 2022, Vol. 42
2022, Vol. 42文章信息
- 窦飞越, 张欢, 吴智宏, 林木青, 束祖飞, 李红英, 寇若玫, 周泽扬, 黄敦元, 肖治术
- DOU Feiyue, ZHANG Huan, WU Zhihong, LIN Muqing, SHU Zufei, LI Hongying, KOU Ruomei, ZHOU Zeyang, HUANG Dunyuan, XIAO Zhishu
- 海拔梯度变化对黄缘蜾蠃筑巢结构及相关生物学特性的影响
- Effects of altitude gradient changes on nesting and biology for the potter wasp Anterhynchium flavomarginatum (Hymenoptera: Vespidae: Eumeninae)
- 生态学报. 2022, 42(8): 3279-3287
- Acta Ecologica Sinica. 2022, 42(8): 3279-3287
- http://dx.doi.org/10.5846/stxb202012303317
-
文章历史
- 收稿日期: 2020-12-30
- 网络出版日期: 2021-12-17





2. 广东车八岭国家级自然保护区管理局, 韶关 512500;
3. 中国科学院动物研究所, 农业虫害鼠害综合治理研究国家重点实验室, 北京 100101;
4. 中国科学院大学, 北京 100049
2. Guangdong Chebaling National Nature Reserve Administration Bureau, Shaoguan 512500, China;
3. State Key Laboratory of Integrated Management of Pest Insects and Rodents, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, China;
4. University of Chinese Academy of Sciences, Beijing 100049, China
地上筑巢的野生独栖蜂类(包括蜜蜂和胡蜂类群)喜欢选择天然存在的空穴材料如石缝、植物茎秆、枯木中的虫洞以及蜗牛壳等场地进行筑巢[1-2]。研究发现适宜的栖息地环境、充足的食物资源以及适合的筑巢材料是这类昆虫繁殖的主要限制因素[3-4]。因此, 对独栖蜂类而言, 在资源有限的自然环境下, 如何最佳利用筑巢资源以提高种群对环境的适应性是非常重要的。研究表明, 在具有雌雄体型性二型现象的物种中, 亲本可以根据现有的筑巢资源丰度调整对子代的投资及性别分配比例[5-6]。例如, 在Osmia rufa筑巢期中, 由于雌性对后代提供的食物效率下降, 导致其后代体型的减少和后代性比雄性偏多[7]。然而到目前为止, 这类研究通常关注的更多的是水平带上的差异[8-9], 对于同一个地区不同海拔梯度下的独栖蜂类是如何调整其筑巢策略以适应环境变化, 还需要进一步研究[10]。
山地生态类环境系统是地球表面具有显著起伏和坡度的三维地貌体[11]。海拔梯度作为山地生境差异的主导因子, 其现象表现为温度、降水、风速、光照及土壤等自然因子随海拔变化而出现规律性的梯度变化, 进而影响物种的多样性及组成和分布[12-13]。目前的研究表明, 不同类群或不同区域下的物种多样性随海拔梯度的分布格局各异[14], 但大致呈现4种格局:即单调递减格局(decreasing patterns)、低平台高原格局(low-elevation plateaus, LP)、偏峰格局(low-elevation plateaus with a mid-elevation peak, LPMP)和中峰格局(mid-elevation peak, MP), 4种格局分别占25%、15%、14%、45%[15]。除此之外, 研究物种的多样性分布格局及组成结构随海拔梯度的变化还有助于理解全球气候变化对物种的影响, 同时为生物多样性的保护提供理论依据[16-20]。在昆虫中, 捕食性和寄生性昆虫在调节猎物和宿主的种群动态中起着至关重要的作用[21], 其中野生蜂类是研究沿海拔梯度变化的模式生物之一[22]。
蜾蠃亚科(Eumeninae)隶属于膜翅目(Hymenoptera)细腰亚目(Apocrita)针尾部(Aculeata)胡蜂科(Vespidae)[23-24], 全世界共有205属3773种, 中国纪录51属267种[25]。研究表明, 蜾蠃亚科多为独栖类群, 喜营自由生活, 雌蜂交配后开始筑巢狩猎并产卵繁殖, 其巢穴类型主要有两种:一种是用泥土修建成中空壶状的巢穴, 另一种是利用天然空穴修建的巢穴[26-27]。该亚科的狩猎方式是雌蜂用蜇针蜇刺鳞翅目幼虫, 使猎物麻痹, 再带回巢穴中储备以供后代取食。研究发现, 黄唇蜾蠃(Rhynchium bruneum)的一个巢穴内多达45条鳞翅目害虫, 因此蜾蠃类群对农林害虫防治能起到一定控制作用[28-29]。此外, 蜾蠃类群的蜂毒因具有麻痹和贮存活体寄主的功能, 所以在蜂毒成分研究和新型麻醉剂的开发应用方面也具有一定的意义[30-32]。
黄缘蜾蠃(Anterhynchium flavomarginatum)隶属于蜾蠃亚科(Eumeninae)缘蜾蠃属(Anterhynchium)[25], 主要分布在我国浙江、上海、江西、湖南、四川、福建、广西、云南等地[33-35]。研究表明, 该蜂自然条件下选择甲虫废弃的地道、树木、竹子以及芦苇等材料中筑巢, 主要以鳞翅目幼虫为食, 是一种重要的捕食性天敌昆虫[9]。
人工巢穴技术(trap-nesting)是利用芦苇、纸筒、竹子等材料制成一端开口一端封闭的巢管, 或直接在木头上钻蛀不同大小的巢孔并放置在野外用以收集独栖蜂类的一种诱集装置[36-39]。1967年, Karl首次利用巢管收集独栖性蜂类[40], 随着该技术的推广和完善, 目前人工巢穴技术常和网扫、黄盘诱集、马来氏网、飞行阻截等采集方法一起对研究区域进行标本采集并作为昆虫多样性分析的依据[41-42], 也广泛用于研究独栖蜂类后代的性比及性别分配机制[5, 9, 43], 还用于传粉昆虫和捕食性天敌的放养等[44-46]。本研究利用芦苇制作的人工巢管, 在广东车八岭国家级自然保护区中诱集不同海拔梯度下的黄缘蜾蠃, 研究海拔梯度对该蜂筑巢策略的影响, 旨在进一步探讨黄缘蜾蠃等独栖性蜂类对不同海拔梯度下特定生境的响应。
1 材料与方法 1.1 研究地概况广东车八岭国家级自然保护区位于中国广东省始兴县东南部, 北纬24°40′29″-24°46′21″, 东经114°07′39″-114°16′46″, 总面积75.45 km2。该区地势西北高东南低, 最低处樟栋水海拔330 m, 最高峰天平架海拔1256 m。保护区年均温度19.6 ℃, 最高温38.4 ℃, 年降水量1, 150-2, 126 mm。保护区植物区系属于南亚热带向中亚热带的过度地带, 是南岭山脉南缘具有代表性的中亚热带常绿阔叶林[47]。
1.2 样地设置本研究自2017年底在广东车八岭国家级自然保护区布设人工巢箱对区内野生蜂类多样性进行监测。人工巢箱布设方案:以整个保护区全境及周边为单位制作1 km × 1 km网格地图, 共设置100个调查网格, 每个调查网格布设3个人工巢箱(其中1个在林内, 2个在林缘), 每个巢箱有2个PVC管组成(PVC规格:长25 cm, 内径110 cm), 每个PVC管内放有约70根巢管(巢管规格:长度在8-20 cm之间, 内径在0.5-1.7 cm之间), 共300个巢箱(图 1)。使用GPS记录所有巢箱的地理信息。野外调查时间从2018年5月初至2019年5月初, 每隔1-2个月收集1次筑巢巢管并带回室内饲养观察, 同时在对应巢箱中补充相同数目的空巢管;室内饲养观察持续到2019年12月。

|
| 图 1 广东车八岭国家级自然保护区巢管位点分布及安装示意图 Fig. 1 Site distribution and installation of trap nests in the Chebaling National Nature Reserve, Guangdong Province 黑色的点代表巢管安装的位点分布 |
室内对黄缘蜾蠃的筑巢巢管进行编号, 用解剖刀把巢管剖开约1/3, 使用电子游标卡尺测量巢管的内部结构大小, 包括:巢管内径、虫室长度、间室长度和前庭长度(相邻隔板间的距离), 并统计每根巢管中的虫室数量和间室数量(图 2), 最后将解剖后的巢管放入无菌玻璃试管内并用药棉堵塞管口, 以便后续观测记录。羽化成虫及其寄生类群用5 mL冻虫管独立分装, 酒精浸泡后放入冰柜中冷冻保存以便后续相关研究。使用电子游标卡尺测量两翅膀基部间的距离(胸宽)。

|
| 图 2 黄缘蜾蠃的巢管结构 Fig. 2 Nest structure of A. flavomarginatum |
广义加合模型(generalized additive model, GAM)的特征是不需要预先设定自变量和因变量之间的具体函数关系, 其可以通过自变量和因变量之间的光滑函数来构建数学关系, 所以能最大程度符合原始数据的规律[48]。该模型的数学表达式如下[49]:
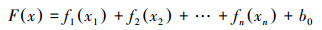
|
式中, Fx为链接函数, fn为解释变量关系的非参数函数, b0为常数截距项。
本研究中, 运用GAM模型分别对巢内径、巢室长度、巢室数量、雌雄数量及雌虫胸宽进行回归分析。数据经过Excel 2016软件统计后, 利用R 3.6.2软件中“mgcv”包中的“gam”函数对数据进行GAM模型分析[50], 通过“plot”函数实现所有的图形可视化[51], 分析结果通过P值和调整判定系数(R2)来表征。其中, P值代表统计结果的显著性水平, 本文的显著性水平取P < 0.05, R2用于判定回归方程的拟合效果, 值域为[0, 1], 值越大拟合效果越好。
2 结果与分析 2.1 海拔梯度对巢管大小选择的影响对野外回收167根黄缘蜾蠃筑巢巢管的内径和长度进行测量发现, 雌虫偏好选择的巢管内径范围在5.63-19.13 mm之间, 平均长度为(11.98 ± 2.20)mm(n=167)。对不同海拔梯度下该蜂的筑巢巢管内径数据进行回归分析发现, 随着海拔梯度增加, 巢管内径呈现先增后减的趋势(R2 =0.28, P < 0.01)(图 3)。

|
| 图 3 基于GAM模型的海拔与巢管内径最优拟合结果 Fig. 3 Results of GAM analysis on altitude and nest inner diameter 实点表示观测的巢管内径, 实线为模型预测的拟合曲线, 虚线表示模型预测的95%置信区间, 纵坐标为平滑函数的值 |
通过室内对黄缘蜾蠃筑巢巢管的相关参数进行测量发现:该蜂筑巢巢管的虫室、间室、前庭的长度范围分别在8.68-45.65 mm、4.63-39.26 mm、3.75-49.98 mm, 平均长度分别为(24.64 ± 7.51)mm(n=302)、(16.21 ± 7.30)mm(n =152)、(19.60 ± 10.39)mm(n=122)。回归结果显示, 虫室、间室和前庭与海拔之间具有相关性, 但解释程度较低, 为弱相关, 模型显示虫室、间室和前庭的长度随着海拔增加总体上都呈现先减后增的趋势(R2=0.14, P < 0.01;R2 =0.16, P < 0.01;R2=0.46, P < 0.01)(图 4)。

|
| 图 4 基于GAM模型的海拔与巢室长度(虫室长度, 间室长度, 前庭长度)最优拟合结果 Fig. 4 Results of GAM analysis on altitude and nest length (brood cell length; intercalary cell length; vestibular cell length) 实点表示观测的巢室长度, 实线为模型预测的拟合曲线, 虚线表示模型预测的95%置信区间, 纵坐标为平滑函数的值 |
对黄缘蜾蠃所有筑巢巢管内的巢室数量统计发现:每个巢管中的虫室数量在1-6之间, 平均为(2.39 ± 1.06)(n=167);间室数量在1-4之间, 平均值为(1.66 ± 0.71)(n=167)。对该蜂不同海拔梯度下的筑巢巢管的虫室数目和间室数目进行回归分析发现, 随着海拔梯度增加, 虫室数和间室数均表现为先增后减的趋势, 但解释程度较低, 为弱相关(R2=0.10, P < 0.001;R2=0.15, P < 0.01)(图 5)。

|
| 图 5 基于GAM模型的海拔与巢室数量(虫室数量, 间室数量)最优拟合结果 Fig. 5 Results of GAM analysis on altitude and nest number (brood cell number; intercalary cell length) 实点表示观测的巢室数量, 实线为模型预测的拟合曲线, 虚线表示模型预测的95%置信区间, 纵坐标为平滑函数的值 |
2018年5月初至2019年5月初期间, 只有46%的羽化蜂被成功收集(在巢管解剖前就有蜂羽化出巢, 或者在室内孵育过程因寄生生物、病虫害造成死亡, 以及未知原因造成的死亡), 共173只, 其中雌性个体55只, 雄性个体118只, 雌雄性比为1∶2.1。对该蜂不同海拔梯度下的雌雄数量进行回归分析发现, 随着海拔梯度增加, 单根巢管内的雄性数量呈现多波峰的非线性关系(R2 =0.14, P=0.02), 而单根巢管内的雌性数量呈先增后减的趋势(R2 =0.28, P < 0.01), 但对单根巢管内的雄性数量解释程度较低, 为弱相关(图 6)。

|
| 图 6 基于GAM模型的海拔与子代雌雄数量(雄性数量, 雌性数量)最优拟合结果 Fig. 6 Results of GAM analysis on altitude and number of male and female offspring (male number; female number) 实点表示观测的子代雌雄数量, 实线为模型预测的拟合曲线, 虚线表示模型预测的95%置信区间, 纵坐标为平滑函数的值 |
对羽化出巢后的雌性黄缘蜾蠃的胸宽进行测量发现:该蜂雌性成虫的胸宽在4.50-6.16 mm, 平均值为(5.23 ± 0.35)mm(n=55)。对该蜂不同海拔梯度下雌性成虫的胸宽大小进行回归分析发现:随着海拔梯度增加, 雌性成虫胸宽总体呈先增后减的趋势, 在海拔400-700 m之间逐渐上升, 在约700 m之后逐渐下降(R2=0.31, P < 0.01)(图 7)。

|
| 图 7 基于GAM模型的海拔与子代雌性成虫胸宽最优拟合结果 Fig. 7 Results of GAM analysis on altitude and female breast wide of offspring 实点表示观测的子代雌性成虫胸宽, 实线为模型预测的拟合曲线, 虚线表示模型预测的95%置信区间, 纵坐标为平滑函数的值 |
独栖蜂类依赖于现存的孔洞筑巢, 一旦雌蜂筑好巢穴, 其后代从卵、幼虫到蛹会一直在巢穴中生活[37], 因此雌蜂对巢穴大小的选择就显得非常重要。目前已有研究表明巢穴尺寸会影响到这类昆虫后代的性别比偏向, 具体而言, 在内径较小的巢穴中, 雄性比例会上升, 而在内径较大的巢穴中, 雌性比例会上升[52]。同样地, 郭鹏飞等人在研究黄缘蜾蠃后代性别的分配时也发现, 随着巢管直径的增加, 雌性比例增加的现象[9]。为了进一步探究这种现象背后的原因, 本研究将海拔梯度作为解释变量, 实验结果发现了, 随着海拔的增加, 黄缘蜾蠃选择的巢管内径尺寸先增后减, 而后代中雌性数量也出现了先增后减的趋势。尽管研究还发现了后代中雄性数量有着与雌性数量变化相反的趋势, 但是模型显示了雄性数量与海拔之间相关性较弱。因此, 本研究认为海拔梯度主要影响了黄缘蜾蠃对巢管内径的选择以及对后代中雌性的投资比例。根据以往的研究结果显示, 巢穴附近的食物丰富度会影响独栖蜂类对后代性别比的分配。Seidelmann等人认为, 在巢管空间不限制下, 亲本将调整对后代的投资至上限, 这种调整将有利于体型较大的雌性个体[1, 53], 而巢穴的内径大小又决定了每个虫室中可投资的食物上限[54]。因此, 在食物可观的情况下, 亲本会选择较大的巢管来繁殖更多的雌性后代。吴艳光在调查长白山地区的昆虫多样性中发现, 在不同海拔高度的垂直带中, 针阔混交林群落相比其它群落具有更高的昆虫物种数和个体数[55]。梁召俊在调查川西高原森林昆虫种类多样性发现, 鳞翅目昆虫在人工林的数量总体少于原始林[56]。而对于黄缘蜾蠃, 该蜂是一种以鳞翅目幼虫为食的独栖性捕食昆虫, 据研究显示, 车八岭植被垂直类型为:从低海拔以常绿阔叶林、针叶林和人工林为主(330-500 m), 中海拔以常绿阔叶林和针叶林为主(500-800 m), 高海拔以灌丛草坡为主(800-1256 m)[57]。因此在该片区, 中海拔的常绿阔叶林和针叶林具有的昆虫多样性更高, 黄缘蜾蠃可以获得更多的食物资源, 从而促使其选择更大内径的巢穴, 以繁殖更多的雌性后代。
独栖蜂的亲本对后代体型有高度控制权, 食物的摄入量与独栖性后代体型大小密切相关[6]。Alonso等人在调查一种泥蜂时发现, 体重更大的泥蜂能捕捉猎物体重的范围更大[52]。此外, 对于这类独栖昆虫而言, 雌性通常扮演者筑巢、捕食和繁殖的角色, 而雄性通常只负责求偶交配。显然, 雌性在种群中具有更多的功能属性。在本研究, 黄缘蜾蠃后代中雌蜂的数量和胸宽都随海拔增加先增后减, 而雄性数量与海拔呈弱相关性。这可能就是因为, 雌性在种群中的重要地位决定的。而雌蜂的这种重要性, 促使亲本在食物更多的中海拔区对雌性子代具有更多的投资, 结果表现为更大体型和更多数量的雌性后代, 以提高种群生存和繁殖的竞争力。相反, 对于雄性的投资就显得不太重要。
独栖蜂类的巢管结构主要包括虫室(brood cells/provisioned cells)、间室(intercalary cell)及前庭(vestibular cell)3个部分[30]。Polidori等人[5]认为, 野生蜂类亲本除了考虑选择适宜的巢管大小外, 还要考虑对筑巢空间的分配, 即雌蜂需要根据现有环境权衡后代大小和数量间的关系, Smith和Fretwell研究发现, 亲本对子代数量上的投资大于后代个体大小的投资, 因为子代数量对整个种群稳定的维持更加显著[58]。而在海拔梯度上, 本研究结果发现, 尽管随着海拔增加, 虫室长度、间室长度、虫室数和间室数分别有对应的变化趋势, 但模型的解释程度较低, 这些因子与海拔之间呈弱相关。相反,前庭长度表现出较高相关性。在Asís等人的研究中发现,较长的前庭不仅能够减少子代在发育最初阶段的死亡率,还能够减少巢穴外剩余空间大小,以防止其它雌性前来筑巢[59]。因此, 结果表明了海拔在筑巢空间上主要体现在对前庭长度的影响。
对独栖蜂类而言, 在资源有限的自然环境下, 如何最佳利用筑巢资源以提高种群对环境的适应性是非常重要的。本研究显示, 海拔主要影响的是黄缘蜾蠃对巢管内径大小的选择, 而不是对巢室长度和数量的分配, 同时, 黄缘蜾蠃对巢管内径的选择可以影响到后代性别比, 这种策略主要体现在亲本对雌性子代的投资, 而不是雄性子代。由此可见黄缘蜾蠃可通过调整其筑巢策略以应对不同海拔下的环境, 但对于其它独栖蜂类而言, 是否也具有一致的调整策略尚未清楚, 仍需进一步研究。
| [1] |
Seidelmann K, Bienasch A, Pröhl F. The impact of nest tube dimensions on reproduction parameters in a cavity nesting solitary bee, Osmia bicornis (Hymenoptera: Megachilidae). Apidologie, 2016, 47(1): 114-122. DOI:10.1007/s13592-015-0380-z |
| [2] |
da Silva Mello B N, Gaglianone M C. Nesting biology of sympatric species of megachilidae bees in a conservation area in Brazilian Atlantic Forest. Sociobiology, 2019, 66(1): 52-60. DOI:10.13102/sociobiology.v66i1.2881 |
| [3] |
Steffan-Dewenter I, Leschke K. Effects of habitat management on vegetation and above-ground nesting bees and wasps of orchard meadows in Central Europe. Biodiversity & Conservation, 2003, 12(9): 1953-1968. |
| [4] |
Morato E F, Martins R P. An overview of proximate factors affecting the nesting behavior of solitary wasps and bees (Hymenoptera: Aculeata) in preexisting cavities in wood. Neotropical Entomology, 2006, 35(3): 285-298. DOI:10.1590/S1519-566X2006000300001 |
| [5] |
Polidori C, Boesi R, Borsato W. Few, small, and male: multiple effects of reduced nest space on the offspring of the solitary wasp, Euodynerus (Pareuodynerus) posticus (Hymenoptera: Vespidae). Comptes Rendus Biologies, 2011, 334(1): 50-60. DOI:10.1016/j.crvi.2010.11.003 |
| [6] |
王丽娜, 闫卓, 欧阳芳, 门兴元, 叶保华. 巢管内径尺寸对凹唇壁蜂(膜翅目: 切叶蜂科)种群参数的影响. 应用昆虫学报, 2018, 55(6): 1001-1006. |
| [7] |
Seidelmann K. Open-cell parasitism shapes maternal investment patterns in the Red Mason bee Osmia rufa. Behavioral Ecology, 2006, 17(5): 839-848. DOI:10.1093/beheco/arl017 |
| [8] |
Herrmann J D, Beye H, de la Broise C, Hartlep H, Diekötter T. Positive effects of the pollinators Osmia cornuta (Megachilidae) and Lucilia sericata (Calliphoridae) on strawberry quality. Arthropod-Plant Interactions, 2019, 13(1): 71-77. DOI:10.1007/s11829-018-9636-7 |
| [9] |
郭鹏飞, 陶玫, 袁峰, 朱朝东. 探究黄缘蜾蠃后代性别的分配. 环境昆虫学报, 2018, 40(3): 645-650. |
| [10] |
Hoiss B, Krauss J, Potts S G, Roberts S, Steffan-Dewenter I. Altitude acts as an environmental filter on phylogenetic composition, traits and diversity in bee communities. Proceedings of the Royal Society B: Biological Sciences, 2012, 279(1746): 4447-4456. DOI:10.1098/rspb.2012.1581 |
| [11] |
郑智, 龚大洁, 孙呈祥. 白水江自然保护区两栖爬行动物物种丰富度和种域海拔梯度格局及对Rapoport法则的验证. 生态学杂志, 2014, 33(2): 537-546. |
| [12] |
岳明, 张林静, 党高弟, 辜天琪. 佛坪自然保护区植物群落物种多样性与海拔梯度的关系. 地理科学, 2002, 22(3): 349-354. DOI:10.3969/j.issn.1000-0690.2002.03.016 |
| [13] |
向琳, 陈芳清, 官守鹏, 王玉兵, 吕坤. 井冈山鹿角杜鹃群落灌木层功能多样性及其随海拔梯度的变化. 生态学报, 2019, 39(21): 8144-8155. |
| [14] |
陶夏秋, 崔绍朋, 蒋志刚, 初红军, 李娜, 杨道德. 李春旺新疆阿勒泰地区爬行动物区系及多样性海拔分布格局. 生物多样性, 2018, 26(6): 578-589. |
| [15] |
刘开明, 郑智, 龚大洁. 物种丰富度的垂直分布格局及其形成机制. 生态学杂志, 2017, 36(2): 541-554. |
| [16] |
Xu M H, Zhang S X, Wen J, Yang X Y. Multiscale spatial patterns of species diversity and biomass together with their correlations along geographical gradients in subalpine meadows. PLoS One, 2019, 14(2)e0211560. |
| [17] |
Streinzer M, Chakravorty J, Neumayer J, Megu K, Narah J, Schmitt T, Bharti H, Spaethe J, Brockmann A. Species composition and elevational distribution of bumble bees (Hymenoptera, Apidae, Bombus Latreille) in the East Himalaya, Arunachal Pradesh, India. Zookeys, 2019, 851: 71-89. DOI:10.3897/zookeys.851.32956 |
| [18] |
Benzina I, Si Bachir A S, Ghazi C, Santoul F, Céréghino R. How altitudinal gradient affects the diversity and composition of benthic insects in arid areas streams of northern East Algeria?. Biologia, 2020, 75(4): 567-577. DOI:10.2478/s11756-019-00326-8 |
| [19] |
彭正兵, 李新建, 张瑞波, 秦莉, 张合理, 陈友平, 刘蕊. 不同去趋势方法的新疆东天山高低海拔雪岭云杉树轮宽度年表对气候的响应. 生态学报, 2019, 39(5): 1595-1604. |
| [20] |
曹仁杰, 尹定财, 田昆, 肖德荣, 李志军, 张绪岗, 李泽辉, 张贇. 丽江老君山海拔上限长苞冷杉(Abies georgei)和云南铁杉(Tsuga dumosa)径向生长对气候变化的响应. 生态学报, 2020, 40(17): 6067-6076. |
| [21] |
Perillo L N, de Siqueira Neves F, Antonini Y, Martins R P. Compositional changes in bee and wasp communities along Neotropical mountain altitudinal gradient. PLoS One, 2017, 12(7): e0182054. DOI:10.1371/journal.pone.0182054 |
| [22] |
Corcos D, Cerretti P, Mei M, Taglianti A V, Paniccia D, Santoiemma G, De Biase A, Marini L. Predator and parasitoid insects along elevational gradients: role of temperature and habitat diversity. Oecologia, 2018, 188(1): 193-202. DOI:10.1007/s00442-018-4169-4 |
| [23] |
Carpenter J M. Phylogenetic relationships and classification of the Vespinae (Hymenoptera: Vespidae). Systematic Entomology, 1987, 12(4): 413-431. DOI:10.1111/j.1365-3113.1987.tb00213.x |
| [24] |
Carpenter J M. A synonymic generic checklist of the Eumeninae (Hymenoptera: Vespidae). Psyche: A Journal of Entomology, 1986, 93: 012489. |
| [25] |
Tan J L, Carpenter J M, van Achterberg C. An illustrated key to the genera of Eumeninae from China, with a checklist of species (Hymenoptera, Vespidae). Zookeys, 2018, 740: 109-149. DOI:10.3897/zookeys.740.22654 |
| [26] |
黄敦元, 何波, 谷平, 余江帆, 彭飞, 许晓风. 日本佳盾蜾蠃的生物学特性观察. 环境昆虫学报, 2014, 36(6): 890-897. |
| [27] |
黄敦元, 何波, 余江帆, 谷平, 彭飞, 黄名广, 朱朝东. 日本佳盾蜾蠃营巢生物学研究. 环境昆虫学报, 2013, 35(6): 783-791. |
| [28] |
周鑫, 李廷景, 陈斌. 蜾蠃亚科(膜翅目: 胡蜂科)分类研究进展. 重庆师范大学学报: 自然科学版, 2011, 28(6): 22-29. |
| [29] |
李参, 王铁伟, 董秀仁. 黄唇蜾蠃蜂及其利用的初步研究. 昆虫学报, 1975, 18(2): 151-155. |
| [30] |
许俊峰, 韩召军. 黄唇蜾蠃蜂蜂毒的毒性特征研究. 南京农业大学学报, 2009, 32(4): 89-93. |
| [31] |
Delatorre P, Olivieri J R, Neto J R, Lorenzi C C B, Canduri F, Fadel V, Konno K, Palma M S, Yamane T, de Azevedo Jr W F. Preliminary cryocrystallography analysis of an Eumenine mastoparan toxin isolated from the venom of the wasp Anterhynchium flavomarginatum micado. Biochimica et Biophysica Acta (BBA)-Protein Structure and Molecular Enzymology, 2001, 1545(1/2): 372-376. |
| [32] |
Konno K, Hisada M, Naoki H, Itagaki Y, Kawai N, Miwa A, Yasuhara T, Morimoto Y, Nakata Y. Structure and biological activities of eumenine mastoparan-AF (EMP-AF), a new mast cell degranulating peptide in the venom of the solitary wasp (Anterhynchium flavomarginatum micado). Toxicon, 2000, 38(11): 1505-1515. DOI:10.1016/S0041-0101(00)00083-0 |
| [33] |
李铁生, 潘秀敏, 关国. 中国经济昆虫志: 第三十册, 膜翅目: 胡蜂总科. 北京: 科学出版社, 1985, 133-134. |
| [34] |
何俊华. 浙江蜂类志[M]. 科学出版社, 2004: 896.
|
| [35] |
董大志, 王云珍. 云南胡蜂志: 膜翅目: 胡蜂总科. 郑州: 河南科学技术出版社, 2017, 175-176. |
| [36] |
da Costa C C F, Gonçalves R B. What do we know about Neotropical trap-nesting bees? Synopsis about their nest biology and taxonomy. Papéis Avulsos de Zoologia, 2019, 59: e20195926. DOI:10.11606/1807-0205/2019.59.26 |
| [37] |
Staab M, Pufal G, Tscharntke T, Klein A M. Trap nests for bees and wasps to analyse trophic interactions in changing environments-a systematic overview and user guide. Methods in Ecology and Evolution, 2018, 9(11): 2226-2239. DOI:10.1111/2041-210X.13070 |
| [38] |
黄敦元, 朱朝东, 肖忠优, 黄名广, 胡冬南. 人工巢穴技术在昆虫学研究中的应用. 安徽农业科学, 2012, 40(10): 5974-5975, 5977. DOI:10.3969/j.issn.0517-6611.2012.10.093 |
| [39] |
何波, 谷战英, 李红英, 黄敦元. 基于DNA条形码的白斑切叶蜂粉源植物种类及多样性分析. 生态学报, 2020, 40(6): 2122-2129. |
| [40] |
Roberts R B. Trap-nesting wasps and bees: life histories, nests, and associates. Systematic Biology, 1967, 16(3): 261-262. |
| [41] |
Sobek S, Tscharntke T, Scherber C, Schiele S, Steffan-Dewenter I. Canopy vs. understory: does tree diversity affect bee and wasp communities and their natural enemies across forest strata?. Forest Ecology and Management, 2009, 258(5): 609-615. DOI:10.1016/j.foreco.2009.04.026 |
| [42] |
Müller A, Praz C, Dorchin A. Biology of Palaearctic Wainia bees of the subgenus Caposmia including a short review on snail shell nesting in osmiine bees (Hymenoptera, Megachilidae). Journal of Hymenoptera Research, 2018, 65: 61-89. DOI:10.3897/jhr.65.27704 |
| [43] |
Mikát M, Benda D, Straka J. Maternal investment in a bee species with facultative nest guarding and males heavier than females. Ecological Entomology, 2019, 44(6): 823-832. DOI:10.1111/een.12768 |
| [44] |
Boyle N K, Artz D R, Lundin O, Ward K, Picklum D, Wardell G I, Williams N M, Pitts-Singer T L. Wildflower plantings promote blue orchard bee, Osmia lignaria (Hymenoptera: Megachilidae), reproduction in California almond orchards. Ecology and Evolution, 2020, 10(7): 3189-3199. DOI:10.1002/ece3.5952 |
| [45] |
魏永平, 袁锋. 果树授粉蜂--凹唇壁蜂人工应用技术. 陕西农业科学, 1997, 43(5): 43-44. |
| [46] |
杨龙龙, 徐环李, 吴燕如. 凹唇壁蜂和紫壁蜂筑巢、访花行为和传粉生态学的比较研究. 生态学报, 1997, 17(1): 1-6. |
| [47] |
徐燕千. 车八岭国家级自然保护区调查研究综合报告//车八岭国家级自然保护区调查研究论文集. 广州: 广东科技出版社, 1993: 1-8.
|
| [48] |
温庆志, 姚蕊, 孙鹏, 张强, 顾然. 变异条件下淮河流域生态径流变化特征及驱动因子. 生态学报, 2020, 40(8): 2621-2635. |
| [49] |
Chen Q X, Ibrahim J G. Semiparametric models for missing covariate and response data in regression models. Biometrics, 2006, 62(1): 177-184. DOI:10.1111/j.1541-0420.2005.00438.x |
| [50] |
Wood S N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. Journal of the Royal Statistical Society Statistical Methodology Series B, 2011, 73(1): 3-36. DOI:10.1111/j.1467-9868.2010.00749.x |
| [51] |
卡巴科弗. R语言实战. 王小宁, 刘撷芯, 黄俊文, 译. 2版. 北京: 人民邮电出版社, 2016: 110-130.
|
| [52] |
Alonso J D S, Silva J F, Garófalo C A. The effects of cavity length on nest size, sex ratio and mortality of Centris (Heterocentris) analis (Hymenoptera, Apidae, Centridini). Apidologie, 2012, 43(4): 436-448. DOI:10.1007/s13592-011-0110-0 |
| [53] |
Seidelmann K. Optimal progeny body size in a solitary bee, Osmia bicornis (Apoidea: Megachilidae). Ecological Entomology, 2014, 39(5): 656-663. DOI:10.1111/een.12145 |
| [54] |
Bosch J. Production of undersized offspring in a solitary bee. Animal Behaviour, 2008, 75(3): 809-816. DOI:10.1016/j.anbehav.2007.06.018 |
| [55] |
吴艳光. 长白山地区访花昆虫多样性及访花行为的研究[D]. 长春: 东北师范大学, 2006.
|
| [56] |
梁召俊. 川西高原森林昆虫种类多样性垂直分布研究[D]. 杨凌: 西北农林科技大学, 2017.
|
| [57] |
陈涛, 缪绅裕, 廖文波, 苏志尧, 唐绍清, 张宏达. 广东车八岭自然保护区植物区系地理研究. 生态科学, 1992(1): 1-28. |
| [58] |
Smith C C, Fretwell S D. The optimal balance between size and number of offspring. The American Naturalist, 1974, 108(962): 499-506. DOI:10.1086/282929 |
| [59] |
Asís J D, Benéitez A, Tormos J, Gayubo S F, Tomé M A M. The Significance of the Vestibular Cell in Trap Nesting Wasps (Hymenoptera: Crabronidae): Does its Presence Reduce Mortality?. Journal of Insect Behavior, 2007, 20: 289-305. DOI:10.1007/s10905-007-9080-5 |