 2022, Vol. 42
2022, Vol. 42文章信息
- 杨波, 曹晓栋, 程洁婕, 余登利, 李光荣, 魏鲁明, 白新祥
- YANG Bo, CAO Xiaodong, CHENG Jiejie, YU Dengli, LI Guangrong, WEI Luming, BAI Xinxiang
- 贵州省宽阔水国家级自然保护区亮叶水青冈群落结构及多样性
- Community structure and diversity of Fagus lucida in Kuankuoshui National Nature Reserve of Guizhou Province, China
- 生态学报. 2022, 42(7): 2685-2697
- Acta Ecologica Sinica. 2022, 42(7): 2685-2697
- http://dx.doi.org/10.5846/stxb202101080083
-
文章历史
- 收稿日期: 2021-01-08
- 网络出版日期: 2021-12-15






2. 贵州省宽阔水国家级自然保护区管理局, 绥阳 563300;
3. 北京森旺林业工程设计有限公司, 北京 100083
2. Kuankuoshui National Nature Reserve, Suiyang 563300, China;
3. Beijing Senwang Forestry Engineering Design Co., Ltd, Beijing 100083, China
亮叶水青冈(Fagus lucida Rehd. et Wils.)属于壳斗科(Fagaceae)水青冈属(Fagus)落叶大乔木[1], 中国特有种, 主要分布在长江北部神农架以南(31.5°N), 南岭山脉以北(24°N)地区, 包括湖北、四川、贵州、湖南、广东等省海拔700m以上的山地[2], 是中国十分重要的林木资源, 常常形成优势群落。亮叶水青冈是第三纪残留的古老物种, 它不仅对古气候、古地理、古生物、古生态的研究具有重大价值, 而且对落叶常绿阔叶混交林生态系统的组成结构、营养结构、能量流动、物质循环以及东亚水青冈生态系统的研究, 均具有较大价值。
贵州亮叶水青冈天然林分布广泛, 面积较大, 保存较完整的主要是在宽阔水国家级自然保护区、梵净山国家级自然保护区以及黔东南等地区[3]。宽阔水自然保护区分布了大量的亮叶水青冈原生性群落, 该区的主要保护对象就是原生性亮叶水青冈林及其生境[4]。目前国内对于水青冈属的植物已经有一定的研究。洪必恭等对中国水青冈属5种特有种的地理分布与5项气候指标的关系进行了分析, 其限制分布的主要气候因子是生长季和降水量[5]。曹铁如等对湖南八大公山亮叶水青冈林的物种组成以及植物区系进行了较深入的分析[6], 得知该群落富集种少、稀疏种多, 而且这些种的多度有一定的规律性特征[7]。张谧等认为神农架地区的米心水青冈及曼青冈占据群落中乔木层不同的高度, 且均不缺乏更新贮备, 因此形成稳定共存的群落[8]。赵睿等通过对天目山国家级自然保护区内米心水青冈种群的调查, 得知该群落多干指数达到96.4%, 表明萌条在种群的更新中起着关键作用[9]。陈志刚等通过资源分配试验证明, 锐齿槲栎和米心水青冈在林隙与林下环境显示出不同的光利用策略[10]。汪正祥等对分布于中国亚热带山地如南山、梵净山、宽阔水、八大公山4地域的亮叶水青冈林进行了植被的比较研究, 根据37个样方调查的资料, 区分出3个群丛6个亚群[11]。赫尚丽等对滇东北乌蒙山自然保护区的亮叶水青冈群落结构和物种多样性进行了研究[12]。对于宽阔水自然保护区内的原生亮叶水青冈群落的研究涉及种—多度结构[13]、数量分类[4]、种群结构与动态[14]等研究, 但时间跨度甚大, 缺乏深入的研究。亮叶水青冈在宽阔水自然保护区属于顶极群落的建群种, 在山地的一定海拔高度上形成了原生性群落与原始森林, 对其研究可以揭示该地区群落的稳定与物种多样性的维持具有重要的意义, 同时也是研究植物系统演化和植物地理学的关键类群之一[2]。但近年来研究发现, 该区亮叶水青冈种群结构老化, 大部分树木种子结实量低甚至不结实, 林下亮叶水青冈幼树缺失, 仅在开阔的林缘地带过渡性地分布一些亮叶水青冈幼树[15], 并且亮叶水青冈林下镶嵌分布有大量的金佛山方竹(Chimonobambusa utilis)和箭竹(Fargesia spathacea)等种群, 它们与其他乔木的幼苗幼树进行资源竞争, 同时还通过影响群落微环境进而对幼苗更新产生影响[16-18]。本文选取宽阔水国家级自然保护区亮叶水青冈群落进行调查, 就群落结构特征及物种多样性进行了分析和总结, 为更深入地揭示宽阔水自然保护区生物多样性和物种共存机制提供参考, 为探索宽阔水保护区森林生态系统的稳定性维持机理提供基础理论, 为制定更有效的物种保护措施提供科学依据。
1 研究区概况宽阔水国家级自然保护区(东经107°02′—107°14′, 北纬28°06′—28°19′)位于贵州省绥阳县北部大娄山山脉向西南延伸地段, 东西横延约19km, 南北纵深约20km, 总面积26231hm2[19], 海拔650—1762m。区内地形起伏多样、切割强烈, 属喀斯特区域中山峡谷地貌[20]。保护区具有低纬度山地湿润气候特点, 属中亚热带湿润季风性气候, 年平均气温11.7—15.2℃, 年降水量在1300—1350mm, 年平均相对湿度超过82%[21]。区内土壤以黄棕壤、黄壤为主;植被为中亚热带常绿落叶阔叶混交林, 森林面积达到保护区总面积的80%以上[22]。贵州省宽阔水国家级自然保护区植物物种丰富, 植物区系较为原始, 在保护区保存有1300hm2的天然亮叶水青冈林, 亮叶水青冈是该原生林的主要建群种[21], 这种原生性亮叶水青冈林具有典型的地带性植被分布特征, 是维持整个保护区乃至黔北地区生态功能的重要支撑。
2 研究方法 2.1 样地设置与调查方法从2019年6月至2020年12月在研究区选取亮叶水青冈分布地段, 设置样地面积共11700m2, 其中面积为20m×20m的样地29个, 10m×10m的幼林样地1个(因太阳山顶地形限制只能设置10m×10m的小样地), 记录每个样地的基本概况(表 1)。根据国家生物多样性观测技术导则(陆生维管植物和植物多样性观测固定样地的设置)的方法, 将标椎样地划分为4个10m×10m的样方, 在顶点设置样桩, 然后将这4个大样方分别划分为4个5m×5m的小样方, 以该小样方作为外业的基本单位。调查乔木层(高度≥5m起测)种类, 记录项目为:树种、株数、树高、胸径、枝下高、冠幅、生活力等;选取上述的每个10m×10m样方的右上角的小样方(5m×5m)作为灌木调查单元进行灌木层调查, 调查内容:树种、株丛数、平均地径、平均高度、生长状况、分布状况和盖度等;在上述灌木样方的右上角位置设置面积为1m×1m的样方作为草本植物观测样方, 调查记录:种类、株数、平均冠径、平均高度、生长情况、盖度等。
| 样地 Site |
优势种 Dominant species |
地点 Location |
海拔/m Altitude |
坡度/(°) Slope |
坡向 Aspect |
| 01 | 亮叶水青冈、云贵鹅耳枥Carpinus pubescens Burk. | 岩脚湾西南148m | 1500m | 8 | 东北 |
| 02 | 亮叶水青冈、多脉青冈Cyclobalanopsis multinervis W.C.Cheng & T.Hong | 岩脚湾西南171m | 1512m | 35 | 北 |
| 03 | 亮叶水青冈、云贵鹅耳枥 | 岩脚湾南61m | 1540m | 20 | 西南 |
| 04 | 亮叶水青冈 | 白石溪 | 1441m | 10 | 东北 |
| 05 | 亮叶水青冈 | 老屋基南137m | 1457m | 2 | 西南 |
| 06 | 亮叶水青冈、多脉青冈、檫木Sassafras tzumu (Hemsl.) Hemsl. | 老屋基东南72m | 1437m | 5 | 西南 |
| 07 | 亮叶水青冈、云贵鹅耳枥、多脉青冈 | 老屋基东南126m | 1478m | 15 | 西南 |
| 08 | 亮叶水青冈 | 老屋基东216m | 1516m | 10 | 北 |
| 09 | 亮叶水青冈、多脉青冈 | 老屋基东北71m | 1457m | 5 | 西坡 |
| 10 | 亮叶水青冈、多脉青冈 | 宽阔东北222m | 1453m | 5 | 东南 |
| 11 | 亮叶水青冈 | 宽阔东北365m | 1464m | 5 | 西南 |
| 12 | 亮叶水青冈 | 岩脚湾西南148m | 1436m | 35 | 北 |
| 13 | 亮叶水青冈 | 凉风南垭东南485m | 1482m | 10 | 东北 |
| 14 | 亮叶水青冈 | 凉风垭南604m | 1477m | 5 | 北 |
| 15 | 亮叶水青冈、多脉青冈 | 凉风垭南641m | 1452m | 10 | 西北 |
| 16 | 亮叶水青冈 | 凉风垭南588m | 1510m | 10 | 西南 |
| 17 | 亮叶水青冈、多脉青冈 | 岩脚弯东150m | 1540m | 5 | 西南 |
| 18 | 亮叶水青冈、多脉青冈、云贵鹅耳枥 | 岩脚湾东南279m | 1570m | 5 | 西南 |
| 19 | 亮叶水青冈、云贵鹅耳枥、多脉青冈 | 岩脚湾东南475m | 1523m | 10 | 西南 |
| 20 | 亮叶水青冈、多脉青冈 | 路好坎北670m | 1457m | 5 | 西 |
| 21 | 亮叶水青冈、多脉青冈、杉木Cunninghamia lanceolata (Lamb.) Hook. | 大鱼塘东594m | 1465m | 10 | 西南 |
| 22 | 亮叶水青冈 | 太阳山 | 1711m | 55 | 南 |
| 23 | 亮叶水青冈、多脉青冈 | 珙桐沟 | 1457m | 30 | 西 |
| 24 | 亮叶水青冈、雷公鹅耳枥Carpinus viminea Lindley、檫木 | 大湾坝子东北183m | 1418m | 5 | 西南 |
| 25 | 亮叶水青冈、云贵鹅耳枥 | 大湾坝子东北165m | 1416m | 30 | 西 |
| 26 | 亮叶水青冈、云贵鹅耳枥、多脉青冈 | 大湾坝子西北67m | 1380m | 5 | 东 |
| 27 | 亮叶水青冈、云贵鹅耳枥 | 大湾坝子北381m | 1439m | 55 | 南 |
| 28 | 亮叶水青冈、云贵鹅耳枥 | 大湾坝子东南142m | 1463m | 20 | 西南 |
| 29 | 亮叶水青冈、半边月 Weigela japonica var. sinica (Rehd.) Bailey |
大湾坝子东南252m | 1447m | 5 | 西北 |
| 30 | 亮叶水青冈、野鸦椿 Euscaphis japonica (Thunb.) Dippel |
大湾坝子东297m | 1459m | 3 | 西北 |
种群是构成群落的基本单位, 年龄结构是种群的重要特征[23], 种群年龄结构的分析是探讨种群动态特征的一种有效可行的方法[24-25]。在许多研究工作中, 由于测定植物种群个体年龄较为困难, 不少学者采用径级结构分析法替代年龄结构, 即以个体的高度(苗期)及胸径的大小(统称为径级, Size class, sc)代替龄级[26]。径级结构也是反映种群结构稳定性的一个重要指标, 研究种群的径级结构能很好的反映群落的动态变化。故本文研究以“空间代时间”、“立木级代替年龄结构”, 采用经典分级[27]的方法进行统计。根据胸径D将亮叶水青冈分为Ⅰ级(D<5cm, 包括幼苗D<5cm, H<0.5m, 和幼树D<5cm, H>0.5m)、Ⅱ级(5cm≤D<10cm)、Ⅲ级(10cm≤D<20cm)、Ⅳ级(20cm≤D<30cm)和Ⅴ级(D≥30cm)共5个等级, 每一个立木级对应一个年龄级。
2.3 重要值统计分析采用重要值(Importance value, IV)作为评价各物种在群落中的重要性指标[28], 反映其在群落中的功能地位和作用, 计算群落立木层树种的重要值, 参考张金屯的计算方法[29]。即:重要值(IV)=(相对密度+相对优势度+相对频度)/3。并使用Excel 2013对计算得到的重要值进行统计并排序。
2.4 群落数量分类植被数量分类是根据各样方或植物种间的相似关系将其分成若干组, 使组内的各样方或植物种相当相似, 而组间的则尽量相异, 从而实现植物群落比较客观的分类, 反映出一定的生态规律[30]。本文以65种植物在30个样方中的重要值矩阵为基础, 利用SPSS 24.0选用组间连接(类间平均距离)方法和欧氏距离为分类标准, 完成数据处理。首先将30个样方合并, 计算群落立木层树种重要值(IV), 将立木层树种的IV值按大小排序, 去除频度小于5%的偶见种, 共计30个物种选为聚类种, 然后构成30×30矩阵, 对样方和物种进行聚类分析[31]。参考吴征镒《中国植被》[27]的植物群落分类和命名原则, 并结合样方内实际的生态现状, 以优势种原则划分和命名。
2.5 物种多样性分析物种多样性(Species diversity)是指物种水平上的生物多样性, 它是用一定空间范围物种数量和分布特征来衡量的。植物群落物种多样性是反映植物群落组成结构和稳定性的重要指标, 是物种丰富度和分布均匀性的综合反映, 能体现群落结构类型、组织水平、发展阶段、稳定程度和生境的差异[28]。在对一个群落或一个区域物种多样性进行研究时, 一般从种的丰富度或多度、种的均匀度或平衡性和种的总多样性这三种概念出发, 综合分析比较物种的多样性及其影响因素[32]。α多样性用来测度一个均质群落内物种组成状况, 包括物种丰富度、物种多样性和物种均匀度3种指数。本文以下面这几个指数来进行本区域的物种多样性研究。
即:
Gleason丰富度指数:
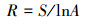
|
Simpson多样性指数:
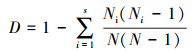
|
Shannon-Wiener多样性指数:
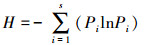
|
Pielou均匀度指数:
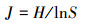
|
式中, S为物种数目, A为单位面积, Ni为第i个物种的个体数, N为所有全部种的个体总数, Pi为频度, 即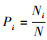
本研究裸子植物采用郑万钧裸子植物分类系统(1978), 被子植物采用恩格勒系统, 蕨类植物采用秦仁昌系统(1978)。通过调查(调查总面积为11700m2), 共计维管植物62科, 111属, 165种;其中蕨类植物4科, 6属, 6种;种子植物58科, 105属, 159种;裸子植物2科, 2属, 2种;被子植物56科, 103属, 157种, 其中单子叶植物2科, 2属, 5种, 双子叶植物54科, 101属, 152种。
乔木层共有59种, 占所有物种的35.76%。其中壳斗科最多, 主要建群种有亮叶水青冈、多脉青冈(Cyclobalanopsis multinervis)、硬壳柯(Lithocarpus hancei)、耳叶柯(Lithocarpus grandifolius)4种。樟科(Lauraceae)其次, 主要建群种有黑壳楠(Lindera megaphylla)和润楠(Machilus nanmu), 其他种相对较少。
灌木层共有91种, 占全部物种的55.15%, 在调查的30个样地中, 有25个出现金佛山方竹, 数目最为庞大, 盖度、密度、高度等都占有绝对的优势。四川大头茶(Polyspora speciosa)、马桑绣球(Hydrangea aspera)、青冈栎(Cyclobalanopsis glauca)、青荚叶(Helwingia japonica)、梾木(Cornus macrophylla)、胡颓子(Elaeagnus pungens)、中国旌节花(Stachyurus chinensis)、南方荚蒾(Viburnum fordiae)、山茶(Camellia japonica)、粉花绣线菊(Spiraea japonica)、半边月(Weigela japonica var. sinica)、野鸦椿(Euscaphis japonica)、杜鹃花(Rhododendron simsii)、山矾(Symplocos sumuntia)等有少量分布。
草本层共有52种, 占全部物种的31.52%, 以冷水花(Pilea notata)和川西鳞毛蕨(Dryopteris rosthornii)居多, 变豆菜(Sanicula chinensis)、绞股蓝(Gynostemma pentaphyllum)、血水草(Eomecon chionantha)、天南星(Arisaema heterophyllum)、蝴蝶花(Iris japonica)、三脉紫菀(Aster trinervius subsp. ageratoides)以及蓟(Cirsium japonicum)等有零星分布。
从表 2统计结果得到科的结构分析, 单种科最为丰富, 在调查的所有群落中占绝对优势, 共28科, 占总科数的45.90%。含2—5种的科较为丰富, 几乎和单种科同等优势, 位居第二, 有25科(47属68种), 占总科数的40.98%。含6—9种的5科(18属30种), 占总科数的8.2%, 分别为山矾科(Symplocaceae)、杜鹃花科(Ericaceae)、山茶科(Theaceae)、百合科(Liliaceae)、五加科(Araliaceae)。含10—15种的3科(17属38种), 仅占总科数的4.92%, 分别为壳斗科、樟科和蔷薇科(Rosaceae)。由此可见该保护区亮叶水青冈群落植物科所含属数、种数的差异明显, 以单种科和寡种科(2—5种)占优势。科内所含种数在10种及以上的科为该区的优势科, 有3科, 共含17属38种。
| 科分级 Family grade |
科名(属/种) Family(Genus/Species) |
数量 Number |
占总科比/% Percentage |
| 含1种的科 Family containing1 species |
紫金牛科、马鞭草科、杉科、省沽油科、桑寄生科、榆科、绣球花科、大风子科、楝科、莎草科、石松科、猕猴桃科、梧桐科、卫矛科、凤仙花科、木通科、唇形科、旌节花科、杜仲科、木兰科、酢浆草科、石杉科、鸢尾科、红豆杉科、紫萁科、漆树科、葫芦科、杨柳科 | 28 | 45.90 |
| 含2—5种的科 Family containing2—5 species |
金粟兰科(1/2)、堇菜科(1/2)、海桐花(1/2)、冬青科(1/2)、杜英科(1/2)、菝葜科(1/2)、十字花科(1/3)、兰科(1/3)、天南星科(1/3)、槭树科(1/4)、菊科(2/2)、安息香科(2/2)、荨麻科(2/2)、大戟科(2/2)、虎耳草科(2/2)、蓼科(2/3)、木犀科(2/3)、桦木科(2/3)、桑科(2/3)、鳞毛蕨科(3/3)、小檗科(3/3)、山茱萸科(3/4)、伞形科(3/3)、忍冬科(3/4)、禾本科(4/4) | 25 | 40.98 |
| 含6—9种的科 Family containing6—9 species |
山矾科(1/6)、杜鹃花科(2/6)、山茶科(4/6)、百合科(5/6)、五加科(6/6) | 5 | 8.20 |
| 含10种以上的科 Family containingmore than 10 species |
壳斗科(5/10)、樟科(6/13)、蔷薇科(6/15) | 3 | 4.92 |
从表 3统计结果得到属的结构分析, 单种属极为丰富, 共78属, 占总属数的70.27%, 对区系结构有着非常重要的影响。含种数2—5种的属丰富程度较低于前者, 但种类总体较丰富, 有31属(75种), 占总属数的27.93%。含6种以上的属较少, 仅有山矾属(Symplocos)和悬钩子属(Rubus), 各有6种, 仅占总属数的1.80%。因此可知该保护区属与科的结构相对一致, 均以单种属和寡种属占优势, 其余等级的属不丰富。
| 属分级 Genus grade |
属名(种数) Genus(Species Number) |
数量 Number |
占总属比/% Percentage |
| 含1种的属 Genus containing1 species |
紫金牛属、柳属、旌节花属、鸢尾属、榆属、野鸦椿属、鼠尾草属、绣球属、莎草属、杉属、香椿属、梧桐属、卫矛属、漆树属、木犀属、钝果寄生属、构属、蓟属、猕猴桃属、含笑属、酢浆草属、山桐子属、桦木属、木荷属、石杉属、楤木属、猫儿屎属、大头茶属、玉簪属、萱草属、山麦冬属、石松属、绞股蓝属、紫菀属、金腰属、黄水枝属、凤仙花属、锦带花属、芒属、树参属、胡颓子属、早熟禾属、方竹属、杜仲属、小檗属、淫羊藿属、大青属、蔷薇属、绣线菊属、吊钟花属、酸模属、梁王茶属、五加属、刺楸属、常春藤属、润楠属、檫木属、野桐属、十大功劳属、水芹属、蓖麻属、鸭儿芹属、变豆菜属、野茉莉属、穇属、白辛树属、紫萁属、耳蕨属、六道木属、贯众属、鳞毛蕨属、楼梯草属、红豆杉属、冷水花属、四照花属、青荚叶属、水青冈属、开口箭属 | 78 | 70.27 |
| 含2—5种的属 Genus containing2—5 species |
山胡椒属(2)、楠属(2)、蓼属(2)、稠李属(2)、石楠属(2)、柯属(2)、鹅耳枥属(2)、锥栗属(2)、梾木属(2)、冬青属(2)、杜英属(2)、榕属(2)、海桐花属(2)、荚蒾属(2)、堇菜属(2)、栎属(2)、金粟兰属(2)、沿阶草属(2)、山茶属(2)、柃属(2)、女贞属(2)、菝葜属(2)、天南星属(3)、虾脊兰属(3)、碎米荠属(3)、木姜子属(3)、李属(3)、青冈属(3)、樟属(4)、槭树属(4)、杜鹃花属(5) | 31 | 27.93 |
| 含6种以上的属 Genus containingmore than 6 species |
山矾属(6)、悬钩子属(6) | 2 | 1.80 |
如图 1所示, 在所调查的30个亮叶水青冈天然林群落样地中, 亮叶水青冈总株数为424株。亮叶水青冈群落种群各龄级呈现出“双峰值”的结构, Ⅰ级和Ⅱ级的个体数分别为58株和133株, 分别占总株数的13.68%和31.37%, 其余三级的个体数分别为96株、25株和112株, 分别占总株数的22.64%、5.90%和26.42%, 三者比例之和高达54.96%。可见, 亮叶水青冈的幼树数量比较多, 但是幼苗数量较少, 大、中型成熟植株较多, 径级呈现双峰曲线, 此结构异常。虽然Ⅰ级幼苗幼树株数较多, 但其主要集中于个别调查到的群落中, 且在群落中的空间结构小, 种类竞争大, 成树可能性较小;对于幼苗幼树来说, 争取到林分上层充足的光照是影响其生产的重要因素, 然而实际调查中发现, 亮叶水青冈群落中金佛山方竹分布较多, 林分郁闭度均较高, 林下光照严重不足, 因此这个有可能是造成亮叶水青冈种群自然更新能力弱的重要原因之一。亮叶水青冈种群内近年缺乏更新, 总体上开始呈现衰退趋势。

|
| 图 1 亮叶水青冈群落的径级结构图 Fig. 1 Diameter class structure diagram of Fagus lucida 根据胸径D将亮叶水青冈分为Ⅰ级(D<5cm, 包括幼苗D<5cm, H<0.5m, 和幼树D<5cm, H>0.5m)、Ⅱ级(5cm≤D<10cm)、Ⅲ级(10cm≤D<20cm)、Ⅳ级(20cm≤D<30cm)和Ⅴ级(D≥30cm)共5个等级, 每一个立木级对应一个年龄级 |
图 2为亮叶水青冈幼苗幼树在30个调查群落中的数量情况, 根据实地调查, 30个样地中有15个样地未发现有亮叶水青冈的幼苗幼树, 在有幼树幼苗的调查样地中, 22号样地中幼树最多13株, 5和11号样地中最少为1株。整体而言, 亮叶水青冈群落的幼苗幼树分布不均, 幼树和中年树相当少, 几乎以大树老树为主, 极有可能出现年龄断层现象。目前研究发现, 宽阔水自然保护区不少种群自然更新能力差, 与人为影响也有较大关系, 方竹笋的经营, 致使方竹的种群数量日渐变大, 更多的夺取了其他种群幼苗幼树的环境资源, 加之其他一些自然灾害等多方因素导致包括亮叶水青冈在内的种群群落正常演替受阻, 让其演替向衰退方向发展。

|
| 图 2 亮叶水青冈幼苗幼树数量分布图 Fig. 2 Distribution of Young Tree in Young Seedlings with Fagus lucida |
从表 4可知, 亮叶水青冈在整个群落中占据重要位置, 优势明显, 是主要优势种。其重要值序在30个样地中, 22个排第一;8个排第二。22号样地中重要值达到58.6939, 但该样方几乎是亮叶水青冈的纯幼林, 且样地受地形限制只能设置10m×10m的样地, 因此该样地重要值应只做参考。此外04号样地达到47.8729为最大, 20号样地最小, 仅为13.2445。此外, 多脉青冈和云贵鹅耳枥(Carpinus pubescens)等在群落中也占据优势, 为主要建群种, 其中云贵鹅耳枥在3个样地中排第1、2个样地中排第2;1个样地中排第3, 多脉青冈在2个样地中排第2。
| 样地 Site |
第一位The first | 第二位The second | 第三位The third | |||||
| 物种 Species |
重要值 Important value |
物种 Species |
重要值 Important value |
物种 Species |
重要值 Important value |
|||
| 01 | 亮叶水青冈 | 28.4667 | 云贵鹅耳枥 | 25.0181 | 多脉青冈 | 10.6761 | ||
| 02 | 亮叶水青冈 | 31.7547 | 多脉青冈 | 30.4631 | 云贵鹅耳枥 | 9.2697 | ||
| 03 | 亮叶水青冈 | 29.9310 | 云贵鹅耳枥 | 23.2202 | 黑壳楠 | 12.2705 | ||
| 04 | 亮叶水青冈 | 47.8729 | 云贵鹅耳枥 | 20.0893 | 多脉青冈 | 12.3135 | ||
| 05 | 亮叶水青冈 | 31.9007 | 多脉青冈 | 10.2204 | 山茶 | 9.8113 | ||
| 06 | 多脉青冈 | 20.9025 | 亮叶水青冈 | 14.1975 | 檫木 | 13.9201 | ||
| 07 | 亮叶水青冈 | 25.4729 | 云贵鹅耳枥 | 14.2013 | 多脉青冈 | 13.2736 | ||
| 08 | 亮叶水青冈 | 32.7453 | 云贵鹅耳枥 | 15.4216 | 山茶 | 10.7312 | ||
| 09 | 亮叶水青冈 | 40.1618 | 多脉青冈 | 21.9929 | 云贵鹅耳枥 | 9.8480 | ||
| 10 | 亮叶水青冈 | 34.3481 | 多脉青冈 | 17.6555 | 西南米槠 | 9.4612 | ||
| 11 | 亮叶水青冈 | 32.3768 | 山茶 | 18.9485 | 多脉青冈 | 12.2381 | ||
| 12 | 亮叶水青冈 | 42.9575 | 山茶 | 21.0052 | 多脉青 | 9.8806 | ||
| 13 | 亮叶水青冈 | 34.0062 | 多脉青冈 | 10.9866 | 云贵鹅耳枥 | 9.3323 | ||
| 14 | 亮叶水青冈 | 40.9854 | 黑壳楠 | 11.4158 | 多脉青冈 | 9.6218 | ||
| 15 | 亮叶水青冈 | 39.7558 | 多脉青冈 | 18.8448 | 黑壳楠 | 11.2432 | ||
| 16 | 亮叶水青冈 | 36.2494 | 云贵鹅耳枥 | 14.8064 | 山矾 | 12.7083 | ||
| 17 | 亮叶水青冈 | 36.3160 | 多脉青冈 | 21.4068 | 云贵鹅耳枥 | 7.1956 | ||
| 18 | 多脉青冈 | 33.4692 | 亮叶水青冈 | 17.9853 | 云贵鹅耳枥 | 16.3452 | ||
| 19 | 亮叶水青冈 | 31.7845 | 云贵鹅耳枥 | 17.4238 | 多脉青冈 | 17.2250 | ||
| 20 | 多脉青冈 | 16.1762 | 多脉青冈 | 13.2445 | 黑壳楠 | 9.2228 | ||
| 21 | 亮叶水青冈 | 31.4203 | 杉木 | 13.2773 | 多脉青冈 | 12.7292 | ||
| 22 | 亮叶水青冈 | 58.6939 | 多脉青冈 | 18.2302 | 云贵鹅耳枥 | 17.9513 | ||
| 23 | 多脉青冈 | 33.1161 | 亮叶水青冈 | 32.7820 | 云贵鹅耳枥 | 8.3369 | ||
| 24 | 亮叶水青冈 | 24.5343 | 雷公鹅耳枥 | 23.3899 | 檫木 | 20.6717 | ||
| 25 | 亮叶水青冈 | 27.5190 | 云贵鹅耳枥 | 22.4533 | 多脉青冈 | 9.1026 | ||
| 26 | 亮叶水青冈 | 28.9509 | 云贵鹅耳枥 | 12.7497 | 多脉青冈 | 10.5851 | ||
| 27 | 云贵鹅耳枥 | 23.8244 | 亮叶水青冈 | 19.8447 | 多脉青冈 | 7.7114 | ||
| 28 | 云贵鹅耳枥 | 36.4896 | 亮叶水青冈 | 33.7128 | 梧桐 | 9.1529 | ||
| 29 | 水马桑 | 30.1418 | 亮叶水青冈 | 21.6915 | 云贵鹅耳枥 | 11.2721 | ||
| 30 | 野鸦椿 | 30.9100 | 亮叶水青冈 | 23.4437 | 云贵鹅耳枥 | 10.7544 | ||
聚类分析得到分类树状图(图 3)。根据分类结果, 结合植物的实际分布情况和群落特征, 按照《中国植被》的命名原则将研究区调查样方植被的主要群落类型进行分类, 共分为以下8个群系类型:

|
| 图 3 植物群落组平均聚类图 Fig. 3 Average cluster diagram of plant community groups |
群系Ⅰ:亮叶水青冈群系(From.Fagus lucida), 包括4、5、7、8、9、10、11、12、13、14、15、16、17、21、22、26号样地共计16个群落, 建群种为亮叶水青冈, 占其绝对优势, 群落总盖度65%—95%。乔木层高度4—30m, 盖度60%—80%, 常见伴生种为云贵鹅耳枥、多脉青冈、杉木(Cunninghamia lanceolata)等, 少有个别群落伴生有构树(Broussonetia papyrifera)、杜英(Elaeocarpus decipiens)、白辛树(Pterostyrax psilophyllus)等。灌木层主要以金佛山方竹为优势种, 仅4和10号样方未见, 其余14个样方均不同程度生长有该种, 其常见伴生种有润楠、黑壳楠、腺柄山矾(Symplocos adenopus)等, 少有伴生槲栎(Quercus aliena)、野桂花(Osmanthus yunnanensis)、异叶梁王茶(Metapanax davidii)等。草本层主要以蕨类为优势种, 次优种为薄片变豆菜(Sanicula lamelligera), 伴生麦冬(Ophiopogon japonicus)、山酢浆草(Oxalis griffithii)、虾脊兰(Calanthe discolor)、白花碎米荠(Cardamine leucantha)等。该类型群系最高海拔1711m;最低海拔1380m;平均海拔1482m, 群落常位于山坡中部或上部, 坡度2°—55°。最高海拔1711m的群落位于太阳山山顶, 太阳山最高峰为保护区海拔最高点, 该样方为亮叶水青冈群系中幼林, 该群落在保护区实属少见。
群系Ⅱ:亮叶水青冈+云贵鹅耳枥群系(From.Fagus lucida+Carpinus pubescens), 包括1、3、19、25、27、28号样地共计6个群落, 建群种为亮叶水青冈和云贵鹅耳枥, 群落总盖度80%—95%。乔木层高度5—25m, 盖度70%—85%, 常见伴生种为檫木、黑壳楠、山茶等, 少有个别群落伴生有大叶青冈(Cyclobalanopsis jenseniana)、山桐子(Idesia polycarpa)、美容杜鹃(Rhododendron calophytum)等。灌木层主要以金佛山方竹、黑壳楠为优势种, 常见伴生种有豪猪刺(Berberis julianae)、川莓(Rubus setchuenensis)、粗糠柴(Mallotus philippensis)等。草本层主要以川西鳞毛蕨为优势种, 伴生粗毛淫羊藿(Epimedium acuminatum)、开口箭(Campylandra chinensis)、玉簪(Hosta plantaginea)等。该类型群系最高海拔1540m;最低海拔1416m;平均海拔1480m, 群落常位于山坡中部或底部, 坡度8°—55°。
群系Ⅲ:亮叶水青冈+多脉青冈群系(From.Fagus lucida+Cyclobalanopsis multinervis), 包括2、18、23号样地共计3个群落, 建群种为亮叶水青冈和多脉青冈, 群落总盖度85%—90%。乔木层高度6—20m, 盖度70%—80%, 常见伴生种为云贵鹅耳枥、黑壳楠、润楠等, 少有伴生四照花(Cornus kousa subsp. chinensis)、梾木等。灌木层主要以金佛山方竹为优势种, 次优种为山矾, 伴生香叶树(Lindera communis)、山莓(Rubus corchorifolius)、光亮山矾(Symplocos lucida)等。草本层仅有几种蕨类。该类型群系最高海拔1570m;最低海拔1457m;平均海拔1513m, 群落常位于山坡中上部, 坡度5°—35°。
群系Ⅳ:亮叶水青冈+多脉青冈+檫木群系(From.Fagus lucida+Cyclobalanopsis multinervis+Sassafras tzumu), 仅有6号样地一个群落, 建群种为亮叶水青冈、多脉青冈和檫木, 群落总盖度90%。乔木层高度7—25m, 盖度75%, 常见伴生种为四川大头茶、西南红山茶(Camellia pitardii)、西南米槠(Castanopsis carlesii)等, 灌木层主要有金佛山方竹、菝葜、细齿叶柃(Eurya nitida)等。草本层主要有川西鳞毛蕨、楼梯草(Elatostema involucratum)、薄片变豆菜等。该类型群系海拔1437m, 群落位于山坡中部, 坡度平缓仅有5°。
群系Ⅴ:多脉青冈+亮叶水青冈群系(From.Cyclobalanopsis multinervis+Fagus lucida), 仅20号样地, 建群种为多脉青冈和亮叶水青冈, 群落总盖度90%。乔木层高度6—25m, 盖度75%, 常见伴生种有四川大头茶、杜鹃、腺柄山矾等。灌木层主要以金佛山方竹为优势种, 伴生山茶、树参(Dendropanax dentiger)、云南樟(Cinnamomum glanduliferum)等。草本层仅有川西鳞毛蕨。该类型群系海拔1437m, 群落位于山坡中上部, 坡度平缓仅有5°。
群系Ⅵ:亮叶水青冈+雷公鹅耳枥+檫木群系(From.Fagus lucida+Carpinus viminea+Sassafras tzumu), 24号样地, 建群种为亮叶水青冈、雷公鹅耳枥和檫木, 群落总盖度90%。乔木层高度6—15m, 盖度80%, 常见伴生种有多脉青冈、光叶海桐(Pittosporum glabratum)、紫楠(Phoebe sheareri)等。灌木层主要以山茶为优势种, 次优种为腺柄山矾, 伴生细齿叶柃、润楠、西南红山茶等。草本层以楼梯草为优势种伴生紫萁(Osmunda japonica)。该类型群系海拔1418m, 群落位于山坡上部, 坡度5°。
群系Ⅶ:水马桑+亮叶水青冈群系(From.Weigela japonica var.sinica+Fagus lucida), 29号样地, 建群种为水马桑和亮叶水青冈, 群落总盖度90%。乔木层高度4—13m, 盖度70%, 常见伴生种有猴樟(Cinnamomum bodinieri)、樱桃(Cerasus pseudocerasus)、稠李(Padus avium)等。灌木层主要以青荚叶(Helwingia japonica)为优势种, 伴生种为小叶女贞(Ligustrum quihoui)、荚蒾(Viburnum dilatatum)、红毛悬钩子(Rubus wallichianus)等。草本层以透茎冷水花(Pilea pumila)和酸模叶蓼(Polygonum lapathifolium)为优势种, 伴生种有深圆齿堇菜(Viola davidii)、碎米荠(Cardamine hirsuta)、鸭儿芹(Cryptotaenia japonica)、三脉紫菀(Aster trinervius subsp. ageratoides)、戟叶蓼(Polygonum thunbergii)、齿果酸模(Rumex dentatus)等。该类型群系海拔1447m, 群落位于山坡底部, 坡度5°。
群系Ⅷ:野鸦椿+亮叶水青冈群系(From.Euscaphis japonica+Fagus lucida), 30号样地, 建群种为野鸦椿和亮叶水青冈, 群落总盖度90%。乔木层高度5—11m, 盖度70%, 常见伴生种有灯台树(Cornus controversa)、八角枫(Alangium chinense)、建始槭(Acer henryi)等。灌木层主要以川莓为优势种, 伴生种为粉花绣线菊(Spiraea japonica)、毛花猕猴桃(Actinidia eriantha)、楤木(Aralia elata)等。草本层以黑鳞耳蕨为优势种, 伴生种有酸模叶蓼、蓟、贯众(Cyrtomium fortunei)、早熟禾(Poa annua)等。该类型群系海拔1459m, 群落位于山坡中部, 坡度3°。
3.5 亮叶水青冈各层片物种多样性特征分析从图 4可知, 在所调查的30个群落样地中, 27号样地的物种最丰富, 30和29号样地次之, Gleason丰富度指数和分别为9.67、8.52、8.51。这3个样地的群落类型、地貌生境相似, 且林下未见金佛山方竹, 其他植物不受其影响。Shannon-wiener指数同样以27号样地最大, 为6.31;而Simpson多样性指数以24号样地最大, 为2.41, 25、27、28、30号样地的多样性也比较高, 均达到了2.30左右;Pielou均匀度指数以24号样地最大, 为2.51。总体趋势上:Gleason物种丰富度, 灌木层>乔木层>草本层;Simpson多样性指数, 乔木层>灌木层>草本层;Shannon-wiener多样性指数, 乔木层>灌木层>草本层;Pielou均匀度指数, 乔木层>草本层>灌木层。亮叶水青冈群落物种多样性相对比较丰富。

|
| 图 4 亮叶水青冈群落各层片物种多样性 Fig. 4 Species diversity in different layers of Fagus lucida glauca community |
从各层片来看, 乔木层物种丰富度最高的是27号样地, 灌木层物种丰富度最高的同样是27号样地, 而草本层最高的是29号样地, 这与植物群落中各层片物种的生物学特性和生态环境变化有关, 同时也和群落承受人为或其它外界干扰能力有重要联系。乔木层植物抗人为或其它外界干扰因素作用的能力强, 其物种丰富度的变化主要是由随海拔变化所导致的气候、土壤等生态环境因素所控制;灌木层、草本层植物对这种抗性作用较弱, 受这种外部干扰因素的影响就较大。Shannon-Wiener指数总体上看以27号样地为最高, Simpson指数和Pielou均匀度指数均在24号样地最高, 说明24号样地植物物种多样, 各物种的个体数量分布均匀, 这两个指数在不同群落中的变化格局十分一致。3个指数的最小值都出现在23号样地, 在实际调查过程中发现, 该样地所处的地形和生境较为特殊, 该区域坡度较陡, 且位于阴坡, 上层乔木较高大, 林下金佛山方竹密集生长, 灌木层植物种类和数量均很少, 草本层仅零星分布个别蕨类植物, 物种集中性较高, 优势种较明显。
4 结论与讨论宽阔水国家级自然保护区亮叶水青冈群落的物种组成总体相对较丰富, 从物种的重要值分析得知, 亮叶水青冈的重要值极高, 在调查的群落中占据明显优势, 另有云贵鹅耳枥、多脉青冈、黑壳楠同是群落中的主要建群种, 在整个森林群落中占有非常重要的地位。灌木层金佛山方竹分布密集, 属主要优势种。草本层优势种为川西鳞毛蕨, 其次是冷水花, 再次是变豆菜。其他植物也分布, 相较之下无优势。通过聚类分析将保护区调查到的群落分为8个群系, 从群系组成及外部特征可以看出, 宽阔水自然保护区亮叶水青冈群落以阔叶林和针阔混交林为主, 符合该区域植被的实际情况。与之前的学者将该保护区森林群落划分为3个类型相比[3], 本研究的划分相对更详细, 类型也更多, 这种情况可能由于选择样地位置和尺度的不同, 以及在时间跨度上的变化造成的。但本研究划分类型中也涵盖了这3种类型, 所以两者研究结果相对一致。
通过对宽阔水自然保护区群落物种多样性研究发现, 各植物群落的物种数量以及建群种、优势种和伴生种的种类等都存在一定程度差异, 但亮叶水青冈在各群落中都占据重要位置, 优势明显。亮叶水青冈群落物种多样性指数及均匀度指数均不低, 主要表现为:Gleason物种丰富度指数为灌木层>乔木层>草本层;Simpson多样性指数及Shannon-Wiener多样性指数均为乔木层>灌木层>草本层;Pielou均匀度指数为乔木层>草本层>灌木层。乔木层多样性指数和均匀度指数相对较高。
物种丰富度、多样性和均匀度指数在各类群落中以及各层片上的分布趋势基本一致, 较好地反映了各类植物群落在物种组成方面的异同。在物种多样性指数的分析中, 经过合理推断导致其指数出现0的情况的因素有以下几个:①调查季节:冬季调查的部分样地, 由于绝大部分草本植物都无法越冬, 一年生草本植物在生长季结束时整体死亡, 多年生的草本植物地上部分死亡, 在冬季调查时均无法有效的记录到, 导致实地调查过程中仅能看到少量的蕨类植物, 如川西鳞毛蕨、狗脊等;②人为干扰:在30个调查样地中, 有25个出现了金佛山方竹, 方竹笋作为当地居民除农耕劳作外的其他收入来源, 在春季草本植物发芽生长时正是人工采摘竹笋的时节, 加上多数草本植物具有相当的药用价值, 采摘、踩踏、砍伐等行为, 导致群落下层植物遭到破坏;③地被物层:主要指死地被物层即林下的枯枝落叶层, 由于枯枝落叶的覆盖, 抑制了苔藓、地衣和草本植物, 特别是对于地面芽植物, 其种子大多粒小, 储蓄养分不多, 幼芽又较纤细, 幼苗穿透能力弱, 致使种子在萌发后, 幼根入土困难, 发生吊根现象而死亡。④林下光照:光照被判定为制约森林植被的盖度和(或)物种丰富度的主要原因[33-34]。林下的光照强度直接决定于上层林冠的结构, 同时林下的温度和湿度又影响了冠层结构, 尤其是林冠层的密度。由于温度和湿度同光照密切相关, 林下层的光照能作为一个综合因子来替代这些微环境的差异[35], 林冠层的分布同时还影响光照环境和净降水量的分布, 从而影响草本植物的生长。
本次调查的11700m2亮叶水青冈群落样地中共有424株亮叶水青冈, 老龄株数占总株数的26.42%, 幼苗占总株数的13.68%, Ⅱ级幼树最多, 占总株数的31.37%, Ⅲ、Ⅳ级相对较少, 分别占总株数的22.64%、5.90%。亮叶水青冈种群年龄呈“双峰值”的结构, 总体数量较多, 但老龄树所占比重较大, 幼树虽较多, 但是年份较短的幼苗很少, 对短时间的种群成长影响可能不明显, 但是从长远看, 天然更新能力开始减弱, 未来成树可能性降低, 亮叶水青冈种群总体开始呈衰退型。此次调查发现, 亮叶水青冈幼苗极少, 在现有分布区内11700m2样地中仅发现58株幼苗, 成树根部也很少萌发, 野外天然亮叶水青冈的更新能力逐渐变弱。由于亮叶水青冈群落中乔木层郁闭度大, 林下金佛山方竹郁闭度高, 透光性较弱, 导致亮叶水青冈及其他物种的幼苗生长缓慢, 竞争力弱。因此, 幼苗不能正常成长和及时更替, 这可能是导致中龄亮叶水青冈较少以及整个群落物种组成不足够丰富的原因之一。从亮叶水青冈种群更新特征及种群生长趋势来看, 幼苗幼树主要集中分布在22号样地, 该样地生境特殊, 位于宽阔水海拔最高的太阳山山顶, 且位于阳坡, 同时上层植被郁闭度不高, 为下层植被更新演替提供了一定条件, 形成小面积初级演替阶段的幼林, 而其他的样地均为顶极群落, 群落结构均较稳定, 但缺乏更新, 群落结构呈衰退趋势。此外, 保护区管理局工作人员发现, 近年很少在此研究区域发现亮叶水青冈的种子和新萌发的幼苗, 根据合理推测有两个原因:其一是区内野生动物过度食用亮叶水青冈种子;其二是亮叶水青冈群落下层枯枝落叶层较厚, 影响其种子入土萌发。除此之外还可能和光照、湿度、土壤等因素有关, 但由于多种条件限制, 无法全面涉及, 还有待进一步深入研究。为了使宽阔水保护区亮叶水青冈群落尤其是亮叶水青冈种群能够良好的保存下来, 建议保护区开展对亮叶水青冈种子萌发和幼苗生长等详细的研究, 必要时进行人工繁殖实验, 扩大其分布范围, 增加个体数量。同时, 尽量减少人为活动, 必要情况下可以适当对上层优势乔木进行开天窗, 降低森林郁闭度, 保护新生幼苗、去劣存优, 扩大群落内的亮叶水青冈种群生存空间, 改善亮叶水青冈种群结构, 为该区植物遗传种质资源的多样性保护创造条件。
对比其他地区亮叶水青冈的研究, 在华中地区南山的光叶水青冈群落中除优势种亮叶水青冈外, 还有多脉青冈、青冈(Cyclobalanopsis glauca)、光枝杜鹃(Rhododendron haofui)等, 灌木层优势种是毛玉山竹(Yushania basihirsuta)、摆竹(Indosasa shibataeaoides)。四川分布的亮叶水青冈群落伴生乔木有水青冈(Fagus longipetiolata)、包果柯(Lithocarpus cleistocarpus)、元宝槭(Acer truncatum)等, 林下主要有筇竹(Chimonobambusa tumidissinoda)、巴山木竹(Arundinaria fargesii)、箭竹。滇东北地区亮叶水青冈群落中出现深裂中华槭(Acer sinense)、中华木荷(Schima sinensis)、扁刺锥(Castanopsis platyacantha)等不同科的共建种。福建地区的亮叶水青冈群落主要以多脉青冈、鹿角杜鹃(Rhododendron latoucheae)、和浙江红山茶(Camellia chekiangoleosa)为优势种。而本研究区分布的亮叶水青冈群落的共建种多为多脉青冈、贵州青冈等壳斗科植物, 以及云贵鹅耳枥、檫木等其他科植物, 林下优势灌木为金佛山方竹和箭竹。该区的亮叶水青冈群落具有自身的特点, 其物种组成既不同于华中、四川、云南、福建等地, 也不同于贵州其他的常绿-落叶阔叶林。亮叶水青冈群落在我国亚热带山地东部东南季风气候区中山地带常形成以其为主要成分的面积较大且较稳定的群落, 但有研究发现亮叶水青冈的其他分布区均不同程度出现更新困难的情况, 群落中林下小径竹类能够降低群落垂直分布的多样性, 直接或间接影响乔木幼苗的更新[36-38]。因而对各地区的亮叶水青冈群落的群落学特征进行分析, 进而深入了解亮叶水青冈群落结构并对其探索保护策略具有重要意义。
| [1] |
傅立国. 中国高等植物(第四卷). 青岛: 青岛出版社, 2001: 689-690.
|
| [2] |
田宇英. 亮叶水青冈落叶阔叶林的群落生态学研究[D]. 厦门: 厦门大学, 2014.
|
| [3] |
朱守谦, 杨业勤. 贵州省宽阔水亮叶水青冈林的数量分类. 生态学杂志, 1985(1): 6-12, 24. |
| [4] |
喻理飞, 陈光平, 余登利. 贵州宽阔水国家级自然保护区生物多样性保护研究. 北京: 中国林业出版社, 2018: 109-110.
|
| [5] |
洪必恭, 安树青. 中国水青冈属植物地理分布初探. Journal of Integrative Plant Biology, 1993, 35(3): 229-233. |
| [6] |
曹铁如, 祁承经, 喻勋林, 龙成良, 周建仁, 张金发. 八大公山亮叶水青冈林及其植物区系的研究. 中南林学院学报, 1993, 13(1): 8-16. |
| [7] |
曹铁如, 祁承经, 喻勋林. 湖南八大公山亮叶水青冈群落物种多样性的研究. 生物多样性, 1997, 5(2): 33-41. |
| [8] |
张谧, 熊高明, 赵常明, 陈志刚, 谢宗强. 神农架地区米心水青冈-曼青冈群落的结构与格局研究. 植物生态学报, 2003, 27(5): 603-609. DOI:10.3321/j.issn:1005-264X.2003.05.004 |
| [9] |
赵睿, 周学峰, 徐娜娜, 赵明水, 刘亮, 陈小勇. 米心水青冈种群萌条更新与高度生长. 生态学报, 2009, 29(7): 3665-3669. DOI:10.3321/j.issn:1000-0933.2009.07.026 |
| [10] |
陈志刚, 樊大勇, 张旺锋, 谢宗强. 林隙与林下环境对锐齿槲栎和米心水青冈种群更新的影响. 植物生态学报, 2005, 29(3): 254-260. |
| [11] |
汪正祥, 雷耘, Kazue Fujiwara, 刘林翰, 薛跃规. 亚热带山地亮叶水青冈林的群落分类及物种组成与更新. 生物多样性, 2006, 14(1): 29-40. DOI:10.3321/j.issn:1005-0094.2006.01.004 |
| [12] |
赫尚丽, 杜凡, 王娟, 罗柏青. 滇东北光叶水青冈群落结构和物种多样性. 西部林业科学, 2016, 45(1): 124-130. |
| [13] |
喻理飞, 朱守谦, 魏鲁明. 贵州喀斯特台原亮叶水青冈林种-多度结构研究. 山地农业生物学报, 1998, 1(1): 9-15. |
| [14] |
朱守谦, 杨业勤. 贵州亮叶水青冈林的结构和动态. 植物生态学与地植物学丛刊, 1985, 9(3): 183-191. |
| [15] |
谢佩耘, 何跃军, 高明浪, 蒋长洪, 杨应, 司建朋, 吴春玉. 金佛山方竹对亮叶水青冈幼树种群数量结构的影响. 热带亚热带植物学报, 2017, 25(3): 225-232. |
| [16] |
Toko Suzaki, Atsushi Kume, Yoshio Ino. Effects of slope and canopy trees on light conditions and biomass of dwarf bamboo under a coppice canopy. Journal of Forest Research, 2005, 10(2): 151-156. DOI:10.1007/s10310-004-0123-x |
| [17] |
Alan H, Taylor, Huang Jinyan, Zhou ShiQiang. Canopy tree development and undergrowth bamboo dynamics in old-growth Abies-Betula forests in southwestern China: a 12-year study. Forest Ecology and Management, 2004, 200(1): 347-360. |
| [18] |
Andrés Holz C., Veblen Thomas T.. Tree regeneration responses to Chusquea montana bamboo die-off in a subalpine Nothofagus forest in the southern Andes. Journal of Vegetation Science, 2006, 17(1): 19-28. |
| [19] |
刘璐. 宽阔水国家级自然保护区黑叶猴生境利用的初步观察[D]. 贵州: 贵州师范大学, 2009.
|
| [20] |
粟海军, 李光容, 陈光平, 姚小刚, 张明明, 胡灿实. 贵州宽阔水自然保护区野生动物红外相机调查初报. 兽类学报, 2018, 38(2): 221-229. |
| [21] |
喻理飞, 谢双喜, 吴太伦. 宽阔水自然保护区综合科学考察集. 贵阳: 贵州科技出版社, 2004.
|
| [22] |
王佳佳, 余志刚, 李筑眉, 蒋鸿, 梁伟. 贵州宽阔水自然保护区鸟类地面巢捕食者的调查. 生态学杂志, 2014, 33(2): 352-357. |
| [23] |
段仁燕, 王孝安, 黄敏毅, 涂云博, 汪超. 太白红杉(Larix chinensis)混交林径级结构与竞争的关系. 生态学报, 2007, 27(11): 4919-4924. DOI:10.3321/j.issn:1000-0933.2007.11.064 |
| [24] |
张文辉, 王延平, 康永祥, 刘祥君. 濒危植物太白红杉种群年龄结构及其时间序列预测分析. 生物多样性, 2004, 12(3): 361-369. DOI:10.3321/j.issn:1005-0094.2004.03.009 |
| [25] |
刘方炎, 李昆, 廖声熙, 崔永忠. 濒危植物翠柏的个体生长动态及种群结构与种内竞争. 林业科学, 2010, 46(10): 23-28. DOI:10.11707/j.1001-7488.20101004 |
| [26] |
高贤明, 王巍, 杜晓军, 马克平. 北京山区辽东栎林的径级结构、种群起源及生态学意义. 植物生态学报, 2001, 25(6): 673-678. DOI:10.3321/j.issn:1005-264X.2001.06.005 |
| [27] |
盘金文, 郭其强, 余大龙, 孙学广, 高超. 不同林龄马尾松人工林优势种径级结构及物种多样性分析. 西北植物学报, 2019, 39(4): 722-728. |
| [28] |
方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6): 533-548. |
| [29] |
张金屯. 数量生态学. 北京: 科学出版社, 2004.
|
| [30] |
阳含熙, 卢泽愚. 植物生态学的数量分类方法. 北京: 科学出版社, 1981.
|
| [31] |
张力. SPSS13.0在生物统计中的应用. 厦门: 厦门大学出版社, 2006.
|
| [32] |
邵丹. 云蒙山国家森林公园植被的多样性研究[D]. 北京: 北京师范大学, 2014.
|
| [33] |
KIRBY K.J.. Changes in the Ground Flora under Plantations on Ancient Woodland Sites. Forestry, 1988, 61(4): 317-338. DOI:10.1093/forestry/61.4.317 |
| [34] |
JENNINGS S B, BROWN N D, SHEIL D. Assessing forest canopies and understorey illumination: canopy closure, canopy cover and other measures. Forestry: An International Journal of Forest Research, 1999, 72(1): 59-74. DOI:10.1093/forestry/72.1.59 |
| [35] |
夏富才, 潘春芳, 赵秀海, 何海燕, 周海城. 长白山原始阔叶红松林林下草本植物多样性格局及其影响因素. 西北植物学报, 2012, 32(2): 370-376. DOI:10.3969/j.issn.1000-4025.2012.02.024 |
| [36] |
王传华, 李俊清, 陈芳清, 杨莹. 鄂东南低丘地区枫香林下枫香幼苗更新限制因子. 植物生态学报, 2011, 35(2): 187-194. |
| [37] |
Panadda Larpkern, Stein R. Moe, Ørjan Totland. Bamboo dominance reduces tree regeneration in a disturbed tropical forest. Oecologia, 2011, 165(1): 161-168. DOI:10.1007/s00442-010-1707-0 |
| [38] |
Becerra P. I., González Rodríguez V., Smith Ramírez C., Armesto J. J.. Spatio-temporal variation in the effect of herbaceous layer on woody seedling survival in a Chilean mediterranean ecosystem. Journal of Vegetation Science, 2011, 22(5): 847-855. DOI:10.1111/j.1654-1103.2011.01291.x |