 2022, Vol. 42
2022, Vol. 42文章信息
- 何雅琴, 史晓洁, 陈国杰, 赖敏英, 曾纪毅, 魏凯, 邓传远
- HE Yaqin, SHI Xiaojie, CHEN Guojie, LAI Minying, ZENG Jiyi, WEI Kai, DENG Chuanyuan
- 滨柃叶功能性状对环境因子的响应
- Response and adaptation of leaf functional traits of Eurya emarginata to environmental factors
- 生态学报. 2022, 42(6): 2418-2429
- Acta Ecologica Sinica. 2022, 42(6): 2418-2429
- http://dx.doi.org/10.5846/stxb202101180189
-
文章历史
- 收稿日期: 2021-01-18
- 网络出版日期: 2021-11-30

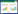




植物功能性状是指植物适应环境变化所形成的形态、生理和物候特征, 能反映植物对不同环境的响应和适应机制[1-2]。植物功能性状不仅影响生态系统功能, 还能将环境、植物和生态系统过程与功能密切联系起来[3]。叶片作为与外界环境进行物质交换的重要器官, 其功能性状易测量、数量多、可塑性较大, 特别是结构型性状(比叶面积、叶厚度、叶干物质含量等), 是植物生长需求对空间异质性或环境变化适应进化的结果[4-6]。近年来, 国内外学者开展了典型植物群落叶结构型功能性状与光照、气候、温度、降水、地形地貌、土壤养分等差异化环境因子间的相关性研究[7-9]。如, 前人研究表明, 植物在林下充足光照环境下倾向于资源获取策略, 在弱光环境中生存策略由资源获取策略转变为资源保守策略[7]。坡向、坡位、海拔等地形因子的变化会间接影响温度、降水、光照等因子产生再分配, 导致气候环境的空间异质性, 影响植物的生长发育并对植物功能性状的变化产生间接作用[9]。因此, 致力于揭示植物叶功能性状与环境因子的关系, 有助于更好理解植物应对环境变化的适应策略及分配利用资源能力。
海岛由于地理位置隔离, 成为独立的生态系统, 相比内陆生态系统, 其更具敏感性和脆弱性。地理隔离显著影响植物的表型性状[10], 如, 黄柳菁等调查了大陆性海岛平潭岛的种子植物, 并分析了其植物功能性状特征, 总结出海岛植物结构功能性状与内陆的区别, 强调海岛植物为了适应强风、干旱、贫瘠的环境, 呈现出以草本植物为主、小型叶植物占比大、叶被毛植物居多等结构功能性状特征[11]。国内一些典型海岛(海南岛和平潭岛)不同演替阶段植物功能性状与环境因子的变化规律已有研究[12-13]。如, 张增可等研究了平潭岛4个不同演替阶段群落植物功能性状对环境因子的响应, 结果表明, 演替早期植物主要分布在土壤pH值、容重高的贫瘠环境, 拥有较高比叶面积(SLA)、叶磷含量(LPC)、茎氮含量、茎磷含量的性状组合;演替后期植物主要分布在土壤养分和水分高的肥沃环境, 拥有较高的茎组织密度、叶干物质含量(LDMC)、叶碳含量(LCC)、叶氮含量(LNC)的性状组合。土壤有机质和全氮是影响海岛植物功能性状的主要土壤因子, 坡位和坡度是影响海岛植物功能性状的主要地形因子[14]。海岛典型植物群落优势种叶功能性状与环境因子的相关性也有研究, 如, 琅岐岛4种优势植物叶功能性状主要受坡度、海拔、土壤电导率、土壤含水量等环境因子的影响[15]。综上所述, 关于海岛不同典型植物群落优势种叶功能性状与环境因子的相关已有研究, 但典型植物群落同一优势种适应不同异化生境叶功能性状的变动还没有研究。
滨柃(Eurya emarginata), 山茶科(Theaceae)柃属(Eurya)常绿灌木。在中国呈地带性间断分布于浙江、福建和台湾等东南沿海及海岛, 且能形成以滨柃为优势种的滨海群落。由于滨柃群落分布特性, 关于滨柃群落结构特征已有研究。聂森研究发现, 滨柃群落物种丰富度和多样性与海拔呈负相关[16]。李加庆对群落优势种滨柃的平均株高、平均地径与环境因子的相关性研究表明, 滨柃的平均株高与pH值、速效钾、全盐量、海拔呈负相关, 滨柃的平均地径与全钾呈负相关[17]。滨柃多生于滨海山坡灌丛中及基岩海岛、海岸边岩石缝中, 在海岛恶劣生境中表现出较强抗逆性和适应性, 因此, 关于滨柃的抗逆性等方面国内外展开了较多研究[18-19], 如, 滨柃在150 mmol/L的盐胁迫环境下能正常生长, 可作为海岛环境修复的耐盐优势树种[18]。滨柃的存活率和生物量与干旱胁迫程度呈负相关[19]。滨柃生境特殊, 在构建海岛特色景观, 改善海岛生态环境上具重要作用。因此, 关于滨柃的组培应用也有研究[20], 但有关滨柃叶功能性状对环境因子的响应、权衡关系及适应机制的研究鲜见报道。因此, 本研究以福建省福州市平潭大练岛、海坛岛和大怀屿三地分布的滨柃为研究对象, 对滨柃叶功能性状指标与环境因子的相关性进行分析, 探究滨柃对资源的分配利用能力、揭示其对异质环境的适应机制, 为滨柃资源的保护及开发利用提供理论参考。
1 研究区概况平潭岛地处福建省东部, 隶属于平潭综合试验区, 地理位置位于25°15′-25°45′N, 119°32′-120°10′E, 由以海坛岛为主的126个岛屿和近千个岩礁组成, 海岸线长408 km, 陆地面积为324.12 km2, 为我国第五大海岛。研究区属于典型的南亚热带海洋性季风气候。年均气温为19.6 ℃, 年均降水量900-2100 mm, 年均日照1700-1980 h, 年均风速9.0 m/s, 一年中125 d以上是大风天气(7级以上), 夏秋季节常受台风影响。土壤类型多为风沙土、盐土、砖红壤, 土层瘠薄养分含量较低[21]。植被调查中发现, 滨柃灌丛在平潭大练岛、海坛岛、大怀屿等海岛相对恶劣环境中均有一定面积和数量的分布, 因此, 本文以大练岛、海坛岛、大怀屿3个海岛的滨柃进行试验, 其中大练岛和海坛岛为有居民海岛, 大怀屿为季节性有居民海岛, 各海岛概况见表 1。
| 海岛名称 Island name |
面积 Area/km2 |
岸线长 Perimeter/km |
海拔 Elevation/m |
地理位置 Geographic location |
年均温 Mean annual temperature/℃ |
年均降雨量 Mean annual precipitation/mm |
年均光照 Mean annual illumination/h |
相对湿度 Relative humidity/% |
| 大练岛 | 9.96 | 20.35 | 238.5 | 119°39′-119°44′E、25°38′-25°40′N | 19.1 | 1100 | 1840.3 | 75 |
| 海坛岛 | 267.13 | 204.5 | 434.6 | 119°40′-119°52′E、25°23′-25°40′N | 19.6 | 1180 | 1919.7 | 83 |
| 大怀屿 | 0.037 | 0.97 | 43.3 | 119°70′E-119°71′E、25°54.13′-25°54.36′N | 19.8 | 1196.2 | 1869.5 | 81 |
2020年7月-9月采用标准样方法对大练岛(围营山)、海坛岛(君山)、大怀屿植物群落中的滨柃叶片进行采集, 共设置30个样方。记录每个样方的经纬度、海拔(Elevation, E)、坡度、坡向及坡位。采用便携式测光仪测定郁闭度(Crown density, CD)。各岛的环境因子概况见表 2。
| 样方编号 Species number |
pH | 水溶性盐总量 S/(g/kg) |
有机质 SOM/ (g/kg) |
全氮 T-N/% |
全磷 T-P/% |
全钾 T-K/% |
碱解氮 A-N/ (mg/kg) |
有效磷 A-P/ (mg/kg) |
速效钾 A-K/ (mg/kg) |
海拔 E/m |
坡度 G/(°) |
坡向 SA |
坡位 SP |
郁闭度 CD |
| W1 | 5.0 | 0.8 | 47.8 | 0.238 | 0.048 | 1.47 | 194.4 | 9.3 | 146 | 123.8 | 29.4 | 3 | 中坡 | >0.7 |
| W2 | 4.8 | 0.6 | 43.9 | 0.219 | 0.032 | 1.54 | 183.2 | 8.7 | 120 | 130.3 | 28.7 | 1 | 中坡 | < 0.1 |
| W3 | 4.9 | 1.0 | 46.8 | 0.230 | 0.031 | 1.31 | 186.9 | 5.9 | 140 | 122.7 | 29.0 | 3 | 中坡 | < 0.1 |
| W4 | 4.3 | 1.0 | 56.9 | 0.329 | 0.046 | 1.72 | 228.1 | 9.5 | 150 | 172.9 | 41.2 | 1 | 上坡 | >0.7 |
| W5 | 4.5 | 0.8 | 64.8 | 0.359 | 0.050 | 1.69 | 280.4 | 10.6 | 176 | 190.2 | 42.0 | 1 | 上坡 | >0.7 |
| W6 | 5.1 | 0.5 | 48.0 | 0.215 | 0.033 | 1.29 | 179.5 | 4.6 | 136 | 131.8 | 29.1 | 1 | 中坡 | < 0.1 |
| W7 | 4.8 | 0.4 | 41.1 | 0.209 | 0.039 | 1.60 | 186.9 | 5.1 | 153 | 137.5 | 28.8 | 1 | 中坡 | < 0.1 |
| W8 | 5.0 | 0.4 | 33.4 | 0.170 | 0.039 | 1.77 | 153.3 | 3.7 | 136 | 160.1 | 31.3 | 8 | 上坡 | 0.3-0.5 |
| W9 | 4.5 | 1.0 | 57.0 | 0.324 | 0.045 | 1.93 | 265.5 | 8.8 | 176 | 156.9 | 26.6 | 1 | 上坡 | >0.7 |
| W10 | 5.5 | 1.3 | 49.1 | 0.229 | 0.040 | 2.24 | 220.6 | 4.4 | 226 | 139.2 | 31.2 | 8 | 中坡 | >0.7 |
| W11 | 4.5 | 1.0 | 56.0 | 0.285 | 0.038 | 1.95 | 246.8 | 5.6 | 190 | 129.0 | 27.3 | 1 | 中坡 | 0.3-0.5 |
| W12 | 4.9 | 0.8 | 47.6 | 0.236 | 0.047 | 1.45 | 195.1 | 9.2 | 145 | 123.6 | 29.3 | 3 | 中坡 | >0.7 |
| CV/% | 6.85 | 34.95 | 16.89 | 22.28 | 20.02 | 16.88 | 18.28 | 34.29 | 18.55 | - | - | - | - | - |
| J1 | 4.6 | 0.6 | 36.0 | 0.172 | 0.022 | 1.18 | 168.3 | 3.4 | 76 | 267.9 | 12.4 | 3 | 上坡 | < 0.1 |
| J2 | 4.6 | 0.2 | 20.3 | 0.105 | 0.018 | 1.22 | 112.2 | 2.5 | 50 | 265.7 | 10.5 | 3 | 上坡 | < 0.1 |
| J3 | 4.8 | 0.7 | 23.2 | 0.119 | 0.016 | 1.03 | 127.1 | 2.4 | 63 | 263.0 | 10.5 | 3 | 上坡 | < 0.1 |
| J4 | 5.3 | 0.05 | 14.8 | 0.080 | 0.016 | 1.13 | 74.8 | 2.3 | 76 | 260.8 | 11.4 | 4 | 上坡 | < 0.1 |
| J5 | 5.1 | 0.5 | 29.9 | 0.146 | 0.025 | 1.23 | 142.1 | 2.5 | 190 | 270.9 | 12.4 | 4 | 上坡 | < 0.1 |
| J6 | 5.0 | 0.5 | 41.7 | 0.208 | 0.030 | 1.35 | 190.7 | 3.1 | 150 | 276.7 | 26.4 | 6 | 上坡 | < 0.1 |
| J7 | 4.8 | 0.4 | 24.5 | 0.118 | 0.019 | 0.92 | 119.6 | 3.8 | 103 | 264.4 | 5.9 | 6 | 上坡 | < 0.1 |
| J8 | 4.8 | 0.4 | 26.0 | 0.118 | 0.023 | 0.94 | 119.6 | 3.4 | 73 | 251.6 | 6.2 | 6 | 上坡 | < 0.1 |
| J9 | 4.9 | 0.2 | 22.6 | 0.111 | 0.020 | 0.91 | 108.4 | 2.4 | 96 | 255.4 | 8.3 | 7 | 上坡 | < 0.1 |
| CV/% | 4.67 | 53.20 | 30.90 | 29.24 | 19.86 | 14.36 | 26.46 | 19.58 | 46.35 | - | - | - | - | - |
| D1 | 7.4 | 1.0 | 38.0 | 0.256 | 0.037 | 2.81 | 179.5 | 9.4 | 116 | 35.1 | 32.0 | 1 | 下坡 | < 0.1 |
| D2 | 8.7 | 0.8 | 32.0 | 0.211 | 0.038 | 2.97 | 145.8 | 9.3 | 246 | 41.3 | 31.4 | 1 | 下坡 | < 0.1 |
| D3 | 4.7 | 1.4 | 54.7 | 0.336 | 0.036 | 2.84 | 224.3 | 11.5 | 140 | 35.8 | 32.8 | 1 | 下坡 | < 0.1 |
| D4 | 7.3 | 0.9 | 37.8 | 0.233 | 0.036 | 2.76 | 178.9 | 9.3 | 117 | 35.7 | 31.9 | 1 | 下坡 | < 0.1 |
| D5 | 8.6 | 0.8 | 31.8 | 0.212 | 0.037 | 2.95 | 145.9 | 9.4 | 247 | 41.2 | 31.5 | 1 | 下坡 | < 0.1 |
| D6 | 4.8 | 1.3 | 54.5 | 0.335 | 0.034 | 2.83 | 224.5 | 11.4 | 139 | 36.1 | 33.2 | 1 | 下坡 | 0.3-0.5 |
| D7 | 4.7 | 0.3 | 24.7 | 0.143 | 0.017 | 2.61 | 127.1 | 4.6 | 136 | 35.3 | 32.5 | 1 | 下坡 | 0.3-0.5 |
| D8 | 5.3 | 0.4 | 17.9 | 0.108 | 0.017 | 2.70 | 93.5 | 6.5 | 146 | 40.6 | 31.9 | 1 | 下坡 | 0.3-0.5 |
| D9 | 5.7 | 0.05 | 20.5 | 0.115 | 0.018 | 2.99 | 104.7 | 3.7 | 166 | 36.5 | 32.7 | 1 | 下坡 | < 0.1 |
| CV/% | 26.07 | 58.37 | 38.34 | 39.48 | 30.19 | 4.51 | 29.95 | 33.45 | 31.26 | - | - | - | - | - |
| D1-D9分别为大怀屿9个滨柃样方;W1-W12分别为大练岛(围营山)12个滨柃样方;J1-J9分别为海坛岛(君山)9个滨柃样方;S: 水溶性盐总量Total content of water-soluble salt;SOM: 土壤有机质Soil organic matter;T-N: 全氮Total nitrogen;T-P: 全磷Total phosphorus;T-K: 土壤全钾Total potassium;A-N: 碱解氮Alkaline hydrolyzable nitrogen;A-P: 有效磷Available phosphorus;A-K: 速效钾Available potassium;G: 坡度Gradient;SA: 坡向Slope aspect;1:北坡North slope 247.5°-292.5°;2:东北坡Northeast slope 292.5°-337.5°;3:西北坡Northwest slope 202.5°-247.5°;4:东坡East slope 337.5°-22.5°;5:西坡West slope 167.5°-202.5°;6:东南坡Southeast slope 22.5°-67.5°;7:西南坡Southwest slope112.5°-167.5°;8:南坡South slope67.5°-112.5°;SP: 坡位Slope position;CD: 郁闭度Crown density;CV: 变异系数Coefficient of variation;-表示无数据 | ||||||||||||||
于每个样方中四角及中心位置选取5株生长发育成熟的滨柃, 每株从东南西北四个方向采样。每一株滨柃取10片成熟、无病虫害、完全展开、边缘完整(均不带叶柄)装入标记好的自封袋, 带回实验室处理。本次共收集到30份滨柃叶片样本, 每份样本含50片, 其中大练岛有样本12份, 海坛岛9份, 大怀屿9份。采用精度为0.001 g电子天平称量叶鲜重, 将叶片于5 ℃的黑暗环境中浸泡12 h后, 用滤纸吸干叶表面水分, 称量叶饱和鲜重[5]。采用精度为0.01 mm电子游标卡尺测量叶厚度(Leaf thickness, LT), 取叶片的叶脉旁上、中、下三个点的平均值。通过CanoScan LiDE 300扫描仪扫描叶片, 运用Image J软件计算叶长(Leaf length, LL)、叶宽(Leaf width, LW)、叶形指数(Leaf index, LI)、叶周长(Leaf perimeter, LP)、叶面积(Leaf area, LA)5个指标。将叶片于85 ℃烘箱烘干至恒重, 48 h后称量叶干重。叶碳含量和叶氮含量采用碳氮元素分析仪测定。叶磷含量采用钼锑抗比色法测定。基于相关文献资料[1, 5-6], 整理各性状指标的计算公式及生态学意义见表 3。
| 叶功能性状 Leaf functional trait |
公式及单位 Formula and unit |
生态学意义 Ecological meaning |
| 叶形指数(LI) Leaf index |
LI(cm)=叶长(cm)/叶宽(cm) | 反映植物叶片生长状况及生长规律 |
| 比叶面积(SLA) Specific leaf area |
SLA(cm2/kg)=叶面积(cm2)/叶干重(kg) | 反映植物获取资源的能力, 与植物光合速率相关 |
| 叶干物质含量(LDMC) Leaf dry matter content |
LDMC(g/kg)=叶干重(g)/叶鲜重(kg) | 反映植物获取资源的能力及防御外界的能力 |
| 比叶重(SLW) Specific leaf weight |
SLW(g/cm2)=叶干重(g)/叶面积(cm2) | 反映植物光合作用性能及获取资源的返还能力 |
| 叶体积(LV) Leaf volume |
LV(cm3)=叶面积(cm2)×叶厚度(cm) | 反映植物对资源的获取及水分的保存 |
| 叶组织密度(LD) Leaf tissue density |
LD(kg/cm3)=叶干重(kg)/叶体积(cm3) | 反映植物水分保持及防御外界的能力 |
| 叶含水量(LWC) Leaf water content |
LWC(%)=(叶鲜重(g)-叶干重(g))/叶鲜重(g)×100% | 反映植物叶片水分状况 |
| 相对水分亏缺(RWD) Relative water deficit |
RWD(%)=(叶饱和重(g)-叶鲜重(g))/(叶饱和重(g)-叶干重(g))×100% | 反映植物持水能力或避免脱水的能力 |
| 叶碳含量(LCC) Leaf carbon content |
LCC(g/kg)=叶片全碳(g)/叶干重(kg) | 是叶片组织的构建成本, 反映植物生长及生理机制调节 |
| 叶氮含量(LNC) Leaf nitrogen content |
LNC(g/kg)=叶片全氮(g)/叶干重(kg) | 与光合速率和植物养分储存相关, 反映植物生长及生理机制调节 |
| 叶磷含量(LPC) Leaf phosphorus content |
LPC(g/kg)=叶片全磷(g)/叶干重(kg) | 与光合速率有关, 反映资源获取能力、生长发育及生理机制调节 |
采用五点交叉取样法进行土壤采样, 钻取5个25 cm深的土壤样品, 将其混合装入编号的自封袋后带回实验室进行处理, 以供土壤化学指标测定。根据鲁如坤[22]测定土壤化学指标的方法, 对土壤的pH值(pH value, pH)、有机质(Soil organic matter, SOM)、全氮含量(Total nitrogen, T-N)、全磷含量(Total phosphorus, T-P)、全钾含量(Total potassium, T-K)、碱解氮(Alkaline hydrolyzable nitrogen, A-N)、有效磷(Available phosphorus, A-P)、速效钾(Available potassium, A-K)、水溶性盐总量(Total content of water-soluble salt, S)9项指标均进行3次重复测定。
2.4 数据处理与分析采用Excel 2019、SPSS 25.0和Canoco 5.0软件进行数据处理和统计。其中采用变异系数(Coefficient of variation, CV)和单因素方差法(One-way analysis of variance, ANOVA)对3个海岛间滨柃各叶性状和土壤因子进行差异显著性分析, 并采用最小显著差异法(Least significant difference, LSD)比较不同数据组间的差异[23]。利用冗余分析法(Redundancy analysis, RDA)分析环境因子对滨柃叶功能性状的影响。
3 结果与分析 3.1 不同海岛土壤因子的差异性根据变异系数法分析结果发现(表 4), 土壤因子各指标CV介于21.61%-54.16%。单因素方差分析表明(表 4), 不同海岛的土壤因子指标间, pH、S、SOM、T-N、T-K、A-N、A-P、A-K存在极显著的差异性(P < 0.01), T-P存在显著的差异性(P < 0.05), 其中S、SOM、T-N、T-P、A-N均以大练岛最高, 以海坛岛最低。T-K、A-P、A-K以大怀屿最高, 海坛岛最低。pH以大怀屿最高, 以大练岛最低。
| 因子 Soil factors |
大练岛 | 海坛岛 | 大怀屿 | CV/% |
| pH值pH value | 4.82±0.33b | 4.88±0.23b | 6.36±1.66a | 21.61 |
| 水溶性盐总量Total content of water-soluble salt/(g/kg) | 0.80±0.28a | 0.39±0.21b | 0.77±0.45a | 54.16 |
| 土壤有机质Soil organic matter/(g/kg) | 49.37±8.34a | 26.56±8.20b | 34.66±13.29b | 36.28 |
| 全氮Total nitrogen/% | 0.25±0.06a | 0.13±0.04b | 0.22±0.08a | 38.65 |
| 全磷Total phosphorus/% | 0.04±0.01a | 0.02±0.00c | 0.03±0.01b | 33.91 |
| 全钾Total potassium/% | 1.66±0.28b | 1.10±0.16c | 2.83±0.13a | 39.37 |
| 碱解氮Alkaline hydrolyzable nitrogen/(mg/kg) | 210.06±38.41a | 129.20±34.18b | 158.24±47.39b | 30.66 |
| 有效磷Available phosphorus/(mg/kg) | 7.12±2.44a | 2.87±0.56b | 8.34±2.80a | 50.20 |
| 速效钾Available potassium/(mg/kg) | 157.83±29.28a | 97.44±45.16b | 161.44±50.47a | 34.98 |
| 同列不同字母表示差异显著(P < 0.05), 相同字母表示差异不显著(P>0.05) | ||||
根据变异系数法分析结果发现(表 5), 滨柃的各叶性状CV介于2.26%-25.78%, 以叶体积(Leaf volume, LV)变异程度最大, 变异程度最小的是叶片含水量(Leaf water content, LWC)。单因素方差分析表明(表 5), 不同海岛滨柃的叶功能性状指标间差异较大, 比叶重(Specific leaf weight, SLW)、相对水分亏缺(Relative water deficit, RWD)、LL、LW、LI、LP、LA、SLA、LV和LPC存在极显著的差异性(P < 0.01), 叶组织密度(Leaf tissue density, LD)、LNC存在显著的差异性(P < 0.05), 说明这些滨柃叶功能性状在群体间多样性丰富, 存在着广泛变异, 具有较好的育种潜力, 其中LL、LW、LT、LP、LA、LV均以大怀屿最高, 以海坛岛最低。SLW以大怀屿为最高, 大练岛最低。RWD以海坛岛最高, 以大怀屿最低。SLA、LPC、LNC以大练岛最高, 大怀屿最低。LD以海坛岛最高, 大练岛最低。不同海岛滨柃的LI、LDMC、LWC和LCC差异不显著。
| 叶功能性状 Leaf functional trait |
大练岛 | 海坛岛 | 大怀屿 | CV/% |
| 叶长Leaf length/cm | 2.90±0.35a | 2.51±0.17b | 3.05±0.11a | 11.67 |
| 叶宽Leaf width/cm | 1.26±0.15a | 1.07±0.07b | 1.28±0.03a | 11.58 |
| 叶形指数Leaf index | 2.34±0.15a | 2.37±0.15a | 2.43±0.09a | 5.47 |
| 叶厚度Leaf thickness/cm | 0.45±0.03b | 0.43±0.03b | 0.54±0.03a | 12.81 |
| 叶周长Leaf perimeter/cm | 7.02±0.77a | 6.04±0.38b | 7.19±0.17a | 10.63 |
| 叶面积Leaf area/cm2 | 3.02±0.65a | 2.23±0.27b | 3.12±0.14a | 20.62 |
| 比叶面积Specific leaf area/(cm2/kg) | 6.96±1.88a | 5.96±0.47b | 5.22±0.46b | 23.14 |
| 叶干物质含量Leaf dry matter content/(g/kg) | 370.97±20.44a | 367.54±6.46a | 372.48±9.05a | 3.77 |
| 比叶重Specific leaf weight/(g/cm2) | 152.18±34.26b | 168.67±12.57b | 205.24±16.95a | 18.68 |
| 叶体积Leaf volume/cm3 | 1.34±0.29b | 0.96±0.16c | 1.65±0.09a | 25.78 |
| 叶组织密度Leaf tissue density/(kg/cm3) | 0.34±0.08b | 0.39±0.01a | 0.37±0.03a | 13.59 |
| 叶片含水量Leaf water content/% | 62.90±2.04a | 63.24±0.65a | 63.11±1.02a | 2.26 |
| 相对水分亏缺Relative water deficit/% | 33.88±5.75b | 40.34±3.94a | 32.27±3.03b | 15.73 |
| 叶碳含量Leaf carbon content/% | 51.10±5.98a | 48.49±5.90a | 53.50±4.08a | 11.03 |
| 叶氮含量Leaf nitrogen content/% | 1.15±0.08a | 1.14±0.10a | 1.05±0.08b | 8.07 |
| 叶磷含量Leaf phosphorus content/(g/kg) | 0.37±0.06a | 0.34±0.03a | 0.31±0.03b | 14.55 |
| 同列不同字母表示差异显著(P < 0.05), 相同字母表示差异不显著(P>0.05) | ||||
采用系统聚类法对3个海岛的30份土壤样品(表 2中的9个土壤因子)进行聚类分析。根据聚类结果绘制聚类树(图 1), 不同海岛样方的9个土壤因子聚为一类。

|
| 图 1 土壤因子组平均聚类树 Fig. 1 Clustering tree group-average strategy of soil factors D1-D9分别为大怀屿9个滨柃样方;W1-W12分别为大练岛(围营山)12个滨柃样方;J1-J9分别为海坛岛(君山)9个滨柃样方 |
采用系统聚类法对3个海岛的30份滨柃叶功能性状(表 6)进行聚类分析。根据聚类结果绘制聚类树(图 2), 在欧氏距离为22时, 3个海岛的30份滨柃样品被分为两大类, 即大练岛、海坛岛的滨柃样品聚为一类(即样方W1-W12, J1-J9), 大怀屿的滨柃样品聚为一类(即样方D1-D9), 其中大练岛、海坛岛为有居民海岛, 在地理位置相互毗邻, 遗传距离相近, 其叶片的功能性状最为接近, 聚为一类。
| 样方编号 Species number |
叶长 LL/cm |
叶宽 LW/cm |
叶形指数 LI/cm |
叶厚度 LT/cm |
叶周长 LP/cm |
叶面积 LA/cm2 |
比叶面积 SLA/ (cm2/kg) |
叶干物 质含量LDMC/ (g/kg) |
比叶重 SLW/ (g/cm2) |
叶体积 LV/ cm3 |
叶组织 密度LD /(kg/cm3) |
叶片含 水量LWC/% |
相对水 分亏缺RWD/% |
全碳含量 LCC/% |
全氮含量 LNC/% |
全磷含量 LPC/ (g/kg) |
| W1 | 3.01 | 1.46 | 2.08 | 0.47 | 7.45 | 3.50 | 9.85 | 376.88 | 101.54 | 1.65 | 0.22 | 62.31 | 40.32 | 54.00 | 1.20 | 0.44 |
| W2 | 2.85 | 1.23 | 2.36 | 0.43 | 6.94 | 2.96 | 5.79 | 359.09 | 172.70 | 1.27 | 0.40 | 64.09 | 23.25 | 53.26 | 1.14 | 0.38 |
| W3 | 2.56 | 1.15 | 2.26 | 0.45 | 6.32 | 2.50 | 5.21 | 394.84 | 192.08 | 1.13 | 0.43 | 60.52 | 32.46 | 51.62 | 1.04 | 0.33 |
| W4 | 3.22 | 1.28 | 2.59 | 0.42 | 7.62 | 3.33 | 6.80 | 358.73 | 147.03 | 1.40 | 0.35 | 64.13 | 30.01 | 42.00 | 1.18 | 0.48 |
| W5 | 3.65 | 1.52 | 2.42 | 0.43 | 8.61 | 4.45 | 10.13 | 318.50 | 98.74 | 1.91 | 0.23 | 68.15 | 28.26 | 42.15 | 1.21 | 0.39 |
| W6 | 2.58 | 1.10 | 2.38 | 0.48 | 6.12 | 2.28 | 5.36 | 382.54 | 186.40 | 1.09 | 0.39 | 61.75 | 37.35 | 50.81 | 1.14 | 0.33 |
| W7 | 2.98 | 1.26 | 2.37 | 0.44 | 7.13 | 3.03 | 5.68 | 384.06 | 175.91 | 1.33 | 0.40 | 61.59 | 30.33 | 56.38 | 0.99 | 0.27 |
| W8 | 2.90 | 1.19 | 2.47 | 0.48 | 7.00 | 2.90 | 5.85 | 378.49 | 170.90 | 1.39 | 0.36 | 62.15 | 34.07 | 47.43 | 1.08 | 0.31 |
| W9 | 3.01 | 1.26 | 2.43 | 0.40 | 7.21 | 3.10 | 7.13 | 355.18 | 140.19 | 1.24 | 0.35 | 64.48 | 32.87 | 46.58 | 1.26 | 0.34 |
| W10 | 2.76 | 1.18 | 2.37 | 0.45 | 6.63 | 2.66 | 5.64 | 382.71 | 177.37 | 1.20 | 0.39 | 61.73 | 33.72 | 51.77 | 1.19 | 0.37 |
| W11 | 2.30 | 1.04 | 2.23 | 0.41 | 5.68 | 1.97 | 6.20 | 383.76 | 161.22 | 0.81 | 0.39 | 61.62 | 43.48 | 63.18 | 1.15 | 0.37 |
| W12 | 3.02 | 1.45 | 2.12 | 0.48 | 7.48 | 3.53 | 9.84 | 376.86 | 102.12 | 1.63 | 0.22 | 62.28 | 40.43 | 53.98 | 1.19 | 0.43 |
| CV/% | 11.89 | 11.78 | 6.21 | 6.32 | 11.03 | 21.68 | 27.04 | 5.51 | 22.51 | 21.78 | 22.18 | 3.25 | 16.98 | 11.69 | 6.70 | 16.05 |
| J1 | 2.27 | 0.99 | 2.32 | 0.42 | 5.44 | 1.79 | 6.07 | 365.94 | 164.69 | 0.75 | 0.39 | 63.41 | 48.21 | 53.06 | 1.21 | 0.34 |
| J2 | 2.47 | 1.16 | 2.15 | 0.41 | 5.95 | 2.26 | 6.47 | 356.97 | 154.60 | 0.93 | 0.38 | 64.30 | 42.50 | 50.57 | 1.29 | 0.38 |
| J3 | 2.73 | 1.09 | 2.56 | 0.46 | 6.29 | 2.37 | 5.62 | 365.34 | 177.89 | 1.09 | 0.39 | 63.47 | 38.19 | 38.26 | 1.20 | 0.30 |
| J4 | 2.40 | 0.98 | 2.45 | 0.44 | 5.67 | 1.93 | 5.57 | 364.69 | 179.59 | 0.85 | 0.41 | 63.53 | 42.71 | 39.51 | 0.99 | 0.38 |
| J5 | 2.78 | 1.20 | 2.36 | 0.47 | 6.67 | 2.72 | 5.55 | 376.98 | 180.15 | 1.28 | 0.38 | 62.30 | 36.66 | 52.07 | 1.04 | 0.37 |
| J6 | 2.48 | 1.05 | 2.44 | 0.43 | 6.05 | 2.19 | 5.92 | 374.12 | 168.95 | 0.94 | 0.39 | 62.59 | 35.55 | 53.77 | 1.14 | 0.33 |
| J7 | 2.40 | 1.06 | 2.28 | 0.43 | 5.98 | 2.23 | 5.94 | 374.95 | 168.34 | 0.96 | 0.39 | 62.50 | 37.21 | 47.86 | 1.15 | 0.30 |
| J8 | 2.62 | 1.02 | 2.57 | 0.44 | 6.43 | 2.37 | 5.57 | 364.76 | 179.58 | 1.04 | 0.41 | 63.52 | 41.26 | 47.64 | 1.18 | 0.32 |
| J9 | 2.40 | 1.11 | 2.20 | 0.36 | 5.88 | 2.20 | 6.93 | 364.07 | 144.27 | 0.79 | 0.40 | 63.59 | 40.81 | 53.64 | 1.05 | 0.38 |
| CV/% | 6.76 | 6.94 | 6.25 | 7.41 | 6.27 | 11.94 | 7.97 | 1.76 | 7.45 | 16.98 | 2.84 | 1.02 | 9.77 | 12.18 | 8.40 | 9.86 |
| D1 | 2.96 | 1.25 | 2.40 | 0.50 | 7.09 | 3.11 | 4.96 | 360.25 | 201.41 | 1.56 | 0.40 | 63.98 | 32.63 | 52.28 | 1.01 | 0.31 |
| D2 | 2.97 | 1.30 | 2.30 | 0.57 | 7.03 | 3.05 | 4.57 | 381.93 | 218.69 | 1.74 | 0.38 | 61.81 | 34.44 | 57.14 | 0.98 | 0.25 |
| D3 | 3.14 | 1.25 | 2.52 | 0.52 | 7.30 | 3.03 | 5.62 | 369.76 | 178.02 | 1.58 | 0.34 | 63.02 | 27.15 | 47.63 | 1.15 | 0.32 |
| D4 | 2.97 | 1.27 | 2.38 | 0.51 | 7.07 | 3.12 | 4.95 | 360.19 | 201.38 | 1.54 | 0.39 | 64.01 | 32.72 | 52.29 | 0.99 | 0.32 |
| D5 | 3.01 | 1.29 | 2.29 | 0.56 | 7.04 | 3.03 | 4.58 | 381.89 | 218.67 | 1.73 | 0.37 | 61.82 | 34.33 | 57.09 | 0.97 | 0.26 |
| D6 | 3.16 | 1.24 | 2.53 | 0.52 | 7.29 | 3.03 | 5.61 | 370.22 | 178.05 | 1.59 | 0.33 | 63.04 | 27.17 | 47.59 | 1.17 | 0.33 |
| D7 | 3.25 | 1.32 | 2.47 | 0.58 | 7.54 | 3.45 | 5.66 | 385.12 | 216.54 | 1.78 | 0.39 | 64.23 | 34.24 | 58.05 | 1.13 | 0.34 |
| D8 | 3.02 | 1.31 | 2.53 | 0.55 | 7.18 | 3.25 | 5.63 | 370.27 | 214.68 | 1.69 | 0.37 | 62.02 | 35.11 | 57.08 | 1.01 | 0.33 |
| D9 | 2.98 | 1.27 | 2.48 | 0.54 | 7.16 | 3.04 | 5.37 | 372.65 | 219.72 | 1.67 | 0.40 | 64.05 | 32.65 | 52.33 | 1.02 | 0.31 |
| CV/% | 3.45 | 2.24 | 3.90 | 5.20 | 2.30 | 4.55 | 8.76 | 2.43 | 8.26 | 5.32 | 6.69 | 1.61 | 9.40 | 7.63 | 7.53 | 10.24 |
| LL: 叶长Leaf length;LW: 叶宽Leaf width;LI: 叶形指数Leaf index;LT: 叶厚度Leaf thickness;LP: 叶周长Leaf perimeter;LA: 叶面积Leaf area;SLA: 比叶面积Specific leaf area;LDMC: 叶干物质含量Leaf dry matter content;SLW: 比叶重Specific leaf weight;LV: 叶体积Leaf volume;LD: 叶组织密度Leaf tissue density;LWC: 叶片含水量Leaf water content;RWD: 相对水分亏缺Relative water deficit;LCC: 叶碳含量Leaf carbon content;LNC: 叶氮含量Leaf nitrogen content;LPC: 叶磷含量Leaf phosphorus content | ||||||||||||||||

|
| 图 2 滨柃叶功能性状组平均聚类树 Fig. 2 Clustering tree group-average strategy of leaf functional traits of Eurya emarginata |
通过对滨柃叶片功能性状与环境因子进行RDA排序, 筛选出CD、T-K、A-P、E、A-K、A-N共6个显著相关的环境因子(P < 0.05), 共同解释量为80.70%, 其中CD、T-K、A-P、E与滨柃叶功能性状呈极显著相关(P < 0.01), 说明CD、T-K、A-P、E是影响滨柃叶功能性状的主要环境因子。由图 3发现: CD、E、A-N分别与LL、LW、LP、LA、SLA、RWD、LWC、LNC、LPC呈正相关, 与其他叶性状参数呈负相关;T-K与LV、LI、LT、SLW、LD呈正相关, 与其他叶性状参数呈负相关;A-P与LL、LW、LI、LP、LA、SLA、LV、LWC、LNC、LPC呈正相关, 与其他叶性状参数呈负相关;A-K与RWD、LDMC、LCC、LD、SLW、LT呈正相关, 与其他叶性状参数呈负相关。

|
| 图 3 滨柃叶功能性状与环境因子的冗余分析排序 Fig. 3 Redundancy analysis (RDA) ordination of leaf functional traits of Eurya emarginata and environmental factors LL, 叶长Leaf length;LW, 叶宽Leaf width;LI, 叶形指数Leaf index;LT, 叶厚度Leaf thickness;LP, 叶周长Leaf perimeter;LA, 叶面积Leaf area;SLA, 比叶面积Specific leaf area;LDMC, 叶干物质含量Leaf dry matter content;SLW, 比叶重Specific leaf weight;LV, 叶体积Leaf volume;LD, 叶组织密度Leaf tissue density;LWC, 叶片含水量Leaf water content;RWD, 相对水分亏缺Relative water deficit;LCC, 叶碳含量Leaf carbon content;LNC, 叶氮含量Leaf nitrogen content;LPC, 叶磷含量Leaf phosphorus content;CD, 郁闭度Crown density;T-K, 土壤全钾Total potassium;A-P, 有效磷Available phosphorus;E, 海拔Elevation;A-K, 速效钾Available potassium;A-N, 碱解氮Alkaline hydrolyzable nitrogen |
海岛生境空间异质化明显, 已有的研究表明, 不同的海拔高度其水热条件不同, 土壤中的水分可能随着海拔升高逐渐降低[14], 海岛典型常绿阔叶林和灌草丛因枯落物产量及分解速度不同, 土壤理化性质也不同[13], 不同坡度的植物群落由于受雨水冲刷程度不同, 土壤理化性质可能差异较大[15]。本研究, 同一海岛不同样方土壤理化因子的变异系数中, 大练岛围营山在6.85%-34.95%之间, 海坛岛君山在4.67%-53.20%之间, 大怀屿在26.07%-58.37%之间, 3个不同海岛全部样方土壤理化因子的变异系数在21.61%-54.16%之间, 同一海岛不同样方间, 3个海岛全部样方间都是pH变异系数最小, S变异系数最大。其他土壤有机质, 土壤养分含量(A-N、A-K、A-P等), 同一海岛不同样方间和3个不同海岛全部样方间也在不同程度变动着, 聚类分析结果表明, 不同海岛样方的9个土壤理化因子可能聚为一类(图 1), 因此, 样方土壤因子的数理统计分析结果表明, 同一海岛的不同样方可能因位置不同而区划为异质土壤生境。冗余分析结果表明, 3个海岛30个全部样方的土壤养分指标(A-N、A-K、A-P)与海拔高度和样方郁闭度具有相关性(图 3)。本研究中, 郁闭度大的群落为常绿阔叶林, 乔木层以台湾相思为建群种, 灌木层以滨柃为优势种, 是海岛植被演替后期的群落类型, 而郁闭度小的群落主要为常绿灌丛群落, 以滨柃为建群种的群落, 是海岛植被演替中后期的群落类型, 有研究表明, 随着演替的进行, 土壤养分和水分逐渐增加, 土壤pH逐渐下降[13, 24]。可以推测, 本研究中相对于常绿灌丛群落, 常绿阔叶林群落中的台湾相思的根瘤菌具有较好的固氮作用, 能增加土壤的氮元素, 台湾相思冠幅大, 降低强风影响, 枯落物产量高且分解快, 有效维持土壤肥力及改善土壤理化性质。因此, A-N和A-P与CD呈正相关关系。而A-N和A-P与海拔呈正相关可能是因为海拔高处的滨柃采样样方刚好是台湾相思林。A-K与CD和海拔呈负相关可能是因为, 海水中富含钾离子, 能以盐雾的形式经风吹散落在土壤表层, 而海拔高可能离海岸线距离更远, 盐雾不易吹落入土壤中, 郁闭度高的常绿阔叶林乔木层的树冠能有效阻断盐雾散落在土壤中。
4.2 滨柃叶功能性状的变动海岛植物叶片作为暴露于大气环境中的植物器官对生境因子的变动比较敏感, 主要表现为叶功能性状的变动, 以适应空间异质性[25]。本研究中, 同一海岛不同样方的滨柃叶功能性状16项指标的变异系数中, 大练岛围营山在3.25%-27.04%之间, 海坛岛君山在1.02%-16.98%之间, 大怀屿在1.61%-10.24%之间, 3个不同海岛全部30个样方滨柃叶功能性状16项指标的变异系数在2.66%-22.99%之间, 同一海岛不同样方间, 3个海岛全部样方间都是LWC变异系数最小, 大练岛围营山12个样方采集的滨柃, 叶功能性状指标变异系数最大的是SLA, 海坛岛君山9个样方采集的滨柃, 叶功能性状指标变异系数最大的是LV, 大怀屿9个样方采集的滨柃, 叶功能性状指标变异系数最大的是LPC, 3个海岛全部30个样方采集的滨柃, 叶功能性状指标变异系数最大的是SLA。相比于30个滨柃采样样方土壤理化因子的变动系数, 30个样方滨柃样品叶功能性状的变异系数更小, 聚类分析结果表明, 大练岛和海坛岛的滨柃样品叶功能性状聚为一类, 大怀屿滨柃样品叶功能性状聚为独立的一类(图 2), 这与不同海岛样方的9个土壤理化因子可能聚为一类不一致。出现这现象的原因在于: (1)相关研究表明功能性状的种内变异系数不超过30%[5, 26], 说明植物在环境中有一定的生态幅, 其性状变异幅度是有限的[27];(2)不同生境因子对叶功能性状的影响不同, 可能一个因子是正向的, 另一个因子却是负向的, 如, 本研究中, 冗余分析表明, A-N和A-P与LL、LW、LP、LA、SLA、LWC、LNC、LPC呈正相关, 而A-K与这些叶功能性状指标呈负相关。
4.3 三个海岛30个样方叶功能性状与土壤、地形因子和群落郁闭度的相关性植物根据环境差异化进行不同功能性状间的调控以形成最佳的性状组合, 来实现最大程度利用资源, 避免逆境影响, 以提高生境适合度和扩宽生态幅[28]。本研究表明, CD、E和A-N表现出强烈的协同性, CD、E和A-N分别与滨柃的LL、LW、LP、LA、SLA、RWD、LWC、LNC、LPC呈正相关, 与滨柃其他叶性状参数(LI、LT、LDMC、SLW、LV、LD和LCC)呈负相关(图 3)。土壤养分(如本研究的A-N)越高, 滨柃叶功能性状倾向于有更大的LA、更高的SLA, 更低的LT、LDMC和LD, 因此, 在养分更充足的情况下, 滨柃倾向于更快速生长的策略。这与许多研究结果类似[5, 15, 29]。值得一提的是, 土壤养分(A-N)的提高, 可能与植物群落结构有关, 本研究中, 部分样方为台湾相思群落, 其中滨柃为灌木层优势种, 上有台湾相思遮荫, 光照较弱, CD高, 而台湾相思因具有固氮菌, A-N可能也相对高, 另外一部分样方为滨柃灌丛, 这些灌丛的滨柃暴露于强光照下, CD低, 群落结构中因无台湾相思, 因而土壤A-N相对较低。同时, 相对于滨柃灌丛群落, 滨柃为灌木层优势种的台湾相思群落中, 台湾相思冠幅大, 枯落物产量高且分解快, 有效提高了养分含量。海拔高度的变化能改变生境的水热条件, 进而影响植物的功能性状, 一般大陆性植物功能性状的研究认为, 海拔升高, 植物倾向于提高养分利用率和水分保存率的保守功能性状[29-30], 但有很多另外[31-32], 因此, 海拔的变动影响植物功能性状首先需要具体分析, 海拔变动与水热条件变化的关系。关于海岛植物功能性状的研究, 张增可等[14]和张秀芳等[15]的研究发现, 叶片性状表现出随海拔上升的资源保守策略。本研究中, 海拔的升高, 对植物功能性状的影响与土壤A-N和CD对植物功能性状的影响高度一致, 随海拔升高, 植物功能性状倾向有利于捕获资源的开放策略, 而不是提高养分利用率和水分保存率的保守策略, 原因可能是, (1)低海拔滨柃的土壤生境更易受到海水盐度的影响, 如海水盐分以盐雾形式通过风作用沉降于土壤[33], 本研究中, 海拔和土壤总水溶性盐呈负相关(偏相关分析, P < 0.05);(2)有研究表明降雨量随海拔升高而增大[34]。因此, 与高海拔滨柃生境相比, 低海拔滨柃生长于风大、干旱、盐分高和贫瘠的恶劣生境, 受人为干扰程度高, 滨柃叶功能性状倾向于具有更小的LA, 更低的SLA和LWC, 更高的LDMC和LD, 这些功能性状的组合有利于防止水分流失和资源消耗。综上所述, 30个研究样方中, 光照不同, 植物群落结构特征可能影响土壤养分, 海拔变动可能导致土壤盐分和水热条件变化, 是CD、E和A-N影响滨柃功能性状指标具有协同性的主要原因。
N、P和K是植物必须的养分元素, 本研究30个样方中, 土壤A-N与LNC, 土壤A-P与LPC呈正相关, 而土壤A-N和A-P含量对植物功能性状的影响也呈现较为一致的协同性, A-N和A-P分别与滨柃的LL、LW、LP、LA、SLA、LWC、LNC、LPC呈正相关, 而与滨柃其他叶性状参数(LT、LDMC、SLW、LD和LCC)呈负相关(图 3), 仅土壤A-N与RWD呈正相关, 而土壤A-P与RWD呈负相关, 土壤A-N与LI和LV呈负相关, 而土壤A-P与LI和LV呈正相关。本研究土壤A-N与RWD呈正相关, 而土壤A-P与RWD呈负相关的原因可能是, 水分与氮磷在叶各项生理活动中的互作方式不同, 有研究表明, 遭受水分胁迫时, 叶氮含量的增加有利于气孔关闭, 增进叶细胞保水[35], 而叶磷含量增加能增大气孔导度, 主要促进光合作用和呼吸作用[36], 具体的机理机制需进一步研究。K, 与N和P一样, 是植物必需的养分元素, 本研究中, 随土壤A-K的增加, 叶保水功能性状增强, 如RWD、LT、LDMC、LD和LCC增大, 而与生长相关的功能性状降低, 如LA和SLA等。土壤中的K作为可为植物吸收的必需元素, 没有与土壤N和P一样, 对叶功能性状的影响表现出一致的协同性, 可能原因在于, 土壤A-K与土壤总水溶性盐具正相关(偏相关分析P < 0.05), 土壤总水溶性盐升高, 植物易遭受水分胁迫, 叶倾向于具有保水分和养分的功能性状[37]。
5 结论在本研究中, 滨柃叶性状具有较大的环境可塑性, 其中以LV变异程度最大, 变异程度最小的是LWC。不同海岛间滨柃大部分叶性状指标差异显著, 各岛的环境因子均存在显著差异。滨柃叶片功能性状通过一定的性状变异和性状组合适应海岛生境的变化, 其中显著影响滨柃各叶性状变异的主要环境因子为CD、T-K、A-P、E、A-K、A-N, 不同海岛的滨柃叶性状对不同异化生境响应和适应程度各不相同, 表明各海岛的滨柃对生境的适应策略存在较大差异, 使得滨柃成为海岛植被中广泛分布的优势群落格局。通过研究滨柃叶功能性状特征及其与环境因子的关系, 有利于更好理解海岛植物适应环境所采取的生态策略, 可为滨柃资源的引种驯化、选育工作及开展其遗传改良等提供理论指导。但由于本文未对人为干扰因子、风因子、土壤含水量、容重等数据收集可能会造成研究结果出现偏差。
| [1] |
张增可, 王齐, 吴雅华, 刘兴诏, 黄柳菁. 基于CiteSpace植物功能性状的研究进展. 生态学报, 2020, 40(3): 1101-1112. |
| [2] |
Zhang S H, Zhang Y, Xiong K N, Yu Y H, Min X Y. Changes of leaf functional traits in karst rocky desertification ecological environment and the driving factors. Global Ecology and Conservation, 2020, 24: e01381. DOI:10.1016/j.gecco.2020.e01381 |
| [3] |
Cornelissen J H C, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich D E, Reich P B, Steege H T, Morgan H D, van der Heijden M G A, Pausas G J, Poorter H. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4): 335-380. DOI:10.1071/BT02124 |
| [4] |
刘贵峰, 刘玉平, 达福白乙拉, 程伟燕, 高学磊, 姜丽丽. 大青沟自然保护区主要森林群落优势种的叶性状. 生态学报, 2017, 37(14): 4646-4655. |
| [5] |
罗达, 史彦江, 宋锋惠, 李嘉诚. 38个榛种质资源叶功能性状与光合特征变异及其相关性. 生态学杂志, 2021, 40(1): 11-22. |
| [6] |
靳莎, 闫淑君, 黄柳菁, 陈莹, 马雯雯, 王云霄, 王喆. 植物叶功能性状间的权衡研究进展. 四川林业科技, 2019, 40(5): 96-103. |
| [7] |
徐海东, 查美琴, 成向荣, 汤胜, 周恒发, 虞木奎. 林下引入耐阴树种功能性状特征及其与生长的关系. 生态学报, 2020, 40(19): 6948-6957. |
| [8] |
Li D D, Zong J Q, Chen J B, Guo H L, Wang Y, Li J J, Liu J X. Leaf photosynthesis and CO2 assimilation in response to low nitrogen supply in bermudagrass (Cynodon dactylon (L.) Pers.). Acta Physiologiae Plantarum, 2020, 42(6): 96. DOI:10.1007/s11738-020-03077-1 |
| [9] |
陈晨, 刘丹辉, 吴键军, 康慕谊, 张金屯, 刘全儒, 梁钰. 东灵山地区辽东栎叶性状与地形因子关系. 生态学杂志, 2015, 34(8): 2131-2139. |
| [10] |
黄勇, 谢一青, 李志真, 王开良, 林萍, 姚小华. 小果油茶表型多样性分析. 植物遗传资源学报, 2014, 15(2): 270-278. |
| [11] |
黄柳菁, 张增可, 郑俊鸣, 黄少君, 林丽丽, 王齐, 邓传远, 刘兴诏. 大陆性海岛野生植物功能性状特征及其演变趋势--以平潭岛为例. 热带亚热带植物学报, 2017, 25(5): 429-437. |
| [12] |
路兴慧, 丁易, 臧润国, 邹正冲, 黄卢标. 海南岛热带低地雨林老龄林木本植物幼苗的功能性状分析. 植物生态学报, 2011, 35(12): 1300-1309. |
| [13] |
张增可, 郑心炫, 林华贞, 林欣, 黄柳菁. 海岛植物不同演替阶段植物功能性状与环境因子的变化规律. 生态学报, 2019, 39(10): 3749-3758. |
| [14] |
张增可, 吴雅华, 王齐, 季凌波, 黄柳菁. 环境因子对海岛植物茎、叶功能性状的影响. 广西植物, 2020, 40(3): 433-442. |
| [15] |
张秀芳, 穆振北, 林美娇, 江淼华, 巩嘉欣, 游巍斌. 琅岐岛4种优势植物叶功能性状及其影响因子. 应用与环境生物学报, 2020, 26(3): 667-673. |
| [16] |
聂森. 滨柃植物群落动态及结构特征研究[D]. 北京: 中国林业科学研究院, 2014.
|
| [17] |
李加庆. 海坛岛典型灌丛群落结构特征及环境解释[D]. 福州: 福建农林大学, 2019.
|
| [18] |
Zheng J L, Zhao L Y, Shen B, Jiang L H, Zhu A Y. Effects of salinity on activity and expression of enzymes involved in ionic, osmotic, and antioxidant responses in Eurya emarginata. Acta Physiologiae Plantarum, 2016, 38(3): 70. DOI:10.1007/s11738-015-2040-3 |
| [19] |
陈闻, 赵颖, 叶正钱, 王美琴, 王晶, 王国明. 干旱胁迫对5个海岛树种生长及生理特性的影响. 浙江农林大学学报, 2013, 30(4): 490-498. |
| [20] |
张椿芳. 三种柃木属植物组织培养技术研究[D]. 南京: 南京林业大学, 2007.
|
| [21] |
何雅琴, 刘健行, 魏凯, 史晓洁, 陈国杰, 曹晟烜, 邓传远. 基于灰色关联度的平潭野生草本花卉开发应用评价. 草业科学, 2020, 37(8): 1497-1507. |
| [22] |
鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 1999. |
| [23] |
刘亚斌, 郭素娟. 迁西地区39份板栗种质叶片的功能性状多样性. 东北林业大学学报, 2020, 48(4): 8-14, 19-19. DOI:10.3969/j.issn.1000-5382.2020.04.002 |
| [24] |
宋光满, 韩涛涛, 洪岚, 张玲玲, 李晓波, 任海. 演替过程中植物功能性状研究进展. 生态科学, 2018, 37(2): 207-213. |
| [25] |
余华, 潘宗涛, 陈志强, 陈志彪, 冯柳俊. 氮添加对侵蚀退化红壤化学性质及芒萁叶功能性状的影响. 应用与环境生物学报, 2021, 27(1): 46-53. |
| [26] |
张凯, 侯继华, 何念鹏. 油松叶功能性状分布特征及其控制因素. 生态学报, 2017, 37(3): 736-749. |
| [27] |
Hallik L, Niinemets V, Wright I J. Are species shade and drought tolerance reflected in leaf-level structural and functional differentiation in northern Hemisphere temperate woody flora?. New Phytologist, 2009, 184(1): 257-274. DOI:10.1111/j.1469-8137.2009.02918.x |
| [28] |
王海珍, 徐雅丽, 代雅琦, 韩路. 胡杨异形叶光合生理特性及其与叶形态的关系. 塔里木大学学报, 2020, 32(3): 35-43. DOI:10.3969/j.issn.1009-0568.2020.03.005 |
| [29] |
刘思文, 艾也博, 刘艳红. 北京松山油松叶功能性状沿海拔梯度的变化及其环境解释. 北京林业大学学报, 2021, 43(4): 47-55. |
| [30] |
杨锐, 张博睿, 王玲玲, 杨建军, 苏文华, 张光飞, 杨波, 周睿. 元谋干热河谷植物功能性状组合的海拔梯度响应. 生态环境学报, 2015, 24(1): 49-56. |
| [31] |
罗璐, 申国珍, 谢宗强, 喻杰. 神农架海拔梯度上4种典型森林的乔木叶片功能性状特征. 生态学报, 2011, 31(21): 6420-6428. |
| [32] |
Zhang L, Luo T X, Liu X S, Wang Y. Altitudinal variation in leaf construction cost and energy content of Bergenia purpurascens. Acta Oecologica, 2012, 43: 72-79. DOI:10.1016/j.actao.2012.05.011 |
| [33] |
侯梦莹, 李芊芊, 袁甜甜, 王文卿, 范志阳, 洪赐在. 南方滨海地区盐雾沉降的时空分布--以福建古雷半岛为例. 生态学杂志, 2019, 38(8): 2524-2530. |
| [34] |
李颖俊, 王尚义, 牛俊杰, 方克艳, 李晓岚, 栗燕, 布文丽, 李玉晗. 芦芽山华北落叶松(Larix principis-rupprechtii)树轮宽度年表对气候因子的响应. 生态学报, 2016, 36(6): 1608-1618. |
| [35] |
陈立松, 刘星辉. 水分胁迫对荔枝叶片氮和核酸代谢的影响及其与抗旱性的关系. 植物生理学报, 1999, 25(1): 49-56. DOI:10.3321/j.issn:1671-3877.1999.01.008 |
| [36] |
刘霞, 张秀清, 张慧, 李宗新, 刘林臣, 翟学旭, 刘书聪, 王庆成. 水分胁迫条件下玉米鲁单818植株体内磷素循环及磷钙关系研究. 山东农业科学, 2014, 46(1): 61-65. |
| [37] |
李群, 赵成章, 赵连春, 王继伟, 文军. 秦王川盐沼湿地芦苇叶片比叶面积与光合效率的关联分析. 生态学报, 2019, 39(19): 7124-7133. |