 2022, Vol. 42
2022, Vol. 42文章信息
- 张乐满, 兰波, 张东升, 刘英杰, 张丽红, 许文锋, 段晨辉, 敦静怡, 刘正学
- ZHANG Leman, LAN Bo, ZHANG Dongsheng, LIU Yingjie, ZHANG Lihong, XU Wenfeng, DUAN Chenhui, DUN Jingyi, LIU Zhengxue
- 三峡水库涪陵-奉节段消落带优势草本植物生态位与种间联结性研究
- Niche and interspecific association of dominant herbaceous plants in the water-level-fluctuating zone of Fuling-Fengjie section of the Three Gorges Reservoir
- 生态学报. 2022, 42(8): 3228-3240
- Acta Ecologica Sinica. 2022, 42(8): 3228-3240
- http://dx.doi.org/10.5846/stxb202011162953
-
文章历史
- 收稿日期: 2020-11-16
- 网络出版日期: 2021-11-04

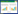




生态位理论是现代生态学的重要理论之一, 其中空间生态位、功能生态位以及多维超体积生态位在生态位理论中最具代表性[1]。生态位理论不仅在解释物种多样性、种群关系、群落定居与进化过程等方面发挥了重要作用[2-3], 而且能从宏观角度即生态循环领域深入理解种群、群落在生态系统中的地位及作用[4-6]。种间联结作为植物生态研究领域的重要组成部分, 能够在空间上反映群落相关性特征、有效表征物种的相互作用关系, 它受到种间竞争、他感作用、群落生境的异质性等多种因素的影响, 在空间上既有相互吸引的协同进化、又有互相排斥的拮抗作用[7], 进而能够很好地反映群落结构的空间稳定性, 对评估群落演替等级具有重要的参考价值[8-9]。因此, 生态位与种间联结二者结合研究, 可从时空尺度上统筹了解物种对资源的利用特征、种间协同与拮抗作用以及群落的稳定性, 且在生态修复中具有一定的实践参考价值, 例如:张静等[10]通过对青海玛多梭罗以礼草的生态位与种间联结分析, 发现其结构简单且多独立分布, 提出梭罗以礼草群落在生态恢复时应减少物种冗余, 即去除对群落的结构与功能恢复影响小的物种; 沈雪梨等[11]对百花山葡萄群落的生态位与种间联结的研究结果表明, 群落进入生殖阶段、竞争压力相对较小, 提出采用人工干预的方式提高物种的资源竞争力; 胡永歌等[12]对信南高速公路边坡植被生态位与种间联结的研究发现, 黑麦草、紫穗槐等平均生态位宽度较高的植物更适合边坡植被的恢复与重建。由此可见, 通过生态位与种间联结分析植物群落配置, 有助于植被生态修复的顺利实施。
近年来, 三峡水库消落带植被生态修复已逐渐成为国内外研究的热点, 研究内容大多集中在耐淹植物筛选、土壤种子库构建、地上植被优势物种配置模式研究等方面, 且从流域尺度进行生态位与种间联结的结合研究鲜有报道[13-16]; 而且, 相邻格子法作为植物生态位研究的常用采样方法, 能更好地揭示植物种群空间分布格局及种间关系、进而反映种群对环境资源的利用状况[17], 但该方法目前仅应用于下段(奉节-秭归)[18-19]、而对于中段(涪陵-奉节)与上段(江津-涪陵)还未见相关报道。因此, 本文在三峡水库中段(涪陵-奉节)消落带设置研究区, 探讨三峡水库消落带优势草本植物的生态位理论与种间联结特征, 以期揭示以下科学问题:(1)植被优势物种的生态位及种间联结性有何特征?(2)优势植物生态位与种间联结存在怎样的关系?相关研究结果能在一定程度上揭示三峡水库消落带优势草本植物的生态学特性、进而推测植被群落的演替趋势, 以期为植被生态恢复与重建提供理论基础或参考建议。
1 研究方法 1.1 小流域概况及研究区设置长江三峡水库涪陵-奉节段小流域(图 1), 地处三峡库区腹心地段, 属于典型的亚热带湿润季风气候, 降雨集中在5-9月, 年日照时数1000-1400 h, 年平均气温相对恒定、介于16-18 ℃, 年活动积温介于5500-6500℃, 土壤类型以黏土与沙壤土为主。综合考虑消落带间隔距离、交通状况、干扰状况、坡度坡向、土壤类型等实际情况, 选择具有一定代表性的一级支流4条, 自涪陵而下分别为梨香溪(涪陵区)、渠溪河(丰都县)、石桥河(万州区)和梅溪河(奉节县); 以每条一级支流与长江干流交汇处为原点分别向干流(纵向)、支流(横向)延伸, 按“就近原则”(距河口约1-2 km左右)各选取1条自然修复缓土坡消落带(坡度25°左右, 人为干扰小)作为研究区, 共8个(图 1、表 1)。

|
| 图 1 小流域行政区划及研究区GIS定位 Fig. 1 Administrative division of small watershed and GEOGRAPHIC INFORMATION SYSTEM orientation of research area |
| 编号 NO. |
坡度/(°) Slope Grade |
坡向 Slope Direction |
土壤类型 Soil type |
地理位置 Geographic location |
| 1 | 22 | 阳坡 | 黏土 | 29°39′56″N-107°11′9″E (梨香溪) |
| 2 | 22 | 阳坡 | 黏土 | 29°42′21″N-107°14′7″E (梨香溪河口附近长江干流) |
| 3 | 21 | 阳坡 | 黏土 | 29°54′2″-107°28′15″E (渠溪河) |
| 4 | 24 | 阴坡 | 黏土 | 29°54′20″N-107°29′37″E (渠溪河河口附近长江干流) |
| 5 | 24 | 阴坡 | 黏土 | 30°32′41″N-108°16′34″E (石桥河) |
| 6 | 23 | 阳坡 | 黏土 | 30°52′92″N-108°27′46″(石桥河河口附近长江干流) |
| 7 | 27 | 阴坡 | 沙壤土 | 31°04′82″N-109°53′09″E (梅溪河) |
| 8 | 25 | 阴坡 | 沙壤土 | 31°03′09″N-109°50′01″E (梅溪河河口附近长江干流) |
2019年6-7月份, 开展植被调查。针对每个研究区, 参考相邻格子法[20], 结合夏季洪汛等因素导致水位变化实际情况, 选取155-175 m高程段消落带, 沿坡面设置面积为16 m×20 m垂向样带, 并按1 m×1 m规格设置48个样方(图 2);8个研究区, 共计384个样方。植被调查指标主要包括植物的种类、数量、频度、盖度等。

|
| 图 2 相邻格子法样方(黄色)设置示意图 Fig. 2 Diagram of adjacent grid method for sampling (Yellow) |
利用Excel、SPSS 20.0进行数据整理、分析。选取频度≥5%的优势植物进行物种重要值、生态位宽度、生态位相似比、生态位重叠值和种间联结系数等分析(表 2)。
| 方法 Method |
公式 Formula |
指数解释 Index to explain |
备注 Remarks |
| 物种重要值(Iv) Species importance value (Iv) |

|
Iv是一个物种所能利用的各种资源总和, 综合表征植物在群落中的重要程度。 | |
| Levins生态位宽度值(Bsw) Levins niche width (Bsw) |
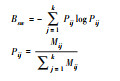
|
生态位宽度是物种对资源开发利用的程度, 数值越大, 对环境的适应能力越强。 | |
| Hurlbert生态位宽度值(Ba) Hurlbert niche width (Ba) |
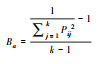
|
||
| 生态位相似比(Cih) Niche proportional similarity (Cih) |
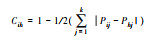
|
两个物种利用资源的相似程度, 判断物种是否存在竞争。 | |
| 生态位重叠值(Lih) Niche overlap(Lih) |
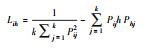
|
两个物种利用相同等级资源而相互叠加的状况, 重叠值越大, 对资源需求越相似, 可能存在激烈竞争(资源充足除外)。 | |
| 卡方检验检验(χ2) Chi-square test (χ2) |
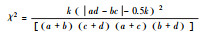
|
χ2值均为正, 当ad>bc, 为正联结; ad<bc, 为负联结。 | 能够体现植物的显著性, 但不可避免的会丧失植物多度等信息。 |
| 种间联结系数(AC) Interspecific association coefficient(AC) |
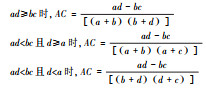
|
AC值进一步检验χ2检验的结果, 值趋近于1, 正联结性越强; 值趋近于-1, 负联结性越强; AC=0时, 种间无联结。 | 利用植物多度等信息判断种间联结的强弱程度。 |
| 共同出现百分率(PC) Cooccurrence percentage(PC) |
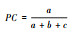
|
PC值测定植物的正联结程度, 结果趋近于1, 正关联紧密; 趋近于-1, 负关联紧密。 | 避免两种植物在同一样方均不出现的偶然性误差, 诠释种间正联结程度。 |
| Pearson相关系数(Px, y) Pearson correlation coefficient (Px, y) |
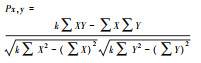
|
Px, y值描述的是两个变量线性相关程度, 绝对值越大, 相关性越强。 | 无法准确定义顺序变量各类别之间的距离, 计算出来的相关系数不是变量间的关联性的真实表示。 |
| Spearman秩相关系数(ρ) Spearman rank correlation coefficient(ρ) |
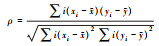
|
分级变量之间的相关程度, 分析植物同时出现的可能性, 对χ2检验进行有效补充。 | 两个变量的观测值是成对的等级评定资料采用的是取值的等级, 而不是取值本身。 |
| 方差比率(VR) Variance ratio(VR) |
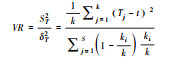
|
VR表征植物总体关联性: VR < 1, 植物负关联; VR=1, 植物无关联性; VR>1, 植物正关联。 |
|
| 统计量(W) Statistic(W) |
W=VR×k | W用来检验VR值偏离1的显著程度, 统计量W落入χ20.95, k < W < χ2(0.05, k), 植物种间关联不显著。 | |
RA、RF、RP对应某种植物的相对密度、相对频度、相对盖度; a、f、p为物种的密度、频度、盖度; k为样方数; Pij与Phj为植物i和植物h对j个资源的利用占该物种对全部资源的利用的百分率; Mij为植物i在样方j的重要值; ki为植物i出现的样方数, t为样方中植物的平均数; X(xi)和Y(yi)分别为植物x和植物y在同一样方中的重要值; 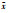 和 和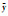 分别为植物x和植物y在同一样方中的平均值; a为植物A与植物B同时出现的样方数, b为只有植物B出现的样方数, c为只有植物A出现的样方数, d为植物A与植物B都不出现的样方数 分别为植物x和植物y在同一样方中的平均值; a为植物A与植物B同时出现的样方数, b为只有植物B出现的样方数, c为只有植物A出现的样方数, d为植物A与植物B都不出现的样方数 | |||
本次植物调查种类计57种、隶属22科44属, 物种数分布最多的科分别为菊科(14种)、禾本科(9种)、莎草科(8种)。根据重要值由大到小排序, 本文将排名前15的植物选作优势植物进一步分析(表 3, 下同):禾本科有5种, 分别是马唐(Digitaria sanguinalis)、狗尾草(Setaria viridis)、狗牙根(Cynodon dactylon)、牛鞭草(Hemarthria altissima)、稗(Echinochloa crusgalli); 菊科有3种, 分别为鬼针草(Bidens pilosa)、苍耳(Xanthium sibiricum)、鳢肠(Eclipta prostrata)。优势植物Levins生态位宽度(Bsw)介于2.28-5.65, Hurlbert生态位宽度(Ba)介于0.04-0.76之间。从总体趋势看, 优势植物Iv值越大(小), 其Ba、Bsw值越大(小), 二者之间呈普遍正相关, 尤其是:15种优势植物中, 有且只有狗牙根、鬼针草和苍耳的Iv值大于0.10, 分别为0.46 ± 0.26、0.12 ± 0.09和0.10 ± 0.09, 显著(P < 0.05)高于其它优势植物; 也有且只有狗牙根、鬼针草和苍耳的Ba与Bsw值分别大于5.00与0.50, 分别为5.65与0.76、5.59与0.64、5.45与0.55, 也显著高于其它优势植物。
| 编号 No. |
植物名称 Species |
科 Science |
重要值±标准差 Iv±SD |
Levins指数 Bsw |
Hurlbert指数 Ba |
| 1 | 狗牙根Cynodon dactylon (Linn.) Pers | 禾本科Gramineae | 0.46±0.26 | 5.65 | 0.76 |
| 2 | 鬼针草Bidens pilosa linn. | 菊科Compositae | 0.12±0.09 | 5.59 | 0.64 |
| 3 | 苍耳Xanthium sibiricum | 菊科Compositae | 0.10±0.09 | 5.45 | 0.55 |
| 4 | 稗Echinochloa crusgalli | 禾本科Gramineae | 0.04±0.08 | 4.59 | 0.20 |
| 5 | 牛鞭草Hemarthria altissima | 菊科Compositae | 0.04±0.11 | 3.82 | 0.10 |
| 6 | 狗尾草Setaria viridis | 禾本科Gramineae | 0.03±0.07 | 4.36 | 0.16 |
| 7 | 空心莲子草Alternanthera philoxeroides | 锦葵科Malvaceae | 0.03±0.06 | 4.42 | 0.18 |
| 8 | 马唐Digitaria sanguinalis | 禾本科Gramineae | 0.03±0.07 | 4.19 | 0.13 |
| 9 | 酸浆Physali alkekengi L. | 酢浆草科Oxalidaceae | 0.02±0.06 | 4.02 | 0.13 |
| 10 | 水蓼Polygonum hydropiper L. | 蓼科Polygonaceae | 0.01±0.04 | 4.86 | 0.10 |
| 11 | 黄花草木樨Melilotus officinalis L. | 豆科Leguminosae | 0.01±0.04 | 4.02 | 0.13 |
| 12 | 鳢肠Eclipta prostrata | 菊科Compositae | 0.01±0.03 | 4.82 | 0.07 |
| 13 | 苘麻Abutilon theophrasti Medicus | 锦葵科Malvaceae | 0.01±0.02 | 3.88 | 0.10 |
| 14 | 香附子Cyperus rotundus | 莎草科Cyperaceae | 0.007±0.02 | 4.24 | 0.08 |
| 15 | 藨草Scirpus triqueter L. | 莎草科Cyperaceae | 0.006±0.03 | 2.28 | 0.04 |
由表 4可知, 优势植物105个种对的生态位相似比(Cih)介于0.00-0.65, 均值为0.16。Cih大于0.50的种对仅有4个, 占总数的3.80%, 分别是狗牙根与鬼针草(0.66)、狗牙根与苍耳(0.54)、鬼针草与苍耳(0.58)、狗尾草与黄花草木樨(0.52), 远高于其它植物组合; Cih高于平均值(0.16)的种对数为45个, 占总数的46.67%;Cih不足0.1的种对数为43个, 占总数的40.95%, 其中藨草与狗尾草(0.003)、藨草与酸浆(0.002)、藨草与黄花草木樨(0.001)的Cih值均远低于其它植物组合。
| 编号No. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
| 1 | 1.00 | 0.66 | 0.54 | 0.29 | 0.11 | 0.20 | 0.22 | 0.18 | 0.25 | 0.18 | 0.14 | 0.20 | 0.18 | 0.19 | 0.05 |
| 2 | 1.00 | 0.58 | 0.30 | 0.08 | 0.30 | 0.20 | 0.23 | 0.15 | 0.29 | 0.20 | 0.17 | 0.12 | 0.13 | 0.03 | |
| 3 | 1.00 | 0.27 | 0.16 | 0.36 | 0.27 | 0.22 | 0.05 | 0.23 | 0.25 | 0.16 | 0.11 | 0.14 | 0.03 | ||
| 4 | 1.00 | 0.08 | 0.18 | 0.29 | 0.38 | 0.03 | 0.36 | 0.08 | 0.16 | 0.35 | 0.06 | 0.03 | |||
| 5 | 1.00 | 0.03 | 0.48 | 0.03 | 0.02 | 0.01 | 0.01 | 0.30 | 0.01 | 0.18 | 0.10 | ||||
| 6 | 1.00 | 0.09 | 0.19 | 0.01 | 0.22 | 0.53 | 0.05 | 0.12 | 0.03 | 0.003 | |||||
| 7 | 1.00 | 0.16 | 0.01 | 0.19 | 0.02 | 0.25 | 0.17 | 0.18 | 0.09 | ||||||
| 8 | 1.00 | 0.02 | 0.39 | 0.08 | 0.11 | 0.22 | 0.05 | 0.01 | |||||||
| 9 | 1.00 | 0.02 | 0.00 | 0.03 | 0.05 | 0.12 | 0.002 | ||||||||
| 10 | 1.00 | 0.09 | 0.15 | 0.16 | 0.03 | 0.06 | |||||||||
| 11 | 1.00 | 0.02 | 0.03 | 0.01 | 0.001 | ||||||||||
| 12 | 1.00 | 0.07 | 0.23 | 0.39 | |||||||||||
| 13 | 1.00 | 0.09 | 0.01 | ||||||||||||
| 14 | 1.00 | 0.08 | |||||||||||||
| 15 | 1.00 | ||||||||||||||
| 各编号所对应的植物见表 3 | |||||||||||||||
由表 5可知, 优势植物生态位重叠值(Lih)的整体变化介于0-0.68之间。Lih大于0.50的种对有7个、占总数的6.67%, 分别是狗牙根与鬼针草(0.68)、狗牙根与苍耳(0.53)、苍耳与鬼针草(0.63)、狗尾草与苍耳(0.51)、牛鞭草与空心莲子草(0.64)、狗尾草与黄花草木樨(0.65)、水蓼与马唐(0.52);Lih处于0.20-0.50之间的有32对, 占总数的30.47%;Lih小于0.20的有66对, 占总数的62.85%, 其中:狗尾草与藨草、黄花草木樨与藨草这两个种对的Lih为0, 表明植物之间相互独立存在。
| 编号No. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
| 1 | 1.00 | 0.68 | 0.53 | 0.25 | 0.09 | 0.17 | 0.18 | 0.17 | 0.46 | 0.12 | 0.24 | 0.25 | 0.26 | 0.30 | 0.09 |
| 2 | 1.00 | 0.63 | 0.30 | 0.10 | 0.41 | 0.22 | 0.29 | 0.23 | 0.32 | 0.32 | 0.20 | 0.18 | 0.19 | 0.09 | |
| 3 | 1.00 | 0.31 | 0.30 | 0.51 | 0.36 | 0.24 | 0.04 | 0.21 | 0.43 | 0.16 | 0.13 | 0.20 | 0.07 | ||
| 4 | 1.00 | 0.07 | 0.17 | 0.29 | 0.46 | 0.02 | 0.43 | 0.08 | 0.13 | 0.44 | 0.03 | 0.04 | |||
| 5 | 1.00 | 0.02 | 0.64 | 0.01 | 0.01 | 0.01 | 0.002 | 0.29 | 0.002 | 0.12 | 0.07 | ||||
| 6 | 1.00 | 0.07 | 0.17 | 0.002 | 0.25 | 0.65 | 0.03 | 0.16 | 0.01 | 0.00 | |||||
| 7 | 1.00 | 0.17 | 0.004 | 0.22 | 0.01 | 0.23 | 0.20 | 0.17 | 0.07 | ||||||
| 8 | 1.00 | 0.01 | 0.52 | 0.06 | 0.08 | 0.18 | 0.01 | 0.01 | |||||||
| 9 | 1.00 | 0.01 | 0.00 | 0.02 | 0.02 | 0.10 | 0.00 | ||||||||
| 10 | 1.00 | 0.07 | 0.08 | 0.22 | 0.01 | 0.09 | |||||||||
| 11 | 1.00 | 0.01 | 0.01 | 0.00 | 0.00 | ||||||||||
| 12 | 1.00 | 0.05 | 0.18 | 0.49 | |||||||||||
| 13 | 1.00 | 0.08 | 0.01 | ||||||||||||
| 14 | 1.00 | 0.05 | |||||||||||||
| 15 | 1.00 |
种对间总体存在正相关关系的种对数为55、负相关关系的种对数为50, 占比分别为52.38%和47.62%。正负关联比为1.09, 正关联数并不明显优于负关联(图 3)。χ2检验的结果显示植被种间关联程度呈现极显著正关联(P < 0.01, 下同)的种对数为22, 占总数的20.95%;显著正关联(P < 0.05, 下同)的种对数为5, 占4.76%;显著负关联(P < 0.05, 下同)的种对数为8, 占7.61%;极显著负关联(P < 0.01, 下同)的种对数为13, 占12.38%。

|
| 图 3 优势植物种间联结性χ2值半矩阵图 Fig. 3 Semi-matrix diagram of interspecies connectivity χ2 of dominant plants |
对于联结程度AC而言, AC值趋近于1, 植物种间的正联结性越强, 植物之间相互作用越有利于互相发展; 相反, AC值趋近于-1, 植物种间的负联结性越强, 植物之间相互作用关系越不利于互相发展; 而AC=0, 物种相互独立。通过图 4, 发现:0.6≤AC的种对数为7, 占比6.67%;0.20≤AC<0.60的种对数为29, 占比27.62%;-0.20≤AC<0.20的种对数为42, 占比40%;-0.60≤AC<-0.20的种对数为16, 占比15.23%;AC<-0.60的种对数为11, 占比10.47%。酸浆与藨草、黄花草木樨与藨草这两个种对间AC值为-1, 实地调查中未发现两组植物同时出现(图 4)。

|
| 图 4 优势植物种间联结性AC值半矩阵图 Fig. 4 Semi-matrix diagram of AC value of interspecific association of dominant plants AC:种间联结系数Interspecific association coefficient |
为了克服AC值受到d值(两种植物都不出现的样方数)影响造成的偶然性误差, 再选用PC值(物种共同出现百分率)进一步分析种间联结程度。PC值趋近于1, 正关联紧密; 趋近于-1, 负关联紧密。发现(图 5):极显著正相关0.7≤PC的种对数为1, 占比4.76%;显著正相关0.40≤PC<0.70的种对数为3, 占比9.52%;不显著正相关0.20≤PC<0.40的种对数为31, 占比5.70%;无相关PC<0.20的种对数为70, 占比5.70%, 整体呈现的正关联性不高。

|
| 图 5 优势植物种间联结性PC值半矩阵图 Fig. 5 Semi-matrix diagram of PC value of interspecific association of dominant plants PC:共同出现百分率Cooccurrence percentage |
Pearson相关系数检验结果(表 6)显示, 呈正相关的种对数为37, 占比35.23%, 其中:呈极显著正相关、显著正相关及不显著正相关的种对数分别为14、6和17, 分别占比13.33%、5.71%和16.19%;呈负相关的种对数为68, 占比64.76%, 其中:呈极显著负相关、显著负相关及不显著负相关的种对数分别为17、20和31, 分别占比16.19%、19.04%和29.52%。正负相关种对的比值为0.54, 负联结占优势。Spearman秩相关系数检验结果(表 6)显示, 呈正相关的种对数为45, 占比42.85%, 其中:呈极显著正相关、显著正相关及不显著正相关的种对数分别为20、6和19, 分别占比19.04%、5.71%和18.09%;呈负相关的种对数为60, 占比57.14%, 其中:呈极显著正相关、显著正相关及不显著正相关的种对数分别为28、11和21, 分别占比26.66%、10.47%和20.00%。正负相关种对比值为0.75, 负联结占优势。
| 编号No. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
| 1 | 1.00 | 0.10 | -0.26 | -0.17 | -0.17 | -0.29 | -0.26 | -0.21 | 0.43 | -0.20 | -0.19 | -0.05 | 0.02 | 0.12 | -0.11 |
| 2 | 0.02 | 1.00 | 0.15 | -0.05 | -0.31 | 0.12 | -0.26 | 0.03 | -0.06 | 0.19 | 0.10 | -0.12 | -0.16 | -0.08 | -0.1 |
| 3 | -0.26 | 0.13 | 1.00 | -0.12 | 0.06 | 0.31 | 0.08 | 0.02 | -0.36 | 0.01 | 0.26 | -0.11 | -0.14 | -0.01 | -0.10 |
| 4 | -0.29 | -0.10 | -0.03 | 1.00 | -0.06 | -0.01 | 0.17 | 0.30 | -0.16 | 0.25 | -0.03 | 0.07 | 0.33 | -0.06 | 0.00 |
| 5 | -0.37 | -0.25 | 0.11 | -0.08 | 1.00 | -0.18 | 0.41 | -0.12 | -0.11 | -0.15 | -0.16 | 0.29 | -0.14 | 0.19 | 0.25 |
| 6 | -0.38 | 0.17 | 0.35 | 0.00 | -0.12 | 1.00 | -0.15 | 0.13 | -0.22 | 0.06 | 0.52 | -0.12 | 0.07 | -0.18 | -0.11 |
| 7 | -0.37 | -0.19 | 0.09 | 0.14 | 0.59 | -0.11 | 1.00 | 0.01 | -0.23 | 0.02 | -0.17 | 0.18 | 0.10 | 0.10 | 0.22 |
| 8 | -0.28 | 0.02 | -0.03 | 0.36 | -0.12 | 0.03 | 0.03 | 1.00 | -0.14 | 0.30 | -0.02 | 0.00 | 0.15 | -0.08 | -0.04 |
| 9 | 0.32 | -0.08 | -0.35 | -0.17 | -0.12 | -0.17 | -0.18 | -0.14 | 1.00 | -0.15 | -0.19 | -0.11 | 0.00 | 0.04 | -0.11 |
| 10 | -0.35 | 0.10 | -0.04 | 0.34 | -0.12 | 0.14 | 0.10 | 0.46 | -0.13 | 1.00 | -0.03 | 0.08 | 0.01 | -0.10 | 0.08 |
| 11 | -0.13 | 0.07 | 0.26 | -0.10 | -0.13 | 0.59 | -0.17 | -0.07 | -0.15 | -0.06 | 1.00 | -0.17 | -0.08 | -0.16 | -0.10 |
| 12 | -0.08 | -0.11 | -0.12 | -0.02 | 0.21 | -0.12 | 0.11 | -0.05 | -0.11 | -0.04 | -0.13 | 1.00 | -0.05 | 0.16 | 0.50 |
| 13 | -0.01 | -0.11 | -0.15 | 0.35 | -0.11 | 0.04 | 0.08 | 0.08 | -0.10 | 0.13 | -0.11 | -0.06 | 1.00 | 0.04 | -0.04 |
| 14 | 0.11 | -0.07 | -0.01 | -0.12 | 0.04 | -0.12 | 0.06 | -0.10 | -0.01 | -0.10 | -0.12 | 0.09 | -0.01 | 1.00 | 0.12 |
| 15 | -0.10 | -0.05 | -0.06 | -0.03 | 0.03 | -0.07 | 0.01 | -0.05 | -0.07 | 0.04 | -0.07 | 0.47 | -0.05 | 0.00 | 1.00 |
| 左下Pearson相关性半矩阵表、右上Spearman相关性半矩阵表 | |||||||||||||||
对优势植物总体关联强度(表 7)分析可知, 优势植物整体呈现不显著正关联(VR=1.09), χ2临界值为χ2(0.95, 384)=339.58、χ2(0.05, 384)=430.69。利用统计量W检测VR值偏离的显著性, W=421.03介于χ2(0.95, 384)与χ2(0.05, 384)之间, 表明三峡水库中段植物的总体关联强度为不显著正关联。再根据优势植物种对间联结性比较(表 8)可了解到:χ2检验、Pearson相关性、Spearman秩相关性这三种检验方法分析结果均表明, 优势植物种间相关性不强、联结性较弱。
| ST2 | δT2 | 方差比率 Variance ratio(VR) |
检查统计量 Statistic(W) |
χ2临界值χ2 threshold | 测定结果 Results |
|
| χ2(0.95, 384) | χ2(0.05, 384) | |||||
| 3.25 | 2.96 | 1.09 | 421.03 | 339.58 | 430.69 | 不显著正关联 |
| 检验方法 Test methods |
正关联Positive correlation | 负关联Negative correlation | |||||||
| 极显著 Distinctly significant (P≤0.01) |
显著 Significant (P≤0.05) |
不显著 Not significant (P>0.05) |
总数 Sum |
极显著 Distinctly significant (P≤0.01) |
显著 Significant (P≤0.05) |
不显著 Not significant (P>0.05) |
总数 Sum |
||
| χ2检验 Chi-square test |
22 | 5 | 28 | 55 | 8 | 13 | 29 | 50 | |
| Pearson相关性 Pearson correlation coefficient |
14 | 6 | 17 | 37 | 17 | 20 | 31 | 68 | |
| Spearma秩相关性 Spearman rank correlation coefficient |
20 | 6 | 19 | 45 | 28 | 11 | 21 | 60 | |
针对优势植物的种间联结性Pearson相关系数、Spearman秩相关系数, 分别与相应的生态位相似比(Cih)、生态位重叠值(Lih)进行回归分析(图 6), 结果发现:均呈现极显著正相关(P < 0.001), 总体上表现为优势植物的种间正(负)联结越强、生态位相似性与重叠程度越大(小)。

|
| 图 6 优势植物种回归分析 Fig. 6 Regression analysis of dominant plant species |
生态位宽度和重要值均为衡量物种在群落中的地位和作用的综合数量指标, 但二者的生态学意义各有侧重[21]:生态位宽度衡量的是物种在群落中所占据的位置及最小生存阈值, 反映了物种对环境的适应能力以及对资源的利用现状; 而重要值是一个物种所能利用的各种资源的总和, 也是种间关系分析的基础, 表征了物种在群落中的优势程度或重要性。本研究结果从总体趋势看, 优势植物Iv值越大(小)、其Ba与Bsw值越大(小), 二者之间呈普遍正相关; 不过也有部分优势植物的重要值与其生态位宽度呈负相关, 如牛鞭草的重要值相对较大(0.04)、但其生态位宽度相对较小(Bsw, 3.82;Ba, 0.10), 而水蓼、鳢肠等重要值相对较小(均为0.01)、却具有较大的生态位宽度(Bsw分别达到4.86和4.82)。由此可见, 从优势植物的重要值与生态位宽度之间的关系看, 三峡水库消落带中段优势植物的重要值是影响其生态位宽度的主要因素但并非唯一因素, 这与大多数学者研究结果一致[22-23], 表明三峡水库消落带优势植物对环境具有较强的生态适应性。
在复杂的自然环境中, 两种或两种以上植物会因生态习性相似而产生生态重叠。通常情况下, 生态位相似比(Cih)越大(小), 生态位重叠值(Lih)越大(小), 种间竞争激烈程度越大(小)(资源充足除外)[24]。本研究结果表明, 狗牙根与鬼针草(Cih=0.66、Lih=0.68)、狗牙根与苍耳(Cih=0.54、Lih=0.53)等种对Cih较大、其Lih也较大, 提示:三峡水库消落带反季节、长时间、高落差等极端环境胁迫, 导致物种多样性急剧下降、环境资源相对充足, 生态习性相似的优势物种之间能够实现稳定共存、协同发展[25]; 狗尾草与藨草(Cih=0.003、Lih=0)、黄花草木樨与藨草(Cih=0.001、Lih=0.007)等种对Cih较小、其Lih也较小, 提示:这些植物在相同环境下, 资源利用的相似性小, 种间竞争不激烈[26]。但也存在Cih较小、其Lih却较大的情况, 如苍耳与狗尾草(Cih=0.31、Lih=0.51)等种对, 这可能与三峡水库消落带优势植物群落结构简单、沿海拔高程呈带状分布(优势植物相对聚集在局部适宜的生境斑块、生态位宽度相对压缩)等因素导有关[13, 27]。
3.2 优势植物种间联结特征种间联结性作为衡量群落结构稳定性的指标, 在植物群落的所有生命周期中发挥着不可替代的作用[28]。植物群落次生演替的初期阶段, 其结构处于不稳定的负联结或无相关状态; 随着演替过程的进行, 优势植物能够发挥自身的生物学特性并利用种间联结使自身处于相对稳定状态[29]。本调查通过对三峡水库消落带中段优势植物种间联结测定结果表明, 优势植物种间整体呈现不显著正关联(VR=1.09), 群落结构不稳定, 个别植物(如酸浆与藨草、黄花草木樨与藨草等种对)表现出相对独立性。χ2值半矩阵图结果显示, 种对间的正负联结比为1.10, 提示:三峡水库消落带优势植物群落仍处于不稳定的动态演替阶段, 其结构相对松散、种间分离程度较大、稳定性不高。
AC值与PC值的Pearson与Spearman秩相关检验, 可以在χ2检验的基础上进一步对调查结果进行验证、反映种间联结性。稗+苘麻、狗尾草+黄花草木樨、马唐+水蓼种对在所有检验中均表现出正相关, 说明在忽略统计误差的前提下这几个种对正联结性优于其它种对。而105个优势植物种对中, 表现出较强正联结(AC≥0.6)的种对数为6、显著正相关(PC≥0.4)的种对数为4, Pearson与Spearman秩相关检验呈现显著正相关的种对数分别为20、26。将χ2检验、AC值、PC值以及Pearson与Spearman秩相关检验的多种检验结果进行统筹分析, 均表征出三峡水库消落带中段植被群落结构不稳定、种间联结性弱、植物独立性强等特点。其可能原因是:三峡水库冬季蓄水造成水位上涨, 长期水淹的极端环境会令大部分植物难以存活; 夏季水位下降, 土壤虽然出露利于植物生长, 但季节性降雨会导致有些植物因高位地表原土冲刷覆盖而出现死亡, 如此就导致了样方内植物多样性有所降低。此外, 周期性年际干湿交替使得消落带原本异质性环境生态趋于相同, 适者生存的生态选择理论使得植物在长期进化中淘汰了大多数原有植物、弱化了群落种间关系, 样方植物明显退化, 植物种间表现出明显的独立性, 这与实际样方调查的结果相一致。而本调查还发现, 种对间χ2、AC值、PC值分析与Pearson、Spearman秩相关系数分析结果之间存在一定差异, 例如狗牙根与鬼针草之间χ2、AC值、PC值均表现出极显著正关联, 而Pearson和Spearman秩相关系数表现出正相关性不强, 可能与不同检验方法导致结果不同有关[28]。总之, 三峡水库消落带中段植物仅极个别种对间表现出正联结, 整体仍表现为种间联结性不强, 表明群落生态系统脆弱。
3.3 优势植物生态位与种间联结的相关性特征植物的生态位与种间联结性之间存在紧密关联:不同植物种间正联结体现了植物占据自然资源的相似性与重叠性, 负联结体现了种间的排斥与分离[30]; 在较强干扰情况下, 生态位与种间联结之间也能相辅相成, 共同表征群落的发展演替趋势[31-32]。三峡水库消落带中段优势植物种间联结与生态位回归分析表明, 二者之间总体上表现为优势植物的种间正(负)联结越强、其生态位相似性与重叠程度越大(小), 这与大多数学者的研究结果一致[23-24]。但也有极少数优势植物种间负联结越强、其Lih不一定小, 如牛鞭草与空心莲子草的种对呈现显著负相关、其Lih高达0.64。因此, 在对三峡水库消落带进行植被修复选择植物配置时, 选择显著正相关的种对可以最大程度提高植被的群落稳定性。
4 结论与生态修复策略综上所述, 三峡水库消落带中段优势植物间生态位差异性较大, 种间关联性较弱, 群落分化程度明显, 消落带植物群落结构仍处于不稳定发展阶段。本文研究结果在一定程度上反映了三峡水库消落带植被退化生态系统现状, 为最大程度提高消落带植被群落稳定性和物种多样性, 针对小流域植被恢复与重建有如下参考建议:在整体上, 可以采用以狗牙根(Cynodon dactylon (Linn.)Pers)、苍耳(Xanthium sibiricum)、鬼针草(Bidens pilosa L.)等为先锋植物, 以狗尾草(Setaria viridis)、稗(Echinochloa crusgalli)、牛鞭草(Hemarthria altissima)等为伴生种的群落配置模式进行宏观布局; 在局部上, 可以采用稗(Echinochloa crusgalli)+苘麻(Abutilon theophrasti Medic.)、狗尾草(Setaria viridis)+黄花草木樨(Melilotus officinalis L.)、马唐(Digitaria sanguinalis)+水蓼(Polygonum hydropiper L.)等种对配置模式进行实地配置。
| [1] |
魏凯, 郑建忠, 杨铭, 何雅琴, 史晓洁, 邓传远. 平潭光幼屿典型植物群落优势种生态位特征. 森林与环境学报, 2020, 40(3): 290-297. |
| [2] |
简尊吉, 马凡强, 郭泉水, 秦爱丽, 肖文发. 三峡水库峡谷地貌区消落带优势植物种群生态位. 生态学杂志, 2017, 36(2): 328-334. |
| [3] |
张金屯. 数量生态学. 北京: 科学出版社, 2011.
|
| [4] |
张爱英, 熊高明, 樊大勇, 谢宗强. 三峡水库运行对淹没区及消落带植物多样性的影响. 生态学杂志, 2016, 35(9): 2505-2518. |
| [5] |
郝志远, 李素清. 阳泉矿区煤矸石山复垦地不同植被下草本植物群落生态关系. 应用与环境生物学报, 2018, 24(5): 1158-1164. |
| [6] |
Sueltenfuss J P, Ocheltree T W, Cooper D J. Evaluating the realized niche and plant-water relations of wetland species using experimental transplants. Plant Ecology, 2020, 221(5): 334-347. |
| [7] |
许宝坤, 许晓岗, 李垚, 李小东, 陈水飞, 丁晖, 蒋晓辉, 苟凌晨, 方炎明. 黄山常绿阔叶林甜槠群落优势种种间关联分析. 南京林业大学学报: 自然科学版, 2019, 43(4): 77-84. |
| [8] |
张明霞, 王得祥, 康冰, 张岗岗, 刘璞, 杜焰玲, 于飞. 秦岭华山松天然次生林优势种群的种间联结性. 林业科学, 2015, 51(1): 12-21. |
| [9] |
简尊吉, 李堂中, 郭泉水, 秦爱丽, 马凡强, 裴顺祥, 肖文发. 三峡水库峡谷地貌区消落带主要植物种间关联性研究. 西北植物学报, 2016, 36(10): 2086-2093. DOI:10.7606/j.issn.1000-4025.2016.10.2086 |
| [10] |
张静, 才文代吉, 索南才仁, 温小成, 王生彩, 杨毛吉, 仁增欧珠, 孙海群. 青海玛多梭罗以礼草生存群落种间关联及生态位. 草业科学, 2019, 36(11): 2752-2765. |
| [11] |
沈雪梨, 吴远密, 朱艺璇, 童玲, 胡巧立, 蒋万杰, 范雅倩, 夏晓飞, 沐先运, 张志翔. 濒危植物百花山葡萄群落物种生态位特征与种间联结研究. 植物科学学报, 2020, 38(2): 195-204. |
| [12] |
胡永歌, 徐恩凯, 陈延永, 曹娓, 田国行. 信南高速公路边坡植物生态位及种间联结性. 四川农业大学学报, 2019, 37(5): 663-669. |
| [13] |
何蕊廷, 杨康, 曾波, 李瑞, 牛汉刚, 史邵华, 阿依巧丽, 苏晓磊. 三峡水库消落区植被在差异性水淹环境中的分布格局. 生态学报, 2020, 40(3): 834-842. |
| [14] |
武帅楷. 三峡水库消落带植物群落生态学研究[D]. 重庆: 重庆大学, 2019.
|
| [15] |
雷波, 王业春, 由永飞, 张晟, 杨春华. 三峡水库不同间距高程消落带草本植物群落物种多样性与结构特征. 湖泊科学, 2014, 26(4): 600-606. |
| [16] |
Zhang L Y, Tian M, Peng C, Fu C, Li T Z, Chen Y, Qiu Y, Huang Y M, Wang H B, Li Z, Yang F M. Nitrogen wet deposition in the Three Gorges Reservoir area: characteristics, fluxes, and contributions to the aquatic environment. Science of the Total Environment, 2020, 738: 140309. DOI:10.1016/j.scitotenv.2020.140309 |
| [17] |
郭燕, 杨邵, 沈雅飞, 肖文发, 程瑞梅. 三峡水库消落带现存植物自然分布特征与群落物种多样性研究. 生态学报, 2019, 39(12): 4255-4265. |
| [18] |
孙鹏飞, 沈雅飞, 王丽君, 杨邵, 郭燕, 肖文发, 程瑞梅. 三峡库区秭归段水位消落带草本植物多样性分析. 林业科学研究, 2020, 33(6): 96-104. |
| [19] |
朱妮妮, 郭泉水, 秦爱丽, 裴顺祥, 马凡强, 朱莉, 简尊吉. 三峡水库奉节以东秭归和巫山段消落带植物群落动态特征. 生态学报, 2015, 35(23): 7852-7867. |
| [20] |
曹静, 白玉宏, 张钦弟, 毕润成. 山西太岳山接骨木群落优势种种间关联性分析. 生态学杂志, 2015, 34(5): 1313-1319. |
| [21] |
李姗泽, 邓玥, 施凤宁, 胡明明, 庞博慧, 王雨春, 李坤, 陈铭, 彭文启, 渠晓东, 包宇飞, 孟晶晶. 水库消落带研究进展. 湿地科学, 2019, 17(6): 689-696. |
| [22] |
袁方, 储寅芳, 朱凤, 孙国俊, 李粉华, 潘云枫, 张海艳, 韩敏, 季敏. 江苏省宜兴市茶园秋季杂草种间生态关系及群落分类. 杂草科学, 2015, 33(2): 18-25. DOI:10.3969/j.issn.1003-935X.2015.02.004 |
| [23] |
吴霖东, 李婷婷, 傅国林, 吴初平, 余裕龙, 王志高, 盛卫星, 朱锦茹, 江波, 焦洁洁, 文香英, 袁位高. 极小种群野生植物浙江安息香的生态位和种间关系. 浙江大学学报: 农业与生命科学版, 2020, 46(4): 459-474. |
| [24] |
Roscher C, Schumacher J, Gubsch M, Lipowsky A, Weigelt A, Buchmann N, Schulze E D, Schmid B. Interspecific trait differences rather than intraspecific trait variation increase the extent and filling of community trait space with increasing plant diversity in experimental grasslands. Perspectives in Plant Ecology, Evolution and Systematics, 2018, 33: 42-50. DOI:10.1016/j.ppees.2018.05.001 |
| [25] |
Konar M, Todd M J, Muneepeerakul R, Rinaldo A, Rodriguez-Iturbe I. Hydrology as a driver of biodiversity: controls on carrying capacity, niche formation, and dispersal. Advances in Water Resources, 2013, 51: 317-325. DOI:10.1016/j.advwatres.2012.02.009 |
| [26] |
董冬, 许小天, 周志翔, 何云核, 王存志, 石方可, 高陆陆. 安徽九华山风景区古树群落主要种群生态位的动态变化. 生态学杂志, 2019, 38(5): 1292-1304. |
| [27] |
刘润红, 陈乐, 涂洪润, 梁士楚, 姜勇, 李月娟, 黄冬柳, 农娟丽. 桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结. 生态学报, 2020, 40(6): 2057-2071. |
| [28] |
Wédjangnon A A, Kuiga N B S, Houêtchégnon T, Ouinsavi C A I N. Spatial distribution and interspecific association patterns between Mansonia altissima A. Chev., Ceiba pentandra (L.) Gaertn and Triplochiton scleroxylon K. Schum. in a moist semi-deciduous forest. Annals of Forest Science, 2020, 77(1): 6. DOI:10.1007/s13595-019-0913-0 |
| [29] |
徐满厚, 刘敏, 翟大彤, 刘彤. 植物种间联结研究内容与方法评述. 生态学报, 2016, 36(24): 8224-8233. |
| [30] |
Rousset O, Lepart J. Positive and negative interactions at different life stages of a colonizing species (Quercus humilis). Journal of Ecology, 2000, 88(3): 401-412. DOI:10.1046/j.1365-2745.2000.00457.x |
| [31] |
王昱, 毕润成, 茹文明. 山西浊漳河北源湿地草本植物群落优势种生态位与种间关系. 草业科学, 2018, 35(2): 276-285. |
| [32] |
李霖, 李素清. 阳泉矿区煤矸石山复垦地不同植被下草本植物群落优势种种间关系及生态位. 中国农学通报, 2019, 35(1): 80-87. |