 2022, Vol. 42
2022, Vol. 42文章信息
- 洪迎新, 刘东升, 马宏海, 陈宇琛, 陈求稳, 施文卿, 朱昊彧
- HONG Yingxin, LIU Dongsheng, MA Honghai, CHEN Yuchen, CHEN Qiuwen, SHI Wenqing, ZHU Haoyu
- 澜沧江梯级开发下鱼类支流生境替代效果
- Effects of fish habitat substitution in tributaries under the cascade hydropower development of Lancang River
- 生态学报. 2022, 42(8): 3191-3205
- Acta Ecologica Sinica. 2022, 42(8): 3191-3205
- http://dx.doi.org/10.5846/stxb202009042303
-
文章历史
- 收稿日期: 2020-09-04
- 网络出版日期: 2021-12-17





2. 中水珠江规划勘测设计有限公司, 广州 510610
2. China Water Resources Pearl River Planning Surveying & Designing Co., Ltd., Guangzhou 510610, China
为应对气候变化, 减少温室气体排放, 水电作为低碳的可再生清洁能源被世界各国广泛利用。然而, 水库大坝的建设改变了河流天然水文过程, 对河流生境造成深远的影响, 导致鱼类群落结构出现演替、多样性大量丧失[1-2], 甚至有些物种走向灭绝[3-4]。为减缓水电开发对土著鱼类的不利影响, 过鱼设施[5-6]、增殖放流[7-8]、多目标调度[9-11]等一系列保护措施被应用于实践, 对鱼类资源的保护起到了积极作用。然而, 对于梯级开发河流(如我国重点开发的十三大水电基地), 开发后的河流生境破碎, 或将难以满足鱼类完整生活史需求, 这就导致以往的保护措施受到限制[12]。因此, 修复和保护具备自然条件属性的栖息地成为近年来研究热点[13], 其中基于“干流开发、支流保护”开发理念的支流生境替代保护, 一经提出便受到了广泛的关注[14-23]。
支流生境替代是指以天然条件为基础, 适当施以人工综合措施, 对支流生境进行维护、修复与重建, 从而为在干流中受到水电开发影响的土著、洄游、特有及濒危保护性鱼类提供补偿生境[15, 18-19, 23-24]。Kruk等[20]认为支流生境保护的重点是保持河流的连通性和栖息地结构的多样性, 以及适当的流量、良好的水质; Marques等[21]研究发现Paraná河上游支流丰富的渔业资源对维持干流鱼类群落多样性意义重大, 可有效降低因干流建坝导致的生物群落均质化趋势; Lopes等[22]通过“标记重补法”, 在巴西São Francisco河干支流监测濒危热带鱼类鲮脂鲤属(Prochilodus)季节性洄游及繁殖过程, 研究表明水库支流对热带洄游性鱼类产卵场的补偿作用; Garcia等[23]发现Congonhas河鱼类群落和生境特征与干流表现出较大相似性, 其洄游性鱼类总数占到干流Paraná河流域的29%, 表明该支流能够为干流受Capivara大坝影响的鱼类提供补偿性生境。近年来, 我国也在鱼类支流生境替代上开展了积极实践。典型案例包括, 长江取消了赤水河10个水电梯级规划项目, 用以保护上游珍稀特有鱼类[14, 24]; 金沙江规划了黑水河作为白鹤滩水电站的鱼类替代生境[24-25]; 澜沧江梯级水电开发预计将对多达11条支流进行生境保护[17]。此外, 高婷[18]、杨青瑞[19]等对支流生境替代基础理论也进行了研究, 探寻了基本原则, 构建了保护效果评价体系; 林俊强等[26]应用模糊相似理论, 提出基于恒定型相似元、时间型相似元和系统相似度的河流生境相似性计算方法, 可为支流替代生境的筛选与最终确定提供参考依据。但是, 针对梯级水电开发下的生境替代支流发挥怎样的保护效果以及干流对其的影响仍有待深入研究, 尤其对于澜沧江这样的国际河流[27]。
澜沧江位于我国西南边陲, 出境后称为湄公河(Mekong River), 干流全长2129 km, 天然落差4583 m, 水能蕴藏量巨大, 迄今为止已建成并运行了11座梯级电站。因其跨境河流的特殊性, 梯级开发所带来的生态环境问题长期受到国内外广泛关注, 尤其是对鱼类资源的影响更是下游湄公河次区域国家关注的焦点。针对澜沧江高坝大库鱼类保护的特殊难题, 在开发过程中探索性采用了支流生境补偿措施, 其中比较典型的包括一级支流罗梭江和基独河。罗梭江又名补远江, 发源于普洱市宁洱县, 主河道长297.8 km, 集水面积7678.9 km2[17, 28]。2007年4月, 罗梭江鱼类自然保护区被批准建立, 通过拆除鱼坝、取消规划电站、设置管理站和增殖中心等多种措施改善了鱼类栖息环境。基独河为较小支流, 发源于大理市兰坪县, 主河道长41.5 km, 集水面积238.0 km2[29]。2012年, 基独河水坝被拆除, 河流连通性得到修复。
本研究选取澜沧江及其一级支流罗梭江、基独河为研究对象, 以各自邻近的南阿河、丰甸河作为对照, 在收集整理历史资料的基础上, 结合现场调查结果, 对澜沧江及其四条一级支流鱼类种类组成、群落结构、生态类型进行对比研究, 并试图阐明梯级开发下鱼类支流生境替代效果以及干流工程建设对其的影响, 以期为水电开发河流鱼类保护提供重要支撑。
1 材料与方法梯级开发后, 澜沧江河流形态由河相转为河相-湖相交替形式。由于2016年新建苗尾大坝的回水影响, 基独河汇入处干流断面平均流速小于0.5 m/s, 成为干流“湖相”下生境替代支流, 相对而言, 罗梭江为干流“河相”下生境替代支流, 其汇入处干流不受回水影响。为分析生境替代支流对鱼类保护效果, 分别以对岸未进行生境修复支流的南阿河和丰甸河为对照, 并设置汇入处干流江段进行关联分析(为避免命名混乱, 罗梭江、南阿河汇入处干流江段命名为下游关联干流, 基独河、丰甸河汇入处干流江段命名为上游关联干流)。对照支流均受人类活动影响较大, 其中南阿河沿岸开垦有大型农场, 河道中修建挡水坝引流灌溉; 而丰甸河运行有两级低水头坝, 河流生境处于破碎化。前期研究表明, 罗梭江与南阿河, 基独河与丰甸河在鱼类区系分别具有类似组成, 且各支流与关联干流鱼类均呈嵌套子集关系[17, 28-29]。
1.1 数据收集鱼类历史数据收集于已公开发表的论文[28-29]与专业报告[30], 并依据《云南鱼类名录》[31]进行同物异名的一致性修订。鱼类现状数据收集以渔获物调查方式进行, 为突出比较分析法的单因子变量原则, 严格控制调查时间的同期性和采样方法的一致性。参考《内陆水域渔业自然资源调查手册》[32]及河流水域分布, 共设置鱼类采样点18个(图 1), 分别为下游关联干流:LC1、LC2、LC3;上游关联干流:LC4、LC5;罗梭江:LS1、LS2、LS3;南阿河:NA1、NA2、NA3;基独河:JD1、JD2、JD3、JD4;丰甸河:FD1、FD2、FD3。基独河、丰甸河及上游关联干流为冷水性鱼类, 而罗梭江、南阿河及下游关联干流鱼类为暖水性鱼类[33], 两个区域鱼类在生态习性上截然不同。因此, 为了对两个区域鱼类进行科学的对比分析, 本研究在采样时间上着重考虑了鱼类生活史上的同期性, 分别选择鱼类在干支流活动频繁的洄游、产卵季节, 即罗梭江、南阿河、下游关联干流为2019年3-4月[28], 基独河、丰甸河、上游关联干流为2019年9-10月[34]。

|
| 图 1 研究区域及鱼类采样河段示意图 Fig. 1 The location of study area and sampling sites FD:丰甸河Fengdian River; JD:基独河Jidu River; LC:澜沧江Lancang River; NA:南阿河Naa River; LS:罗梭江Luosuo River; FD1-FD3、JD1-JD4、LC1-LC5、NA1-NA3、LS1-LS3:采样河段Sampling sites |
为了捕获到不同水层及不同大小的鱼类, 采用多种规格渔具及渔法。罗梭江、南阿河及各干流河段以定制刺网、撒网为主, 辅以饵钩、地笼等, 尺寸大小分别为:刺网网目3-15 cm、网高2-15 m、网长20-150 m, 撒网网目2 cm、高4 m、直径6 m, 饵钩长度300 m, 地笼长宽高分别为20 m、50 cm、50 cm。采集为全天进行, 每次放置刺网10张、地笼2个、饵钩一条, 早、晚各收集一次; 此外撒网2副, 每天傍晚作业2 h。基独河、丰甸河因属于高落差山区河流, 水流湍急、河床粗糙, 为保证采样数据的准确性, 采用2人组背负式电捕鱼器(电瓶:20 A, 12 V; 电鱼器:3000 W)采样, 每日一次, 每个样点采样长度为500 m。每个采样江段采样时间为3-5 d, 采集到的鱼类现场拍照后, 进行分类鉴定及生物学测定, 其中濒危鱼类、洄游鱼类、土著鱼类及特有鱼类分别依据《中国濒危动物红皮书(鱼类)》[35]、《云南湿地》[33]及相关文献[36]进行判定。由于本研究区域以中下层鱼类为主, 因此水样采集于水体中下层。使用哈希(HQ40D, USA, Hash)便携式水质分析仪现场测定水样的温度、溶解氧及酸碱度(pH), 采用碱性过硫酸钾消解紫外分光光度法(GB11894-89)、钼酸铵分光光度法(GB11893-89)实验室分别测定水体总氮、总磷指标。
1.2 数据处理与分析 1.2.1 优势种采用相对重要性指数(IRI)分析鱼类群落优势种[37-38]:
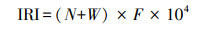
|
(1) |
式中, N和W分别为每种鱼占所捕总量的数量百分比和重量百分比; F为出现频率, 即某一种鱼出现的样点数占所有采样点数的比例。一般认为IRI大于1000时为优势种, 大于2000时为显著优势种[39]。
1.2.2 群落相似性基于鱼类的种类丰度, 以Bray-Curtis相似性系数为基础构建不同采样河段的相似性矩阵, 为了消除丰度极端值的影响, 在分析前对所有丰度值进行log(x+1)转化[40]。采用平均聚类(Cluster analysis, CA)和非度量多维标度排序方法(Non-metric multidimensional scaling, NMDS)分析各监测河段鱼类群落结构差异, 进一步地使用单因素相似性分析(one-way analysis of similarities, one-way ANOSIM)对CA结果进行差异检验[41-42]。NMDS分析结果的优劣采用胁强系数来检验[43]:当胁强系数(Stress) < 0.2时, 表示图形有一定的解释意义; 胁强系数 < 0.1时, 是一个好的排序; 胁强系数 < 0.05时, 具有很好的代表性。以上分析与绘图均在PRIMER 7.0中进行。
1.2.3 群落生态类型基于鱼个体生态学指数, 以鱼类生物量占比计算群落对环境的需求[44-45]:
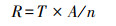
|
(2) |
式中, T为鱼类群落矩阵; A为鱼类个体生态类型矩阵; n为鱼类种类数; R为鱼类群落对环境需求因子的需求度。鱼类个体生态学矩阵依据流速偏好、水深偏好、底质偏好、产卵类型、营养类型以及洄游习性等18个生态指标的建立, 偏好数据主要来自在线数据库(www.fishbase.org)、专业书籍[46-48]以及相关期刊论文[49-50], 当某种鱼类某些生态特征不明确时, 则在矩阵中取平均值。
2 结果与分析 2.1 水环境调查期间, 各采样河流水体pH、水温、溶解氧、总氮、总磷如图 2所示。罗梭江除总氮、总磷指标低于南阿河外, 其余水环境指标类似。其中罗梭江总氮、总磷分别为(0.548±0.042) mg/L、(0.023±0.009) mg/L, 近似Ⅱ类水, 与干流接近; 而南阿河虽然总磷为Ⅱ类水标准, 但总氮达到(1.301±0.234) mg/L, 为Ⅳ类水。基独河水体温度与干流接近, 而高于丰甸河, 其余水环境参数二者均与干流类似, 其特征是碱性、高溶解氧和低营养盐。

|
| 图 2 研究区域水环境指标 Fig. 2 Water environment indexes in the study area |
罗梭江、南阿河及下游关联干流共统计渔获物510.58 kg, 7899尾, 鉴定出种类80种, 隶属于5目21科61属。其中下游关联干流种类数最多, 有5目15科48属55种, 占物种总数的68.8%;罗梭江次之, 有3目16科39属51种, 占63.8%;南阿河相对较低, 有3目14科34属41种, 占51.3%(附表 1)。总体而言, 罗梭江各河段物种丰富度与下游关联干流接近, 却高于南阿河(图 3)。

|
| 图 3 采样河段鱼类种类组成 Fig. 3 Fish species composition in the sampling sites |
基独河、丰甸河及上游关联干流共统计渔获物61.74kg, 1522尾, 鉴定出种类19种, 隶属于3目9科18属。种类数同样以干流最多, 保护支流次之, 对照支流相对较低, 但三者数量差距不大, 分别为15种、13种和11种。支流各河段仅邻近河口段(JD1、FD1)物种数与干流接近, 其余均低于干流(图 3)。
鱼类区系组成在各调查河段大致相同, 以鲤形目、鲇形目、鲈形目为主, 其余魟形目、脂鲤目仅在LC1江段各有1种(图 3)。而河流优势种则在保护支流、对照支流、关联干流之间差异显著(表 1)。如罗梭江以云南吻孔鲃、长臀鲃属等土著鱼类为主, 而南阿河以入侵鱼类下口鲇和小型鱼类南鳅属为主; 基独河、丰甸河鱼类以条鳅科、鮡科为主, 而其关联干流鱼类则以裂腹鱼属为主。
| 河流 Rivers |
鱼类相对重要性指数/IRI The Relative Importance Index of Fish |
| 罗梭江Luosuo River | 云南吻孔鲃(4527.2)、长臀鲃(2294.5)、细尾长臀鲃(1656.4)、花[鱼骨](886.2)、高体鳑鲏(261.3) |
| 南阿河Naa River | 下口鲇(3537.3)、大头南鳅(2173.1)、湄南南鳅(847.3)、丽色低线鱲(620.6)、长臀鲃(536.9) |
| 下游关联干流 Associated down-main stream |
中华鲱鲇(1926.9)、野结鱼(1225.3)、云南吻孔鲃(719.1)、高体鳑鲏(532.1)、尼罗罗非鱼(486.7) |
| 基独河Jidu River | 拟鳗荷马条鳅(13450.8)、长须纹胸鮡(2126.2)、短尾高原鳅(185.0)、麦穗鱼(79.0)、长腹华沙鳅(40.7) |
| 丰甸河Fengdian River | 长须纹胸鮡(1413.8)、短尾高原鳅(1364.7)、麦穗鱼(1338.6)、泥鳅(760.3)、拟鳗荷马条鳅(480.4) |
| 上游关联干流 Associated Up-mian Stream |
光唇裂腹鱼(8128.2)、澜沧裂腹鱼(5644.1)、张氏间吸鳅(1738.5)、鲤鱼(456.1)、麦穗鱼(182.5) |
| IRI:相对重要性指数The Relative Importance Index | |
本次调查到濒危鱼类4种, 分别为双孔鱼、短须粒鲇、线足鲈、湄南细丝鲇。调查到的澜沧江特有鱼类共28种, 采集到特有鱼类种类数最多的是罗梭江, 为16种; 其次为南阿河与下游关联干流, 均为12种; 基独河、丰甸河及上游关联干流相对较少, 分别为3种、2种、5种(附表 1)。具有洄游习性鱼类共采集到26种, 采集到洄游鱼类种类数最多的是下游关联干流, 为23种; 其次为罗梭江与南阿河, 分别为15种、10种; 而基独河、丰甸河和及上游关联干流采集到的鱼类均为定居习性。
2.4 保护鱼类物种数变化2007年自然鱼类保护区建立后, 罗梭江保护鱼类物种数呈现上升趋势。其中土著鱼类由30种增长为42种, 特有鱼类由9种增长为16种, 洄游鱼类由10种增长为15种(图 4), 濒危鱼类保持1种不变, 但2006年3-4月为魾、2010年4月和8月的为裂峡鲃, 本次调查的为双孔鱼。较保护区建立前, 本次新增的土著鱼类有:大斑纹胸鮡、滨河缺鳍鲇、异斑小鲃、长体间吸鳅、缺须墨头鱼、伯氏似鳞头鳅、斑尾墨头鱼、大头南鳅、异颌南鳅、斑鳍连穗沙鳅、大鳞半餐、鲮鱼、澜沧江爬鳅、彭氏间吸鳅、克氏南鳅、双孔鱼、鲤; 减少的有魾、柬埔寨墨头鱼、长嘴鱲、黑线安巴沙鳅、南方翅条鳅。

|
| 图 4 罗梭江和基独河保护鱼类物种数历史变化 Fig. 4 Historical changes of the protected fish species in the Luosuo River and Jidu River |
基独河在拆坝后的第一年, 土著鱼类物种数就迅速增加, 2012年9月监测为6种, 而后增长为10种, 2014年9月与2015年9月均为13种, 至2019年9月, 监测的物种数出现减少现象, 为10种(图 4), 但特有鱼类数量一直保持上升趋势, 由1种增长为3种。相较于拆坝前, 至2015年9月增加的种类有:后背鲈鲤、鲤鱼、奇额墨头鱼、长腹华沙鳅、穗缘异齿鰋、长胸异鮡、长须纹胸鮡, 而2019年9月较前者减少的种类有:后背鲈鲤、鲤鱼、奇额墨头鱼、穗缘异齿鰋、长胸异鮡, 新增的有无斑褶鮡、澜沧裂腹鱼。
2.5 群落结构罗梭江、南阿河及下游关联干流采样河段鱼类群落可以分为两大类, 其中LC1、LC2、LC3、LS1、LS2、LS3为一个相似类群, 而NA1、NA2、NA3为另一个相似类群, 两大类群Bray-Curtis相似性为21.76%(图 4), One-way ANOSIM检验显示这两个类群的群落结构在统计学上差异显著(r=1.00, P=0.1% < 0.05)。NMDS图则表示罗梭江鱼类群落与干流下游LC1河段差异程度较小, 而与景洪坝下的LC2、LC3河段差异程度较大(Stress=0.08 < 0.1)(图 5)。

|
| 图 5 罗梭江、南阿河及下游关联干流各河段鱼类群落CA及NMDS图 Fig. 5 CA and NMDS maps of fish community structures in the sampling sites of Luosuo River、Naa River and its down-main stream CA:平均聚类Cluster analysis; NMDS:非度量多维标度排序Non-metric multidimensional scaling |
剔除丰甸河无鱼河段FD2、FD3, 基独河、丰甸河、上游关联干流各采样河段鱼类群落也可划分为两个相似类群:类群Ⅰ包括LC4、LC5、FD1、JD1;类群Ⅱ包括JD2、JD3、JD4(图 6)。两大类群Bray-Curtis相似性仅为10.73%, One-way ANOSIM检验显示这两个类群的群落结构在统计学上差异显著(r=0.99, P=0.1% < 0.05)。NMDS图则表示了干流鱼类群落与各支流靠近河口段(JD4、FD3)差异程度较小, 而与支流上游河段差异程度较大, (Stress=0.01 < 0.05)。

|
| 图 6 基独河、丰甸河及上游关联干流各河段鱼类群落CA及NMDS图 Fig. 6 CA and NMDS maps of fish community structures in the sampling sites of Jidu River、Fengdian River and its up-mian stream |
罗梭江、南阿河及下游关联干流的鱼类群落均是以杂食性(0.75 < R < 0.90)、缓流型(0.54 < R < 0.78), 产粘性及粘沉性卵(0.69 < R < 0.76)、喜沙砾\\卵石底质(0.41 < R < 0.44)的鱼类为主。具有差异的是, 罗梭江与下游关联干流的鱼类是以中下层栖息(0.67 < R < 0.74)为主, 而南阿河以底层栖息(R=0.78)为主; 支流鱼类以定居性(0.69 < R < 0.82)为主, 而干流以半洄游性(R=0.73)为主。
基独河、丰甸河及上游关联干流的鱼类物种均是以产粘沉性卵(0.56 < R < 0.95)、底层栖息(0.55 < R < 0.97)、喜沙砾\\卵石底质(0.48 < R < 0.54)、生活在急流型水体(0.56 < R < 0.96)、具有定居习性(R=1.00)的鱼类为主(图 7)。具有差异的是, 丰甸河及上游关联干流的鱼类是以杂食性(0.53 < R < 0.97)为主, 而基独河是以肉食性(R=0.92)为主。

|
| 图 7 鱼类群落环境需求度 Fig. 7 Life history requirements of fish community |
水电梯级开发改变了河流水文情势, 将河流形态由“河相”转变为“河相-湖相交替”形式, 对其鱼类造成了潜在影响。研究表明, 澜沧江梯级开发以来土著鱼类、洄游鱼类、特有鱼类种群规模不断减少、物种数大量丧失[36, 51-52], 一些濒危鱼类由于所需生境的特殊性, 更是逐渐走向灭绝。然而, 与之相对的是罗梭江河流生境修复后, 土著鱼类、特有鱼类不断增加, 分别由30种增加至42中, 9种增长为16种(图 4), 甚至栖息于此的不乏有中国结鱼、中华鲱鲇、双孔鱼、大鳞半餐、滨河缺鳍鲇、大斑纹胸鮡、澜沧江爬鳅、彭氏间吸鳅、斑尾墨头鱼、斑鳍连穗沙鳅等这些在干流开始变得稀少[36, 51]的物种。同样基独河生境修复后的第一年内, 其鱼类物种丰度、密度、多样性也迅速增加[29], 2013年9月监测到栖息于基独河的干流土著鱼类6种, 2015年9月上升到13种(图 4)。另一方面, 通过鱼类资源现状对比发现, 生境修复的支流鱼类物种丰度和群落数量均明显高于对照支流, 如罗梭江物种数甚至与干流接近(图 3), 这意味着生境修复后的支流变得更适宜鱼类栖息, 支流替代生境建设对鱼类具有显著保护效果。
值得注意的是, 罗梭江新增种类除大鳞半餐、异斑小鲃等少数中上层鱼类外, 其余均为急流底栖鱼类, 且以肉食性为主, 如双孔鱼、斑鳍连穗沙鳅、异颌南鳅、长体间吸鳅、彭氏间吸鳅、澜沧江爬鳅、大斑纹胸鮡等。这主要是因为肉食性底栖鱼类以昆虫幼虫、软体动物或其他大型、微型无脊椎底栖动物为食[53], 而这些底栖生物通常受到沉积物粒度、栖息地组成和时空异质性变化的影响[54]。罗梭江汇入口上游于2009年修建大坝, 干流水体透明度和水文节律显著变化, 致使底栖动物种类及生物量下降。在此背景下, 底栖肉食性鱼类出现食物资源衰减、栖息地质量下降的情况[55], 它们更倾向于进行避难迁徙, 以寻求合适的替代生境。
3.2 干流环境条件变化对支流生境替代效果的影响与罗梭江土著鱼种类逐渐增多截然相反, 本次调查基独河土著鱼类较2015年9月监测减少了5种, 分别为鲤鱼以及后背鲈鲤、奇额墨头鱼、穗缘异齿鰋、长胸异鮡这些急流型鱼类。研究表明, 对较小支流而言, 鱼类群落结构的稳定依赖于干流鱼类向支流的资源补充效应[25, 29]。而本次调查期间, 干流苗尾电站已于2016年蓄水运行, 基独河汇入干流的河口区成为了“湖相”状态; 又加之鱼类对于生存区域的高度选择性, 可以主动规避不利的空间和栖息环境[56], 即急流型鱼类往往本能地避开静水环境而自主选择流水栖息。由此, “湖相”状态致使干流中原急流型的鱼类被大量“驱离”至库尾, 如本次在库尾河相段(LC5)就采集到大量张氏间吸鳅、奇额墨头鱼、裂腹鱼等急流型鱼类。这种现象也发生在金沙江下游河段, 向家坝电站的修建使得鱼类群落出现河湖分区, 流水性鱼类大量聚集库尾[45]。基独河在失去干流对这些鱼类的资源补充后, 其鱼类多样性也随之下降。另一方面, 通过与丰甸河的对比分析发现, 虽然目前栖息于基独河各河段的鱼类物种数均相对较多(图 3), 但就种群规模而言, 基独河可能仅对拟鳗荷马条鳅、长须纹胸鮡有着保护效果, 因为调查期间采集到鱼类数量, 除二者较多以外(分别为779尾和263尾), 其余种类采集量均低于10尾, 而在干流占据群落绝对优势的裂腹鱼属采集量更是只有5尾幼稚鱼。NMDS图也显示干流鱼类群落与支流相似度低于10%, 且与基独河下游河段的相似度显著高于其上游河段(图 6), 这表明基独河上游河段并不适合干流鱼类栖息。可以预见, 干流“湖相”下的基独河正处于一个封闭狭小的替代生境状态, 加之与干流低相似性的鱼类群落, 其未来对干流鱼类的保护效果日趋微小。
研究表明, 许多鱼类具有向干流或支流的某些河段进行产卵迁徙或索饵迁徙的特征[57-58], 这些特征也将导致位于迁徙路线上的河段鱼类在洄游季节具有更高的群落相似性。在本研究中, 通过与南阿河的对比发现, 罗梭江与干流鱼类群落具有更明显的聚类现象和更高的相似性(图 5), 这表明罗梭江的干支流鱼类迁徙更为频繁, 其更适宜干流鱼类栖息; 此外, NMDS图还显示澜沧江下游河段(LC1)的鱼类群落相较于景洪坝下河段(LC2、LC3), 与罗梭江相似性更高(图 3), 考虑到澜沧江现有鱼类洄游均为上溯性洄游[59-60], 这意味着罗梭江或已成为澜沧江洄游鱼类繁殖或觅食的关键栖息地。需要注意的是, 鱼类迁徙活动除需保持洄游路线联通之外, 其洄游过程更需要流速引导[60], 若处于洄游通道的干流失去“河相”状态, 罗梭江对干流鱼类的替代保护效果也必将大幅降低。
3.3 鱼类支流替代生境建设的建议自2012年环保部明确提出“开展干流和支流开发与保护”生态补偿试点以来, 支流生境修复举措在我国水电开发中被广泛应用。在本研究中, 通过对生境修复与未修复支流的对比分析, 发现支流生境修复可为干流土著鱼类提供有效的替代生境。但此类保护措施的效果持续性也存在一个前提条件, 即关联干流需要保持“河相”, 特别是对于存在大量洄游性鱼类的河段。同样, 对于目前基独河替代效果的下降趋势, 也可以通过开展生态调度使关联干流保持“河相”来改善。
支流生境替代作为补偿性保护措施, 同样需要以经济代价与保护效应合理为基础[18], 一些支流生境修复可能耗资巨大, 譬如需要拆除支流水电站等[24, 29]。因此需要避免盲目清除支流小水电, 因为即使清除“湖相区”支流的小水电, 鱼类保护效果可能也一般, 投入-效益比较低。因此, 本研究建议今后开展此类保护措施应优先选择库尾河相区支流。
4 结论以澜沧江生境替代支流罗梭江(河相干流)和基独河(湖相干流)为研究对象, 并设置对照组, 通过鱼类种类组成与群落结构的差异分析以及历史数据比较, 阐明了澜沧江支流生境替代建设对鱼类的保护成效, 并初步揭示了干流对支流生境替代保护效果的影响机制。主要结论如下:
(1) 罗梭江和基独河作为支流替代生境对干流澜沧江的鱼类具有保护效果, 支流生境修复后鱼类多样性明显提高, 替代生境能为澜沧江土著鱼类甚至一些濒危特有及洄游鱼类提供完成生活史的关键栖息地。
(2) 干流环境条件变化对支流替代生境的保护效果具有重要影响, 罗梭江替代生境因澜沧江干流建坝而出现对底栖肉食性鱼类的偏向性, 基独河替代生境因河口段干流成为“湖相”使得替代效果出现下降趋势。
(3) 支流河口段的干流保持“河相”是支流生境替代保护效果可持续性的关键, 建议今后开展此类保护措施时优先选择河相区支流。
| 种类 Fish Species |
罗梭江 Luosuo River |
南阿河 Naa River |
下游关联干流 Associated down main-stream |
基独河 Jidu River |
丰甸河 Fengdian River |
上游关联干流 Associated up mian-stream |
|||||||||||||||||
| LS1 | LS2 | LS3 | NA1 | NA2 | NA3 | LC1 | LC2 | LC3 | JD1 | JD2 | JD3 | JD4 | FD1 | FD2 | FD3 | LC4 | LC5 | ||||||
| I魟形目MYLIOBATIFORMES | |||||||||||||||||||||||
| (1) 魟科Dasyatidae | |||||||||||||||||||||||
| 1老挝魟Dasyatis laosensis | 1 | ||||||||||||||||||||||
| II鲤形目CYPRINIFORMES | |||||||||||||||||||||||
| (2) 双孔鱼科Gyrinocheilidae | |||||||||||||||||||||||
| 2双孔鱼Gyrinocheilus aymonieri* | 1 | 1 | |||||||||||||||||||||
| (3) 沙鳅科Botiidae | |||||||||||||||||||||||
| 3长腹华沙鳅Sinibotia longiventralis | 1 | 1 | |||||||||||||||||||||
| 4斑鳍连穗沙鳅Syncrossus beauforti* | 1 | 1 | 1 | ||||||||||||||||||||
| (4) 鳅科Cobitidae | |||||||||||||||||||||||
| 5伯氏似鳞头鳅Lepidocephalichthys berdmorei | 1 | 1 | 1 | ||||||||||||||||||||
| 6泥鳅Misgurnus anguillicaudatus | 1 | 1 | 1 | ||||||||||||||||||||
| 7大鳞副泥鳅Paramisgurnus dabryanus▲ | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||
| (5) 爬鳅科Balitoridae | |||||||||||||||||||||||
| 8澜沧江爬鳅Balitora lancangjiangensis* | 1 | 1 | |||||||||||||||||||||
| 9长体间吸鳅Hemimyzon elongatus* | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||
| 10彭氏间吸鳅Hemimyzon pengi* | 1 | 1 | 1 | 1 | |||||||||||||||||||
| 11张氏间吸鳅Hemimyzon tchangi* | 1 | 1 | 1 | 1 | |||||||||||||||||||
| (6) 条鳅科Nemacheilidae | |||||||||||||||||||||||
| 12拟鳗荷马条鳅Homatula anguillioides | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||
| 13南方翅条鳅Pteronemacheilus meridionalis | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||
| 14鼓颊南鳅Schistura bucculenta* | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||
| 15横纹南鳅Schistura fasciolata | 1 | 1 | 1 | ||||||||||||||||||||
| 16湄南南鳅Schistura kengtungensis | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||
| 17克氏南鳅Schistura kloetzliae | 1 | ||||||||||||||||||||||
| 18宽纹南鳅Schistura latifasciata* | 1 | 1 | |||||||||||||||||||||
| 19大头南鳅Schistura macrocephalus | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||
| 20异颌棱唇条鳅Sectoria heterognathos* | 1 | ||||||||||||||||||||||
| 21短尾高原鳅Triplophysa brevicauda | 1 | 1 | 1 | ||||||||||||||||||||
| (7) 鲤科Cyprinidae | |||||||||||||||||||||||
| 22金线[鱼丹]Danio chrysotaeniatus* | 1 | ||||||||||||||||||||||
| 23金长须[鱼丹]Esomus metallicus | 1 | ||||||||||||||||||||||
| 24玫瑰短[鱼丹]Brachydanio rosea | 1 | 1 | |||||||||||||||||||||
| 25丽色低线鱲Barilius pulchellus | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||
| 26马口鱼Opsariichthys bidens | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||
| 27高体鳑鲏Rhodeus ocellatus▲ | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||
| 28罗碧鱼Paralaubuca barroni* | 1 | 1 | 1 | ||||||||||||||||||||
| 29大鳞半Hemiculterella macrolepis* | 1 | ||||||||||||||||||||||
| 30Hemiculter leucisculus▲ | 1 | 1 | 1 | ||||||||||||||||||||
| 31团头鲂Megalobrama amblycephala▲ | 1 | ||||||||||||||||||||||
| 32红鳍原鲌Cultrichthys erythropterus▲ | 1 | 1 | 1 | ||||||||||||||||||||
| 33花鱼骨Hemibarbus maculatus | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||
| 34麦穗鱼Pseudorasbora parva▲ | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||
| 35棒花鱼Abbotina rivularis▲ | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||
| 36草鱼Ctenopharyngodon idella▲ | 1 | 1 | |||||||||||||||||||||
| 37中国结鱼Tor sinensis* | 1 | 1 | 1 | ||||||||||||||||||||
| 38野结鱼Tor tambra | 1 | 1 | 1 | 1 | |||||||||||||||||||
| 39异斑小鲃Puntius ticto | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||
| 40南方白甲鱼Onychostoma gerlachi | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||
| 41少鳞舟齿鱼Scaphiodonichthys acanthopterus | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||
| 42南腊方口鲃Cosmochilus nanlaensis* | 1 | ||||||||||||||||||||||
| 43爪哇鲃鲤Puntioplites waandersi | 1 | ||||||||||||||||||||||
| 44云南吻孔鲃Poropuntius huangchuchieni | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||
| 45爪哇无名鲃Barbonymus gonionotus | 1 | ||||||||||||||||||||||
| 46黄尾短吻鱼Sikukia flavicaudata* | 1 | 1 | 1 | ||||||||||||||||||||
| 47短吻鱼Sikukia gudgeri* | 1 | 1 | 1 | 1 | |||||||||||||||||||
| 48长须短吻鱼Sikukia longibarbata* | 1 | ||||||||||||||||||||||
| 49细尾长臀鲃Mystacoleucus lepturus* | 1 | 1 | 1 | ||||||||||||||||||||
| 50长臀鲃Mystacoleucus marginatus* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||
| 51大鳞高须鱼Hypsibarbus vernayi | 1 | 1 | 1 | ||||||||||||||||||||
| 52脂孟加拉鲮Bangana lippa | 1 | 1 | |||||||||||||||||||||
| 53麦瑞加拉鲮Cirrhinus mrigala▲ | 1 | 1 | |||||||||||||||||||||
| 54长背鲃Labiobarbus leptocheila* | 1 | 1 | |||||||||||||||||||||
| 55鲮Cirrhinus molitorella | 1 | ||||||||||||||||||||||
| 56斑尾墨头鱼Garra fasciacauda | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||
| 57缺须墨头鱼Garra imberbaimberba | 1 | 1 | 1 | ||||||||||||||||||||
| 58奇额墨头鱼Garra mirofronits* | 1 | 1 | |||||||||||||||||||||
| 59澜沧裂腹鱼Schizothorax lantsangensis* | 1 | 1 | 1 | 1 | |||||||||||||||||||
| 60光唇裂腹鱼Schizothorax lissolabiatus | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||
| 61鲤Cyprinus carpio | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||
| 62鲫Carassius auratus | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||
| III脂鲤目CHARACIFORMES | |||||||||||||||||||||||
| (8) 脂鲤科Characidae | |||||||||||||||||||||||
| 63短盖肥脂鲤Piaractus brachypomus▲ | 1 | ||||||||||||||||||||||
| IV鲇形目SILURIFORMES | |||||||||||||||||||||||
| (9)甲鲇科Loricariidae | |||||||||||||||||||||||
| 64下口鲇Hypostomus plecostomus▲ | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||
| (10)鲿科Bagridae | |||||||||||||||||||||||
| 65黄颡鱼Pelteobagrus fulvidraco▲ | 1 | ||||||||||||||||||||||
| 66瓦氏黄颡鱼Pelteobagrus vachelli▲ | 1 | 1 | 1 | ||||||||||||||||||||
| 67丝尾鳠Hemibagrus wyckioides | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||
| (11)鲇科Siluridae | |||||||||||||||||||||||
| 68鲇Silurus asotus | 1 | ||||||||||||||||||||||
| 69湄南细丝鲇Micronema moorei | 1 | ||||||||||||||||||||||
| 70滨河亮背鲇Phalacronotus bleekeri | 1 | 1 | 1 | ||||||||||||||||||||
| 71叉尾鲇Wallago attu | 1 | ||||||||||||||||||||||
| (12)锡伯鲇科Schilbidae | |||||||||||||||||||||||
| 72中华鲱鲇Clupisoma sinense* | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||
| (13) 鮰科Ictaluridae | |||||||||||||||||||||||
| 73斑点叉尾鮰Ictalurus punctatus▲ | 1 | ||||||||||||||||||||||
| (14) 鮡科Sisoridae | |||||||||||||||||||||||
| 74巨魾Bagarius yarrelli | 1 | 1 | 1 | 1 | |||||||||||||||||||
| 75纺锤纹胸鮡Glyptothorax fuscus | 1 | 1 | 1 | 1 | |||||||||||||||||||
| 76老挝纹胸鮡Glyptothorax laosensis* | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||
| 77大斑纹胸鮡Glyptothorax macromaculatus* | 1 | 1 | 1 | ||||||||||||||||||||
| 78长须纹胸鮡Glyptothorax longinema | 1 | 1 | 1 | 1 | |||||||||||||||||||
| 79无斑褶鮡Pseudecheneis immaculatus* | 1 | 1 | |||||||||||||||||||||
| 80细尾鮡Pareuchiloglanis gracilicaudata* | 1 | ||||||||||||||||||||||
| (15) 粒鲇科Akysidae | |||||||||||||||||||||||
| 81短须粒鲇Akysis brachybarbatus* | 1 | 1 | |||||||||||||||||||||
| (16) 刺鳅科Mastacebelidae | |||||||||||||||||||||||
| 82大刺鳅Mastacembelus armatus | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||
| V鲈形目PERCIFORMES | |||||||||||||||||||||||
| (17) 丽鱼科Cichlidae | |||||||||||||||||||||||
| 83莫桑比克罗非鱼Oreochromis mossambica▲ | 1 | 1 | |||||||||||||||||||||
| 84尼罗罗非鱼Oreochromis nilotica▲ | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||
| 85奥利亚罗非鱼Oreochromis aureus▲ | 1 | ||||||||||||||||||||||
| (18) 沙塘鳢科Odontobutidae | |||||||||||||||||||||||
| 86小黄[鱼幼]鱼Micropercops swinhonis▲ | 1 | 1 | |||||||||||||||||||||
| (19) 鰕虎鱼科Gobiidae | |||||||||||||||||||||||
| 87子陵吻鰕虎鱼Rhinogobius giurinus▲ | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||
| (20) 斗鱼科Belontiidae | |||||||||||||||||||||||
| 88线足鲈Trichogaster trichopterus | 1 | ||||||||||||||||||||||
| (21) 鳢科Channidae | |||||||||||||||||||||||
| 89宽额鳢Channa gachua | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||
| 90线鳢Channa striata | 1 | 1 | 1 | 1 | |||||||||||||||||||
| (22) 塘鳢科Eleotridae | |||||||||||||||||||||||
| 91云斑尖塘鳢Oxyeleotris marmorata▲ | 1 | 1 | 1 | ||||||||||||||||||||
| 种类数合计 | 34 | 33 | 35 | 23 | 27 | 26 | 36 | 31 | 32 | 13 | 3 | 1 | 1 | 11 | 0 | 0 | 10 | 11 | |||||
| “1”代表采集到鱼类; “*”代表特有鱼类, “▲”代表外来鱼类。FD:丰甸河Fengdian River; JD:基独河Jidu River; LC:澜沧江Lancang River; NA:南阿河Naa River; LS:罗梭江Luosuo River | |||||||||||||||||||||||
| [1] |
Agostinho A A, Gomes L C, Santos N C L, Ortega J C G, Pelicice F M. Fish assemblages in Neotropical reservoirs: colonization patterns, impacts and management. Fisheries Research, 2016, 173: 26-36. DOI:10.1016/j.fishres.2015.04.006 |
| [2] |
Liermann C R, Nilsson C, Robertson J, Ng R Y. Implications of dam obstruction for global freshwater fish diversity. BioScience, 2012, 62(6): 539-548. DOI:10.1525/bio.2012.62.6.5 |
| [3] |
Best J. Anthropogenic stresses on the world's big rivers. Nature Geoscience, 2019, 12(1): 7-21. DOI:10.1038/s41561-018-0262-x |
| [4] |
谢平. 长江的生物多样性危机--水利工程是祸首, 酷渔乱捕是帮凶. 湖泊科学, 2017, 29(6): 1279-1299. |
| [5] |
Bao J H, Li W W, Zhang C S, Mi X Y, Li H T, Zhao X J, Cao N, Twardek W M, Cooke S J, Duan M. Quantitative assessment of fish passage efficiency at a vertical-slot fishway on the Daduhe River in Southwest China. Ecological Engineering, 2019, 141: 105597. DOI:10.1016/j.ecoleng.2019.105597 |
| [6] |
Mulligan K B, Haro A, Towler B, Sojkowski B, Noreika J. Fishway entrance gate experiments with adult American shad. Water Resources Research, 2019, 55(12): 10839-10855. DOI:10.1029/2018WR024400 |
| [7] |
王继隆, 刘伟, 杨文波, 李培伦, 王维坤. 绥芬河大麻哈鱼增殖放流效果评估. 应用生态学报, 2019, 30(10): 3572-3578. |
| [8] |
Milot E, Perrier C, Papillon L, Papillon L, Dodson J J, Bernatchez L. Reduced fitness of Atlantic salmon released in the wild after one generation of captive breeding. Evolutionary Applications, 2013, 6(3): 472-485. DOI:10.1111/eva.12028 |
| [9] |
Jiang Z Q, Liu P, Ji C M, Zhang H R, Chen Y Y. Ecological flow considered multi-objective storage energy operation chart optimization of large-scale mixed reservoirs. Journal of Hydrology, 2019, 577: 123949. DOI:10.1016/j.jhydrol.2019.123949 |
| [10] |
匡亮, 张鹏, 杨洪雨, 梁瑞峰, 王远铭, 李克锋. 梯级水库叠梁门分层取水水温改善效果的衰减. 长江流域资源与环境, 2019, 28(5): 1244-1251. |
| [11] |
党莉, 马超, 练继建. 水库调节对下游鱼类栖息地适宜性的影响. 天津大学学报: 自然科学与工程技术版, 2018, 51(6): 566-574. DOI:10.3969/j.issn.1008-4339.2018.06.012 |
| [12] |
严灿, 胡加琴, 路波, 刘国勇, 胡晓, 王继保, 石小涛. 水电背景的鱼类救护工程与投资回报: 中国鱼类资源前景堪忧. 长江流域资源与环境, 2016, 25(S1): 44-49. |
| [13] |
王超. "西南水电高坝大库梯级开发的生态保护与恢复"研究构想. 工程科学与技术, 2017, 49(1): 19-26. |
| [14] |
曹文宣. 长江上游特有鱼类自然保护区的建设及相关问题的思考. 长江流域资源与环境, 2000, 9(2): 131-132. |
| [15] |
曹文宣. 有关长江流域鱼类资源保护的几个问题. 长江流域资源与环境, 2008, 17(2): 163-164. DOI:10.3969/j.issn.1004-8227.2008.02.001 |
| [16] |
陈求稳, 张建云, 莫康乐, 陈宇琛, 关铁生, 王国庆, 林育青. 水电工程水生态环境效应评价方法与调控措施. 水科学进展, 2020, 31(5): 793-810. |
| [17] |
黄光明, 王海龙, 王伟营, 陈豪, 丁诚志, 熊合勇, 李坤. 澜沧江流域鱼类栖息地保护实践. 水生态学杂志, 2015, 36(6): 86-92. |
| [18] |
高婷, 李翀, 廖文根. 实施支流生境替代保护的基本原则. 中国水利水电科学研究院学报, 2012, 10(4): 267-272. DOI:10.3969/j.issn.1672-3031.2012.04.006 |
| [19] |
杨青瑞, 陈声威, 何建宽, 祁文龙. 支流生境替代保护效果评价指标体系与评价方法研究. 中国水利水电科学研究院学报, 2015, 13(6): 408-413, 420-420. |
| [20] |
Kruk A, Ciepłucha M, Zięba G, Błońska D, Tybulczuk S, Tszydel M, Marszał L, Janic B, Pietraszewski D, Przybylski M, Penczak T. Spatially diverse recovery (1986-2012) of fish fauna in the Warta River, Poland: the role of recolonizers' availability after large-area degradation. Ecological Engineering, 2017, 102: 612-624. DOI:10.1016/j.ecoleng.2017.01.019 |
| [21] |
Marques H, Dias J H P, Perbiche-Neves G, Kashiwaqui E A L, Ramos I P. Importance of dam-free tributaries for conserving fish biodiversity in Neotropical reservoirs. Biological Conservation, 2018, 224: 347-354. DOI:10.1016/j.biocon.2018.05.027 |
| [22] |
de Magalhães Lopes J, Pompeu P S, Alves C B M, Peressin A, Prado I G, Suzuki F M, Facchin S, Kalapothakis E. The critical importance of an undammed river segment to the reproductive cycle of a migratory Neotropical fish. Ecology of Freshwater Fish, 2019, 28(2): 302-316. DOI:10.1111/eff.12454 |
| [23] |
Garcia D A Z, Vidotto-Magnoni A P, Costa A D A, Casimiro A C R, Jarduli L R, Ferraz J D, De Almeida F S, Orsi M L. Importance of the Congonhas River for the conservation of the fish fauna of the Upper Paraná basin, Brazil. Biodiversitas Journal of Biological Diversity, 2019, 20(2): 474-481. DOI:10.13057/biodiv/d200225 |
| [24] |
邱阳凌. 黑水河生态河貌模拟及鱼类替代生境评价[D]. 重庆: 重庆交通大学, 2018.
|
| [25] |
杨志, 唐会元, 龚云, 朱迪, 赵娜. 产卵迁徙对金沙江黑水河下游鱼类群聚结构变动的影响. 湖泊科学, 2018, 30(3): 753-762. |
| [26] |
林俊强, 彭期冬, 任杰, 柏海霞, 赵利. 赤水河与金沙江下游河段鱼类生境条件的相似性分析. 淡水渔业, 2014, 44(6): 93-99. DOI:10.3969/j.issn.1000-6907.2014.06.016 |
| [27] |
何大明, 汤奇成, 刘国才, 宿华, 李丽娟, 赵楚年, 王又丰, 刘天仇, 杨明, 冯彦. 中国国际河流. 北京: 科学出版社, 2000.
|
| [28] |
康斌, 胡文娴, 祈文龙, 杨春明, 李江红. 补远江鱼类多样性研究. 渔业科学进展, 2010, 31(3): 6-14. DOI:10.3969/j.issn.1000-7075.2010.03.002 |
| [29] |
Ding C Z, Jiang X M, Wang L E, Fan H, Chen L Q, Hu J M, Wang H L, Chen Y F, Shi X T, Chen H, Pan B H, Ding L Y, Zhang C, He D M. Fish assemblage responses to a low-head dam removal in the Lancang River. Chinese Geographical Science, 2019, 29(1): 26-36. DOI:10.1007/s11769-018-0995-x |
| [30] |
水利部中国科学院水工程生态研究所. 小黑江回龙山水电站水生生态影响评价专题报告. 武汉: 水利部中国科学院水工程生态研究所, 2010.
|
| [31] |
陈小勇. 云南鱼类名录. 动物学研究, 2013, 34(4): 281-337. |
| [32] |
张觉民, 何志辉. 内陆水域渔业自然资源调查手册. 北京: 农业出版社, 1991.
|
| [33] |
杨岚, 李恒. 云南湿地. 北京: 中国林业出版社, 2010: 479-521.
|
| [34] |
孙洁. 基于耳石微结构的澜沧江光唇裂腹鱼产卵时间和仔鱼生长研究[D]. 昆明: 云南大学, 2018.
|
| [35] |
汪松, 乐佩琦, 陈宜瑜. 中国濒危动物红皮书--鱼类. 北京: 科学出版社, 1998.
|
| [36] |
刘明典, 陈大庆, 段辛斌, 王珂, 刘绍平. 澜沧江云南段鱼类区系组成与分布. 中国水产科学, 2011, 18(1): 156-170. |
| [37] |
Pinkas L, Oliphant M S, Iverson I L K. Fish Bulletin 152. Food habits of albacore, bluefin tuna, and bonito in California waters//California Department of Fish and Game, ed. Fish Bulletin. Sacramento: California Department of Fish and Game, 1971: 1-10.
|
| [38] |
李凡, 徐炳庆, 吕振波, 王田田. 莱州湾鱼类群落优势种生态位. 生态学报, 2018, 38(14): 5195-5205. |
| [39] |
王朝阳, 黄德才, 胡茂林, 柳俊, 苗春, 王焘, 简少卿. 赣江下游一级支流鱼类多样性比较分析. 水生态学杂志, 2019, 40(3): 99-106. |
| [40] |
Clarke K R, Warwick R M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation. 2nd ed. Plymouth: Plymouth Marine Laboratory, 2001.
|
| [41] |
Majewski A R, Lynn B R, Lowdon M K, Williams W J, Reist J D. Community composition of demersal marine fishes on the Canadian Beaufort Shelf and at Herschel Island, Yukon Territory. Journal of Marine Systems, 2013, 127: 55-64. DOI:10.1016/j.jmarsys.2013.05.012 |
| [42] |
王雪辉, 林昭进, 杜飞雁, 邱永松, 孙典荣, 王跃中, 梁新, 黄硕琳. 南海西北部陆架区鱼类的种类组成与群落格局. 生态学报, 2013, 33(7): 2225-2235. |
| [43] |
Rohlf F J. Paleontological Data Analysis. By Øyvind Hammer and, David A T Harper. Malden (Massachusetts): Blackwell Publishing. $89.95 (paper). xi+351 p; ill; index ISBN: 1‐4051‐1544‐0.2006. The Quarterly Review of Biology, 2007, 82(4): 411-411. |
| [44] |
Suen J P, Herricks E E. Investigating the causes of fish community change in the Dahan River (Taiwan) using an autecology matrix. Hydrobiologia, 2006, 568(1): 317-330. DOI:10.1007/s10750-006-0209-x |
| [45] |
李婷, 唐磊, 王丽, 安磊, 王骏, 莫康乐, 陈求稳. 水电开发对鱼类种群分布及生态类型变化的影响--以溪洛渡至向家坝河段为例. 生态学报, 2020, 40(4): 1473-1485. |
| [46] |
褚新洛, 陈银瑞. 云南鱼类志(上册). 北京: 科学出版社, 1989.
|
| [47] |
褚新洛, 陈银瑞. 云南鱼类志(下册). 北京: 科学出版社, 1990.
|
| [48] |
Poulsen A F, Hortle K G, Valbo-Jorgensen J, Chan S, Chhuon C K, Viravong S, Bouakhamvongsa K, Suntornratana U, Yoorong N, Nguyen T T, Tran B Q. Distribution and ecology of some important riverine fish species of the Mekong River Basin. Phnom Penh: Mekong River Commission, 2004.
|
| [49] |
申安华, 李光华, 赵树海, 杨光清, 刘跃天, 吴敬东, 李威, 宋建宇, 丁登虎, 熊燕. 光唇裂腹鱼胚胎发育与仔鱼早期发育的研究. 水生态学杂志, 2013, 34(6): 76-80. DOI:10.3969/j.issn.1674-3075.2013.06.013 |
| [50] |
薛晨江, 雷春云, 符世伟. 中华鲱鲇繁殖生物学研究. 水生态学杂志, 2017, 38(6): 102-106. |
| [51] |
洪迎新, 施文卿, 陈宇琛, 刘东升, 马宏海, 朱昊彧, 陈求稳. 水电梯级开发进程中澜沧江干流鱼类群落演变特征. 生态学报, 2021, 41(1): 235-253. |
| [52] |
Zhang C, Ding L Y, Ding C Z, Chen L Q, Sun J, Jiang X M. Responses of species and phylogenetic diversity of fish communities in the Lancang River to hydropower development and exotic invasions. Ecological Indicators, 2018, 90: 261-279. DOI:10.1016/j.ecolind.2018.03.004 |
| [53] |
Fugi R, Agostinho A A, Hahn N S. Trophic morphology of five benthic-feeding fish species of a tropical floodplain. Revista Brasileira de Biologia, 2001, 61: 27-33. DOI:10.1590/S0034-71082001000100005 |
| [54] |
Quinn J M, Hickey C W. Magnitude of effects of substrate particle size, recent flooding, and catchment development on benthic invertebrates in 88 New Zealand rivers. New Zealand Journal of Marine and Freshwater Research, 1990, 24(3): 411-427. DOI:10.1080/00288330.1990.9516433 |
| [55] |
Granzotti R V, Miranda L E, Agostinho A A, Gomes L C. Downstream impacts of dams: shifts in benthic invertivorous fish assemblages. Aquatic Sciences, 2018, 80(3): 28. DOI:10.1007/s00027-018-0579-y |
| [56] |
马巍, 彭静, 彭文启, 黄伟, 骆辉煌. 河流栖息地适合度曲线与分级评价标准研究. 中国水利水电科学研究院学报, 2016, 14(1): 23-28. |
| [57] |
Nunn A D, Copp G H, Vilizzi L, Carter M G. Seasonal and diel patterns in the migrations of fishes between a river and a floodplain tributary. Ecology of Freshwater Fish, 2010, 19(1): 153-162. DOI:10.1111/j.1600-0633.2009.00399.x |
| [58] |
Da Silva P S, Makrakis M C, Miranda L E, Makrakis S, Assumpção L, Paula S, Dias J H P, Marques H. Importance of reservoir tributaries to spawning of migratory fish in the Upper Paraná River. River Research and Applications, 2015, 31(3): 313-322. DOI:10.1002/rra.2755 |
| [59] |
杨君兴, 陈小勇, 陈银瑞. 中国澜沧江鱼芒科鱼类种群现状及洄游原因分析. 动物学研究, 2007, 28(1): 63-67. DOI:10.3321/j.issn:0254-5853.2007.01.011 |
| [60] |
Hortle K G, Nam S. Mitigation of the Impacts of Dams on Fisheries - A Primer. Vientiane: Mekong River Commission, 2017.
|