 2022, Vol. 42
2022, Vol. 42文章信息
- 张续同, 李卫明, 张坤, 熊伟唯, 陈圣盛, 刘子健
- ZHANG Xutong, LI Weiming, ZHANG Kun, XIONG Weiwei, CHEN Shengsheng, LIU Zijian
- 长江宜昌段桥边河大型底栖动物功能摄食类群时空分布特征
- Spatiotemporal distribution of macroinvertebrate functional feeding groups in Qiaobian River, a tributary of Yangtze River in Yichang
- 生态学报. 2022, 42(7): 2559-2570
- Acta Ecologica Sinica. 2022, 42(7): 2559-2570
- http://dx.doi.org/10.5846/stxb202107061806
-
文章历史
- 收稿日期: 2021-07-06
- 网络出版日期: 2022-02-13






大型底栖动物是河流生物群落中分布最普遍的类群之一, 也是河流生态系统食物链的中间环节, 对河流生态系统的能量流动和物质循环具有重要意义[1]。大型底栖动物功能摄食类群指河流生态系统中食性基本相同的底栖动物群落, 基于食物网关系建立的功能群落, 能更精确地研究底栖动物群落结构和功能以及与环境因子的相关性[2]。随着全球工业化和城市化的迅猛发展, 生态环境最终无可避免地成为经济增长的受害者, 其中河流等水体往往成为各类污染的最终受纳者, 使其水质和生物群落受到威胁[3]。因此, 研究大型底栖动物功能摄食类群的时空分布特征, 建立功能摄食类群与环境因子的响应关系, 对有效指示河流生态系统健康和河流生态修复具有重要意义[4]。
根据摄食方式的不同可将大型底栖动物划分为五种功能类群:刮食者(Scrapers, SC), 滤食者(Filter-collectors, FC), 收集者(Gather-collectors, GC), 撕食者(Shredders, SH), 捕食者(Predators, PR)[5—6]。早期研究大型底栖动物主要基于传统的形态分类方法[7], 大多采用底栖动物群落结构参数对河流生态系统进行一系列的评价。但有研究指出:基于传统形态分类方法对大型底栖动物群落结构的研究忽略了类群间在生态功能上的差异, 提供的信息有限, 而物种功能群是联系群落结构和生态系统的框架[8—9]。因此研究功能摄食类群对于底栖动物的群落结构和功能组成具有重要作用[10]。Park等利用人工神经网络(Artificial Neural Network)评价了环境变量对大型底栖动物功能摄食类群的影响[11];Rawer-Jost等通过德国11条河流大型底栖动物功能摄食类群的不同指标体系, 测试其检测河流受损的准确性[12];Gayraud等研究了人类活动对底栖动物功能摄食组成的影响[13];Jiang等检验了大型底栖动物功能摄食类群与河流连续体(River Continuum Concept, RCC)的对应程度[14]。
目前, 国外对于大型底栖动物功能摄食类群的研究多集中于建立基于功能摄食类群的评价体系和应用以及关于大型底栖动物功能摄食类群对人类活动的响应等方面[8, 15]。在我国针对河流生境状况的时间和空间变化对大型底栖动物功能摄食类群影响的研究还不够深入, 且大型底栖动物功能摄食类群的结构特征对判断流域整体生境的时空状况具有重要意义[16]。基于此, 本研究以长江一级支流桥边河为研究对象, 根据2020—2021年采样调查的结果, 研究大型底栖动物功能摄食类群的时间和空间分布特征, 探讨大型底栖动物功能摄食类群与河流环境因子之间的相关性, 尝试应用基于功能摄食类群的生态参数评估桥边河生态系统健康, 为其水质的生物评价和河流生态修复提供理论基础。
1 材料与方法 1.1 研究区域及点位设置桥边河为长江宜昌段一级支流, 发源于宜昌市点军区土城乡竹蒿山, 流经点军区土城乡、桥边镇、点军街道和高新区电子信息产业园, 于点军街道朱市街社区流入长江。桥边河流域面积295 km2, 主河道长40 km。流域属丘区地形, 流域内山势平缓, 最高海拔1168 m, 最低海拔55 m, 最大高差1113 m。流域地势为西北高东南低, 河流总体流向由西北向东南, 穿流于低山丘陵之间。随着流域内社会经济的发展、村镇人口数量的增加等因素, 导致桥边河水质不断受到威胁。
本研究于夏季(2020年8月)、冬季(2021年1月)和春季(2021年4月)分别对桥边河流域的大型底栖动物进行采样调查, 同时现场测定水体的理化指标, 记录河流和岸边生境等相关情况。从桥边河源头到入江口, 根据河流生境特点分上、中、下3个河段, 共设置8个采样点(图 1)。其中上游段源头—土城(Q1—Q2), 有车溪自然保护风景区, 水流速度相对较快, 两岸较窄, 底质主要以大石块为主;中游段土城—桥边(Q3—Q5), 河面逐渐变宽, 两岸植被茂密, 底质主要以卵石为主;下游段桥边—点军(Q6—Q8), 水流较缓, 岸边杂草多生, 底质主要以粗细沙为主。

|
| 图 1 桥边河采样点分布 Fig. 1 Distribution of sampling points in Qiaobian River Q: 代表桥边河 |
依据《生物多样性观测技术导则淡水底栖大型无脊椎动物》(HJ 710.8—2014)[17]的规定, 在每个采样点两岸上下游100 m范围内利用索伯网(Surber net, 网口尺寸为30 cm×30 cm, 网口孔径为500 μm)随机采集3个平行样方, 然后将3个样方混合成一个样品, 总采样面积为0.27 m2。在现场将采集到的大型底栖动物样品过40目筛网, 通过初步粗挑后放入300 mL样品瓶内, 然后用10%甲醛溶液固定带回实验室, 在显微镜下进行分类鉴定和计数[18], 尽可能地鉴定至最低分类单元, 通常为属或种级。最后将底栖动物表面的液体用滤纸吸干, 用万分之一电子天平称重(湿重), 计算其密度(个/m2)和生物量(g/m2)。
1.2.2 环境因子采集根据《地表水和污水监测技术规范》(HJ/T91—2002)[19]进行环境因子的采集, 在采样现场使用多参数水质监测分析仪(Hydrolab DS5, 美国)测定pH、水温(T)、溶解氧(DO)、电导率(Cond)、浊度(Tur)和水深(Dep);使用Vector声学多普勒点式流速仪(挪威)测定流速(Vel)。同时于每个采样点采集2瓶500 mL水样, 低温保存, 在24 h内送回实验室, 依据《水和废水监测分析方法》(第四版)[20]测定总氮(TN)、硝态氮(NO3--N)、铵态氮(NH4+-N)、总磷(TP)、正磷酸盐(PO43-)、化学需氧量(CODMn)等理化因子。
1.3 大型底栖动物功能摄食类群生态系统属性按照大型底栖动物各功能摄食类群的组成及结构特征, 参考前人的研究成果[21—22], 基于功能摄食类群的参数从物质循环、物质纵向输送能力、沿岸物质输入等方面对桥边河流域的生态系统状况进行分析(表 1)。
| 基于功能摄食类群的参数 Metrics based on functional feeding groups |
生态系统属性 Ecosystem attributes |
| 物质循环Material cycling | |
| F1刮食者密度 Density of scrapers |
初级生产力 |
| F2刮食者密度与滤食者、收集者之和的比率 Ratio of scrapers to filterers and gather-collectors |
自养作用/异养作用 |
| F3撕食者密度和收集者密度之和 Density of shredders and gather-collctors |
分解能力 |
| F4生物量 Total biomass |
次级生产力 |
| 物质纵向输送能力Longitudinal transport | |
| F5滤食者密度 Density of filterers |
纵向输送能力 |
| F6滤食者与撕食者和收集者之和的比率 Ratio of filterers to shredders and gather-collectors |
相对纵向输送能力 |
| 沿岸物质的输入Lateral input | |
| F7撕食者密度 Density of shredders |
沿岸物质输入 |
| F8撕食者密度与总密度之比 Ratio of shredders to total density |
相对物质输入量 |
| 其它Others | |
| F9撕食者与滤食者和收集者之和的比率 Ratio of shredders to filterers and gather-collectors |
粗有机颗粒输入量/细有机颗粒输入量 |
| F10捕食者密度与总密度之比 Ratio of predators to total density |
捕食者的下行效应 |
| F11刮食者和滤食者之和与撕食者和收集者之和的比率 Ratio of scrapers and filterers to total shredders and gather-collectors |
生境稳定性 |
大型底栖动物群落结构采用Shannon-Wiener多样性指数(H)、Pielou均匀度指数(J)和Margalef物种丰富度指数(D)研究, 采用优势度值(Y)确定大型底栖动物的优势种, 计算公式如下:
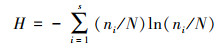
|
(1) |
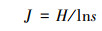
|
(2) |
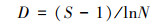
|
(3) |
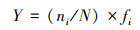
|
(4) |
式中, ni为第i中的密度, N为大型底栖动物的总密度, S为物种种类数, fi为第i种出现的频率。当Y≥0.02时, 确定该物种为优势种[23]。
1.4.2 统计分析利用束缚型排序(Constrained Ordination)识别环境因子和功能摄食类群之间的关系[24]。对大型底栖动物功能摄食类群进行除趋势对应分析(Detrended correspondence analysis, DCA), 假设得出物种的单峰响应值小于2, 则选择冗余分析(Redundancy analysis, RDA), 若大于2, 则进行典范对应分析(Canonical correspondence analysis, CCA)。
基础数据统计与处理在Excel 2016中, 利用Origin 2017绘制柱状图, Canoco 5.0软件进行DCA和RDA分析, 用ArcMap 10.2绘制采样点位图。除pH值以外, 其余所有水环境因子数据和底栖动物功能摄食类群数据均在SPSS中进行对数转换[log (x+1)]。
2 结果 2.1 环境因子特征桥边河流域不同季节和不同河段的水环境因子如表 2所示, 三个季节中夏季水体温度最高, 冬季最低, 不同河段pH范围为7.82—8.46, 水体呈弱碱性。氮、磷相关环境因子沿河流整体有增大趋势, 溶解氧、电导率、水深等环境因子存在季节性差异。
| 环境因子 Environmental factors |
春季Sping | 夏季Summer | 冬季Winter | ||||||||
| 上游 Upstream |
中游 Midstream |
下游 Downstream |
上游 Upstream |
中游 Midstream |
下游 Downstream |
上游 Upstream |
中游 Midstream |
下游 Downstream |
|||
| TN/(mg/L) | 0.48±0.04 | 0.77±0.14 | 0.83±0.12 | 0.74±0.03 | 0.87±0.11 | 0.96±0.10 | 0.75±0.06 | 0.74±0.34 | 0.70±0.13 | ||
| NO3--N/(mg/L) | 0.37±0.01 | 0.44±0.10 | 0.49±0.01 | 0.69±0.03 | 0.57±0.05 | 0.58±0.08 | 0.48±0.03 | 0.45±0.17 | 0.47±0.08 | ||
| NH4+-N/(mg/L) | 0.01±0 | 0.03±0.01 | 0.09±0.05 | 0.03±0.01 | 0.07±0.04 | 0.07±0.01 | 0.01±0 | 0.04±0.03 | 0.09±0.03 | ||
| TP/(mg/L) | 0.02±0.01 | 0.08±0.07 | 0.09±0.02 | 0.02±0 | 0.06±0.05 | 0.07±0.02 | 0.02±0 | 0.05±0.06 | 0.05±0.03 | ||
| PO43-/(mg/L) | 0.01±0 | 0.03±0.01 | 0.03±0.01 | 0.01±0 | 0.02±0 | 0.02±0 | 0.02±0 | 0.02±0.02 | 0.02±0.02 | ||
| CODMn/(mg/L) | 1.59±0.32 | 2.06±0.24 | 2.38±0.31 | 1.81±0.12 | 1.58±0.41 | 1.61±0.22 | 0.89±0.11 | 1.40±0.54 | 2.29±1.38 | ||
| T/℃ | 15.88±0.32 | 14.83±0.99 | 16.68±0.64 | 20.78±1.09 | 23.37±1.25 | 24.41±2.05 | 11.77±0.65 | 11.02±1.37 | 7.91±1.03 | ||
| pH | 7.82±1.11 | 8.38±0.16 | 8.02±0.25 | 8.19±0.16 | 8.27±0.04 | 8.46±0.09 | 8.45±0.16 | 8.15±0.12 | 7.99±0.26 | ||
| Tur/NTU | 6.24±4.12 | 4.08±1.95 | 24.14±14.81 | 10.90±11.46 | 18.33±12.57 | 29.60±25.98 | 7.31±7.58 | 8.78±7.71 | 33.19±8.48 | ||
| DO/(mg/L) | 12.58±0.66 | 10.21±1.17 | 8.71±0.71 | 8.32±1.06 | 7.50±0.93 | 7.13±0.50 | 14.33±1.12 | 14.29±0.62 | 11.76±1.02 | ||
| Cond/(ms/m) | 530.02±9.90 | 534.67±29.69 | 583.33±29.91 | 415.5±20.51 | 427.33±3.51 | 423.00±45.74 | 600.51±44.55 | 541.00±72.92 | 796.67±137.70 | ||
| Vel/(m/s) | 0.48±0.06 | 0.27±0.18 | 0.32±0.32 | 0.48±0.21 | 0.57±0.06 | 0.41±0.18 | 0.16±0.08 | 0.28±0.20 | 0.37±0.17 | ||
| Dep/m | 0.17±0.06 | 0.39±0.05 | 0.77±0.66 | 0.28±0.05 | 0.61±0.50 | 1.37±0.95 | 0.09±0.02 | 0.24±0.06 | 0.63±0.60 | ||
| TN: 总氮Total nitrogen; NO3--N: 硝态氮Nitrate nitrogen; NH4+-N: 铵态氮Ammonium nitrogen; TP: 总磷Total phosphorus; PO43-: 正磷酸盐Phosphate anion; CODMn: 化学需氧量Chemical oxygen demand; T: 温度Temperature; pH: 酸碱度Potential of hydrogen; Tur: 浊度Turbidity; DO: 溶解氧Dissolved oxygen; Cond: 电导率Conductivity; Vel: 流速Flow velocity; Dep: 水深Water depth | |||||||||||
2020年8月、2021年1月和4月三次野外调查共采集到大型底栖动物1225只, 隶属于3门6纲16目38种。其中以节肢动物门种类最多, 有昆虫纲17种、甲壳纲6种;软体动物门次之, 有腹足纲7种、双壳纲6种;环节动物门最少, 有寡毛纲和蛭纲各1种(图 2)。分析桥边河大型底栖动物的各功能摄食类群组成, 发现滤食者相对丰度最大, 为52.8%;刮食者、捕食者和撕食者的相对丰度分别为21.8%、10.1%和9.3%;收集者相对丰度最少, 为6.1%, 功能摄食类群物种数依次为9种、8种、12种、6种和4种。由表 3可知各季节大型底栖动物功能摄食类群的优势种, 三个季节的共同优势种为河蚬、闪蚬、方格短沟蜷和铜锈环棱螺, 除此之外, 春季的优势种还有卵萝卜螺(Y=0.03)、河蟌(Y=0.03)和秀丽白虾(Y=0.09), 冬季的优势种还有黄色羽摇蚊(Y=0.03)。

|
| 图 2 桥边河大型底栖动物群落结构 Fig. 2 Community structure of macroinvertebrates in Qiaobian River |
| 功能摄食类群 Functional feeding groups |
优势种 Dominant species |
春季(Y)
Spring |
夏季(Y)
Summer |
冬季(Y)
Winter |
| 滤食者FC Filter-collectors | 河蚬Corbicula fluminea | 0.33 | 0.45 | 0.30 |
| 闪蚬Corbicula nitens | 0.07 | 0.04 | 0.07 | |
| 刮食者SC Scrapers | 方格短沟蜷Semisulcospira cancellata | 0.02 | 0.03 | 0.03 |
| 铜锈环棱螺Bellamya aeruginosa | 0.09 | 0.06 | 0.11 | |
| 卵萝卜螺Radix ovata | 0.03 | — | — | |
| 捕食者PR Predators | 河蟌Calopterygidae | 0.03 | — | — |
| 撕食者SH Shredders | 秀丽白虾Exopalaemon modestus | 0.09 | — | — |
| 收集者GC Gather-collectors | 黄色羽摇蚊Chironomus flaviplumus | — | — | 0.03 |
根据图 3可知, 桥边河流域3个季节的大型底栖动物群落Shannon-Wiener多样性指数(H)、Pielou均匀度指数(J)和Margalef物种丰富度指数(D)的平均值分别为1.66、0.82和1.87。这三项多样性指数的季节性差异均不显著(P>0.05), 且Pielou均匀度指数(J)和Margalef物种丰富度指数(D)在空间分布上也不显著(P>0.05), 只有Shannon-Wiener多样性指数(H)在空间分布上差异显著(P=0.02)。

|
| 图 3 桥边河流域大型底栖动物多样性指数 Fig. 3 Biodiversity indices of the macroinvertebrates community in Qiaobian River |
图 4为桥边河大型底栖动物功能摄食类群相对丰度时空变化。从图中可知滤食者功能群的相对丰度在各河段和各季节最高, 为主要的优势类群, 最大相对丰度为65.9%, 出现在冬季中游河段, 在春季上游河段出现最小值25.8%;刮食者最大相对丰度为29.0%, 出现在冬季下游河段, 在春季上游河段为0;收集者的相对丰度最大值为20.5%, 出现在夏季下游河段, 在春季中游河段出现最小值1.4%;撕食者和捕食者的最大相对丰度都出现在春季上游河段, 分别为32.6%和36.0%, 最小值都出现在下游河段, 分别为冬季的1.6%和春季的1.0%。

|
| 图 4 大型底栖动物功能摄食类群相对丰度时空变化 Fig. 4 Spatiotemporal variation of relative abundance of functional feeding groups of macroinvertebrates |
从季节分布上来看, 滤食者始终处于优势地位, 相对丰度最大值为58.0%, 出现在夏季, 在春季出现最小值47.3%;刮食者在3个季节的相对丰度变化不大, 最大值和最小值分别为22.8%和20.5%, 分别出现在冬季和夏季;收集者最大相对丰度为11.0%, 出现在冬季, 在春季出现最小值3.8%;撕食者和捕食者的最大相对丰度都出现在春季, 分别为14.8%和11.6%, 最小值都在冬季出现, 分别为3.9%和7.8%。
2.3.2 密度时空分布特征图 5为桥边河大型底栖动物功能摄食类群密度时空分布格局。滤食者在夏季中游河段有最大密度181个/m2, 在夏季下游河段有最小密度37个/m2;刮食者最大密度为76个/m2, 也出现在夏季中游河段, 在春季上游河段密度为0;收集者在夏季下游河段有最大密度21个/m2, 最小密度出现在夏季上游河段, 为4个/m2;撕食者和捕食者的最大密度都出现在春季上游河段, 分别为54个/m2和60个/m2, 最小值分别出现在冬季下游河段和春季下游河段, 都为3个/m2。

|
| 图 5 大型底栖动物功能摄食类群密度时空变化 Fig. 5 Spatiotemporal variation of density of functional feeding groups of macroinvertebrates |
从季节分布上来看, 优势类群滤食者密度在夏季出现最大值120个/m2, 冬季出现最小值71个/m2;刮食者最大密度出现在春季, 为52个/m2, 冬季出现最小值30个/m2;收集者最大密度出现在冬季, 为15个/m2, 春季出现最小值9个/m2;撕食者和捕食者的最大密度都出现在春季, 分别为35个/m2和27个/m2, 冬季出现最小值, 分别为6个/m2和11个/m2。
2.4 大型底栖动物功能摄食类群生态系统评价表 4是基于功能摄食类群生态系统特征值对大型底栖动物时空分布格局进行的分析, 结果表明:在物质循环方面, 底栖动物初级生产力F1在桥边河中游大于上下游, 春季大于其他季节;自养作用/异养作用F2从上游到下游逐渐增高, 春季大于其他季节;底栖动物分解能力F3在上游大于中下游, 春季大于其他季节;底栖动物次级生产力F4在中游最大, 春季和夏季大于冬季。物质纵向能力输送方面, 纵向输送能力F5和相对纵向输送能力F6在中游都明显高于上下游, 夏季高于其他季节。在沿岸物质的输入方面, 沿岸物质输入量F7和相对物质输入量F8在上游大于其他区域, 春季高于其他季节。在其他方面, 粗有机颗粒输入量/细有机颗粒输入量F9在上游高于中下游, 春季高于其他季节;捕食者的下行效应F10沿河流逐步减弱, 在各季节间差异性较小;生境稳定性F11在中游最大, 夏季大于其他季节。
| 基于功能摄食类群的参数 Metrics based on functional feeding groups |
上游 Upstream |
中游 Midstream |
下游 Downstream |
春季 Spring |
夏季 Summer |
冬季 Winter |
|
| 物质循环 | F1 | 12.96 | 55.97 | 45.27 | 51.85 | 42.13 | 29.63 |
| Material cycling | F2 | 0.14 | 0.42 | 0.44 | 0.44 | 0.32 | 0.35 |
| F3 | 37.65 | 21.81 | 30.45 | 43.06 | 24.54 | 19.44 | |
| F4 | 39.06 | 43.87 | 33.58 | 46.20 | 41.20 | 26.02 | |
| 物质纵向输送能力 | F5 | 81.48 | 125.93 | 86.01 | 109.26 | 119.44 | 70.83 |
| Longitudinal transport | F6 | 2.16 | 5.77 | 2.82 | 2.54 | 4.87 | 3.64 |
| 沿岸物质的输入 | F7 | 27.78 | 15.64 | 12.76 | 34.26 | 13.43 | 5.09 |
| Lateral input | F8 | 0.16 | 0.07 | 0.08 | 0.15 | 0.07 | 0.04 |
| 其它Others | F9 | 0.30 | 0.12 | 0.12 | 0.29 | 0.10 | 0.06 |
| F10 | 0.25 | 0.07 | 0.04 | 0.12 | 0.10 | 0.08 | |
| F11 | 6.51 | 8.34 | 4.31 | 10.74 | 6.58 | 5.17 |
基于功能摄食类群特征值的时空变化对桥边河生态系统健康状况进行评估, 在空间尺度上, 桥边河大型底栖动物生物量在中上游要大于下游, 生境稳定性在中游最大;在时间尺度上, 包括生物量在内的多项参数都在春季较高, 生境稳定性在春季也最强。
2.5 大型底栖动物功能摄食类群与水环境因子的相关性分析依据对桥边河大型底栖动物功能摄食类群进行的除趋势对应分析(DCA), 得出物种的单峰响应值为1.2, 冗余分析(RDA)适合本研究。因此选择冗余分析对各类底栖动物功能摄食类群密度同TN、NO3--N、NH4+-N、TP、PO43-、CODMn、T、pH、DO、Cond、Tur、Dep和Vel共13个水环境因子进行分析。
通过蒙特卡罗检验(Monte Carlo Permutation Test), 排除贡献小的因子(P>0.05), 对比发现不同季节主要影响大型底栖动物功能摄食类群的环境参数存在异同。RDA分析结果显示(图 6), 春季主要影响桥边河流域大型底栖动物功能摄食类群的环境因子为DO(P=0.01)和NH4+-N(P=0.04);夏季主要影响桥边河流域大型底栖动物功能摄食类群的环境因子为NO3--N(P=0.02)、pH(P=0.02)和Dep(P=0.04);冬季主要影响桥边河流域大型底栖动物功能摄食类群的环境因子为T(P=0.01)和NO3--N(P=0.03)。

|
| 图 6 桥边河大型底栖动物功能摄食类群与环境因子的RDA分析 Fig. 6 Relationships of macroinvertebrates FFGs and environmental factors obtained by Redundancy analysis in Qiaobian River FC: 滤食者Filter-collectors; SC: 刮食者Scrapers; PR: 捕食者Predators; SH: 撕食者Shredders; GC: 收集者Gather-collectors; NH4+-N: 铵态氮Ammonium nitrogen; DO: 溶解氧Dissolved oxygen; NO3--N: 硝态氮Nitrate nitrogen; pH: 酸碱度Potential of hydrogen; Dep: 水深Water depth; T: 温度Temperature; Q1—Q8: 采样点Sampling sites |
桥边河流域大型底栖动物功能摄食类群中滤食者密度明显大于其他类群, 处于优势地位;收集者密度最小, 处于劣势地位。这与国内其他一些学者的研究存在差异, 如香溪河流域底栖动物以收集者占优势, 撕食者处于劣势[10];漓江流域底栖动物刮食者处于优势地位, 撕食者处于劣势地位[25]。研究对比发现, 在不同的研究区域大型底栖动物功能摄食类群优劣势类群存在异同, 其主要原因可能在于各个研究区域的地理尺度不同, 河流生境状况各异, 并且底栖动物各功能摄食类群的物种组成和分布是其对河流栖息地不同生境因子的综合反应。桥边河属山区向平原过渡型河流, 河床底质具有显著的空间异质性, 上游河段底质主要以大石块为主, 中游河段底质主要以卵石为主, 下游河段底质主要以粗细沙为主。因此不同的地理尺度和生境状况使得处于优劣势的功能摄食类群不同。滤食者主要依靠水流的作用来获取食物, 桥边河水浅流急, 给滤食者提供了适宜的滤食流速生境, 保证了其取食和呼吸[26], 使之成为优势功能群。河流中颗粒物的自然沉降是收集者获取食物的主要来源, 桥边河过快的流速使得沉积的有机碎屑较少, 不能满足收集者的摄食要求[27], 使得收集者处于劣势地位。
3.2 大型底栖动物功能摄食类群时空分布依据河流连续体概念(River Continuum Concept, RCC), 从上游到下游撕食者的相对丰度逐步减小, 收集者的相对丰度逐步增大, 刮食者的相对丰度在中游最大[28]。这与本研究的结果基本一致。大型底栖动物功能摄食类群的空间分布受到河流底层基质的异质性[29]、其他环境因素和人类活动的共同影响。在上游河段, 河岸植被通过遮阴减少自养产生, 并对河流输入大量有机质碎屑;在中下游河段, 水流减缓和人类活动影响使得有机物在河床沉积, 这保证了底栖动物足够的食物来源。除与河流连续体相关的功能群外, 滤食者和捕食者功能群的相对丰度在河流空间上存在差异。滤食者的相对丰度在桥边河上游最低, 中下游较高。原因可能在于滤食者的优势种闪蚬和河蚬均为耐污种, 而桥边河上游有王家坝水库饮用水源保护区和车溪自然保护风景区, 生态环境保护较好, 外源污染物少[30], 水质较清洁。在中下游有土城乡、桥边镇和点军工业园区等农村和城镇活动区, 村镇生活污水、畜牧业养殖沼泽液、乡村农药化肥残留等污染物直接或间接输移到河流中, 造成水体富营养化, 导致水体污染严重, 使得滤食者的相对丰度增大, 底栖动物群落结构主要以耐污种群为主。捕食者的相对丰度在上游最大, 并且沿河流减小, 原因可能在于捕食者的优势种河蟌为敏感类群, 常栖息于山溪等流水水域, 上游水源保护区为其创造了良好的生境条件。
比较桥边河流域3个季节采集到的大型底栖动物, 发现其密度在春季最高, 夏季次之, 冬季最低。大型底栖动物功能摄食类群密度的季节变化与其种类生殖习性和种间关系等有关[31—32]。滤食者主要以悬浮的细有机颗粒为食, 存在适宜的滤食流速[33], 本研究中滤食者的密度在夏季最大, 原因可能在于夏季桥边河流量大, 水流速度较快, 为其提供了较为适宜的流速生境。相反, 收集者的主要食物为有机碎屑, 易在流速较低的生境沉积, 因此在流速较缓的冬季收集者密度最大。捕食者的密度在春季最大, 因为春季是各类底栖动物的主要繁殖季节, 许多新幼体进入群落, 为捕食者创造了有利的捕食条件。撕食者的密度在春季明显大于其他季节, 因为撕食者功能群的主要优势种为秀丽白虾, 秋冬季河流储备了河岸植被向其输入的大量有机质, 而春季水温逐渐升高, 是撕食者活动能力最旺盛的季节[34], 通过摄食沉积的有机质使其密度达到最大值。刮食者的密度在春季最大, 冬季最小, 原因在于刮食者的食物主要是附石性藻类[35], 而春季为着生藻类的生长提供了充足的光照等适宜条件。
3.3 大型底栖动物功能摄食类群与环境因子的关系环境因子是组成河流生态系统的重要成分, 其直接或间接影响水生生物的群落结构和时空分布[36], 分析环境因子对大型底栖动物功能摄食类群的影响对评估河流生态系统健康具有重要作用[37]。国内外研究指出, 不同区域不同季节影响大型底栖动物功能摄食类群的环境因子存在差异[38—39]。通过RDA分析得出, DO、pH、Dep、T、NO3--N、NH4+-N等6个物理和化学因子对桥边河大型底栖动物功能摄食类群群落结构和时空分布具有显著影响。春季DO与大型底栖动物功能摄食类群相关性最强, 原因在于春季是大多数底栖动物繁殖最旺盛的季节, 其新陈代谢活动加强、取食和呼吸作用会消耗水中大量溶解氧。除此之外, 河流中耗氧有机质的分解也会抑制底栖生物, 从而对底栖动物功能摄食类群的组成和分布产生影响[40]。NH4+-N会对大型底栖动物产生毒害作用, 有研究指出[41], 水体中的NH4+-N会直接影响底栖动物的免疫反应和部分机体组织, 含量过高会导致其死亡。夏季pH与大型底栖动物功能摄食类群相关性较强, 因为底栖动物主要生存在酸碱度适宜的水环境中, 而桥边河夏季pH最高值达到8.60, 超出了底栖动物的适合生存范围, 对其生存产生有害影响。有研究表明收集性状通常与水深Dep呈负相关[42], 在本研究中刮食者和收集者与水深呈负相关, 这是因为随着水深逐渐增大, 河流中营养物质含量相对减少, 食物的减少导致底栖动物密度也相应递减。夏季和冬季NO3--N与大型底栖动物功能摄食类群同时具有较强的相关性, 这可能是因为水中硝态氮含量的增加会促进水草生长进而影响底栖藻类生长[22], 使得硝态氮与表征河流初级生产力的刮食者密度呈负相关。冬季水温对底栖动物功能摄食类群有直接的影响, 桥边河冬季水温最低为7.29℃, 温度过低对底栖动物的生长和繁殖具有抑制作用。
综合分析表明, 不同时空尺度影响桥边河各功能摄食类群的主要环境因子存在差别, 主要可以归纳为物理因子、化学因子、生物因子三类, 其中物理因子主要为水深、水温和流速等;化学因子主要为pH、溶解氧、营养盐和有机物等;生物因子主要为河岸带植被、水生植物以及河流生态系统中的竞争和捕食行为等。
4 结论(1) 桥边河大型底栖动物共分为5种功能摄食类群, 其中滤食者占绝对优势, 其次为刮食者、捕食者和撕食者, 收集者处于劣势地位。
(2) 通过分析大型底栖动物功能摄食类群的组成和时空分布, 可知各功能摄食类群的分布特征受河流时空条件的约束, 同时也能够表征流域底栖动物不同时空序列受人为活动影响的程度。
(3) 不同季节影响大型底栖动物功能摄食类群的环境因子存在差异, 春季的主要影响因子为DO和NH4+-N, 夏季为Dep、NO3--N和pH, 冬季为T和NO3--N。
| [1] |
Edema C U, Ayeni J O, Aruoture A. Some observations on the zooplankton and macrobenthos of the Okhuo River, Nigeria. Journal of Aquatic Sciences, 2002, 17(2): 145-149. |
| [2] |
Ma T W, Huang Q H, Wang H, Wang Z J, Wang C X, Huang S B. Selection of benthic macroinvertebrate-based multimetrics and preliminary establishment of biocriteria for the bioassessment of the water quality of Taihu Lake, China. Acta Ecologica Sinica, 2008, 28(3): 1192-1200. DOI:10.1016/S1872-2032(08)60038-4 |
| [3] |
Charkhabi A H, Sakizadeh M, Bayat R. Land use effects on heavy metal pollution of river sediments in Guilan, southwest of the Caspian sea. Gaspian Journal of Environmental Sciences, 2008, 6(2): 133-140. |
| [4] |
Scheibler E E, Campón F F, Silnik S L, Wellnitz T. Seasonal influence and local factors affecting macroinvertebrate structure in a high-altitude Andean stream. Journal of Mountain Science, 2020, 17(6): 1374-1386. DOI:10.1007/s11629-019-5813-0 |
| [5] |
Cummins K W. Structure and Function of Stream Ecosystems. BioScience, 1974, 24(11): 631-641. DOI:10.2307/1296676 |
| [6] |
Riva-Murray K, Bode R W, Phillips P J, Wall G L. Impact source determination with biomonitoring data in New York State: concordance with environmental data. Northeastern Naturalist, 2002, 9(2): 127-162. DOI:10.1656/1092-6194(2002)009[0127:ISDWBD]2.0.CO;2 |
| [7] |
陈博, 李卫明, 陈求稳, 刘德富. 夏季漓江不同底质类型和沉水植物对底栖动物分布的影响. 环境科学学报, 2014, 34(7): 1758-1765. |
| [8] |
Bazzanti M, Bella V D. Functional feeding and habit organization of macroinvertebrate communities in permanent and temporary ponds in central Italy. Journal of Freshwater Ecology, 2004, 19(3): 493-497. DOI:10.1080/02705060.2004.9664924 |
| [9] |
Córdova-Tapia F, Zambrano L. Fish functional groups in a tropical wetland of the Yucatan Peninsula, Mexico. Neotropical Ichthyology, 2016, 14(2): e150162. |
| [10] |
蒋万祥, 蔡庆华, 唐涛, 渠晓东. 香溪河水系大型底栖动物功能摄食类群生态学. 生态学报, 2009, 29(10): 5207-5218. DOI:10.3321/j.issn:1000-0933.2009.10.005 |
| [11] |
Park Y S, Lek S, Chon T S, Verdonschot P. Evaluation of environmental factors to determine the distribution of functional feeding groups of benthic macroinvertebrates using an artificial neural network. Journal of Ecology and Environment, 2008, 31(3): 233-241. DOI:10.5141/JEFB.2008.31.3.233 |
| [12] |
Rawer-Jost C, Böhmer J, Blank J, Rahmann H. Macroinvertebrate functional feeding group methods in ecological assessment. Hydrobiologia, 2000, 422-423: 225-232. DOI:10.1023/A:1017078401734 |
| [13] |
Gayraud S, Statzner B, Bady P, Haybachp A, Schöll F, Usseglio-Polatera P, Bacchi M. Invertebrate traits for the biomonitoring of large European rivers: an initial assessment of alternative metrics. Freshwater Biology, 2003, 48(11): 2045-2064. DOI:10.1046/j.1365-2427.2003.01139.x |
| [14] |
Jiang X M, Xiong J, Xie Z C, Chen Y F. Longitudinal patterns of macroinvertebrate functional feeding groups in a Chinese river system: a test for river continuum concept (RCC). Quaternary International, 2011, 244(2): 289-295. DOI:10.1016/j.quaint.2010.08.015 |
| [15] |
Soldner M, Stephen I, Ramos L, Angus R, Wells N C, Grosso A, Crane M. Relationship between macroinvertebrate fauna and environmental variables in small streams of the Dominican Republic. Water Research, 2004, 38(4): 863-874. DOI:10.1016/S0043-1354(03)00406-8 |
| [16] |
王博涵, 吴丹, 张吉, 殷旭旺, 赵长森, 窦同文. 济南河流大型底栖动物摄食功能群多样性及时空动态. 生态学报, 2017, 37(21): 7128-7139. |
| [17] |
中华人民共和国环境保护部. HJ 710.8-2014生物多样性观测技术导则淡水底栖大型无脊椎动物. 北京: 中国环境科学出版社, 2014.
|
| [18] |
段学花, 王兆印, 徐梦珍. 底栖动物与河流生态评价. 北京: 清华大学出版社, 2010.
|
| [19] |
国家环境保护总局. HJ/T 91-2002地表水和污水监测技术规范. 北京: 中国环境科学出版社, 2002.
|
| [20] |
国家环境保护总局. 水和废水监测分析方法(第四版). 北京: 中国环境科学出版社, 2002.
|
| [21] |
Yoshimura C, Tockner K, Omura T, Moog O. Species diversity and functional assessment of macroinvertebrate communities in Austrian rivers. Limnology, 2006, 7(2): 63-74. DOI:10.1007/s10201-006-0170-4 |
| [22] |
孙小玲, 蔡庆华, 李凤清, 杨顺益, 谭路. 春季昌江大型底栖无脊椎动物群落结构及功能摄食类群的空间分布. 应用与环境生物学报, 2012, 18(2): 163-169. |
| [23] |
芦康乐, 武海涛. 三江平原沼泽湿地底栖无脊椎动物群落特征. 中国环境科学, 2020, 40(2): 832-838. DOI:10.3969/j.issn.1000-6923.2020.02.044 |
| [24] |
邢圆, 吴小平, 欧阳珊, 张君倩, 徐靖, 银森录, 谢志才. 赣江水系大型底栖动物多样性与受胁因子初探. 生物多样性, 2019, 27(6): 648-657. |
| [25] |
朱晨曦, 莫康乐, 唐磊, 吴焱, 李婷, 林育青, 陈求稳. 漓江大型底栖动物功能摄食类群时空分布及生态效应. 生态学报, 2020, 40(1): 60-69. |
| [26] |
傅小城. 小水电站对香溪河底栖动物的影响[D]. 武汉: 中国科学院水生生物研究所, 2007.
|
| [27] |
汪峰, 单彪, 于涛, 王帅帅. 小清河流域春季底栖动物功能摄食类群与水环境因子的关系. 河北渔业, 2019(12): 41-46. DOI:10.3969/j.issn.1004-6755.2019.12.013 |
| [28] |
Vannote R L, Minshall G W, Cummins K W, Sedell J R, Cushing C E. The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences, 1980, 37(1): 130-137. DOI:10.1139/f80-017 |
| [29] |
Kerakova M, Varadinova E. Influence of the river bottom substrate and sediment organic component on the macrozoobenthos functional feeding groups. Comptes Rendus De l'Acad'Emie Bulgare Des Sciences, 2020, 73(1): 66-72. |
| [30] |
李金京, 李卫明, 粟一帆, 孙徐阳, 胡威. 长江支流桥边河大型底栖动物群落结构及水质生物评价. 环境科学学报, 2020, 40(8): 3020-3027. |
| [31] |
沈洪艳, 曹志会, 刘军伟, 王文欢, 张远. 太子河流域大型底栖动物功能摄食类群与环境要素的关系. 中国环境科学, 2015, 35(2): 579-590. |
| [32] |
郑金秀, 陈明秀, 胡菊香. 苏皖交界河网区后生浮游动物群落结构及其影响因子分析. 水生态学杂志, 2014, 35(6): 53-60. DOI:10.3969/j.issn.1674-3075.2014.06.008 |
| [33] |
粟一帆, 李卫明, 李金京, 孙徐阳, 胡威. 桥边河大型底栖动物生境适宜性. 生态学报, 2020, 40(16): 5844-5854. |
| [34] |
温周瑞, 张大文, 谢平, 徐军. 太湖日本沼虾、秀丽白虾现存量的周年动态研究--以梅梁湾、贡湖湾为例. 长江流域资源与环境, 2011, 20(9): 1035-1039. |
| [35] |
贾兴焕, 吴乃成, 唐涛, 蔡庆华. 香溪河水系附石藻类的时空动态. 应用生态学报, 2008, 19(4): 881-886. |
| [36] |
Frissell C A, Liss W J, Warren C E, Hurley M D. A hierarchical framework for stream habitat classification: viewing streams in a watershed context. Environmental management, 1986, 10(2): 199-214. DOI:10.1007/BF01867358 |
| [37] |
耿雷华, 刘恒, 钟华平, 刘翠善. 健康河流的评价指标和评价标准. 水利学报, 2006, 37(3): 253-258. DOI:10.3321/j.issn:0559-9350.2006.03.001 |
| [38] |
Bêche L A, Mcelravy E P, Resh V H. Long‐term seasonal variation in the biological traits of benthic‐macroinvertebrates in two Mediterranean‐climate streams in California, U.S.A. Freshwater Biology, 2010, 51(1): 56-75. |
| [39] |
蒋万祥, 贾兴焕, 周淑婵, 李凤清, 唐涛, 蔡庆华. 香溪河大型底栖动物群落结构季节动态. 应用生态学报, 2009, 20(4): 923-928. |
| [40] |
宋晓娜, 于涛, 张远, 张彦, 尹秀英. 利用三维荧光技术分析太湖水体溶解性有机质的分布特征及来源. 环境科学学报, 2010, 30(11): 2321-2331. |
| [41] |
洪美玲, 陈立侨, 顾顺樟, 刘超, 龙章强, 张伟. 氨氮胁迫对中华绒螯蟹免疫指标及肝胰腺组织结构的影响. 中国水产科学, 2007, 14(3): 412-418. DOI:10.3321/j.issn:1005-8737.2007.03.010 |
| [42] |
Frainer A, Johansen K S, Siwertsson A, Mousavi S K, Brittain J E, Klemetsen A, Knudsen R, Amundsen P A. Variation in functional trait composition of benthic invertebrates across depths and seasons in a subarctic lake. Fundamental and Applied Limnology, 2016, 188(2): 103-112. DOI:10.1127/fal/2016/0839 |