changes and protected areas: a global overview. Biological Conservation, 2015, 191: 367-374. DOI:10.1016/j.biocon.2015.07.028 |
| [7] |
Stuart S N, Chanson J S, Cox N A, Young B E, Rodrigues A S L, Fischman D L, Waller R W. Status and trends of amphibian declines and extinctions worldwide. Science, 2004, 306(5702): 1783-1786. DOI:10.1126/science.1103538 |
| [8] |
Pimm S L, Jenkins C N, Abell R, Brooks T M, Gittleman J L, Joppa L N, Raven P H, Roberts C M, Sexton J O. The biodiversity of species and their rates of extinction, distribution, and protection. Science, 2014, 344(6187): 1246752. DOI:10.1126/science.1246752 |
| [9] |
IUCN. The IUCN red list of threatened speciesTM(2020). https://www.iucnredlist.org.
|
| [10] |
江建平, 谢锋, 臧春鑫, 蔡蕾, 李成, 王斌, 李家堂, 王杰, 胡军华, 王燕, 刘炯宇. 中国两栖动物受威胁现状评估. 生物多样性, 2016, 24(5): 588-597. |
| [11] |
蔡波, 李家堂, 陈跃英, 王跃招. 通过红色名录评估探讨中国爬行动物受威胁现状及原因. 生物多样性, 2016, 24(5): 578-587. |
| [12] |
Cordier J M, Aguilar R, Lescano J N, Leynaud G C, Bonino A, Miloch D, Loyola R, Nori J. A global assessment of amphibian and reptile responses to land-use changes. Biological Conservation, 2021, 253: 108863. DOI:10.1016/j.biocon.2020.108863 |
| [13] |
Spatz D R, Zilliacus K M, Holmes N D, Butchart S H M, Genovesi P, Ceballos G, Tershy B R, Croll D A. Globally threatened vertebrates on islands with invasive species. Science Advances, 2017, 3(10): e1603080. DOI:10.1126/sciadv.1603080 |
| [14] |
Thompson M E, Nowakowski A J, Donnelly M A. The importance of defining focal assemblages when evaluating amphibian and reptile responses to land use. Conservation Biology, 2016, 30(2): 249-258. DOI:10.1111/cobi.12637 |
| [15] |
Rubbo M J, Kiesecker J M. Amphibian breeding distribution in an urbanized landscape. Conservation Biology, 2005, 19(2): 504-511. DOI:10.1111/j.1523-1739.2005.000101.x |
| [16] |
王卿, 阮俊杰, 沙晨燕, 黄沈发, 王敏. 人类活动对上海市生物多样性空间格局的影响. 生态环境学报, 2012, 21(2): 279-285. DOI:10.3969/j.issn.1674-5906.2012.02.014 |
| [17] |
高明新, 潮洛濛, 阿斯茹. 内蒙古地区城市化对生物多样性的影响及维护措施. 环境与发展, 2016, 28(4): 1-8. |
| [18] |
王鸿媛. 北京鱼类和两栖·爬行动物志. 北京: 北京出版社, 1993.
|
| [19] |
费梁, 叶昌媛, 江建平. 中国两栖动物及其分布彩色图鉴. 成都: 四川科学技术出版社, 2012.
|
| [20] |
张孟闻, 宗愉, 马积藩. 中国动物志·第一卷·爬行纲, 总论龟鳖目鳄形目. 北京: 科学出版社, 1998.
|
| [21] |
赵尔宓, 黄美华, 宗愉. 中国动物志·第三卷·爬行纲, 有鳞目蛇亚目. 北京: 科学出版社, 1998.
|
| [22] |
赵尔宓, 赵肯堂, 周开亚. 中国动物志·第二卷·爬行纲, 有鳞目蜥蜴亚目. 北京: 科学出版社, 1999.
|
| [23] |
蒋志刚, 江建平, 王跃招, 张鹗, 张雁云, 李立立, 谢锋, 蔡波, 曹亮, 郑光美, 董路, 张正旺, 丁平, 罗振华, 丁长青, 马志军, 汤宋华, 曹文宣, 李春旺, 胡慧建, 马勇, 吴毅, 王应祥, 周开亚, 刘少英, 陈跃英, 李家堂, 冯祚建, 王燕, 王斌, 李成, 宋雪琳, 蔡蕾, 臧春鑫, 曾岩, 孟智斌, 方红霞, 平晓鸽. 中国脊椎动物红色名录. 生物多样性, 2016, 24(5): 500-551. |
| [24] |
陈卓, 刘典, 何玉晓, 朱艳军, 可灿, 冯琦琦, 徐海根, 陈永杰. 鼎湖山两栖动物多样性的时空格局变异. 河南师范大学学报: 自然科学版, 2020, 48(6): 76-83. |
| [25] |
张利, 何玲, 闫丰, 陈亚恒. 基于图论的两栖类生物栖息地网络规划--以黑斑侧褶蛙为例. 应用生态学报, 2021, 32(3): 1054-1060. |
| [26] |
姚冲学, 王智红, 梁良, 吕婷, 黄元, 陈明勇. 云南玉龙雪山两栖动物多样性时空格局的观测. 生态与农村环境学报, 2020, 36(6): 726-730. |
| [27] |
高武. 北京地区两栖动物区系及生态分布. 北京师范学院学报: 自然科学版, 1989, 10(2): 28-34. |
| [28] |
高武, 陈卫, 傅必谦. 北京的两栖爬行动物研究. 绿化与生活, 2004(6): 43. |
| [29] |
刘研, 何玉晓, 诸冠彦, 高朔, 徐海根, 李宏. 湖南南岳衡山两栖动物物种多样性及其时空格局. 生态与农村环境学报, 2020, 36(8): 975-981. |
| [30] |
Díaz-García J M, Pineda E, López-Barrera F, Moreno C E. Amphibian species and functional diversity as indicators of restoration success in tropical montane forest. Biodiversity and Conservation, 2017, 26(11): 2569-2589. DOI:10.1007/s10531-017-1372-2 |
| [31] |
Xu H G, Cao M C, Wu Y, Cai L, Cao Y, Ding H, Cui P, Wu J, Wang Z, Le Z F, Lu X Q, Liu L, Li J Q. Optimized monitoring sites for detection of biodiversity trends in China. Biodiversity and Conservation, 2017, 26(8): 1959-1971. DOI:10.1007/s10531-017-1339-3 |
| [32] |
Kaiser J. Building a case for biological corridors. Science, 2001, 293(5538): 2199. DOI:10.1126/science.293.5538.2199 |
| [33] |
段菲, 李晟. 黄河流域鸟类多样性现状、分布格局及保护空缺. 生物多样性, 2020, 28(12): 1459-1468. |