 2022, Vol. 42
2022, Vol. 42文章信息
- 付翔, 付成, 肖玲韬, 樊捷, 付世建
- FU Xiang, FU Cheng, XIAO Lingtao, FAN Jie, FU Shijian
- 活跃性和社会性对中华倒刺鲃集群行为的影响
- Effects of activity and sociability on shoaling behavior of Spinibarbus sinensis
- 生态学报. 2022, 42(8): 3206-3215
- Acta Ecologica Sinica. 2022, 42(8): 3206-3215
- http://dx.doi.org/10.5846/stxb202106021452
-
文章历史
- 收稿日期: 2021-06-02
- 网络出版日期: 2021-12-17

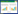




个性指不同时间和环境条件下动物表现出的一致且可重复的个体行为差异[1]。描述动物个性的特征有活跃性、社会性等[2-3], 前者是指个体在自发运动时的行为特征[4], 常用指标有自发游泳速度中值等[5-6]。社会性是指除了攻击性互动之外, 动物对同种类群亲密互动或者回避的反应[7], 常用指标包括与刺激鱼群的距离等[7-8]。研究发现包括鱼类在内的动物的个性特征在一定时间内具较好的稳定性[1], 例如鲫(Carassius auratus)幼鱼在间隔15 d前、后的活跃性和好斗性具有较好的重复性[9], 而蓝鳃太阳鱼(Lepomis macrochirus)的勇敢性在间隔3个月依旧具有较好的稳定性[10]。这种稳定的个性差异有利于种群在变化的环境中生存, 而群体成员的个体异质性可能有利于集体协作的进化[11-13]。
多数动物种类在其生活史的特定或全部阶段会偏好集群生活, 集群行为是动物在长期自然选择中进化形成的生存策略[4, 11]。在集群行为中个体被认为是“自我推进的粒子”, 相互作用被称为“社会力量”, 个体行为的微小变化可能会影响个体的空间定位, 并强烈的影响群体的运动特征[12]。研究群体行为的工作多热衷于探讨群体成员如何调整自己的行为, 从而使群体行为保持一致[13]。近年来, 研究者日益关注不同个性成员对群体行为产生影响的种内差异。这些研究表明, 个性特征可能与机体在群体决策发挥作用的大小密切关联[14-16]。例如, 有研究表明孔雀鱼(Poecilia reticulata)的群体探索行为主要受制于群体中最不活跃的群体成员[17], 而勇敢的河鲈(Perca fluviatilis)个体在群体行为中往往发挥着至关重要的影响力[18]。由于群体成员普遍存在个性和生理状况的差异, 探讨集群生活的个体如何在维持群体凝聚力的基础上权衡牺牲个性和维持特定的行为独立性, 具有重要的理论意义[12, 17]。
目前有关个性与鱼类集群行为的研究较少, 主要在孔雀鱼、三刺鱼(Gasterosteus aculeatus)等少数高度社会化且认知能力较高的鱼类中开展[12, 17]。这些研究发现, 鱼类的集群运动和群体探索倾向与群体成员的活跃性和社会性关联较为密切[17, 19]。鲤科鱼类是我国的第一大淡水鱼类类群, 且多数种类喜好集群, 但有关鲤科鱼类个性特征及其对集群行为的研究却鲜见报道。因此, 本研究选取偏好集群的鲤科鱼类-中华倒刺鲃(Spinibarbus sinensis)为研究对象[20-23], 首先考察其活跃性和社会性等个性特征及其稳定性。在此基础上, 本研究还旨在探究不同活跃性和社会性组成对中华倒刺鲃群体水平运动特征的影响及其内在机制, 以及个性特征与集群时的行为特征是否存在关联。本研究结果不仅有助于理解以中华倒刺鲃为代表的鲤科鱼类集群行为的形成机制, 还可能为鱼类物种保护和资源利用提供参考。
1 材料与方法 1.1 实验鱼来源与驯养实验鱼于2019年4月购买于重庆永川渔场, 将买回的实验鱼用1%-2%的氯化钠溶液消毒后放入实验室的自净化循环控温系统(每个养殖单元的体积约为250 L, 容纳125尾实验鱼)中驯养六周, 驯养期间无实验鱼死亡现象。驯养时水温控制在(25±1)℃, 水中溶氧维持在7 mg/L以上, 养殖用水使用充分曝气的自来水, 日换水量约10%, 光周期为12L: 12D。驯养期间, 每天在10:00时用商业颗粒饲料进行饱足投喂, 30 min后, 用虹吸管清理残存的食物和鱼类排泄物。驯养结束后, 选取大小适宜的实验鱼进行实验。
1.2 实验方案首先选择大小适宜的实验鱼336尾(体重(7.99±0.22)g, 体长(7.48±0.07)cm), 腹腔注射PIT(Passive integrated transponder, 直径2 mm, 长7 mm)体内标签(研究表明体内标签不会对鱼类的行为和生理状态造成影响[24])。待实验鱼恢复2周之后(标记及恢复过程无实验鱼死亡现象), 对每尾实验鱼依次进行活跃性和社会性的测定。测定结束后, 分别以实验鱼的活跃性和社会性为依据, 从高到低进行排序。将位于中间个性的个体除去, 分为高个性组、低个性组和混合个性组(高、低个性1:1混合), 每种个性组重复14组。提前一天对实验鱼进行体表的颜色标记(研究表明体表颜色标记不会对鱼类的个性造成影响[9, 25]), 第二天进行集群行为拍摄, 分别记录视频中6尾实验鱼群(以往研究发现, 中华倒刺鲃在个体数量为4尾及其以上时, 就会表现出明显的集群现象, 因此, 本实验选择6尾实验鱼组成群体)的运动轨迹。集群行为拍摄结束后, 实验鱼放回到原养殖系统进行饲养, 养殖条件不变。实验结束时随机选择80尾实验鱼, 重复测定活跃性和社会性, 用以衡量其个性的稳定性。所有的行为测定均在视频拍摄完成后清洗装置并换水。
1.3 实验参数的测定 1.3.1 个性(活跃性、社会性)的测定活跃性和社会性的测定使用的是实验室自制的行为测定仪, 为了消除人为活动对实验测定的影响, 所有实验装置被白色的幕布所遮盖。实验装置的水温等条件与驯养期间一致。为消除昼夜节律的影响, 所有拍摄工作在08:00-17:00之间进行。活跃性的测定装置为一个长方形水槽(图 1;55 cm×35 cm×35 cm), 装置由有机玻璃板组成, 整个装置内壁和底部用白色广告纸覆盖。装置内水深为10 cm。装置的正上方固定有一个连接计算机的网络摄像头(罗技Webcam C9000)。测定时将实验鱼带水转移至活跃性测定装置中适应10 min, 随后对实验鱼的自发活跃性进行10 min的拍摄(15帧/s)。拍摄的视频由QQ影音转码后, 再用idTracker提取实验鱼质心运动的轨迹用以后续分析。活跃性的测量参数为速度中值(cm/s), 计算公式如下:
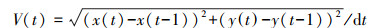
|
(1) |

|
| 图 1 用于活跃性、社会性和集群行为测定的实验装置示意图 Fig. 1 The structure of the devices used to observe the activity, sociality and shoaling behavior in the present study 图中a, c为刺激区; b为选择区; d, e为透明隔板 |
式中, x(t)和y(t)分别是实验鱼质心位置的横坐标和纵坐标, x(t-1)和y(t-1)分别是前一帧实验鱼质心位置的横坐标和纵坐标; dt是连续两帧之间的时间间隔(1/15 s)。
社会性测定装置为“鱼类行为二元选择观测仪”, 该测定装置同样也是由有机玻璃板组成的长方形水槽(图 1;70 cm×35 cm×35 cm), 水槽内壁和底部被白色广告纸所覆盖。整个装置被两块透明的有机玻璃板分隔为三部分:位于装置中间的选择区(40 cm×35 cm×35 cm)和位于两侧的两个刺激区(15 cm×35 cm×35 cm)。装置内水深为10 cm。测定时首先随机选取6尾中华倒刺鲃幼鱼置于实验装置的任一刺激区, 而另一刺激区空白。待刺激鱼群在实验装置中适应20 min后, 将单尾实验鱼带水转入选择区并立即进行视频拍摄, 拍摄时长为17 min(分析时前2 min不纳入统计)。社会性的测量参数为实验鱼与刺激鱼群的距离(cm), 该值越大表明实验鱼的社会性越低, 计算公式如下:
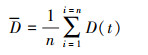
|
(2) |
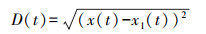
|
(3) |
式中, x(t)、x1(t)分别表示目标实验鱼及刺激鱼群一侧隔板(即图 1中的隔板e)在t时刻的横坐标, n表示视频的总帧数。
1.3.2 集群行为的测定集群行为使用直径为50 cm、高为25 cm的圆形装置完成测定(图 1), 装置内壁和底部均用白色广告纸覆盖。实验装置的水温等条件与驯养期间一致。为消除昼夜节律的影响, 所有拍摄工作在08:00到17:00之间进行。集群行为拍摄前一天, 先将实验鱼轻度麻醉(MS-222: NaHCO3=80 mg/L: 160 mg/L)后进行体外颜色标记:高、低个性(活跃性和社会性)的个体分别在背部标记不同的颜色。集群行为测定时将高个性组、低个性组和混合个性组依次放入上述装置中, 每组6尾实验鱼。实验鱼适应10 min后, 使用网络摄像头(罗技Webcam C9000)进行集群行为拍摄, 持续10 min。所有视频(15帧/s)用QQ影音转码后, 用idTracker软件进行分析, 得到每一尾鱼的运动轨迹。随后根据坐标点计算鱼群的集群运动和空间位置等参数。群体运动特征指标包括速度、同步性和运动时间比, 集群时每尾个体运动特征参数包括距离群体质心的距离、速度中值、运动时间比以及速度同步性。相关参数的计算方法参照以往相关研究[4, 12, 26-27]。相关参数的计算方法如下:
鱼群的质心(xG, yG):
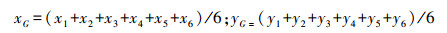
|
(4) |
距离鱼群质心的距离(D, cm):
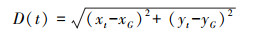
|
(5) |
式中, xG、yG分别为t时刻鱼群的质心坐标; x1、x2、x3、x4、x5、x6和y1、y2、y3、y4、y5、y6分别为t时刻6尾鱼的横、纵坐标。xt、yt为t时刻某一尾鱼的横纵坐标。距离鱼群质心的距离用于评价鱼群中某一尾鱼的集群倾向, 即凝聚力。
速度中值的计算公式同公式(1)。
速度同步性(Sv):

|
(6) |
式中, vi和vj是鱼群中任意两尾实验鱼在t时刻的游泳速度。用于评价鱼群运动时速度上协调性。运动时间比(PTM, %):速度大于1.75 cm/s的帧数除以总帧数[21]。
1.3.3 中华倒刺鲃幼鱼个性的稳定性测定集群行为拍摄结束以后, 实验鱼放回到原养殖系统进行饲养, 养殖条件不变。实验结束时随机选择80尾实验鱼, 重复测定其活跃性和社会性, 并与首次测定结果对比, 以此验证中华倒刺鲃的个性是否稳定。
1.4 数据统计分析本实验数据采用Excel 2010进行常规计算后, 使用SPSS 22.0进行统计分析, 所有统计值用平均值±标准误(Mean ± SE)表示, 显著性水平P < 0.05。首先检验实验数据的正态性和方差齐性, 如果不满足, 则使用非参数检验。活跃性和社会性稳定性的分析采用重复测量的可靠性分析。集群行为参数采用单因素方差分析(One-way ANOVA)和LSD多重比较检验差异显著性; 活跃性、社会性和集群参数的关联采用Pearson或Spearman相关性分析。
2 结果 2.1 中华倒刺鲃幼鱼个性的重复性中华倒刺鲃的活跃性和社会性存在较大的个体差异(Cv分别为54.3%和29.5%)。可靠性分析表明实验过程中(时间间隔约1个月)活跃性和社会性指标均具有较高的稳定性(活跃性:R=0.706, P < 0.001, 社会性:R=0.564, P=0.006, 图 2)。

|
| 图 2 活跃性和社会性的重复性 Fig. 2 The repeatability of activity and sociabilit |
高活跃性鱼群的速度同步性和运动时间比均显著高于低活跃性群体(P < 0.05;图 3), 而混合活跃性鱼群的上述两参数与高、低活跃性组间均无显著差异。

|
| 图 3 鱼群个性组成对中华倒刺鲃幼鱼集群行为特征的影响 Fig. 3 The effect of group personality composition on shoaling behavior characteristics in juvenile Qingbo (Mean±S.E. N=14) a、b不同字母表示不同个性组之间差异显著(P < 0.05) |
不同社会性群体在速度同步性上存在显著差异(P < 0.05), 其中高社会性群体显著小于低社会性组和混合社会性群体(P < 0.05;图 3)。
2.3 个性与群体中单尾实验鱼运动特征之间的关联对于活跃性而言, 个性测定时活跃性的指标(速度中值)与群体中单尾实验鱼的速度中值、运动时间比以及速度同步性均呈现显著正相关(P < 0.001, 表 1, 图 4)。
| 参数 Parameter |
距离质心的距离 The distance from the center/cm |
速度中值 Median speed/cm/s |
运动时间比 Percent time spent moving/% |
速度同步性 Synchronization of speed |
| 活跃性 | N=240 | N=240 | N=240 | N=240 |
| Activity | R=0.102 | R=0.248 | R=0.319 | R=0.263 |
| P=0.114 | P < 0.001** | P < 0.001** | P < 0.001** | |
| 社会性 | N=246 | N=246 | N=246 | N=246 |
| Sociability | R=0.159 | R=0.137 | R=0.204 | R=0.278 |
| P=0.013* | P=0.031* | P=0.001** | P < 0.001** | |
| (P < 0.05)表示实验鱼的个性与集群中每尾实验鱼运动特征参数之间具有显著关联 | ||||

|
| 图 4 个性与集群中单尾实验鱼运动特征参数之间的关联 Fig. 4 The relationships between personality traits and corresponding moving characteristics of individual fish during shoaling |
对于社会性而言, 社会性指标(与刺激鱼群的距离)不仅与速度中值(P=0.031)、运动时间比(P=0.001)和速度同步性显著正相关(P < 0.001), 还与距离质心的距离显著正相关(P=0.013)(表 1, 图 4)(即相关指标与社会性负相关)。
3 讨论 3.1 中华倒刺鲃幼鱼的个性及其对集群行为的影响中华倒刺鲃幼鱼的活跃性和社会性在一定时间内表现出较好的稳定性, 这与鲫、大西洋鳕(Gadus morhua)等鱼类的研究较为一致[9, 28]。本研究还发现中华倒刺鲃幼鱼的活跃性和社会性均表现出较大的个体差异, 这与大多数的其他鲤科鱼类较为类似[7, 26, 29]。最近的研究发现, 宽鳍鱲(Zacco platypus)的活跃性变异系数为21.93%, 社会性的变异系数高达52.5%[30]。研究者认为, 这种个性特征的差异有利于物种在不断变化的环境条件下生存, 具有重要进化和生态意义[31]。对中华倒刺鲃而言, 这种个性差异使得其在面临栖息地生境剧烈波动时, 至少种群的部分个体能够存活并适应新的环境条件, 进而该物种能够得以保全[23, 32]。其次, 大量研究表明中华倒刺鲃是一种群居性极强的鱼类[21, 33], 个性之间的这种差异可能有利于集体协作的形成, 提高了群体运动或决策的一致性[34-35]。
高活跃性群体在运动时间比上比低活跃性群体高出约24%, 这说明中华倒刺鲃鱼群个性组成的差异能够引起群体层面运动特征的改变, 这与Brown等人研究结果一致[4, 17]。其次, 高活跃性群体在速度同步性上比低活跃性群体高出约2.7%, 表明越活跃的鱼群, 其协调性越强, 这与鳊(Parabramis pekinensis)等鲤科鱼类研究相同[33, 36]。高社会性群体在速度同步性上比低社会性群体低了约4.1%, 这可能是因为高社会性群体的活动水平下降造成的(运动时间比较低社会性群体下降14%)。这表明社会性越强的个体, 其群体运动协调性可能越差。总之, 与其他鱼类相似[17, 19], 中华倒刺鲃的个性也会对其集群行为产生影响。个性对集群运动特征的影响可能进一步影响鱼群群体功能水平的性能表现。例如, 关于孔雀鱼的研究表明, 群体中不同勇敢性的组成对群体总体的觅食行为有重要影响; 并且在群体中, 勇敢的个体可能会使得害羞的个体变得勇敢, 从而提高群体的觅食效率[37]。
有关鲤科鱼类个性特征研究较少, 主要集中在鳊、鲫和中华倒刺鲃等少数鱼类[5-6]。这些研究发现, 鲤科鱼类个性特征存在较高的种间差异。在更多种类上开展不同个性鲤科鱼类个性对集群行为影响的比较研究可能产生具有重要理论价值的研究发现。
3.2 中华倒刺鲃幼鱼个性对集群行为影响的内在机制群体成员在完成协调一致的集群运动时, 往往需要形成群体决策(Group decision)或者说达成群体共识[38-39]。有研究表明, 在动物中某些少数个体的特定个性可能主要决定了群体运动的特征, 即少数决定机制(Minority-determined mechanism)[4, 17], 而某些个性是以所有群体成员该个性的整体状况来影响群体运动特征, 即平均决策机制(Average-determined mechanism)[40-41]。在本实验中, 我们发现在以活跃性划分的群体中, 群体的速度同步性和运动时间比在高、低活跃性群体上存在显著差异, 但混合活跃性群体与高、低群体却无显著差异。另外, 混合活跃性群体上述两个参数在数值上均介于高、低活跃性群体之间。这些结果表明, 对于中华倒刺鲃而言, 群体所有成员活跃性的状况共同影响了鱼群的集群行为的特征。这种平均决策机制在食蚊鱼(Gambusia affinis)等的研究中也得到证实[40]。特别值得一提的是, 在以社会性划分的不同群体中, 高社会性群体的速度同步性显著小于低社会性群体和混合社会性群体, 而后两者之间没有差异, 这表明鱼群中较低社会性的个体可能对群体运动特征表现出主导作用。另外, 虽然没有显著差异, 不同社会性群体在运动时间比上存在与速度协调性一致的趋势(低社会性和混合型群体数值上更为接近)。因此, 群体成员中低社会性的关键个体可能对中华倒刺鲃群体行为产生重要影响, 即少数决定机制[17-18]。在三刺鱼的研究也发现, 社会性较弱的个体出现在群体前面的几率更高, 因此更多的引导群体运动, 并且更有可能在觅食过程中率先发现食物[12, 42]。将来开展决定机制性质与鱼类种类、认知能力甚至生境之间的关联的深入研究可能对理解鱼类个性的产生和维持机制以及集群行为的进化具有重要意义。
实验鱼个体单独测定个性时的速度中值与集群运动时个体的速度中值、运动时间比以及速度同步性均呈显著正相关, 这表明活跃性即游泳速度在集群时能够在较大程度上得以保留。因此, 中华倒刺鲃鱼群的运动状态是由群体中所有个体共同决定的, 这很好的解释了上述活跃性对群体运动特征影响的平均决定机制。有趣的是, 实验鱼的社会性与速度中值、运动时间比以及速度同步性均呈显著负相关, 表明社会性越高的实验鱼在群体中的运动能力和协调性表现越差, 这可能同样是上述提到的高社会性鱼类较低的运动水平导致。特别值得一提的是, 与活跃性不同, 实验鱼的社会性还与群体凝聚力呈显著正相关, 这表明社会性越高的个体, 其凝聚力越强[43]。群体凝聚力是集群行为的重要特征, 很大程度上决定了群体的结构甚至分群、合群等重要行为[44-45]。因此, 社会性特征与凝聚力的关联可能是社会性对群体的影响为少数决定机制的重要原因。本研究首次把个性对鱼群行为特征影响及其与集群时每个成员行为特征的关联结合起来进行探讨, 发现鱼类集群行为特征和个性特征关联是鱼群决定机制的基础。该发现为鲤科鱼类的特有机制还是普遍存在多数喜好集群的鱼类类群, 值得研究者进一步深入探讨。
总之, 本研究发现中华倒刺鲃鱼群的活跃性和社会性组成均对集群行为产生重要影响, 但活跃性对群体运动特征的影响表现为平均决定机制, 而社会性对群体运动性能的影响表现为少数决定机制。这种内在机制的差异可能与个性特征在群体运动时的独立性的维持和妥协程度有关。在自然环境中, 鱼群可能会根据生态环境的变化自发进行分类, 形成具有不同个性特征的群体。因此, 中华倒刺鲃稳定的个体间个性变异、个性组成与鱼群运动特征的密切关联及其内在机制的复杂性可能有利于该物种在多变的环境条件下完成不同的生理活动, 具有重要的生态意义。
| [1] |
Biro P A, Stamps J A. Do consistent individual differences in metabolic rate promote consistent individual differences in behavior?. Trends in Ecology & Evolution, 2010, 25(11): 653-659. |
| [2] |
Réale D, Reader S M, Sol D, McDougall P T, Dingemanse N J. Integrating animal temperament within ecology and evolution. Biological Reviews, 2007, 82(2): 291-318. DOI:10.1111/j.1469-185X.2007.00010.x |
| [3] |
Sih A, Giudice M D. Linking behavioural syndromes and cognition: a behavioural ecology perspective. Philosophical Transactions of the Royal Society B: Biological Sciences, 2012, 367(1603): 2762-2772. DOI:10.1098/rstb.2012.0216 |
| [4] |
Tang Z H, Fu S J. Qingbo (Spinibarbus sinensis) personalities and their effect on shoaling behavior. Acta Ethologica, 2019, 22(2): 135-144. DOI:10.1007/s10211-019-00321-x |
| [5] |
Liu S, Fu S J. Effects of food availability on metabolism, behaviour, growth and their relationships in a triploid carp. Journal of Experimental Biology, 2017, 220(24): 4711-4719. |
| [6] |
曾令清, 付成, 奚露, 彭姜岚, 付世建. 鳊幼鱼能量代谢和个性行为的个体变异及表型关联. 生态学报, 2017, 37(14): 4807-4816. |
| [7] |
Killen S S, Fu C, Wu Q Y, Wang Y X, Fu S J. The relationship between metabolic rate and sociability is altered by food deprivation. Functional Ecology, 2016, 30(8): 1358-1365. DOI:10.1111/1365-2435.12634 |
| [8] |
Conrad J L, Weinersmith K L, Brodin T, Saltz J B, Sih A. Behavioural syndromes in fishes: a review with implications for ecology and fisheries management. Journal of Fish Biology, 2011, 78(2): 395-435. DOI:10.1111/j.1095-8649.2010.02874.x |
| [9] |
陈李雪, 冷霞, 曾令清. 颜色标记对鲫幼鱼个性的影响. 重庆师范大学学报: 自然科学版, 2020, 37(5): 28-34. |
| [10] |
Wilson A D M, Godin J G J. Boldness and behavioral syndromes in the bluegill sunfish, Lepomis macrochirus. Behavioral Ecology, 2009, 20(2): 231-237. DOI:10.1093/beheco/arp018 |
| [11] |
Miller N, Garnier S, Hartnett A T, Couzin I D. Both information and social cohesion determine collective decisions in animal groups. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(13): 5263-5268. DOI:10.1073/pnas.1217513110 |
| [12] |
Jolles J W, Boogert N J, Sridhar V H, Couzin I D, Manica A. Consistent individual differences drive collective behavior and group functioning of schooling fish. Current Biology, 2017, 27(18): 2862-2868. DOI:10.1016/j.cub.2017.08.004 |
| [13] |
Dall S R X, Houston A I, Mcnamara J M. The behavioural ecology of personality: consistent individual differences from an adaptive perspective. Ecology Letters, 2004, 7(8): 734-739. DOI:10.1111/j.1461-0248.2004.00618.x |
| [14] |
Ward A J W, Thomas P, Hart P J B, Krause J. Correlates of boldness in three-spined sticklebacks (Gasterosteus aculeatus). Behavioral Ecology and Sociobiology, 2004, 55(6): 561-568. DOI:10.1007/s00265-003-0751-8 |
| [15] |
Jolles J W, Fleetwood-Wilson A, Nakayama S, Stumpe M C, Johnstone R A, Manica A. The role of social attraction and its link with boldness in the collective movements of three-spined sticklebacks. Animal Behaviour, 2015, 99: 147-153. DOI:10.1016/j.anbehav.2014.11.004 |
| [16] |
Jolles J W, Laskowski K L, Boogert N J, Manica A. Repeatable group differences in the collective behaviour of stickleback shoals across ecological contexts. Proceedings of the Royal Society B: Biological Sciences, 2018, 285(1872): 20172629. DOI:10.1098/rspb.2017.2629 |
| [17] |
Brown C, Irving E. Individual personality traits influence group exploration in a feral guppy population. Behavioral Ecology, 2014, 25(1): 95-101. DOI:10.1093/beheco/art090 |
| [18] |
Magnhagen C, Bunnefeld N. Express your personality or go along with the group: what determines the behaviour of shoaling perch?. Proceedings of the Royal Society B: Biological Sciences, 2009, 276(1671): 3369-3375. DOI:10.1098/rspb.2009.0851 |
| [19] |
Sumpter D J T, Szorkovszky A, Kotrschal A, Kolm N, Herbert-Read J E. Using activity and sociability to characterize collective motion. Philosophical Transactions of the Royal Society B: Biological Sciences, 2018, 373(1746): 20170015. DOI:10.1098/rstb.2017.0015 |
| [20] |
丁瑞华. 四川鱼类志. 成都: 四川科学技术出版社, 1994.
|
| [21] |
Tang Z H, Wu H, Huang Q, Kuang L, Fu S J. The shoaling behavior of two cyprinid species in conspecific and heterospecific groups. PeerJ, 2017, 5: e3397. DOI:10.7717/peerj.3397 |
| [22] |
王蕾, 唐金玉, 覃英莲, 曾令清, 彭姜岚, 付世建. 饥饿对中华倒刺鲃幼鱼代谢、个性和集群的影响. 生态学报, 2019, 39(3): 1095-1104. |
| [23] |
付翔. 个性对中华倒刺鲃集群行为及生长性能的影响[D]. 重庆: 重庆师范大学, 2021: 1-40.
|
| [24] |
郝雅宾, 刘金殿, 郭爱环, 张爱菊, 周志明. PIT标记在水生动物研究中的应用研究进展. 海洋渔业, 2019, 41(2): 242-249. |
| [25] |
孙春艳, 曾令清. 颜色标记对两种鱼类有氧运动能力的影响. 重庆师范大学学报: 自然科学版, 2020, 37(3): 67-71. |
| [26] |
Tang J Y, Fu S J. The relationship between personality and the collective motion of schooling fish. Journal of Ethology, 2020, 38(3): 333-341. DOI:10.1007/s10164-020-00655-1 |
| [27] |
Herbert-Read J E, Krause S, Morrell L J, Schaerf T M, Krause J, Ward A J W. The role of individuality in collective group movement. Proceedings of the Royal Society B: Biological Sciences, 2013, 280(1752): 20122564. DOI:10.1098/rspb.2012.2564 |
| [28] |
Villegas-Ríos D, Réale D, Freitas C, Moland E, Olsen E M. Individual level consistency and correlations of fish spatial behaviour assessed from aquatic animal telemetry. Animal Behaviour, 2017, 124: 83-94. DOI:10.1016/j.anbehav.2016.12.002 |
| [29] |
曾令清, 付世建, 曹振东. 鲫幼鱼(Carassius auratus)标准代谢个体差异与力竭后代谢特征及行为的关联. 生态学报, 2016, 36(6): 1746-1755. |
| [30] |
唐中华. 捕食压力对乌江流域不同宽鳍鱲种群的标准代谢、个性和集群的影响[D]. 重庆: 重庆师范大学, 2018: 1-48.
|
| [31] |
Wolf M, Weissing F J. Animal personalities: consequences for ecology and evolution. Trends in Ecology & Evolution, 2012, 27(8): 452-461. |
| [32] |
Réale D, Garant D, Humphries M M, Bergeron P, Careau V, Montiglio P O. Personality and the emergence of the pace-of-life syndrome concept at the population level. Philosophical Transactions of the Royal Society B: Biological Sciences, 2010, 365(1560): 4051-4063. DOI:10.1098/rstb.2010.0208 |
| [33] |
吴慧, 唐中华, 黄清, 况璐, 付世建. 群体成员大小差异对不同生境鲤科鱼类集群行为的影响. 水生生物学报, 2017, 41(3): 552-559. |
| [34] |
Spiegel O, Leu S T, Bull C M, Sih A. What's your move? Movement as a link between personality and spatial dynamics in animal populations. Ecology Letters, 2017, 20(1): 3-18. DOI:10.1111/ele.12708 |
| [35] |
Villegas-Ríos D, Réale D, Freitas C, Moland E, Olsen E M. Personalities influence spatial responses to environmental fluctuations in wild fish. Journal of Animal Ecology, 2018, 87(5): 1309-1319. DOI:10.1111/1365-2656.12872 |
| [36] |
Fu S J. Effects of group size on schooling behavior in two cyprinid fish species. Aquatic Biology, 2016, 25: 165-172. DOI:10.3354/ab00667 |
| [37] |
Dyer J R G, Croft D P, Morrell L J, Krause J. Shoal composition determines foraging success in the guppy. Behavioral Ecology, 2009, 20(1): 165-171. DOI:10.1093/beheco/arn129 |
| [38] |
Ward A, Webster M. Development, ontogeny and parasite-mediated changes in social behaviour//Sociality: the Behaviour of Group-Living Animals. Berlin, Germany: Springer, 2016.
|
| [39] |
Hein A M, Gil M A, Twomey C R, Couzin I D, Levin S A. Conserved behavioral circuits govern high-speed decision-making in wild fish shoals. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(48): 12224-12228. DOI:10.1073/pnas.1809140115 |
| [40] |
Cote J, Fogarty S, Brodin T, Weinersmith K, Sih A. Personality-dependent dispersal in the invasive mosquitofish: group composition matters. Proceedings of the Royal Society B: Biological Sciences, 2011, 278(1712): 1670-1678. DOI:10.1098/rspb.2010.1892 |
| [41] |
Planas-Sitjà I, Nicolis S C, Sempo G, Deneubourg J L. The interplay between personalities and social interactions affects the cohesion of the group and the speed of aggregation. PLoS One, 2018, 13(8): e0201053. DOI:10.1371/journal.pone.0201053 |
| [42] |
Bumann D, Krause J. Front individuals lead in shoals of three-spined sticklebacks (Gasterosteus aculeatus) and juvenile roach (Rutilus rutilus). Behaviour, 1993, 125(3/4): 189-198. |
| [43] |
Mcdonald N D, Rands S A, Hill F, Elder C, Ioannou C C. Consensus and experience trump leadership, suppressing individual personality during social foraging. Science Advances, 2016, 2(9): e1600892. |
| [44] |
柏杨, 覃英莲, 彭姜岚, 付世建. 群体大小对麦穗鱼幼鱼凝聚力和协调性的影响. 重庆师范大学学报: 自然科学版, 2016, 33(4): 22-26. |
| [45] |
Herbert-Read J E, Perna A, Mann R P, Schaerf T M, Sumpter D J T, Ward A J W. Inferring the rules of interaction of shoaling fish. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(46): 18726-18731. DOI:10.1073/pnas.1109355108 |