 2022, Vol. 42
2022, Vol. 42文章信息
- 郝庆丽, 任卓菲, 刘刚, 王立雄, 于娟, 郁泽君
- HAO Qingli, REN Zhuofei, LIU Gang, WANG Lixiong, YU Juan, YU Zejun
- 光和噪声污染胁迫下城市生态斑块鸟类风险评价
- Bird risk assessment in urban ecological patch under light pollution and noise pollution stress
- 生态学报. 2022, 42(6): 2186-2201
- Acta Ecologica Sinica. 2022, 42(6): 2186-2201
- http://dx.doi.org/10.5846/stxb202102220491
-
文章历史
- 收稿日期: 2021-02-22
- 网络出版日期: 2021-11-26

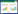




2. 天津市建筑物理环境与生态技术重点实验室, 天津 300072
2. Tianjin Key Laboratory of Architectural Physical Environment and Ecological Technologies, Tianjin 300072, China
人类活动产生的光污染和噪声污染快速增长、扩张, 侵入鸟类生存和发展的生态斑块, 改变其赖以生存的自然光、声环境[1-2]。研究表明, 光、声污染胁迫可以通过改变鸟类对环境的感知进而影响鸟类行为和生理[3]。鸟类基于自然光进化出复杂、互补的光感知系统和磁场感知系统[4-5]。人工光会影响鸟类光和磁场感知系统, 进而改变鸟类生理节律[6-7]、基因表达[8]、激素分泌[9]、能量代谢[10]等方面。另一方面, 鸟类的听觉与鸣叫行为密切相关, 正常的听觉输入对于鸟类鸣叫正常水平的维持至关重要[11]。交通噪声会损害鸟类听觉信号感知, 改变鸟类鸣声特性(振幅和频率), 从而破坏鸟类交流能力[12]以及觅食或躲避掠食者的能力[13], 影响鸟类生理和繁殖行为[14], 造成鸟类栖息地丧失或破碎[15], 降低鸟类种群密度和物种丰度[16]。人工光和交通噪声或将随城市扩张而继续蔓延, 对鸟类的影响也会不断扩大。因此, 评价光污染和噪声污染造成鸟类生态风险是必要而迫切的。
在光污染生态风险评价研究领域, 对微观的鸟种“光刺激-行为反应”的定量化实验和宏观的光污染生态风险评价两方面均有一定研究。在鸟类光刺激实验研究方面, 国内外学者获得了鸟类行为和光环境特性(光谱、光照强度等)之间的定量化规律[17]。Poot等[18]研究发现, 夜间迁徙的鸟类依赖于蓝光和绿光定向, 而容易被红光诱陷;Evans[19]和Zhao等[20]研究表明, 蓝光、绿光会对鸟类飞行产生风险。Wolfgang Wiltschko等[21]研究发现, 565 nm绿光辐照度达到15 mW/m2时, 澳大利亚绣眼鸟在该光照的影响下迁徙方向发生变化。Raap等[7]研究表明1.6 lx LED照明会改变雌性大山雀的睡眠节律。但目前, 鸟类“光刺激-行为反应”实验中光环境的评价指标与宏观夜光遥感数据的评价指标不一致;同时, 实验光环境与真实夜间光环境的光谱能量分布存在差异, 故实验研究结论难以全面反映城市环境中鸟类受到人工光的影响。在宏观光污染生态风险评价方面, 夜光遥感为大范围评估光污染的生态风险提供了可能。Horton等[22]结合VIIRS/DNB夜光数据和雷达监测的候鸟飞行数量, 研究了美国候鸟每年暴露在光污染下最多的位置和时间;Xue等[23]利用Jilin 1-3B夜光数据结合模拟获得的鸟类活动区域分布图, 通过自然断点法推测了鸟类活动区域的光污染等级。然而, 上述光污染评价研究未与鸟类敏感响应特征建立直接关系, 采用自然断点法的城市区域光污染等级划分未必能反映鸟类面临的光污染生态风险程度。
在声污染生态风险评价领域, 道路噪声对鸟类影响机理及宏观的声环境评价均有研究。已有学者通过研究获得了鸟类敏感的声压级阈值范围, 多项研究表明, 超过45 dBA SPL(re 20 μPa)的交通噪音会改变鸟类鸣声特征、繁殖、应激激素水平和物种丰富度[14]。另有研究表明, 不同鸟类对于环境的声压级偏好各不相同[24], 因此需要针对本土典型鸟种开展噪声敏感基础机理研究。同时, 城市环境中道路噪声分布复杂, 仅有鸟类敏感声压级阈值还不能直观评价城市内交通噪声对鸟类造成的生态风险。在宏观的声环境评价研究方面, 已有学者开展城市区域尺度的声环境评价研究。Wang等[25]提出了基于空间细分的波束跟踪法来模拟和评估城市建筑噪声;Gao等[26]通过现场实测和仿真模拟, 探究了快速检测城市噪声、评价降噪效果的方法;Aumond等[27]基于克里格法的空间插值方法, 利用城市区域声压级测量值绘制了声环境插值地图。上述研究主要通过实测、模拟和预测获得城市局部区域的声环境分布, 其中模拟是可便捷获取噪声分布的方法, 但是模拟结果缺乏空间信息及数值信息。此外, 与光污染类似, 研究仅定性地评价了声环境的分布或噪声改造情况, 但声环境分布情况无法体现城市噪声对鸟类形成的实际风险。
完善的生态风险评价需要构建完整的技术链条[28], 针对光污染和噪声污染生态风险评价领域存在的不足, 本文搭建了全链条研究方法: 探究微观层面光、声污染胁迫下鸟类生态风险阈值, 同时获取宏观光、声污染分布数据;将微观风险阈值和宏观污染分布数据联合, 开展生态风险评价;搭建生态风险评价平台并进行生态风险可视化展现。具体而言, 在微观层面, 本文探究了光污染对鸟类寤寐节律及声污染对鸟类退避行为的生态风险阈值;由于鸟类所处的真实环境中, 人工光多以混合光的形式长期稳定存在, 因此本文光污染风险研究围绕混合光对鸟类造成的生态风险进行评价。本文建立的全链条研究方法可为城市光、声污染生态风险的快速诊断、评价及防控提供方法支撑。
1 研究方法开展环境污染生态风险评价研究的首要条件是获取两类数据, 即典型受体的风险评价阈值和环境污染空间分布数据。前者主要来自污染源刺激实验获得的典型受体风险阈值, 后者来自于实测、建模、遥感等方式。
1.1 生态风险评价阈值量化 1.1.1 光污染生态风险评价阈值量化本文探究了混合光影响黄雀和栗鹀寤寐节律的光强风险阈值。团队前期基于规范化实验方法, 采用常规LED混合光及五种单色光(包括紫光、蓝光、绿光、黄光和橙红光)作为实验光源, 以辐照度作为实验光强的衡量指标, 选取入睡时刻、清醒时刻、睡眠时长及惊醒率作为鸟类寤寐节律观测指标, 开展人工光干扰下鸟类寤寐节律实验研究[29]。本文通过总结该实验结果, 发现以下规律。对黄雀而言, 5 mW/m2人工光推迟入睡时刻、提前清醒时刻及缩短睡眠时长均超过1 h, 并显著提升惊醒率;当人工光从较低辐照度升至5 mW/m2时, 四项寤寐节律观测指标变化显著, 而随着辐照度继续增大, 观测指标变化趋势减缓。对栗鹀而言, 10 mW/m2人工光推迟入睡时刻、缩短睡眠时长均超过1 h, 且推迟清醒时刻, 与较低辐照度人工光提前清醒时刻的现象形成鲜明对比。通过对比不同辐照度下六种光色对鸟类寤寐节律观测指标的综合影响程度, 本文初步确定黄雀和栗鹀的人工光辐照度风险阈值分别为5 mW/m2和10 mW/m2。
1.1.2 室外夜间光污染生态风险评价阈值转换实验场景难以还原复杂的室外夜间光环境光谱分布, 且实验研究常采用的光环境评价指标(照度、辐照度)与宏观夜光遥感数据的评价指标(辐亮度)不一致, 因此本文提出根据特定鸟种辐照度风险阈值估算符合研究区域混合光光谱能量分布的辐亮度风险阈值的方法, 具体如下:
(1) 开展实验室单色光辐亮度定标。因研究团队前期开展的光刺激实验获得的鸟类风险指标为辐照度(mW/m2), 而可用于大范围生态风险评价的夜光遥感影像单位是经辐射校正后的辐亮度(nW cm-2 sr-1)。若想利用夜光遥感影像进行大范围光污染生态风险评价, 则需要通过光强定标将实验获得的辐照度阈值转换为辐亮度阈值。本文采用峰值波段位置精准、于可见光范围内分布较为均匀的六种定制单色光, 包括蓝色光(448 nm)、青色光(501 nm)、黄绿色光(542—554 nm)、琥珀色光(594 nm)、橙红色光(618 nm)、红色光(660 nm)。实验人员将六种单色光置于标准实验箱装置内形成人工光环境, 使用拓普康SR-3AR分光辐射度计, 分别测量了六种单色光工况下实验鸟站立平面辐照度为5、10 mW/m2时对应的实验装置底部漫反射表面的辐亮度。以10 mW/m2辐照度实验工况为例, 各单光色光谱能量分布如图 1所示。

|
| 图 1 6种单色光光谱能量分布及峰值波长 Fig. 1 Spectral power distribution (SPD) and peak wavelength of six monochromatic lights |
(2) 确定研究区光环境光谱能量分布。通过调研发现, 道路照明、建筑照明、景观照明是对夜间光环境影响较大的几种照明类型。使用CL-500A分光辐射照度计可以采集夜间光环境光谱能量分布, 获得典型光谱能量分布特征。
(3) 划分单色光代表波段。根据6种单色光光谱能量分布(图 1), 以峰值波长相邻的两种单色光光谱能量分布曲线交点对应的波长为界, 将可见光范围(360—780 nm)分为6个波段, 各波段中光谱辐照度高的单色光作为主光色, 从而将单色光与代表波段一一对应。以相邻的蓝色光和青色光为例, 两者的光谱能量分布曲线在473 nm处相交, 且在波段360—473 nm内蓝色光的光谱辐照度高, 因此确定蓝色光所代表的蓝色光波段为360—473 nm。同理, 青色光代表波段为473 nm至青色光和黄绿色光光谱能量分布曲线交点对应的波长, 其他单色光代表波段范围以此类推。
(4) 确定真实光环境中各单色光波段具有的能量权重。权重系数是真实光环境光谱能量分布中, 不同单色光所在波段范围内对应的辐照度与真实光环境总辐照度的比值。
(5) 估算符合真实光环境的辐亮度阈值。单色光权重系数乘以该单色光在实验室环境下辐照度阈值定标对应的辐亮度值, 并求和, 即可估算得符合真实光环境可见光波段的辐亮度阈值。
公式1总结了(4)、(5)的计算方法。
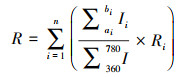
|
(1) |
式中, n为单色光的个数, 第i个单色光代表波段范围为ai-bi, I为测量的研究区域可见光波段真实光环境光谱辐照度, Ii为单色光i代表波段ai-bi在真实夜间光环境中的光谱辐照度, Ri为单色光i在实验室特定辐照度定标的辐亮度, R为估算获得的符合真实光环境的辐亮度阈值。
1.1.3 噪声污染生态风险评价阈值量化本文探究了道路噪声影响画眉鸟退避率的声压级风险阈值。画眉鸟具有生性胆小、对噪声敏感的特征, 因此适宜作为典型物种开展实验室噪声干扰实验。退避行为是鸟类受噪声影响的典型反应之一, 以退避率作为量化指标。研究定义某声压级下的退避率为该声压级下画眉鸟退避数量除以实验鸟总数量的百分比。研究团队在条形实验室两端播放野外采集的噪声源, 模拟野外道路噪声环境, 以画眉鸟作为典型风险受体, 观察各声压级下画眉鸟的分布情况, 并以退避率作为因变量y, 声压级作为自变量x, 建立了一元回归方程(公式2)[30]:
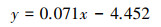
|
(2) |
夜间光环境地图的来源主要有三种, 实测光环境地图、无人机航拍图和夜光遥感影像。对于城市区域尺度光污染生态风险评价, 夜光遥感影像更具有优势。本文中, 光环境地图来自Luojia 1-01卫星(LJ1-01)和Jilin 1-7B卫星(JL1-07B)获取的夜光遥感影像, 两者空间分辨率分别为130 m[31]和0.92 m[23]。其中, LJ1-01图像来自湖北高分辨率地球观测系统数据和应用网络(http://59.175.109.173:8888/app/login_zh.html, 于2020年3月31日访问), 拍摄于2018年8月21日。JL1-07B夜光遥感影像拍摄于2020年4月9日。根据图 2所示流程, 基于ArcGIS 10.2对两种夜光遥感影像进行了数据几何纠正、辐射校正、中值滤波, 并将WGS84基准的通用横轴墨卡托投影应用于遥感图像。通过一系列处理, 获得研究区域的夜间光环境地图。

|
| 图 2 光环境地图及光污染风险地图处理流程 Fig. 2 The processing flow of light environment map and light pollution risk map |
基于官方提供的辐射校正方法, 使用公式(3)将LJ1-01图像的DN值转换为辐亮度。

|
(3) |
式中, R为绝对辐射校正后的辐亮度(W cm-2 sr-1), DN为像素的灰度值, w为带宽[32]。为便于可视化表现, 下文所提及的辐亮度值单位多为nW cm-2 sr-1。
对于JL1-07B夜光遥感影像, 根据元数据中提供的方法对其单波段影像进行辐射校正。灰度亮度计算方法见公式4:
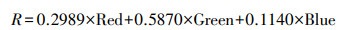
|
(4) |
式中, Red、Green、Blue为经过辐射校正后的JL1-07B单波段影像。
在光环境地图的基础上, 以特定鸟种风险阈值作为分类值, 利用ArcGIS重分类工具进行重分类, 将研究区域划分为安全区和风险区, 获得光污染风险地图。
1.2.2 声环境地图及噪声污染风险地图处理交通噪声现已成为噪声污染的最显著来源之一。因此, 本文以交通噪声作为主要噪声源, 通过噪声环境模拟、实测声压级校核、ArcGIS数据处理获得噪声地图, 方法如下。
(1) 实测校核的噪声环境模拟
① 噪声声压级实测: 在研究区域的主要交通干路及支路设置监测点, 使用声级计测量声压级, 同时通过拍摄视频获取车流量信息, 每测点测量10分钟, 取平均声压级, 并排除汽鸣声等极端噪声的干扰。每日分别在早高峰(7:00—8:00)、早平峰(9:00—10:00)、晚高峰(18:00—19:00)、晚平峰(21:00—22:00)四个时间段进行监测。通过上述操作, 实测获得不同时间段内显著噪声源的声压级及车流量。
② 声环境仿真模拟: 使用软件SoundPLAN进行研究区域建模和声环境模拟。建模考虑因素包括建筑形状、高度及分布, 道路表面材料、宽度及车流量, 并设定研究区域中的林地、草地及水域, 同时使用实测声压级对模拟数据进行校核, 最终输出符合真实环境的声环境模拟图。
(2) 声环境地图处理
由于模拟输出的数据为RGB图片, 图内可直接读取数据仅为R、G、B三个通道的灰度值, 缺少空间及数值属性。为解决这一问题, 本文提出基于ArcGIS的声环境模拟地图处理技术, 将位置属性及声压级数值属性赋予声环境仿真模拟图像, 生成声环境地图及噪声污染风险地图(图 3)。处理流程如下:

|
| 图 3 声环境地图及风险地图处理流程 Fig. 3 The processing flow of noise environment map and risk map |
① 图像预处理: 将声环境模拟地图导出为灰度图像, 通过颜色识别, 获取并记录与声压级分类一一对应的灰度值(0—255)。②赋予位置属性: 将声环境模拟图加载到ArcGIS, 并设置为墨卡托投影坐标系, 利用地理配准工具以谷歌地图作为底图对声环境模拟图进行地理配准, 使其获得空间属性。③赋予声压级数值属性: 将声环境模拟图R、G、B任意一波段加载到ArcGIS, 使用重分类工具, 以灰度值为旧值, 以对应声压级作为新值, 重新赋值该波段。至此, 新生成的栅格图像则为具有位置属性和声压级数值属性的声环境地图, 图中每像元灰度值均为声压级。④栅格重分类: 以特定鸟种噪声污染风险阈值作为分类值, 对声环境地图进行重分类, 获得噪声污染风险地图。
1.3 评价平台搭建评价平台主要包括客户端前端、管理端前端、后端及数据库。(1)客户端前端以网络地图为支撑, 可视化环境污染源分布地图及特定物种生态风险评价地图。(2)管理端前端服务于管理人员, 实现便捷化数据上传、管理、查询、调取、更新等功能。(3)后端通过接口与前端进行数据交互, 进行业务逻辑处理工作(数据的增、删、改、查等), 并保存于数据库中。(4)数据库主要用于存储污染源数据、物种生态风险评价数据等。
2 结果与分析 2.1 示范研究区概况福建省厦门市拥有丰富的鸟类资源, 鸟类种数占全国总种数的19.5%, 主要为鸣禽和涉禽。其中, 黄雀、栗鹀和画眉鸟, 是厦门市三种常见鸣禽[33]。位于厦门本岛中西部的筼筜湖区(图 4), 经过多年的环境修复和治理, 俨然成为厦门的“城市会客厅”、鸟类栖息的天堂, 是典型的城市生态斑块。经调研发现, 图 4中a—e区域均存在鸟类频繁活动的现象。但筼筜湖区及周边是厦门市的政治、文化、金融中心, 城市化水平高, 导致夜间光污染和噪声污染较为严重, 逐渐成为影响生态斑块内鸟类生存的潜在风险因素。因此, 本文以筼筜湖区域这一典型城市生态斑块作为示范研究区, 探讨如何将宏观光污染和噪声污染数据与微观鸟类阈值数据结合进行城市生态斑块生态风险评价。

|
| 图 4 筼筜湖地图 Fig. 4 Map of Yundang Lake |
调研发现, 筼筜湖区环湖建筑照明是主要光污染源, 建筑照明形式包括整体照明、局部照明、轮廓照明等。该区域建筑的平均高度约20 m, 高层建筑约占总建筑数量的17%, 其建筑立面亮化强度及垂直高度较高, 对整体光环境影响大。因此, 筼筜湖环湖建筑照明光谱分布可以一定程度代表该区域综合光谱分布。实验人员在湖心岛(图 4)南北侧码头采集环湖建筑立面光谱, 测点距离最近的建筑约220 m。通过CL-500A分光辐射照度计进行建筑立面光谱采集, 获得代表筼筜湖区的夜间光谱能量分布(图 5)。

|
| 图 5 筼筜湖夜间光环境光谱能量分布 Fig. 5 SPD of nighttime light environment in Yundang Lake |
根据阈值量化提出的估算混合光谱风险评价阈值的方法, 最终获得符合筼筜湖区典型光谱能量分布的辐亮度阈值: 黄雀人工光风险阈值5 mW/m2对应的辐亮度阈值为118.28 nW cm-2 sr-1, 栗鹀人工光风险阈值10 mW/m2对应的辐亮度阈值为218.88 nW cm-2 sr-1。
2.2.2 噪声污染生态风险评价阈值根据研究团队获得的画眉鸟退避率与声压级关系式可知, 当声压级为62.7 dB(A)时, y=0, 即此时画眉鸟出现退避行为的概率为0;当声压级大于62.7 dB(A)时, 画眉鸟即开始出现退避行为, 且概率随声压级增加而增大。因此, 为确保无任何画眉鸟出现退避行为, 本文将画眉鸟的噪声风险阈值定于62.7 dB(A)。
2.3 生态风险评价及分析 2.3.1 夜间光环境地图及光污染风险评价通过1.2.1操作, 获得筼筜湖区域的JL1-07B(图 6a)和LJ1-01(图 6b)夜间光环境地图。在此基础上, 以实验及计算获得的黄雀人工光风险阈值118.28 nW cm-2 sr-1、栗鹀人工光风险阈值218.88 nW cm-2 sr-1作为辐亮度阈值, 对光环境地图进行重分类, 继而生成光污染风险地图(图 6c、d、e、f)。图 7e中选取四个夜间光污染典型区域: g为市政建筑照明(厦门市政府和人民会堂), h为住宅建筑群照明, i为商业建筑群照明(万象城), j为道路及沿湖景观照明。

|
| 图 6 光环境地图及特定鸟种光污染风险地图 Fig. 6 Light environment maps and light pollution risk map of specific bird species a: JL1-07B夜光地图;b: LJ1-01夜光地图;c: 黄雀JL1-07B光污染风险地图;d: 黄雀LJ1-01光污染风险地图;e: 栗鹀JL1-07B光污染风险地图;f: 栗鹀LJ1-01光污染风险地图;g-j: 选取的典型光污染区域的JL1-07B图像 |

|
| 图 7 闽三角城市群夜间光环境地图及生态风险地图 Fig. 7 Nighttime light environment map and ecological risk map in Fujian Triangle a: 闽三角区域LJ1-01夜间光环境地图;b: 黄雀LJ1-01光污染风险地图;c: 栗鹀LJ1-01光污染风险地图;d: 厦门市黄雀LJ1-01光污染风险地图 |
由筼筜湖夜间光环境地图可见(图 6), 该区域两类夜光影像表征的夜间光环境分布特征相近, 基本以道路及建筑为中心形成集中照明区域, 并且在道路周边建筑区域形成局部光污染热点。由于分辨率的差异以及溢散光的影响, LJ1-01地图的整体辐亮度水平高于JL1-07B。以栗鹀风险地图为例, 研究区域中, LJ1-01地图显示光污染风险区域面积6.31 km2, 而JL1-07B地图显示风险区域面积仅为0.96 km2。对比两者风险区域, 发现风险重合区域与JL1-07B风险区域的面积比为84.0%, 可见两类夜光影像对于光污染核心风险区域的识别是相似的。
JL1-07B夜光影像分辨率高, 可以精细表现夜间照明分布, 是较好的城市区域尺度夜间光环境数据来源。因此, 针对厦门市筼筜湖区域, 本文以JL1-07B光污染风险地图对鸟类风险区进行描述: 超出黄雀风险阈值的光污染风险区域占研究区域的24.6%, 主要为湖区道路照明和市政、商业、住宅等建筑群装饰性照明;相比之下, 栗鹀超出风险阈值部分相对较小, 占比6.9%, 主要来源于建筑装饰性照明和景观照明。本文推测对黄雀和栗鹀造成风险的高辐亮度区域缘于照明设施缺乏截光措施。
LJ1-01夜光影像虽分辨率相对较低, 难以细致地对城市区域尺度光环境进行评价, 但其每景影像覆盖范围广, 因此更适用于大尺度夜间光环境生态风险评价。本文利用LJ1-01夜光数据绘制了厦门市、漳州市、泉州市在内的闽三角城市群夜间光环境地图, 并分别对该区域的黄雀和栗鹀的光污染生态风险进行了评价(图 7)。由图可见, 闽三角城市群绝大部分区域属于光污染生态安全区, 光污染生态风险区集中分布在城市中心和东南沿海区域。黄雀和栗鹀的光污染风险区分别占闽三角城市群面积的0.8%和0.4%, 其中, 漳州市和泉州市的光污染风险较小, 黄雀和栗鹀光污染风险区域在漳州市分别为0.3%和0.2%, 在泉州市分别为0.7%和0.2%。而厦门市的光污染风险最高, 黄雀和栗鹀的风险区域面积占比分别为6.0%和3.2%。
2.3.2 夜间光污染风险分布模式道路照明、建筑照明、景观照明是需重点关注的照明类型, 三者的夜间光照水平与土地利用类型密切相关[34]。探究城市土地类型与夜间光污染之间的关系, 有助于为防治生态光污染、降低生态风险提供支撑。
根据卫星地图及实地调研, 本文获得筼筜湖区域土地利用类型分布(图 8), 包括商业、住宅、行政、绿化及其他共五类用地。其中, 住宅所占比例最大, 商业次之。为评价不同类型用地对夜间光污染的贡献, 本文通过叠加土地利用类型图和JL1-07B夜光影像, 计算获得各分区斑块的总辐亮度, 即各斑块内所有像素辐亮度之和(图 9和图 11)。根据各斑块总辐亮度分布图及箱型图可以看出, 五类土地利用类型中, 住宅用地总辐射亮度较高, 商业其次, 这两类占地面积相对较大, 因此对整体光环境贡献的夜间光照也最多;而行政、绿化、其他三类面积占比相对较小, 对光污染的贡献也相对较小。为了区分不同土地利用的光照强度, 进一步计算了各土地利用斑块的平均辐亮度(图 10和图 12), 平均辐亮度为各斑块总辐亮度与总像元数之比。结果显示, 商业用地各斑块的平均辐亮度水平最高;行政用地次之, 存在局部过亮的照明;住宅的光照强度相对均衡。

|
| 图 8 研究区城市土地利用分布图 Fig. 8 Urban landuse distribution map in the study area |

|
| 图 9 各斑块总辐射亮度分布 Fig. 9 Total radiance distribution in each block |

|
| 图 10 各斑块平均辐射亮度分布 Fig. 10 Average radiance distribution in each block |

|
| 图 11 各土地利用类型总辐射亮度分布 Fig. 11 Total radiance distribution of each land use type |

|
| 图 12 各土地利用类型平均辐射亮度分布 Fig. 12 Average radiance distribution of each land use type |
本文统计了五类功能分区中黄雀和栗鹀的光污染风险区域占比(即各类分区中超出阈值的面积与该类区域的面积之比, 图 13)。五类分区中, 黄雀和栗鹀风险区域占比相差较大, 但总趋势一致。其中, 商业和行政区域超过阈值部分占比最高, 商业区域中, 黄雀和栗鹀风险区域占比分别为55.0%和24.9%;行政区域中, 两者占比分别为44.8%和19.4%。其次, 住宅区域的风险区域占比也相对较高, 黄雀和栗鹀风险区分别占34.5%和10.4%。本文推测, 这是由于商业和行政区域的建筑及景观室外照明设计不合理, 导致较多光线直接射向天空, 或照射到建筑表面和路面而反射到天空, 被卫星传感器捕捉。且调研发现, 该区域住宅多为高层, 顶部装饰照明普遍, 且缺乏截光措施。

|
| 图 13 黄雀和栗鹀光污染风险区域在各功能分区占比 Fig. 13 The proportions of light pollution risk areas of Eurasian Siskin and Chestnut Bunting in each functional zone |
本文根据前文1.2.2描述的声环境地图处理方法, 获得筼筜湖区声环境地图, 以晚高峰阶段为例进行表现(图 14)。并根据典型风险受体画眉鸟的退避风险阈值62.7 dB(A)对噪声污染进行评价, 生成噪声污染风险地图(图 14)。

|
| 图 14 筼筜湖区晚高峰声环境地图及噪声污染风险地图 Fig. 14 Nighttime peak noise map and noise pollution risk map |
多项鸟类研究表明, 噪声水平高于45 dB(A), 鸟类鸣声特征、繁殖、应激激素水平和物种丰富度会发生变化[14]。根据筼筜湖区域噪声模拟结果显示, 距单向道路噪声源近1300 m处, 环境声压级降至约45 dB(A)。道路围合的筼筜湖湖心声压级大约为55 dB(A)。画眉鸟风险区占研究区面积的34.7%。
2.3.4 噪声污染风险分布模式为深入了解该区域噪声沿道路分布模式, 对声环境地图进行声压级提取及分析。沿垂直于道路的方向设置声压级提取路径(图 15中红线), 设置提取路径与道路中心交叉点为中心点, 沿远离道路中心点方向每隔10m设置一个提取点。筼筜湖区湖滨东路、湖滨西路为一级道路, 白鹭洲路及湖滨中路为二级道路。根据噪声声压级随距道路中心线距离的变化曲线(图 16), 声压级在道路中心处最高, 甚至超过80 dB(A)。随着距离的增加, 噪声受到障碍物遮挡而衰减, 声压级逐渐降低。对于画眉鸟而言, 距离一级道路约150 m、距离二级道路约50 m以内为画眉鸟噪声污染风险区。针对风险受体主要活动区域与噪声污染风险预警区发生重叠的问题, 相关保护组织可以考虑通过在道路附近设置声屏障或增加绿化林带的方式缓解噪声对风险受体的影响, 运用SoundPLAN模拟调整后的噪声分布情况, 验证规划建议的有效性, 最终提出改进方案。

|
| 图 15 研究区域声压级数据提取 Fig. 15 Data extraction of the sound pressure level (SPL) in the study area |

|
| 图 16 声压级随距道路中心线距离变化图 Fig. 16 The SPL changes with the distance from the center line of the road |
在夜间照明和交通晚高峰的重叠时间段, 光污染和噪声污染同时作用于鸟类, 并可能产生单日最大叠加风险。经调研发现, 研究区道路照明开启时间与日落时间基本一致, 建筑及景观照明随后开启, 晚高峰时段会随日落时间变化、作息时间调整而相应偏移。研究区全年夜间照明时段会与交通晚高峰时段完全或部分重叠。以团队11月调研结果为例, 道路照明随日落大约在17:20开启, 交通晚高峰时段约为18:00—19:00, 光污染和噪声污染时段完全重合。因此, 本文对夜间黄雀、栗鹀光污染风险和画眉鸟噪声污染风险开展了综合评价(图 17)并获得以下结果。风险区集合的总面积为4.99 km2, 约占研究区域的44.32%。将生态风险区集合划分为三个等级: 仅一种物种遭受风险的区域定义为轻度污染区域(占地2.55 km2, 约占总生态风险区域的51.10%), 两种物种生态风险重合的区域定义为中度污染区域(1.91 km2, 38.28%), 三种物种风险区域重合的区域定义为重度污染区域(0.53 km2, 10.62%)。结果表明, 重度污染区域面积有限, 主要分布于靠近道路且照明水平高的区域;中度污染及轻度污染区域, 覆盖范围相对较广。风险水平高的区域亟需政府相关部门给予重点关注, 降低物种的光污染和噪声污染生态风险。

|
| 图 17 光、声污染多重风险地图 Fig. 17 Multiple risk map of light and noise pollution |
网页客户端界面如图 18所示, 关键功能是由用户选择不同物理环境污染源及特定物种生态风险评价地图进行可视化展示, 并叠加于开源地图Open Street Map (OSM)上方。当前物理环境污染源包括光污染和噪声污染。在网页客户端平台中, 可调节光环境、声环境地图及污染风险地图的不透明度, 用于查看光污染和噪声污染生态风险的叠加效果, 据此综合评价生态风险, 为政府相关部门决策及环境治理提供依据。

|
| 图 18 平台网页客户端界面及功能展示 Fig. 18 The web-based client interface and function |
本文在GIS技术的支持下, 将微观的风险阈值与宏观的污染地图结合, 以典型城市生态斑块筼筜湖区为例, 进行了光污染和噪声污染胁迫下的鸟类生态风险评价, 得出以下结论并展开讨论:
(1) 微观风险阈值
在光污染研究中, 本文根据团队前期研究, 初步总结了黄雀和栗鹀的人工光辐照度风险阈值分别为5 mW/m2和10 mW/m2, 对应的实验LED混合光照度风险阈值分别为1.78 lx和3.38 lx。该风险阈值与已有研究中对鸟类产生影响的混合光光强相近。例如, 1.5 lx人工光使城市大山雀(Parus major)日间活动开始时刻提前55分钟[35];1.6 lx人工光可能扰乱大山雀睡眠[7, 36];3.2 lx人工光可能影响灌丛鸦(Aphelocoma californica)褪黑素分泌[37], 或进一步影响寤寐节律;10 mW/m2人工光会推迟绣眼鸟(Silvereyes)入睡行为[38];在平均照度为3.91 lx的重度光污染区, 美国知更鸟的晨鸣时间相比于轻度光污染区大幅度提前[39]。此外, 本文提出了根据特定鸟种实验室辐照度阈值估算符合真实光环境混合光谱能量分布下辐亮度阈值的方法, 并获得符合筼筜湖区典型光谱能量分布的黄雀和栗鹀辐亮度风险阈值, 分别为118.28 nW cm-2 sr-1、218.88 nW cm-2 sr-1。不同鸟种对于人工光强度的敏感度和耐受力存在差异, 在未来研究中有待开展更多鸟种风险机理研究。
在噪声污染研究中, 画眉鸟呈现0%退避率时, 风险阈值为62.70 dB(A)。相关研究表明, 70 dB(A)或更高水平的噪声会迫使鸟类产生退避行为[40]。该噪声水平微高于本文中的风险阈值62.70 dB(A), 推测原因有二: 其一是鸟种间差异[24], 其二是实验设置方法不同。
(2) 宏观污染地图及风险评价
在光污染研究中, 本文利用鸟类风险阈值结合JL1-07B夜光遥感影像进行了筼筜湖区光污染风险评价, 发现黄雀和栗鹀在该区域的光污染风险区占比分别为24.6%和6.9%。在筼筜湖区, LJ1-01与JL1-07B表征的栗鹀风险区域重合率达84.0%, 说明LJ1-01与JL1-07B表征的光污染风险分布相近。基于两类夜光数据各自特征, 本文认为JL1-07B夜光地图适用于城市区域尺度光污染风险评价, LJ1-01更适用于城市群大尺度评价。以闽三角城市群为例, LJ1-01风险评价地图显示, 黄雀和栗鹀的光污染风险区域占比为0.8%和0.4%, 其中厦门市的光污染风险最高, 分别占全部风险区域的6.0%和3.2%。对比Xue等[23]研究中关于鸟类栖息地光污染等级划分, 本文获得的黄雀光污染风险阈值(118.28 nW cm-2 sr-1)对应该研究中的初级光污染(89.40—151.84 nW cm-2 sr-1), 栗鹀光污染风险阈值(218.88 nW cm-2 sr-1)对应中级光污染(151.85—334.60 nW cm-2 sr-1)。该研究虽然没有参考鸟类风险阈值, 但是光污染分级可与本文实验鸟种风险阈值对应, 说明采用自然断点法获得的光污染等级一定程度上能反映典型人工光对鸟类的影响情况。但是, 由于不同鸟种对人工光的敏感程度存在差异, 自然断点法获得的光污染等级能否代表各典型鸟种面临的光污染风险尚不明确, 因此仍需开展鸟类对人工光响应机理研究, 从而为光污染风险等级的合理划分提供支撑。
在噪声污染研究中, 本文通过SoundPLAN模拟、实测校核以及ArcGIS赋予空间及数值属性, 获取声环境地图, 结合画眉鸟噪声风险阈值, 发现画眉鸟噪声风险区域占筼筜湖区的34.7%。噪声污染可能会导致以画眉鸟为代表的城市鸟类出现退避行为而放弃栖息地, 进而可能造成城市生态斑块栖息地功能退化。
在光、声污染综合评价中, 筼筜湖区光、声污染风险区域总面积4.99 km2, 占筼筜湖区域面积的44.32%。轻度污染区域2.55 km2(占污染风险区域的51.10%);中度污染区域1.91 km2(38.28%), 重度污染区域0.53 km2(10.62%)。在评价平台客户端中, 可通过调节地图的不透明度查看光、声污染生态风险的叠加效果, 据此进行综合生态风险评价。相关研究表明, 在光污染和噪声同时存在的夜晚, 人工光对城市鸟类造成的不良影响将被进一步放大;而对于郊区鸟类, 人工光和噪声的影响也将叠加[35]。因此, 夜间光污染和噪声污染的风险区域如出现重叠, 则表明该重叠区域的城市生态环境亟待改善。
(3) 风险分布模式
在光污染研究中, 五类土地利用类型中的商业区域光污染风险最多, 黄雀和栗鹀风险区域占比分别为55.0%和24.9%;行政和住宅区域其次。调研发现, 在非节假日夜间, 筼筜湖区的大部分建筑外立面装饰照明在22:30关闭, 但商业区低层沿街商铺照明以及基础设施照明基本通宵运行。因此, 光污染的防治措施需从功能和时间两个角度考虑。在噪声污染研究中, 距离一级道路约150m以及距离二级道路约50m以内的区域为画眉鸟噪声污染风险区。道路中心处声压级最高, 达到80 dB(A)以上。筼筜湖区大部分区域噪声水平超过55 dB(A), 高于世界卫生组织(WHO)认定的人类长期接触的噪声安全水平。
(4) 光、声污染控制建议
从控制光污染的角度, 本文建议城市管理部门根据鸟类活动频率、绿化率等因素重点关注商业区域和行政区域的光照水平, 对于鸟类光污染风险区采取降低光照强度、减少光源数量、缩短照明时间等措施以减少光污染生态风险。从控制声污染的角度, 建议在鸟类活动、栖息及繁殖的区域, 相关机构需采取必要措施适当减少道路噪声的影响, 例如限制车速、增加绿化、增建隔音设施等。
| [1] |
Lyytimäki J. Avoiding overly bright future: the systems intelligence perspective on the management of light pollution. Environmental Development, 2015, 16: 4-14. DOI:10.1016/j.envdev.2015.06.009 |
| [2] |
施慧雄, 焦海峰, 尤仲杰, 王亚军, 李松海, 徐继林, 杨家锋. 船舶噪声对鲈鱼和大黄鱼血浆皮质醇水平的影响. 生态学报, 2010, 30(14): 3760-3765. |
| [3] |
Senzaki M, Barber J R, Phillips J N, Carter N H, Cooper C B, Ditmer M A, Fristrup K M, Mcclure C J W, Mennitt D J, Tyrrell L P, Vukomanovic J, Wilson A A, Francis C D. Sensory pollutants alter bird phenology and fitness across a continent. Nature, 2020, 587(7835): 605-609. DOI:10.1038/s41586-020-2903-7 |
| [4] |
Sheppard C, Phillips G. Bird-Friendly Building Design. 2nd ed. The Plains, VA: American Bird Conservancy, 2015: 60-60.
|
| [5] |
Dominoni D M. The effects of light pollution on biological rhythms of birds: an integrated, mechanistic perspective. Journal of Ornithology, 2015, 156(1): 409-418. |
| [6] |
Raap T, Sun J C, Pinxten R, Eens M. Disruptive effects of light pollution on sleep in free-living birds: season and/or light intensity-dependent?. Behavioural Processes, 2017, 144: 13-19. DOI:10.1016/j.beproc.2017.08.011 |
| [7] |
Raap T, Pinxten R, Eens M. Artificial light at night disrupts sleep in female great tits (Parus major) during the nestling period, and is followed by a sleep rebound. Environmental Pollution, 2016, 215: 125-134. DOI:10.1016/j.envpol.2016.04.100 |
| [8] |
Mishra I, Knerr R M, Stewart A A, Payette W I, Richter M M, Ashley N T. Light at night disrupts diel patterns of cytokine gene expression and endocrine profiles in zebra finch (Taeniopygia guttata). Scientific Reports, 2019, 9(1): 15833. DOI:10.1038/s41598-019-51791-9 |
| [9] |
Nagashima S, Osawa M, Matsuyama H, Ohoka W, Ahn A, Wakamura T. Bright-light exposure during daytime sleeping affects nocturnal melatonin secretion after simulated night work. Chronobiology International, 2018, 35(2): 229-239. DOI:10.1080/07420528.2017.1394321 |
| [10] |
Welbers A A M H, van Dis N E, Kolvoort A M, Ouyang J, Visser M E, Spoelstra K, Dominoni D M. Artificial light at night reduces daily energy expenditure in breeding great tits (Parus major). Frontiers in Ecology and Evolution, 2017, 5: 55. DOI:10.3389/fevo.2017.00055 |
| [11] |
蔡超. 道路噪声对鸟类习鸣质量的影响因子研究——以金丝雀为例[D]. 天津: 天津大学, 2012.
|
| [12] |
Habib L, Bayne E M, Boutin S. Chronic industrial noise affects pairing success and age structure of ovenbirds Seiurus aurocapilla. Journal of Applied Ecology, 2007, 44(1): 176-184. |
| [13] |
Slabbekoorn H, Ripmeester E A P. Birdsong and anthropogenic noise: implications and applications for conservation. Molecular Ecology, 2008, 17(1): 72-83. DOI:10.1111/j.1365-294X.2007.03487.x |
| [14] |
Shannon G, McKenna M F, Angeloni L M, Crooks K R, Fristrup K M, Brown E, Warner K A, Nelson M D, White C, Briggs J, McFarland S, Wittemyer G. A synthesis of two decades of research documenting the effects of noise on wildlife. Biological Reviews, 2016, 91(4): 982-1005. DOI:10.1111/brv.12207 |
| [15] |
Ware H E, Mcclure C J W, Carlisle J D, Barber J R. A phantom road experiment reveals traffic noise is an invisible source of habitat degradation. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(39): 12105-12109. DOI:10.1073/pnas.1504710112 |
| [16] |
Kociolek A V, Clevenger A P, Clair C C S, Proppe D S. Effects of road networks on bird populations. Conservation Biology, 2011, 25(2): 241-249. |
| [17] |
Muheim R, Bäckman J, Åkesson S. Magnetic compass orientation in European robins is dependent on both wavelength and intensity of light. Journal of Experimental Biology, 2002, 205(24): 3845-3856. DOI:10.1242/jeb.205.24.3845 |
| [18] |
Poot H, Ens B J, de Vries H, Donners M A H, Wernand M R, Marquenie J M. Green light for nocturnally migrating birds. Ecology and Society, 2008, 13(2): 47. DOI:10.5751/ES-02720-130247 |
| [19] |
Evans W R, Akashi Y, Altman N S, Manville II A M. Response of night-migrating songbirds in cloud to colored and flashing light. North American Birds, 2007, 60(4): 476-488. |
| [20] |
Zhao X B, Zhang M, Che X L, Zou F S. Blue light attracts nocturnally migrating birds. The Condor, 2020, 122(2): duaa0022. |
| [21] |
Wiltschko W, Wiltschko R, Munro U. Light-dependent magnetoreception in birds: the effect of intensity of 565-nm green light. Naturwissenschaften, 2000, 87(8): 366-369. DOI:10.1007/s001140050742 |
| [22] |
Horton K G, Nilsson C, van Doren B M, La Sorte F A, Dokter A M, Farnsworth A. Bright lights in the big cities: migratory birds' exposure to artificial light. Frontiers in Ecology and the Environment, 2019, 17(4): 209-214. DOI:10.1002/fee.2029 |
| [23] |
Xue X Y, Lin Y, Zheng Q M, Wang K, Zhang J, Deng J S, Abubakar G A, Gan M Y. Mapping the fine-scale spatial pattern of artificial light pollution at night in urban environments from the perspective of bird habitats. Science of the Total Environment, 2020, 702: 134725. DOI:10.1016/j.scitotenv.2019.134725 |
| [24] |
Patón D, Romero F, Cuenca J, Escudero J C. Tolerance to noise in 91 bird species from 27 urban gardens of Iberian Peninsula. Landscape and Urban Planning, 2012, 104(1): 1-8. DOI:10.1016/j.landurbplan.2011.09.002 |
| [25] |
Wang H B, Cai M, Cui H J. Simulation and analysis of road traffic noise among urban buildings using spatial subdivision-based beam tracing method. International Journal of Environmental Research and Public Health, 2019, 16(14): 2491. DOI:10.3390/ijerph16142491 |
| [26] |
Gao C, Gao S, Zhao G F. Research on rapid detection and control mechanism of urban traffic noise impact based on Soundplan software simulation. IOP Conference Series: Earth and Environmental Science, 2019, 233(5): 052050. |
| [27] |
Aumond P, Can A, Mallet V, de Coensel B, Ribeiro C, Botteldooren D, Lavandier C. Kriging-based spatial interpolation from measurements for sound level mapping in urban areas. The Journal of the Acoustical Society of America, 2018, 143(5): 2847-2857. DOI:10.1121/1.5034799 |
| [28] |
陈春丽, 吕永龙, 王铁宇, 史雅娟, 胡文友, 李静, 张翔, 耿静. 区域生态风险评价的关键问题与展望. 生态学报, 2010, 30(3): 808-816. |
| [29] |
刘芳博. 人工光干扰下黄雀和黄眉鹀的寤寐节律研究[D]. 天津: 天津大学, 2018.
|
| [30] |
刘刚, 刘芳博, 鲁世伟. 道路噪声与画眉鸟退避率的定量关系. 生态学杂志, 2018, 37(12): 3685-3690. |
| [31] |
Jiang W, He G J, Long T F, Guo H X, Yin R Y, Leng W C, Liu H C, Wang G Z. Potentiality of using Luojia 1-01 nighttime light imagery to investigate artificial light pollution. Sensors, 2018, 18(9): 2900. DOI:10.3390/s18092900 |
| [32] |
Li X, Li X Y, Li D R, He X J, Jendryke M. A preliminary investigation of Luojia-1 night-time light imagery. Remote Sensing Letters, 2019, 10(6): 526-535. DOI:10.1080/2150704X.2019.1577573 |
| [33] |
厦门观鸟协会. 厦门市鸟类名录2019V1.0. (2019-03-12)[2021-01-23]. https://xmbirds.org/bbs/index.php?showtopic=47761.
|
| [34] |
Guk E, Levin N. Analyzing spatial variability in night-time lights using a high spatial resolution color Jilin-1 image-Jerusalem as a case study. ISPRS Journal of Photogrammetry and Remote Sensing, 2020, 163: 121-136. DOI:10.1016/j.isprsjprs.2020.02.016 |
| [35] |
Dominoni D, Smit J A H, Visser M E, Halfwerk W. Multisensory Pollution: artificial Light at night and anthropogenic noise have interactive effects on activity patterns of great tits (Parus major). Environmental Pollution, 2020, 256: 113314. DOI:10.1016/j.envpol.2019.113314 |
| [36] |
Raap T, Pinxten R, Eens M. Light pollution disrupts sleep in free-living animals. Scientific Reports, 2015, 5: 13557. DOI:10.1038/srep13557 |
| [37] |
Schoech S J, Bowman R, Hahn T P, Goymann W, Schwabl I, Bridge E S. The effects of low levels of light at night upon the endocrine physiology of western scrub-jays (Aphelocoma californica). Journal of Experimental Zoology Part A: Ecological Genetics and Physiology, 2013, 319(9): 527-538. |
| [38] |
Liu G, Peng X T, Ren Z F, Liu M, Dang R, Chen Y Q, Liu F B. The effect of artificial light with different SPDs and intensities on the sleep onset of silvereyes. Biological Rhythm Research, 2019, 50(5): 787-804. DOI:10.1080/09291016.2018.1498201 |
| [39] |
Miller M W. Apparent effects of light pollution on singing behavior of American robins. The Condor, 2006, 108(1): 130-139. DOI:10.1093/condor/108.1.130 |
| [40] |
Liu Q X, Slabbekoorn H, Riebel K. Zebra finches show spatial avoidance of near but not far distance traffic noise. Behaviour, 2020, 157(3-4): 333-362. DOI:10.1163/1568539X-bja10004 |