 2022, Vol. 42
2022, Vol. 42文章信息
- 潘玉梅, 唐赛春, 韦春强, 李象钦, 吕仕洪
- PAN Yumei, TANG Saichun, WEI Chunqiang, LI Xiangqin, Lü Shihong
- 3种本地植物与入侵植物紫茎泽兰的竞争
- Competition between three native plants and invasive Ageratina adenophora
- 生态学报. 2022, 42(6): 2394-2404
- Acta Ecologica Sinica. 2022, 42(6): 2394-2404
- http://dx.doi.org/10.5846/stxb202102240509
-
文章历史
- 收稿日期: 2021-02-24
- 网络出版日期: 2021-11-26





外来生物入侵破坏自然生态系统的结构完整, 威胁生物多样性平衡发展, 给入侵地带来严重经济和生态损失, 通过外来入侵植物的入侵机制探讨对其合理的控制方法是防控外来入侵植物的重要途径, 也是入侵生态学研究的热点问题。一般认为外来入侵植物比本地植物具有较强的竞争力是其成功入侵的主要原因之一[1-2], 因此针对其竞争力学者们提出了生物替代控制方法, 该方法就是根据植物种间竞争关系, 利用有生态效益或经济价值的植物取代入侵植物的一种生态控制方法, 具有持效久、生态和经济效果良好、利于生态修复等特性[3-4], 其关键就是替代目标植物的筛选。新式“武器”假说即化感作用被认为是外来植物成功入侵的另一主要机制[5-6], 且有研究表明入侵植物的化感作用可以协同入侵植物竞争促进其入侵[7-8]。此外, 近年来一些研究者发现土壤微生物群落尤其是土壤丛枝菌根真菌(AMF)有利于入侵植物对本地植物的竞争[9-10]。因此, 在进行生物替代控制时, 考虑入侵植物的化感物质和土壤AMF对入侵植物与目标植物竞争关系的影响可以为探讨提高替代控制效率的方法提供重要信息。
紫茎泽兰(Ageratina adenophora) 是菊科紫茎泽兰属一种多年生半灌木植物, 原产于中美洲, 自20世纪40年代从缅甸入侵我国云南以来, 由于其较高的繁殖能力已在我国云南、四川、广西、贵州等西南地区广泛分布。该植物的入侵降低了当地的生物多样性、破坏了生态群落结构、影响和危害农林畜牧业发展, 给入侵地造成巨大经济和生态损失, 是我国最重要的外来入侵植物之一, 也是我国环保总局确定的最重要的16种外来入侵生物之一[11]。紫茎泽兰具有强的化感作用和竞争力[12-13];可以改变入侵地土壤的养分元素或化学性状及微生物群落结构, 使其利于自身生长而抑制或降低相邻植物的生长和竞争能力[14-15]。针对紫茎泽兰的严重入侵, 学者们对其进行了人工机械防治、化学防治[16]、引入天敌控制[17]及生物替代控制[18]等多种防控方法研究, 其中由于生物替代控制方法较生态环保且具有可持续性, 一直被研究关注。目前已筛选出一些对紫茎泽兰具有竞争力的潜在目标植物, 如非洲狗尾草(Setaria sphacelata)[19]、紫穗槐(Amorpha fruticosa)[4]和黄花蒿(Artemisia annua)[20]等。由于紫茎泽兰入侵生境复杂多样, 与不同生境中当地物种间的竞争关系也复杂多样, 因此针对紫茎泽兰在不同地区的入侵及替代控制还需开展更多的种间竞争关系相关研究, 尤其是其与本地植物间的竞争关系, 以筛选出更多的适合不同生境的本地替代目标物种。
活性炭由于具有吸附化感有机物质的特性, 可以削弱或消除入侵植物的化感物质对相邻植物的影响, 经常被用作验证入侵植物的化感作用或降低其化感作用的添加剂[2, 21]。有研究表明活性炭可以影响加拿大一枝黄花(Solidago canadensis)的化感作用而降低其竞争力[22]。土壤真菌可以与植物形成共生关系, 直接或间接影响土壤环境、养分形态从而影响植物根系对养分、水分的吸收及植物生长, 其中的AMF普遍存在于多数植物根系土壤中, 多数研究表明AMF可以影响入侵植物与本地植物的竞争关系[23-24]。李立青等[25]研究发现AMF可以在紫茎泽兰与本地植物竞争中起到显著促进作用;宋洁蕾等[26]研究发现添加AMF可以促进紫茎泽兰的生长; 李会娜等[27]研究发现添加杀真菌剂可以明显降低紫茎泽兰对伴生植物的竞争力。可见, 活性炭和杀真菌剂可能会影响紫茎泽兰与替代目标植物的竞争。
在野外调查中, 发现在紫茎泽兰入侵区域常有南酸枣(Choerospondias axillaris)、假地豆(Desmodium heterocarpon)和狗尾草(Setaria viridis)等植物存在, 这3种植物是在我国西南山地常见的本地植物, 它们对当地生境适应性强、生长快且具有一定的经济、生态或药用价值。因此, 本研究提出以下两个问题: (1) 这3种本地植物是否可以作为替代控制紫茎泽兰的目标植物;(2) 添加活性炭或杀真菌剂是否可以影响紫茎泽兰与这3种本地植物的竞争力从而促进本地植物的生长。其目的一方面为筛选替代控制紫茎泽兰的本地植物提供科学依据, 另一方面探讨能够提高替代控制入侵植物效率的方法。
1 实验材料与方法 1.1 实验材料实验所需的紫茎泽兰种子于2017年4月份采自广西隆林县紫茎泽兰严重入侵山地(105°7′55″E, 24°39′5″N, 1521 m a.s.l), 该地为退耕还林地, 土壤贫瘠, 林木覆盖率较低, 以马尾松为主, 林下紫茎泽兰覆盖率达85%以上, 伴有稀疏本地草本及灌木, 采集时每个植株至少间隔20 m。本地植物南酸枣、假地豆和狗尾草的种子在入侵区域采集, 其中南酸枣的种子在2016年的10—11月采集并沙埋储存, 假地豆和狗尾草的种子在2016年的9—11月采集, 装于信封置干燥室温储存。
1.2 实验方法实验在广西植物研究所生物入侵生态学实验基地采用盆栽法进行(位于桂林市雁山区雁山镇, 25°04′49.6″N, 110°18′01.8″E, 海拔170 m)。该地属中亚热带季风气候, 年平均气温17.8℃, 最冷月1月平均温度5.8℃, 最热月7月平均温度28℃;年平均降水量1949.5 mm。2017年4月中旬在温室大棚内进行入侵植物紫茎泽兰与本地植物育苗, 一个月后选择株高、基茎基本一致的幼苗移植到装有相同土壤的塑料盆内(内径24 cm, 底径16 cm, 高18 cm), 每盆装土约5 kg。根据Eller和Oliveira[28]将入侵植物与本地植物分别设置单种和混种处理, 单种为每种植物每盆种植2株植物, 混种处理为每盆种植1株入侵植物与1株本地植物。为了检测活性炭和杀真菌剂是否对入侵植物与本地植物的竞争有影响, 在入侵植物与本地植物混种时又分别设置添加活性炭(Activated carbon, AC, 纯活性炭, 固体粉末)、杀真菌剂(Fungicide, FC, 甲基硫菌灵, 有效成分含量70%, 可湿性粉剂, 日本日友商社(香港)有限公司生产)及同时添加活性炭和杀真菌剂(AC+FC)3种处理, 以不添加任何物质为对照处理(A0F0)。活性炭以20 mL/L土(约200 g/盆)的量添加[29], 杀真菌剂以0.15 g/kg土的量添加, 二者均分两次添加, 间隔15 d, 每种种植处理设置7个重复, 共种植112(4物种单种×7个重复+3物种对×4处理×7个重复)盆。实验期间每天浇足量的水, 保持盆内土壤湿润, 及时清除杂草和防治病虫害, 并每周随机移动花盆1次以避免位置效应。植物生长4个月左右进行收获, 记录每种植物的株高, 将地上茎叶和地下根分开收获, 并分别在60℃下烘干至恒重用电子天平称重(精确度为0.001), 计算其总生物量、地上生物量比(地上生物量/总生物量)和根冠比(地下生物量/地上生物量, R/C)。采用相对产量(RY)和竞争攻击力系数(A)[30]衡量物种间的竞争关系, 通过混种时添加以上物质处理下入侵植物与本地植物的生长变量与对照的差异衡量这些物质对入侵植物与本地植物竞争的影响。相对产量(RY)和竞争攻击力系数(A)根据以下公式计算:
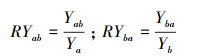
|
(1) |
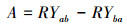
|
(2) |
式中, Yab为紫茎泽兰与本地种混种时单株生物量, Ya为紫茎泽兰单种时单株生物量;Yba为本地种与紫茎泽兰混种时单株生物量, Yb为本地种单种时单株生物量。
RYab < 1时, 紫茎泽兰对本地种的竞争力小于对紫茎泽兰本身的竞争力, 紫茎泽兰受本地物种的竞争影响比其本身大, 即种间竞争大于种内竞争;RYab=1时, 2个种的竞争力水平相当, 即种间竞争等于种内竞争;RYab>1时, 紫茎泽兰对本地物种的竞争力大于对紫茎泽兰本身的竞争力, 紫茎泽兰受本地植物的竞争影响比其本身小, 即种内竞争大于种间竞争。
A < 0表明紫茎泽兰比本地植物竞争力弱;A=0表明紫茎泽兰与本地种竞争力相当;A>0表明紫茎泽兰比本地植物竞争力强。本地植物对紫茎泽兰的竞争攻击力系数与紫茎泽兰对本地植物的竞争攻击力系数相反, 在此不再作详细说明。
1.3 数据分析所有数据分析均在SPSS 17.0软件中进行, 利用One-way ANOVA方差分析并结合最小显著差异法LSD检验不添加任何物质条件下紫茎泽兰在与不同本地植物混种与单种时的形态和生长变量的差异性及混种时入侵植物与本地植物在添加活性炭或杀真菌剂处理下的生长变量与对照的差异性, 当检验变量方差不齐时对其进行平方根或自然对数转换;利用独立样本t-test检验本地植物的生长变量在混种与单种时的差异性;利用单样本t检验分析相对产量RY与1及攻击力系数A与0之间的差异显著性, 以上分析差异性显著水平均为0.05。利用Excel 2007软件进行作图。
2 研究结果 2.1 不添加任何物质条件下本地植物与紫茎泽兰混种时的生长变量紫茎泽兰分别与南酸枣、假地豆和狗尾草混种时, 其株高均显著低于单种(P < 0.05), 分别比单种时降低了14.3%、52.07%和31.97%;在与南酸枣混种时, 紫茎泽兰的生物量显著高于单种(P < 0.05), 比单种时增加了43.54%, 而在与假地豆和狗尾草混种时, 其生物量均显著低于单种(P < 0.05), 分别比单种时降低了90.03%和61.67%;紫茎泽兰的地上生物量比仅在与狗尾草混种时显著低于单种(P < 0.05), 其根冠比在与假地豆和狗尾草混种时均显著高于单种(P < 0.05)(图 1)。

|
| 图 1 不添加任何物质条件下紫茎泽兰单种与混种时的生长变量 Fig. 1 Growth variables of A. adenophora when grown alone and grown with native plants under no addition of activated carbon or fungicide treatment Z紫茎泽兰A. adenophora; N: 南酸枣C. axillaris; J: 假地豆D. heterocarpon; G: 狗尾草S. viridis; 不同小写字母表示不同处理间差异显著(P < 0.05), 图中数据为平均值±标准误 |
南酸枣与紫茎泽兰混种时, 其生物量和地上生物量比均显著低于单种(P < 0.05), 分别比单种时降低了28.82%和15.35%;而根冠比显著高于单种(P < 0.05), 比单种时增加了29.34%。假地豆与紫茎泽兰混种时, 其生物量与单种时无明显差异;株高和地上生物量比均显著低于单种(P < 0.05), 分别比单种时降低了29.89%和18.05%;而根冠比显著高于单种, 比单种时增加了87.82%。狗尾草与紫茎泽兰混种时, 其株高和生物量均显著高于单种(P < 0.05), 分别比单种时增加了21.43%和173.53%;地上生物量比和根冠比均与单种无显著差异(P>0.05)(图 2)。

|
| 图 2 不添加任何物质条件下本地植物单种与混种时的生长变量 Fig. 2 Growth variables of native plants when grown alone and grown with A. adenophora under no addition of activated carbon or fungicide treatment 不同小写字母表示同一物种在单种与混种间的差异显著性(P < 0.05), 图中数据为平均值±标准误 |
南酸枣与紫茎泽兰混种时各自的竞争参数: 南酸枣对紫茎泽兰的竞争参数RY显著小于1(P < 0.05), 表现为种间竞争大于种内竞争;紫茎泽兰的RY显著大于1(P < 0.05), 表现为种内竞争大于种间竞争;南酸枣对紫茎泽兰的竞争参数A显著小于0(P < 0.05), 相反紫茎泽兰的A显著大于0(P < 0.05)(图 3), 说明紫茎泽兰的竞争力强于南酸枣。

|
| 图 3 不添加任何物质条件下本地植物与紫茎泽兰的竞争参数 Fig. 3 Competitive parameters of A. adenophora and native plants under no addition of activated carbon or fungicide treatment *表示相对产量RY或竞争攻击力系数A分别与1或0相比差异显著(P < 0.05), **表示RY或A分别与1或0相比差异极显著(P < 0.01); 图中数据为平均值±标准误 |
假地豆和狗尾草分别与紫茎泽兰混种时各自的竞争参数: 假地豆和狗尾草对紫茎泽兰的竞争参数RY均明显大于1(P < 0.05), 表现为种内竞争大于种间竞争;紫茎泽兰对它们的竞争参数RY均显著小于1(P < 0.05), 表现为种间竞争大于种内竞争;假地豆和狗尾草对紫茎泽兰的竞争参数A均显著大于0(P < 0.05), 相反紫茎泽兰的A均显著小于0(P < 0.05)(图 3), 说明紫茎泽兰的竞争力弱于假地豆和狗尾草。
2.3 活性炭和杀真菌剂对混种时本地植物与紫茎泽兰的生长变量的影响南酸枣与紫茎泽兰混种时, 与对照相比, AC、FC和AC+FC处理均显著降低了紫茎泽兰的地上生物量比(P < 0.05), 增加了其根冠比, 但仅AC和AC+FC处理增加的显著(P < 0.05);3种处理均降低了南酸枣的生物量, 且AC和FC处理降低的显著(P < 0.05)(图 4)。假地豆与紫茎泽兰混种时, 与对照相比, 仅FC处理显著增加了紫茎泽兰的株高(P < 0.05);AC和AC+FC处理均显著增加了假地豆的株高和生物量(P < 0.05), 同时, 这两种处理增加了假地豆的地上生物量比, 降低了其根冠比, 但仅AC+FC处理增加或降低的显著(P < 0.05) (图 5)。狗尾草与紫茎泽兰混种时, 与对照相比, AC和AC+FC处理均显著降低了紫茎泽兰的地上生物量比、增加了其根冠比(P < 0.05);3种处理均降低了狗尾草的生物量和根冠比, 但均不显著(P>0.05)(图 6)。

|
| 图 4 活性炭和/或杀真菌剂对紫茎泽兰和南酸枣混种时生长变量的影响 Fig. 4 Effects of activated carbon and/or fungicide on the growth variables of A. adenophora and C. axillaris in mixculture A0F0: 不添加任何物质No addition; AC: 活性炭Activated carbon; FC: 杀真菌剂Fungicide; 不同小写字母表示同一物种在不同处理间的差异显著(P < 0.05), 图中数据为平均值±标准误 |

|
| 图 5 活性炭和/或杀真菌剂对紫茎泽兰和假地豆混种时生长变量的影响 Fig. 5 Effects of activated carbon and/or fungicide on the growth variables of A. adenophora and D. heterocarpon in mixculture 不同小写字母表示同一物种在不同处理间的差异显著(P < 0.05), 图中数据为平均值±标准误 |

|
| 图 6 活性炭和/或杀真菌剂对紫茎泽兰和狗尾草混种时生长变量的影响 Fig. 6 Effects of activated carbon and/or fungicide on the growth variables of A. adenophora and S. viridis in mixculture 不同小写字母表示同一物种在不同处理间的差异显著(P < 0.05), 图中数据为平均值±标准误 |
一般认为外来入侵植物具有较强的竞争能力是其在新的入侵群落中成功定居、扩散从而成为单优种群的主要原因之一[31-32]。寻找竞争力较强、能够替代控制入侵植物的本地植物对入侵地的生态修复具有重要意义。株高和生物量能够体现植物吸收利用资源的能力, 在一定程度上可以反映植物竞争能力的大小[33-34]。在本研究中, 入侵植物紫茎泽兰的株高和生物量被本地植物假地豆和狗尾草显著抑制;而假地豆的生物量不受紫茎泽兰的竞争影响;狗尾草的株高和生物量在竞争时被显著促进, 反映出这两个本地植物对资源吸收利用的能力强于紫茎泽兰。同时它们对紫茎泽兰的竞争攻击力系数均显著大于0, 也说明它们的竞争能力强于紫茎泽兰。在紫茎泽兰与南酸枣混种时, 紫茎泽兰的生物量显著高于单种, 且其对南酸枣的竞争攻击力系数显著大于0, 而南酸枣的生物量显著低于单种, 说明南酸枣的资源吸收利用能力和竞争力均弱于紫茎泽兰。
生物替代控制是根据植物间竞争关系的原理实施的一种生态控制方法, 其关键就是找到能够竞争抵御入侵植物的物种。目前已有较多学者通过研究物种间的竞争能力筛选可以作为替代控制入侵植物的目标植物, 如甘薯(Ipomoea batatas)可替代控制薇甘菊(Mikania micrantha)[35]、马唐(Digitaria sanguinalis)和牛筋(Eleusine indica)可替代控制黄顶菊[30]、杂交狼尾草(Pennisetum americanum×P. purpureum)和木豆(Cajanus cajan)可替代控制飞机草(Eupatorium odoratum)[36]。但卢向阳等[18]发现, 竞争力强于紫茎泽兰的非洲狗尾草(Setaria anceps)和多年生黑麦草(Lolium perenne), 由于生态适应性差而导致其对紫茎泽兰的控制效果大大减弱。因此认为, 替代控制入侵植物的目标植物不仅要有强的竞争力, 还应具有优良的生态适应性、覆盖性、化感耐受性、可持续性、经济或生态价值等特征。本研究中, 假地豆是一种本地的豆科灌木或亚灌木, 适应于山坡、荒山、草地等生境, 具直立和平卧生长的特性, 叶片茂盛、枝条多而密集, 覆盖性较好, 种子产量较多, 易形成稳定的种群, 且之前的研究发现该植物对入侵植物飞机草的竞争力也较强[37], 因此认为该植物可以作为替代控制紫茎泽兰的目标植物。狗尾草是本地丛生型禾本科植物, 多生于荒田、荒地、山坡等生境, 种子产量多, 适应性强, 常作为牲畜饲料, 还具有一定的药用价值, 且周泽建等[38]研究表明该植物在不同肥力条件下对紫茎泽兰的控制效果都较好。因此, 在清除紫茎泽兰后的荒田荒地种植该植物, 其可以凭借优先占据生态位和强的竞争力在一定程度上控制紫茎泽兰的生长及二次入侵。南酸枣竞争力表现较弱, 可能与该植物为乔木, 在幼苗早期相对于草本植物紫茎泽兰的生长速率较慢有关, 因此, 该植物幼苗期不宜用于替代控制紫茎泽兰。
3.2 活性炭对本地植物与紫茎泽兰竞争的影响化感作用是多数入侵植物具有的共性, 且这一特性可以协同促进入侵植物对其他植物的竞争[8, 39]。研究表明活性炭可以在一定程度上吸附化感物质, 减弱入侵植物的化感作用, 提高本地植物的生长和竞争能力[29, 40]。本研究中, 在不同本地植物与紫茎泽兰混种时, 添加活性炭降低了南酸枣的生物量, 增加了假地豆的株高和生物量, 而对狗尾草的生长变量无明显影响;对紫茎泽兰的株高和生物量均无明显影响, 仅在紫茎泽兰与南酸枣或狗尾草混种时, 降低了紫茎泽兰的地上生物量比、增加了其根冠比。可见, 活性炭对混种时的紫茎泽兰和不同本地植物生长的影响作用不同。这与前者研究发现活性炭可以吸附化感物质降低入侵植物的竞争力或促进本地植物生长的研究结果不完全一致。其原因一方面可能是不同物种对入侵植物化感物质的敏感度不同, 有些本地植物对入侵植物的化感物质具有耐受性, 甚至本地植物也可以释放化感物质影响入侵植物的生长或竞争[37, 41], 因而活性炭在入侵植物与不同本地植物竞争时表现的影响作用就不同。Zheng等[42]研究也表明活性炭处理的不同本地植物的群落对飞机草的入侵响应结果不一致。另一方面入侵植物与本地植物竞争时是否会释放化感物质及释放多少与其竞争对象、生长时间及竞争时土壤养分条件等多种因素相关, 如反枝苋(Amaaranthus retroflexus)的化感作用以幼苗期最强, 现蕾期次之, 成熟期最弱[43], 且其与相邻植物竞争时在低肥作用下的化感作用强于高肥下的[29];紫茎泽兰化感作用在生长早期很弱, 随着发育期的推进而逐渐增强[44]。本研究中紫茎泽兰生长期为4个月, 可能在与不同竞争力的本地植物混种时分泌的化感物质量不同。此外, Nolan等[45]研究认为活性炭能降低入侵植物的生长是由于植物与土壤微生物相互作用调节导致的, 而不是直接由于降低化感物质所致, 说明活性炭在入侵植物与本地植物竞争中的作用可能还与土壤微生物活动相关。Lau等[46]研究认为活性炭除了可以吸附化感物质外, 还可以提高土壤中氮及其他养分的浓度从而影响植物生长, 利用活性炭对植物生长的影响推断化感作用是复杂的。可见, 活性炭对外来物种的入侵及其与本地植物竞争的影响与多种因素相关, 是十分复杂的。在替代控制入侵植物时能否将其用于提高替代控制效率还需要针对具体的目标物种展开详细的研究。
3.3 杀真菌剂对本地植物与紫茎泽兰竞争的影响土壤真菌可以通过菌丝网络将营养物质在不同植物间交流, 从而影响群落中植物生长和物种的组成, 其中的AMF种群被多数研究证实对外来植物的入侵与竞争有正反馈调节作用[47-49], 被认为是外来植物成功入侵的重要驱动因子[23, 50]。研究表明土壤AMF可以与紫茎泽兰形成共生关系, 促进紫茎泽兰对本地植物的竞争[25]; 还可以通过菌根网络缓解营养元素的限制增加紫茎泽兰对养分的竞争力[51]; 添加杀真菌剂可以明显降低紫茎泽兰对伴生植物的竞争力[27]。然而, 本研究中, 在紫茎泽兰与不同本地植物竞争时, 添加杀真菌剂对混种时的紫茎泽兰与本地植物的多数生长变量均未产生显著影响, 甚至在紫茎泽兰与南酸枣混种时还降低了本地植物南酸枣的生物量, 在与假地豆混种时增加了紫茎泽兰的株高。这与之前的研究结果不符, 分析其原因, 一方面可能是土壤AMF与植物生长的相互反馈及其对植物间竞争关系的影响作用受多种因素影响, 如入侵植物本身(发育时期、入侵阶段等)、AMF本身(AMF种类、起源等)、伴生植物种类及土壤肥力等[49]。Barto等[52]发现, 土壤AMF可以与本地植物形成正反馈抵御入侵植物的入侵;Waller等[53]发现在入侵植物矢车菊(Centaurea solstitialis) 与北美本地植物针茅(Stipa pulchra) 竞争时, 添加AMF抑制了该入侵植物的生物量, 认为在竞争时减少菌根共生可能也会对入侵植物有利;Chen等[54]研究发现土壤AMF仅在低磷下可以提高入侵植物的竞争优势。另一方面, 杀真菌剂处理中的紫茎泽兰失去了与之共生的土壤真菌后, 可能会改变或调节土壤中其他微生物维持其对本地植物的竞争。有研究表明紫茎泽兰的根系生命活动能促进根际微生物的生长繁殖、活性增强, 从而促进紫茎泽兰的生长和竞争[55]。此外, 杀真菌剂除了消除对入侵植物有正反馈作用的真菌外, 可能也消除了一些对植物生长有负反馈作用的病原真菌, 尽管有研究认为入侵植物在入侵地可以快速积累本地病原菌使本地植物生长受到抑制, 从而提高其相对竞争优势, 即“本地病原菌积累假说”[56-57], 还有研究表明入侵植物可以携带病原菌感染本地植物从而增加自身的竞争优势[58], 但也有少部分研究认为本地病原菌对入侵植物也有一定的致病作用[59]。然而, 本研究在未添加杀真菌剂的对照处理中未发现病原菌对植物的影响症状, 因此病原菌影响本文研究结果的可能性较小。Zheng等[42]研究也发现添加杀真菌剂对飞机草的入侵没有影响。可见, 添加杀真菌剂是否可以影响入侵植物与本地植物的竞争需要根据具体的竞争对象及入侵生境等多种因素做深入分析研究。
4 结论紫茎泽兰是一种具有较强竞争力和化感作用且难以清除的入侵植物, 本研究中3种本地植物对紫茎泽兰的竞争影响不同, 假地豆和狗尾草的竞争可以降低紫茎泽兰的株高和生物量, 其竞争力强于紫茎泽兰, 根据它们的生长特征和生态适应特性, 可以在一定区域替代控制紫茎泽兰。南酸枣在幼苗期竞争力弱于紫茎泽兰, 因此, 幼苗期的南酸枣不宜用于抵御紫茎泽兰。活性炭和杀真菌剂在紫茎泽兰与不同本地植物竞争中对不同植物生长的影响不同, 说明活性炭和杀真菌剂是否能够提高替代控制紫茎泽兰的控制效率取决于具体的目标物种。总之, 研究结果回答了开始提出的两个问题, 也达到了预期的目的。此外, 由于本实验在单种处理中, 没有设置添加活性炭和杀真菌剂的处理, 因此, 仅评估了添加活性炭和杀真菌剂对紫茎泽兰和本地植物混种时的形态和生长的影响, 未能评估这些物质对竞争指标如RY和A的影响。但研究可以为筛选替代控制紫茎泽兰的本地植物及提高替代控制效率的方法提供科学依据。
| [1] |
Blossey B, Nötzold R. Evolution of increased competitive ability in invasive nonindigenous plants: a hypothesis. Journal of Ecology, 1995, 83(5): 887-889. DOI:10.2307/2261425 |
| [2] |
Gruntman M, Pehl A K, Joshi S, Tielbörger K. Competitive dominance of the invasive plant Impatiens glandulifera: using competitive effect and response with a vigorous neighbor. Biological Invasions, 2014, 16(1): 141-151. DOI:10.1007/s10530-013-0509-9 |
| [3] |
马杰, 皇甫超河, 易津, 杨殿林. 4种替代植物对黄顶菊入侵土壤养分和酶活性的影响. 生态环境学报, 2011, 20(5): 805-812. DOI:10.3969/j.issn.1674-5906.2011.05.003 |
| [4] |
卢向阳, 王秋霞, 刘冰, 俞晶山, 杨万吉. 紫穗槐替代控制对撂荒山地紫茎泽兰的影响. 西南农业学报, 2013, 26(5): 1893-1898. |
| [5] |
Callaway R M, Ridenour W M. Novel weapons: invasive success and the evolution of increased competitive ability. Frontiers in Ecology and the Environment, 2004, 2(8): 436-443. DOI:10.1890/1540-9295(2004)002[0436:NWISAT]2.0.CO;2 |
| [6] |
Inderjit, Evans H, Crocoll C, Bajpai D, Kaur R, Feng Y L, Silva C, Carreón J T, Valiente-Banuet A, Gershenzon J, Callaway R M. Volatile chemicals from leaf litter are associated with invasiveness of a Neotropical weed in Asia. Ecology, 2011, 92(2): 316-324. DOI:10.1890/10-0400.1 |
| [7] |
Qin R M, Zheng Y L, Valiente-Banuet A, Callaway R M, Barclay G F, Pereyra C S, Feng Y L. The evolution of increased competitive ability, innate competitive advantages, and novel biochemical weapons act in concert for a tropical invader. New Phytologist, 2013, 197(3): 979-988. DOI:10.1111/nph.12071 |
| [8] |
Zheng Y L, Feng Y L, Zhang L K, Callaway R M, Valiente-Banuet A, Luo D Q, Liao Z Y, Lei Y B, Barclay G F, Silva-Pereyra C. Integrating novel chemical weapons and evolutionarily increased competitive ability in success of a tropical invader. New Phytologist, 2015, 205(3): 1350-1359. DOI:10.1111/nph.13135 |
| [9] |
Paudel S, Baer S G, Battaglia L L. Arbuscular mycorrhizal fungi (AMF) and success of Triadica sebifera invasion in coastal transition ecosystems along the northern Gulf of Mexico. Plant and Soil, 2014, 378(1/2): 337-349. |
| [10] |
Zhang F J, Li Q, Yerger E H, Chen X, Shi Q, Wan F H. AM fungi facilitate the competitive growth of two invasive plant species, Ambrosia artemisiifolia and Bidens pilosa. Mycorrhiza, 2018, 28(8): 703-715. DOI:10.1007/s00572-018-0866-4 |
| [11] |
肖正清, 周冠华, 权文婷. 恶性外来入侵植物紫茎泽兰在云南的分布格局. 自然灾害学报, 2009, 18(5): 82-87. DOI:10.3969/j.issn.1004-4574.2009.05.013 |
| [12] |
马金虎, 杨文秀, 孙亮亮, 陈皓, 赵倩, 杨小环. 紫茎泽兰提取物对3种杂草的化感胁迫作用. 生态学报, 2018, 38(10): 3514-3523. |
| [13] |
雷桂生, 王五云, 蒋智林, 邓丹丹, 刘万学, 桂富荣, 李正跃. 紫茎泽兰与伴生植物小藜的竞争效应及其生理生化特征. 生态环境学报, 2014, 23(1): 16-21. DOI:10.3969/j.issn.1674-5906.2014.01.003 |
| [14] |
牛红榜, 刘万学, 万方浩. 紫茎泽兰(Ageratina adenophora)入侵对土壤微生物群落和理化性质的影响. 生态学报, 2007, 27(7): 3051-3060. DOI:10.3321/j.issn:1000-0933.2007.07.047 |
| [15] |
Yu W Q, Liu W X, Liu W Z, Ren Y J, Wan F H, Zhang L L. Positive feedback of soil fungi, including arbuscular mycorrhizal fungi, to the invasive weed Ageratina adenophora: evidence from field studies. Journal of Biosafety, 2014, 23(1): 4-12. |
| [16] |
翟鸣, 熊鑫, 田茂娟. 紫薇清除治紫茎泽兰林间药效试验. 贵州林业科技, 2018, 46(1): 39-42. |
| [17] |
Niu Y F, Feng Y L, Xie J L, Luo F C. Noxious invasive Eupatorium adenophorum may be a moving target: implications of the finding of a native natural enemy, Dorylus orientalis. Chinese Science Bulletin, 2010, 55(33): 3743-3745. DOI:10.1007/s11434-010-4117-0 |
| [18] |
卢向阳, 张锦华, 左相兵, 刘冰, 郑雅婧. 几种替代植物对入侵杂草紫茎泽兰的防控效果. 贵州农业科学, 2012, 40(6): 103-106, 109-109. DOI:10.3969/j.issn.1001-3601.2012.06.028 |
| [19] |
蒋智林, 刘万学, 万方浩, 李正跃. 非洲狗尾草与紫茎泽兰的竞争效应. 中国农业科学, 2008, 41(5): 1347-1354. DOI:10.3864/j.issn.0578-1752.2008.05.012 |
| [20] |
孙娜娜, 谭永钦, 马洪菊, 周兴苗, 王永模. 黄花蒿对紫茎泽兰竞争效应的影响. 华中农业大学学报, 2015, 34(1): 38-44. |
| [21] |
Del Fabbro C, Güsewell S, Prati D. Allelopathic effects of three plant invaders on germination of native species: a field study. Biological Invasions, 2014, 16(5): 1035-1042. DOI:10.1007/s10530-013-0555-3 |
| [22] |
Yuan Y G, Tang J J, Leng D, Hu S J, Yong J W H, Chen X. An invasive plant promotes its arbuscular mycorrhizal symbioses and competitiveness through its secondary metabolites: indirect evidence from activated carbon. PLoS One, 2014, 9(5): e97163. DOI:10.1371/journal.pone.0097163 |
| [23] |
Dong L J, Ma L N, He W M. Arbuscular mycorrhizal fungi help explain invasion success of Solidago canadensis. Applied Soil Ecology, 2021, 157: 103763. DOI:10.1016/j.apsoil.2020.103763 |
| [24] |
Zhang F J, Li Q, Chen F X, Xu H Y, Inderjit, Wan F H. Arbuscular mycorrhizal fungi facilitate growth and competitive ability of an exotic species Flaveria bidentis. Soil Biology and Biochemistry, 2017, 115: 275-284. DOI:10.1016/j.soilbio.2017.08.019 |
| [25] |
李立青, 张明生, 梁作盼, 肖博, 万方浩, 刘万学. 丛枝菌根真菌促进入侵植物紫茎泽兰的生长和对本地植物竞争效应. 生态学杂志, 2016, 35(1): 79-86. |
| [26] |
宋洁蕾, 李胜, 林莹, 罗石文, 段艳茹, 桂富荣. 丛枝菌根真菌对紫茎泽兰生长与繁殖的影响. 生物安全学报, 2019, 28(1): 44-48. DOI:10.3969/j.issn.2095-1787.2019.01.009 |
| [27] |
李会娜, 刘万学, 万方浩. 杀真菌剂对入侵植物紫茎泽兰与伴生植物生长的反馈作用. 中国农业科学, 2011, 44(11): 2269-2275. DOI:10.3864/j.issn.0578-1752.2011.11.008 |
| [28] |
Eller C B, Oliveira R S. Effects of nitrogen availability on the competitive interactions between an invasive and a native grass from Brazilian cerrado. Plant and Soil, 2017, 410(1/2): 63-72. |
| [29] |
鲁萍, 干珠扎布, 梁慧, 白雅梅, 徐广惠, 田秋阳, 陈春锋, 王宏燕. 不同供氮水平下反枝苋的化感作用与资源竞争能力. 生态学杂志, 2011, 30(8): 1590-1597. |
| [30] |
吕远, 王贵启, 郑丽, 倪汉文. 入侵植物黄顶菊与本地植物的竞争. 生态学杂志, 2011, 30(4): 677-681. |
| [31] |
Gioria M, Osborne B A. Resource competition in plant invasions: emerging patterns and research needs. Frontiers in Plant Science, 2014, 5: 501. |
| [32] |
Kuebbing S E, Nuñez M A. Invasive non-native plants have a greater effect on neighbouring natives than other non-natives. Nature Plants, 2016, 2(10): 16134. DOI:10.1038/nplants.2016.134 |
| [33] |
Keddy P, Nielsen K, Weiher E, Lawson R. Relative competitive performance of 63 species of terrestrial herbaceous plants. Journal of Vegetation Science, 2002, 13(1): 5-16. DOI:10.1111/j.1654-1103.2002.tb02018.x |
| [34] |
李慧燕, 陈冬青, 王慧, 杨殿林, 皇甫超河. 不同混种密度下杀真菌剂对黄顶菊与反枝苋种间竞争的影响. 生态学杂志, 2015, 34(4): 1013-1018. |
| [35] |
Shen S C, Xu G F, Clements D R, Jin G M, Chen A D, Zhang F D, Kato-Noguchi H. Suppression of the invasive plant mile-a-minute (Mikania micrantha) by local crop sweet potato (Ipomoea batatas) by means of higher growth rate and competition for soil nutrients. BMC Ecology, 2015, 15(1): 1. DOI:10.1186/s12898-014-0033-5 |
| [36] |
岳茂峰, 崔烨, 冯莉, 张纯, 田兴山. 入侵植物飞机草与4种牧草的竞争效应. 生物安全学报, 2016, 25(4): 270-274. DOI:10.3969/j.issn.2095-1787.2016.04.006 |
| [37] |
潘玉梅, 唐赛春, 韦春强, 李象钦, 吕仕洪, 王云波. 两种本地植物种子萌发对飞机草的化感耐受性及其相互竞争. 生态学报, 2019, 39(19): 7302-7310. |
| [38] |
周泽建, 周美兰, 刘万学. 三种植物在不同肥力下对紫茎泽兰的生态控制效果. 中国农学通报, 2006, 22(6): 361-364. DOI:10.3969/j.issn.1000-6850.2006.06.092 |
| [39] |
Ridenour W M, Callaway R M. The relative importance of allelopathy in interference: the effects of an invasive weed on a native bunchgrass. Oecologia, 2001, 126(3): 444-450. DOI:10.1007/s004420000533 |
| [40] |
Tian Y H, Feng Y L, Liu C. Addition of activated charcoal to soil after clearing Ageratina adenophora stimulates growth of forbs and grasses in China. Tropical Grasslands, 2007, 41(4): 285-291. |
| [41] |
潘玉梅, 唐赛春, 韦春强, 林春华. 土著植物黄荆条水提取液对飞机草生长和光合特性的化感效应. 生态学杂志, 2013, 32(2): 351-357. |
| [42] |
Zheng Y L, Burns J H, Liao Z Y, Li Y P, Yang J, Chen Y J, Zhang J L, Zheng Y G. Species composition, functional and phylogenetic distances correlate with success of invasive Chromolaena odorata in an experimental test. Ecology Letters, 2018, 21(8): 1211-1220. DOI:10.1111/ele.13090 |
| [43] |
刘爽, 马丹炜. 不同发育期反枝苋对黄瓜根缘细胞的化感作用. 生态学报, 2009, 29(8): 4392-4396. DOI:10.3321/j.issn:1000-0933.2009.08.045 |
| [44] |
韩利红, 冯玉龙. 发育时期对紫茎泽兰化感作用的影响. 生态学报, 2007, 27(3): 1185-1191. DOI:10.3321/j.issn:1000-0933.2007.03.045 |
| [45] |
Nolan N E, Kulmatiski A, Beard K H, Norton J M. Activated carbon decreases invasive plant growth by mediating plant-microbe interactions. AoB Plants, 2015, 7: plu072. DOI:10.1093/aobpla/plu072 |
| [46] |
Lau J A, Puliafico K P, Kopshever J A, Steltzer H, Jarvis E P, Schwarzländer M, Strauss S Y, Hufbauer R A. Inference of allelopathy is complicated by effects of activated carbon on plant growth. New Phytologist, 2008, 178(2): 412-423. DOI:10.1111/j.1469-8137.2007.02360.x |
| [47] |
Menzel A, Hempel S, Klotz S, Moora M, Pyšek P, Rillig M C, Zobel M, Kühn I. Mycorrhizal status helps explain invasion success of alien plant species. Ecology, 2017, 98(1): 92-102. DOI:10.1002/ecy.1621 |
| [48] |
Awaydul A, Zhu W Y, Yuan Y G, Xiao J, Hu H, Chen X, Koide R T, Cheng L. Common mycorrhizal networks influence the distribution of mineral nutrients between an invasive plant, Solidago canadensis, and a native plant, Kummerowa striata. Mycorrhiza, 2019, 29(1): 29-38. DOI:10.1007/s00572-018-0873-5 |
| [49] |
于文清, 周文, 万方浩, 刘万学. 丛枝菌根真菌(AMF)对外来植物入侵反馈机制的研究进展. 生物安全学报, 2012, 21(1): 1-8. |
| [50] |
Dawson W, Schrama M. Identifying the role of soil microbes in plant invasions. Journal of Ecology, 2016, 104(5): 1211-1218. DOI:10.1111/1365-2745.12619 |
| [51] |
Shen K P, Cornelissen J H C, Wang Y J, Wu C B, He Y J, Ou J, Tan Q Y, Xia T T, Kang L L, Guo Y, Wu B L. Am fungi alleviate phosphorus limitation and enhance nutrient competitiveness of invasive plants via mycorrhizal networks in karst areas. Frontiers in Ecology and Evolution, 2020, 8: 125. DOI:10.3389/fevo.2020.00125 |
| [52] |
Barto K, Friese C, Cipollini D. Arbuscular mycorrhizal fungi protect a native plant from allelopathic effects of an invader. Journal of Chemical Ecology, 2010, 36(4): 351-360. DOI:10.1007/s10886-010-9768-4 |
| [53] |
Waller L P, Callaway R M, Klironomos J N, Ortega Y K, Maron J L. Reduced mycorrhizal responsiveness leads to increased competitive tolerance in an invasive exotic plant. Journal of Ecology, 2016, 104(6): 1599-1607. DOI:10.1111/1365-2745.12641 |
| [54] |
Chen E J, Liao H X, Chen B M, Peng S L. Arbuscular mycorrhizal fungi are a double-edged sword in plant invasion controlled by phosphorus concentration. New Phytologist, 2020, 226(2): 295-300. DOI:10.1111/nph.16359 |
| [55] |
刘海, 王玉书, 焦玉洁, 彭丽媛, 郭明全, 王勇, 陈玉蓝, 袁玲. 三种土壤条件下紫茎泽兰根际的酶活性及细菌群落状况. 生态学报, 2018, 38(23): 8455-8465. |
| [56] |
Eppinga M B, Rietkerk M, Dekker S C, De Ruiter P C, van der Putten W H. Accumulation of local pathogens: a new hypothesis to explain exotic plant invasions. Oikos, 2006, 114(1): 168-176. DOI:10.1111/j.2006.0030-1299.14625.x |
| [57] |
Mangla S, Inderjit, Callaway R M. Exotic invasive plant accumulates native soil pathogens which inhibit native plants. Journal of Ecology, 2008, 96(1): 58-67. |
| [58] |
Li H, Zhang X M, Zheng R S, Li X, Elmer W H, Wolfe L M, Li B. Indirect effects of non-native Spartina alterniflora and its fungal pathogen (Fusarium palustre) on native saltmarsh plants in China. Journal of Ecology, 2014, 102(5): 1112-1119. DOI:10.1111/1365-2745.12285 |
| [59] |
Nijjer S, Rogers W E, Siemann E. Negative plant-soil feedbacks may limit persistence of an invasive tree due to rapid accumulation of soil pathogens. Proceedings of the Royal Society B: Biological Sciences, 2007, 274(1625): 2621-2627. DOI:10.1098/rspb.2007.0804 |