 2022, Vol. 42
2022, Vol. 42文章信息
- 李茂萍, 缪宁, 刘世荣
- LI Maoping, MIAO Ning, LIU Shirong
- 固氮树种旱冬瓜对退化林地土壤修复和林下植被重建的生态驱动效应
- Effects of nitrogen-fixing tree species Alnus nepalensis on the degraded soils and understory restoration in the upper reaches of the Jinsha River, China
- 生态学报. 2022, 42(6): 2321-2330
- Acta Ecologica Sinica. 2022, 42(6): 2321-2330
- http://dx.doi.org/10.5846/stxb202101120121
-
文章历史
- 收稿日期: 2021-01-12
- 网络出版日期: 2021-11-26





2. 四川大学生命科学学院, 教育部生物资源与生态环境重点实验室, 成都 610065;
3. 中国林业科学研究院森林生态环境与保护研究所, 国家林业和草原局森林生态环境重点实验室, 北京 100091
2. Key Laboratory of Bio-Resource and Eco-Environment of Ministry of Education, College of Life Sciences, Sichuan University, Chengdu 610065, China;
3. Key Laboratory of Forest Ecology and Environment of National Forestry and Grassland Administration, Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China
土壤退化是指土壤肥力衰退, 其结构改变、养分元素和有机质含量下降, 土壤的生产能力或土地利用和环境调控潜力下降甚至完全丧失[1]。植树造林是修复退化土壤的重要措施之一[2];一般而言, 与固氮微生物共生产生固氮功能的固氮植物[3]对土壤的修复往往比非固氮植物见效更快[4-6]。因此, 造林中常常利用固氮植物改良土壤[7-9]。固氮树种往往能显著增加土壤有机质和土壤氮素[10-11], 比如Wang等研究表明, 固氮树种人工林中表层土有机质含量和氮含量比非固氮树种人工林分别高40%-50%和20%-50%[12]。固氮树种除了由于其固氮能力而直接影响土壤外, 还可以通过其自身与土壤间的作用影响其他植物, 如: 固氮植物可通过根系分泌[13]或者凋落物分解[14]而为其他植物提供丰富的可利用氮素[15], 进而促进其他植物的生长[16], 从而提高生产力[17], 促进植被的次生演替[18]等, 在生态系统中有着重要的生态功能。
桤木属(Alnus spp.)树种因其较高的固氮速率和对土壤的改善作用而被认为是重要的固氮植物[19], 主要通过根系分泌物[20]、土壤中游离的固氮相关微生物[19]及叶片凋落物、根系和根瘤的降解三种方式提升土壤的氮元素含量。有研究发现桤木属树种叶片氮含量比其他植物叶片氮含量高, 叶片凋落物降解后会给土壤提供大量氮元素, 可提升土壤肥力[20]。桤木属树种与土壤间的这种反馈能对其他植物产生正效应, Binkley等研究发现红桤木(A. rubra)能提升杉木林的净生产力和生物量, 促进杉木生长[21];Delver和Post的研究发现欧洲桤木(A. glutinosa)能提升苹果树叶片氮的含量, 促进其嫩芽的生长[22]。作为森林生态系统的重要组分, 桤木林的林下植物能降低土壤侵蚀和保土保肥[23]、维持森林多样性和稳定性[24]及调节能量流动[25]。
旱冬瓜(Alnus nepalensis D. Don)是分布印度、不丹、尼泊尔及中国西南和广西的乡土树种, 因其高效的固氮能力而在印度人工林中得到较为广泛的应用[26-28]。中国是世界上人工林面积最大的国家[29], 作为乡土树种, 旱冬瓜在云南省人工林中得到了推广应用[30]。以往主要研究旱冬瓜对土壤理化性质的影响[27-28]、土壤微生物及酶活性的影响[31]、旱冬瓜凋落物降解及营养周转[26], 但对旱冬瓜在不同混交模式、年龄结构和人工与天然起源森林中的固氮效应及其变化规律, 特别是旱冬瓜对退化林地土壤修复过程中如何影响林下植被的生长等方面研究较少。
金沙江上游的云南省富源县占马地林区, 森林植被及土壤退化严重、特征明显, 乡土树种旱冬瓜作为固氮树种是该地区退化植被及土壤恢复的优良树种。然而, 旱冬瓜林对严重退化土壤功能修复与改善的效果缺乏系统研究, 特别是旱冬瓜-土壤的反馈作用是否促进着林下植被的恢复、提升保土保肥能力、提高生物多样性的作用尚不清楚。因此, 研究旱冬瓜与土壤间的相互作用关系, 以期阐明旱冬瓜对严重退化土壤功能的修复和林下植被生长的影响, 为长江上游土壤严重退化地区植被恢复树种选择提供理论基础。
1 研究方法 1.1 研究区域概况研究区位于云南省富源县中安镇占马地小流域, 该区域地处金沙江上游, 海拔1920-2213 m, 属高原季风气候, 干湿季分明, 5-10月为雨季, 年降雨量1100-1300 mm, 年平均日照时数1820-1900 h, 年均气温为13.8℃。土壤主要为玄武岩和石灰岩发育而来的黄红壤, 土壤偏酸性(如本研究的土壤pH值在4.02-6.02之间), 土层深厚。由于沟深坡陡, 耕地较多, 水土流失严重[32-33]。该区原生植被退化较为严重, 现有森林以云南松(Pinus yunnanensis Franch.)次生林和栎(Quercus spp.)林为主, 不少林地退化为灌木林地和草地, 有机质含量下降, 营养元素减少, 土壤及植被退化特征明显。为修复退化土壤与植被, 重建生态系统, 该地区1998年以来实施了封山育林和人工造林及退耕还林等一系列生态恢复措施。
1.2 旱冬瓜的土壤修复实验设计及野外调查为探究不同旱冬瓜种植模式对土壤和林下植被的影响, 于2001年选耕种历史及土壤基本条件一致的耕地, 采用旱冬瓜与云南松的实生苗, 分别按不同比例混交种植: 旱冬瓜纯林、旱冬瓜5∶云南松5及云南松纯林。为研究旱冬瓜林龄对土壤和林下植被的影响, 分别于2010年和2013年在立地条件相近的荒草地上种植旱冬瓜纯林。为进一步验证旱冬瓜对改善土壤和林下植被生长的影响, 选取了旱冬瓜为优势种的天然林、旱冬瓜和云南松混交林及云南松为优势种的三种天然林林分, 探究天然起源的林分中旱冬瓜对土壤和林下植被的影响。
2018年4月开展了野外调查和采样, 选取2001年退耕地上种植的人工林中选取旱冬瓜纯林(AP)、旱冬瓜5∶云南松5(MP)及云南松纯林(PP)三种林分类型的样地, 研究旱冬瓜不同种植模式对退化土壤和林下植被的影响。选取2010年和2013年在荒草地上造林的旱冬瓜纯林及同条件的荒草地作为对照样地, 研究旱冬瓜不同林龄对土壤和林下植被的影响。在旱冬瓜为优势种的天然次生林(NAF)、旱冬瓜与云南松混交的天然次生林(NMF)及云南松天然次生林(NPF)中各选取一块样地, 探究天然林中旱冬瓜对土壤及林下植被的影响, 样地概况见表 1。在每块均不小于2 hm2的样地中分别设置8个8 m×4 m的样方, 调查其中所有的乔木树种、胸径、树高及存活状况, 并调查灌木物种、地径、高度和萌株数。每个样方中分别随机取3个表层(0-5 cm)土样, 均匀混合后一分为二, 一份现场称重, 用于测量土壤含水率, 另一份用于分析其理化性质。每个样方的四角及中心分别设置一个1 m×1 m小样方, 测量草本物种种类、多度及地上生物量[34], 五个小样方物种累计值计算每个样方中草本物种多样性;五个小样方的平均值作为每个样方中地上生物量数据。
| 林分组别 Forest group |
林分类型 Forest type |
优势种 Dominant species |
林龄/a Stand age |
林地起源 Forest origin |
旱冬瓜重要值 Importance value of A. nepalensis |
旱冬瓜材积 Timber volume of A. nepalensis /m3 |
| 旱冬瓜混交配置 | 旱冬瓜人工林 | 旱冬瓜 | 17 | 退耕地 | 0.88 | 5.64 |
| Species configuration | 人工混交林 | 旱冬瓜5:云南松5 | 17 | 退耕地 | 0.35 | 1.51 |
| 云南松人工林 | 云南松 | 17 | 退耕地 | 0.1 | 0.38 | |
| 林分年龄 | 荒草地 | 荒草地 | - | 荒草地 | 0 | 0 |
| Stand age | 5年生林分 | 旱冬瓜 | 5 | 荒草地 | 1 | 0.42 |
| 8年生林分 | 旱冬瓜 | 10 | 荒草地 | 1 | 0.64 | |
| 优势种 | 旱冬瓜天然林 | 旱冬瓜 | - | 天然林 | 0.61 | 5.58 |
| Dominant species | 天然林混交 | 旱冬瓜与云南松 | - | 天然林 | 0.39 | 2.74 |
| 云南松天然林 | 云南松 | - | 天然林 | 0.03 | 0.01 |
采用重铬酸钾氧化容量法测定土壤有机质[35];凯氏定氮法测定总氮、速效氮[36];依据鲍士旦的《土壤农化分析》中钼锑抗比色法测定土壤总磷、速效磷, 碱熔-火焰光度法测定总钾, NH4OAc浸提, 火焰光度法测定速效钾[37], 并根据土壤鲜重和干重测定了土壤含水量。本研究中采用上述8个指标计算的土壤肥力综合指标(Integrated Fertility Index, IFI)反映土壤养分综合质量, 其计算公式如下:
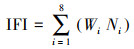
|
(1) |
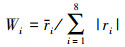
|
(2) |
式(1)中Wi是第i个土壤理化指标的权重, Ni是第i个土壤理化指标的隶属度;式(2)中ri为第i个土壤理化指标相关系数平均值, |ri|为第i个土壤理化指标与其他7个指标相关关系的绝对值。
某一指标的权重指该指标在土壤质量水平衡量中作用大小的体现, 这里土壤指标权重采用相关系数法确定[38-39]。某项土壤指标的隶属度值, 则表示该指标的状态值。本研究中所选取的8个土壤指标的作物效应曲线呈S型[38], 因此, 其对应的隶属度函数也采用S型, 并建立如下隶属度函数:
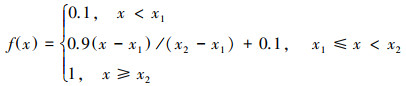
|
式中f(x)是隶属度函数, x是土壤指标的实际测定值, x1和x2是函数的拐点值, 根据研究区土壤的实际情况, 本研究利用88份合格的土壤养分指标计算x1和x2这两个拐点值, 各营养组分的拐点值见表 2。
| 有机质 OM/(g/kg) |
总氮 TN /(g/kg) | 总磷 TP /(g/kg) | 总钾 TK /(g/kg) | 速效氮 AN /(mg/kg) | 速效磷 AP/(mg/kg) | 速效钾 AK /(g/kg) | 土壤含水率 SWC |
|
| x1 | 20.78 | 0.20 | 0.16 | 4.95 | 204.24 | 2.11 | 0.11 | 0.13 |
| x2 | 66.15 | 0.51 | 0.83 | 16.36 | 595.73 | 7.32 | 0.66 | 0.25 |
| x1是土壤养分隶属度函数的拐点值x1;x2是土壤养分隶属度函数的拐点值x2;OM,有机质 Organic matters; TN,总氮 Total nitrogen; TP,总磷 Total phosphorus; TK,总钾 Total potassium; AN,速效氮 Available nitrogen; AP,速效磷 Available phosphorus; AK,速效钾 Available potassium; SWC,土壤含水率 Soil water content ratio | ||||||||
利用旱冬瓜树高及胸径, 根据《云南省一元立木材积表》[40], 估计旱冬瓜单株材积, 并计算每个样方中旱冬瓜的材积总和, 据此定量评价旱冬瓜总材积对土壤及林下植被的影响。本研究测定了每个样方中小样方对应的草本地上部分干重, 作为草本生长状况的指标, 并计算草本的Shannon-Wiener指数。根据灌木的地径、高度及萌株数量估算每个样方中灌木的总蓄积量, 再根据蓄积量与生物量的换算系数计算灌木生物量[41], 以样方灌木生物量与各样方灌木生物量均值间的差值除以该均值所得的比值作为衡量灌木生长状况的系数。根据物种和多度计算各样方中灌木的Shannon-Wiener指数。
灌木生长状况计算公式如下:
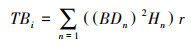
|
(4) |
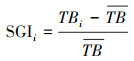
|
(5) |
式(4)中, TBi为第i个样方中的灌木生物量, BDn是样方i中第n株灌木的地径, Hn是样方i中第n株灌木的株高, r是灌木材积换算生物量的系数;式(5)中SGIi是第i个样方中的灌木生长系数, TBi是i个样方中的灌木生物量, TB是样地中8个样方灌木生物量的均值。
Shannon-Wiener指数计算公式如下:
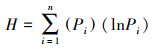
|
(6) |
式中, H是样方中灌木(或草本)的Shannon-Wiener指数;Pi是样方中第i个物种的个体数占样方中总个体数的比例。
1.5 统计分析利用单因素方差分析分别对比各组不同样地中总有机质、总N、速效N及土壤养分综合指标的大小, 分析旱冬瓜对林地土壤的影响;此外, 用Spearman相关分析分析了旱冬瓜材积与上述各指标间的线性关系。利用单因素方差分析比较了各组不同样地中草本地上部分干重、草本多样性指数、灌木生长状况及灌木多样性;用Spearman相关分析分析了各组样地中土壤养分与林下植被生长、多样性间的相关关系。为进一步探究旱冬瓜-土壤反馈对林下植被生长的影响, 本研究以立地条件为控制因子, 用偏相关分析(Partial correlation)法分析了所有样地旱冬瓜材积、土壤养分和林下植被生长间的相关关系。考虑立地条件的前提下, 用结构方程模型(Structural Equation Model, SEM)分析了研究区内旱冬瓜材积、土壤养分对林下植被生长的影响。SEM模型中以土壤有机质、土壤总氮及土壤肥力综合指标反映土壤养分状况。
所有分析基于R 3.3.2、SPSS 16.0和Excel 2007完成, 制图和制表基于Origin Pro 8.5及Excel 2007完成。显著性水平α=0.05。
2 研究结果 2.1 旱冬瓜对土壤养分和肥力的影响对比分析各不同旱冬瓜种植模式的土壤养分, 发现不同模式间土壤有机质、总氮及速效氮无显著差异(P>0.05)(图 1), 且这三种土壤养分不随因种植比例不同而造成的旱冬瓜材积变化而呈相应的线性变化(图 1)。但是, 旱冬瓜纯林的土壤肥力综合指标显著高于混交林和云南松纯林(P < 0.05), 同时土壤肥力综合指标随旱冬瓜材积的增加而增大(图 1)。与旱冬瓜种植模式对土壤的影响有所不同, 旱冬瓜种植时间对土壤养分的影响较为明显。林龄8 a的旱冬瓜纯林土壤有机质、总氮、速效氮及土壤肥力综合指标都显著大于林龄5 a的旱冬瓜纯林和荒草地(P < 0.05)(图 1), 而且这些养分指标都与旱冬瓜材积呈显著的线性正相关(图 1)。在天然林中, 除速效氮在三个林分中无显著差异外(P>0.05), 旱冬瓜纯林的土壤有机质、总氮及土壤肥力综合指标都显著大于旱冬瓜和云南松的混交林, 云南松林中的这三个指标最小(P < 0.05)(图 1);样方中旱冬瓜材积与土壤有机质、总氮及土壤肥力综合指标均显著正相关(图 1)。

|
| 图 1 旱冬瓜对土壤养分的影响 Fig. 1 The impact of A. nepalensis on the soil nutrient OM: 有机质 Organic matte;TN: 总氮 Total nitrogen;AN: 速效氮 Available nitrogen;IFI: 土壤肥力综合指标 The soil integrated fertility index; AP: 旱冬瓜人工林 A. nepalensis plantation; MP: 人工混交林 Mixed plantation; PP: 云南松人工林 P. yunnanensis Plantation; 8a: 种植8年的旱冬瓜林 8-years A. nepalensis plantation; 5a: 种植5年的旱冬瓜林 5-years A. nepalensis plantation; NAF: 旱冬瓜天然林 Natural A. nepalensis forest; NMF: 旱冬瓜-云南松混交天然林 Natural mixed forest; NPF: 云南松天然林 Natural P. yunnanensis forest; ATV: 旱冬瓜总材积 Total volume of A. nepalensis; 不同小写字母表示多重比较结果中两组数据在0.05水平上差异显著 |
旱冬瓜的种植模式、种植时间及天然林中旱冬瓜材积与林下植被的关系并非简单的线性关系(图 2)。旱冬瓜不同种植模式的比较, 发现旱冬瓜纯林和旱冬瓜+云南松混交林中草本植物地上干重显著大于云南松纯林(P < 0.05);草本多样性、灌木生长和灌木多样性则在三个模式之间无显著差异(P>0.05)(图 2)。草本生长、草本多样性及灌木多样性均与旱冬瓜材积无显著线性关系, 仅灌木生长与旱冬瓜材积呈显著正相关(图 2)。草本生长和多样性均在荒草地中最好, 并不随旱冬瓜种植时间的增加而变化, 与旱冬瓜材积也无显著线性关系(图 2);同时, 灌木生长在三个不同种植时间的林分中无显著差异, 而灌木多样性表现为林龄8a的旱冬瓜纯林显著高于林龄5a的林分(图 2)。

|
| 图 2 旱冬瓜对林下植被的影响 Fig. 2 The impacts of A. nepalensis on the understory vegetation ADMG: 草本地上部分干重 The above-ground dried mass of herb species; SGI: 灌木生长指数 The growth index of shrub; G-Shannon: 草本多样性指数 Shannon-Wiener index of herb species; S-Shannon: 灌木多样性指数 Shannon-Wiener index of shrub species; 不同小写字母表示多重比较结果中两组数据在0.05水平上差异显著 |
天然林中, 旱冬瓜对草本无显著影响(图 2);旱冬瓜林中灌木生长显著优于云南松林(P < 0.05), 且灌木生长与旱冬瓜材积呈显著正关系, 而混交林中的灌木多样性显著高于旱冬瓜林和云南松林(P < 0.05)(图 2)。在退耕地与天然林中, 灌木生长与OM、TN及土壤养分综合指标呈现显著的正相关(P < 0.05)(表 3)。
| 林型组别 Forest type |
林下植被 Understory vegetation |
有机质 OM |
总氮 TN |
速效氮 AN |
土壤肥力综合指标 IFI |
| 旱冬瓜混交配置 | 草本地上干重 | 0.25 | 0.29 | 0.26 | 0.29 |
| Species configuration | 草本多样性 | 0.30 | 0.37 | 0.25 | 0.37 |
| 灌木生长 | 0.46* | 0.42* | 0.28 | 0.58** | |
| 灌木多样性 | 0.02 | 0.04 | -0.03 | 0.20 | |
| 林分年龄 | 草本地上干重 | -0.07 | 0.01 | -0.21 | -0.05 |
| Stand age | 草本多样性 | -0.28 | -0.28 | -0.4 | -0.30 |
| 灌木生长 | 0.39 | 0.31 | 0.34 | 0.34 | |
| 灌木多样性 | 0.33 | 0.27 | 0.31 | 0.34 | |
| 群落优势种 | 草本地上干重 | 0.15 | 0.20 | -0.35 | 0.23 |
| Dominant species of community | 草本多样性 | -0.03 | -0.01 | 0.21 | -0.06 |
| 灌木生长 | 0.50* | 0.49* | 0.10 | 0.51* | |
| 灌木多样性 | 0.03 | 0.08 | 0.39 | 0.17 | |
| *: P < 0.05;**: P < 0.01;IFI:土壤肥力综合指标Integrated fertility index | |||||
旱冬瓜材积与OM、TN及IFI都有极显著的正相关关系(R=0.30, P=0.011;R=0.36, P=0.002;R=0.49, P < 0.001), 但与速效氮则没有显著相关关系(P>0.05) (图 1)。灌木生长与旱冬瓜材积、OM、TN及IFI呈显著的正相关关系(R=0.51, P < 0.001;R=0.23, P=0.049;R=0.27, P=0.024;R=0.27, P=0.024)(图 2, 表 3)。SEM模型中, 旱冬瓜材积与土壤养分综合指标和灌木生长均显著正相关(P < 0.001), 而草本地上生物量与旱冬瓜材积无显著相关关系, 各土壤养分指标和林下草本地上生物量及灌木生长状况间的关系均不显著(P>0.05)(图 3)。

|
| 图 3 旱冬瓜对林下植物的影响 Fig. 3 The response of understory vegetation growth to A. nepalensis-soil feedback SEM模型的参数为:Fisher′s C=1.46; P=0.23; AIC=29.46. AIC, akaike information criterion. 实线箭头表示显著的关系(P < 0.05); 黑色虚线表示无显著的关系(P>0.05); 路径系数为标准化后的系数; 用Shipley′s C检验各路径(P>0.05说明模型中无路径丢失, 拟合良好), 用AIC衡量模型拟合优良性 |
氮是土壤中的重要营养元素, 常常成为限制植物生长的土壤因子。以往的研究表明固氮树种能提升土壤氮含量, 增加土壤有机质, 同时改良土壤[7, 11-12, 42-43]。桤木属植物作为能与弗兰克氏菌共生形成根瘤的固氮树种能显著改善土壤[19, 44]。本研究亦发现, 旱冬瓜对退化土壤的改善作用较为显著: SEM模型分析表明, 旱冬瓜材积与土壤性质显著正相关, 其路径系数达0.47。此外, 旱冬瓜材积与土壤有机质、土壤总氮及土壤养分综合指标均有显著的正相关关系。
土壤肥力综合指数随旱冬瓜优势度增加而增大, 这说明旱冬瓜优势度的增加提升了退化土壤的修复效果。有研究表明, 林地中土壤总氮及土壤总有机碳也会随着固氮植物的优势度增加而增加[45-46], 当旱冬瓜优势度增大的时候, 土壤中氮素营养输入量与输出量间的顺差增大, 因而土壤获得的净氮素量增加。虽然土壤有机质与土壤总氮在天然林中随旱冬瓜优势度的增大而增加, 但是在退耕地的人工林中土壤有机质与土壤总氮并未随旱冬瓜种植比例的增加而呈现显著变化, 这有可能与立地条件有关。Orozco-Aceves等研究发现, 土壤基质会影响植物与土壤的反馈和植物对土壤的改良效果[47], 退耕地受施肥影响, 土壤养分性质与未受施肥影响的天然林有所不同, 这可能导致旱冬瓜与土壤间的长期反馈关系与天然林也有所不同。另外, 鉴于退耕地旱冬瓜的造林时间晚于天然林中旱冬瓜的定植时间, 这也可能导致旱冬瓜对土壤的影响呈现人工林不如天然林。作为对照的云南松天然林, 其土壤总氮显著小于退耕地对照的云南松人工林(P < 0.05), 即由于耕种过程中的施肥, 退耕地中土壤N含量可能高于天然林, 这可能是退耕地中旱冬瓜对土壤总氮影响相对较弱的原因之一。由于施肥耕种导致退耕地中的土壤总氮水平较高, 为旱冬瓜个体生长提供了充足的氮素环境, 因此降低了旱冬瓜获取土壤N的需求, 进而降低了旱冬瓜的生物固氮功能。如Boring等发现固氮植物的固氮效率与土壤中有效氮源负相关[48], 高有效氮源环境中固氮植物的固氮功能反而降低[5]。此外, 固氮植物提升土壤氮的同时也增加了土壤的碳固持[11], 因此, 退耕地中土壤有机质不随旱冬瓜优势度增加而增加, 而天然林中土壤有机质随旱冬瓜优势度增加而显著增加。虽然植物共生固氮需要土壤供应大量的磷元素[49], 但是退耕地土壤N∶P的比值显著低于天然林(P < 0.05), 因而土壤磷元素可能并非是退耕地中旱冬瓜改善土壤的限制因子。而这可能是由于林分发展到一定阶段后, 旱冬瓜根瘤固氮活动较低[27], 不足以对土壤速效氮造成显著影响。
林龄在桤木与土壤养分间的相互作用中也较为关键[26, 28, 50]。本研究发现8年生旱冬瓜林的土壤TN、OM及土壤养分肥力均显著大于5年生旱冬瓜林的这三个指标。在5个林龄段(7-56 a)旱冬瓜人工林中, 土壤TN和OC均与林龄呈显著正相关[28], 说明随旱冬瓜林龄增大, 土壤养分性状越优良。此外, AN随林龄增大而趋于增加, 可能是由于8年生的旱冬瓜其生理和生长对AN的需求量比5年生的旱冬瓜大, 需要增加土壤中的AN供给[50]。Sharma等发现, 旱冬瓜的年固氮量在5-15 a人工林阶段随林龄而增加, 而15-40 a的旱冬瓜林中随林龄增大逐渐降低[50], 特别是在低林龄阶段, 旱冬瓜的固氮活动十分活跃[27, 49]。
3.2 林下植被对旱冬瓜-土壤间反馈作用的响应林下植被能增加生态系统多样性及稳定性[24]且保土保肥[23], 同时土壤养分往往被看作是林下植物多样性[51]和生长状况[52-53]的重要解释因子。固氮树种可能会通过凋落物降解、根系分泌等途径向土壤中释放更多的营养物质[8], 提升土壤肥力, 增加土壤中植物可利用的营养, 促进其他植物生长[17, 22, 54], 因而作为固氮树种的旱冬瓜会影响林下植被的多样性, 促进林下植被生长。通过SEM模型分析发现, 林下灌木与旱冬瓜材积有显著的正相关关系(P < 0.05), 偏相关分析也表明, 灌木生长与旱冬瓜材积呈正显著相关关系(P < 0.05), 这可能是由于旱冬瓜增加了土壤中灌木生长所需的营养;同时, 土壤总氮、总有机质及肥力综合指标与灌木生长显著正相关(P < 0.05)进一步说明这一点。此外, 有研究发现, 占马地流域相对于耕地、荒草地及疏林地而言, 灌木林持水保肥效果最好[32], 而旱冬瓜对灌木的这一正影响, 会消减土壤侵蚀, 进一步促进旱冬瓜对土壤的改善。
旱冬瓜通过增加土壤氮素助益植物生长的现象在旱冬瓜不同优势度的林分和不同林龄的旱冬瓜林中均有呈现。退耕地中, 旱冬瓜材积与灌木生长显著正相关, 这可能是旱冬瓜改善了土壤肥力的结果。退耕地中土壤养分与灌木生长显著正相关, 正是灌木生长对旱冬瓜-土壤间关系的正反馈[54]。天然林中, 灌木生长随旱冬瓜优势度增加而增加, 这是由于天然林中旱冬瓜改善了土壤养分, 进而促进了灌木的生长。对草本而言, 旱冬瓜不同种植模式中, 云南松人工纯林中草本的地上生物量最小, 可能是旱冬瓜改变了土壤肥力所致[54]。
4 结论固氮树种旱冬瓜的种植显著提升了退化林地土壤总氮和总有机质, 显著改良了土壤肥力, 为其他植物的发展提供更优越的土壤环境。旱冬瓜对土壤的修复程度受其优势度和林龄的影响, 作为土壤氮素来源之一, 旱冬瓜优势度较大时, 林地中的土壤获得净氮素量趋于增加;在旱冬瓜幼龄林阶段, 随着林龄增大, 土壤得到了改良和修复, 趋于正向生态演替。旱冬瓜改善土壤环境的作用为林下植被生长提供更丰富的养分, 促进了林下灌木的生长, 对金沙江上游退化土壤修复和林下植被重建具有生态演替的驱动效应。
致谢: 感谢云南省林业科学研究院孟广涛和李品荣对本研究的帮助, 感谢姜健和刘刚对野外样地调查中给予的帮助。| [1] |
张桃林, 王兴祥. 土壤退化研究的进展与趋向. 自然资源学报, 2000, 15(3): 280-284. DOI:10.3321/j.issn:1000-3037.2000.03.016 |
| [2] |
陈小雪, 李红丽, 董智, 陈鹏, 仇苏倩, 高娅. 不同造林模式下火烧迹地的土壤修复效果及植物多样性变化. 水土保持学报, 2019, 033(003): 332-339. |
| [3] |
Sprent J I, Parsons R. Nitrogen fixation in legume and non-legume trees. Field Crops Research, 2000, 65(2/3): 183-196. |
| [4] |
Binkley D, Giardina C. Why do tree species affect soils? The warp and woof of tree-soil interactions. Biogeochemistry, 1998, 42(1/2): 89-106. DOI:10.1023/A:1005948126251 |
| [5] |
Van Breemen N, Finzi A C. Plant-soil interactions: ecological aspects and evolutionary implications. Biogeochemistry, 1998, 42(1/2): 1-19. DOI:10.1023/A:1005962124317 |
| [6] |
Spears J D H, Lajtha K, Caldwell B A, Pennington S B, Vanderbilt K. Species effects of Ceanothus velutinus versus Pseudotsuga menziesii, Douglas-fir, on soil phosphorus and nitrogen properties in the Oregon cascades. Forest Ecology and Management, 2001, 149(1/3): 205-216. |
| [7] |
Chen X P, Yang J N, Zhu X E, Liang X, Lei Y R, He C Q. N-fixing trees in wetland restoration plantings: effects on nitrogen supply and soil microbial communities. Environmental Science and Pollution Research, 2016, 23(24): 24749-24757. DOI:10.1007/s11356-016-7454-y |
| [8] |
Forrester D I, Bauhus J, Cowie A L, Vanclay J K. Mixed-species plantations of Eucalyptus with nitrogen-fixing trees: a review. Forest Ecology and Management, 2006, 233(2/3): 211-230. |
| [9] |
Chaer G M, Resende A S, Campello E F C, De Faria S M, Boddey R M. Nitrogen-fixing legume tree species for the reclamation of severely degraded lands in Brazil. Tree Physiology, 2011, 31(2): 139-149. DOI:10.1093/treephys/tpq116 |
| [10] |
Hoogmoed M, Cunningham S C, Baker P, Beringer J, Cavagnaro T R. N-fixing trees in restoration plantings: effects on nitrogen supply and soil microbial communities. Soil Biology and Biochemistry, 2014, 77: 203-212. DOI:10.1016/j.soilbio.2014.06.008 |
| [11] |
Johnson D W, Curtis P S. Effects of forest management on soil C and N storage: meta-analysis. Forest Ecology and Management, 2001, 140(2/3): 227-238. |
| [12] |
Wang F M, Li Z A, Xia H P, Zou B, Li N Y, Liu J, Zhu W X. Effects of nitrogen-fixing and non-nitrogen-fixing tree species on soil properties and nitrogen transformation during forest restoration in southern China. Soil Science and Plant Nutrition, 2010, 56(2): 297-306. DOI:10.1111/j.1747-0765.2010.00454.x |
| [13] |
Paynel F, Murray P J, Cliquet J B. Root exudates: a pathway for short-term N transfer from clover and ryegrass. Plant and Soil, 2001, 229(2): 235-243. DOI:10.1023/A:1004877214831 |
| [14] |
Høgh-Jensen H, Schjoerring J K. Below-ground nitrogen transfer between different grassland species: direct quantification by 15N leaf feeding compared with indirect dilution of soil 15N. Plant and Soil, 2000, 227(1/2): 171-183. DOI:10.1023/A:1026535401773 |
| [15] |
Bellingham P J, Walker L R, Wardle D A. Differential facilitation by a nitrogen-fixing shrub during primary succession influences relative performance of canopy tree species. Journal of Ecology, 2001, 89(5): 861-875. DOI:10.1046/j.0022-0477.2001.00604.x |
| [16] |
Png G K, Lambers H, Kardol P, Turner B L, Wardle D A, Laliberté E. Biotic and abiotic plant-soil feedback depends on nitrogen-acquisition strategy and shifts during long-term ecosystem development. Journal of Ecology, 2019, 107(1): 142-153. DOI:10.1111/1365-2745.13048 |
| [17] |
Thomas B D, Bowman W D. Influence of N2-fixing Trifolium on plant species composition and biomass production in alpine tundra. Oecologia, 1998, 115(1/2): 26-31. |
| [18] |
Titus J H, Bishop J G. Propagule limitation and competition with nitrogen fixers limit conifer colonization during primary succession. Journal of Vegetation Science, 2014, 25(4): 990-1003. DOI:10.1111/jvs.12155 |
| [19] |
Tarrant R F, Trappe J M. The role of Alnus in improving the forest environment. Plant and Soil, 1971, 35(1): 335-348. DOI:10.1007/BF02661862 |
| [20] |
Stewart W D P. A quantitative study of fixation and transfer of nitrogen in Alnus. Journal of Experimental Botany, 1962, 13(2): 250-256. DOI:10.1093/jxb/13.2.250 |
| [21] |
Binkley D, Sollins P, Bell R, Sachs D, Myrold D. Biogeochemistry of adjacent conifer and alder-conifer stands. Ecology, 1992, 73(6): 2022-2033. DOI:10.2307/1941452 |
| [22] |
Delver P, Post A. Influence of alder hedges on the nitrogen nutrition of apple trees. Plant and Soil, 1968, 28(2): 325-336. DOI:10.1007/BF01880250 |
| [23] |
Nearing M A, Jetten V, Baffaut C, Cerdan O, Couturier A, Hernandez M, Bissonnais Y L, Nichols M H, Nunes J P, Renschler C S, Souchère V, van Oost K. Modeling response of soil erosion and runoff to changes in precipitation and cover. Catena, 2005, 61(2/3): 131-154. |
| [24] |
Lehman C L, Tilman D. Biodiversity, stability, and productivity in competitive communities. The American Naturalist, 2000, 156(5): 534-552. DOI:10.1086/303402 |
| [25] |
Gilliam F S. The ecological significance of the herbaceous layer in temperate forest ecosystems. Bioscience, 2007, 57(10): 845-858. DOI:10.1641/B571007 |
| [26] |
Sharma E, Ambasht R S. Litterfall, decomposition and nutrient release in an age sequence of Alnus nepalensis plantation stands in the eastern Himalaya. Journal of Ecology, 1987, 75(4): 997-1010. DOI:10.2307/2260309 |
| [27] |
Sharma E, Ambasht R S. Seasonal variation in nitrogen fixation by different ages of root nodules of Alnus nepalensis plantations, in the eastern Himalayas. Journal of Applied Ecology, 1984, 21(1): 265-270. DOI:10.2307/2403052 |
| [28] |
Sharma E, Ambasht R S, Sing M P. Chemical soil properties under five age series of Alnus nepalensis plantations in the Eastern Himalayas. Plant and Soil, 1985, 84(1): 105-113. DOI:10.1007/BF02197871 |
| [29] |
Liu S R, Wu S R, Wang H. Managing planted forests for multiple uses under a changing environment in China. New Zealand Journal of Forestry Science, 2014, 44(S1): S3. |
| [30] |
陈宏伟, 李大伟, 史富强, 杨斌, 陈伟, 李江, 孟梦, 陈绍安. 旱冬瓜用材林优树选择标准的研究. 西部林业科学, 2010, 39(1): 6-10. DOI:10.3969/j.issn.1672-8246.2010.01.002 |
| [31] |
Kayang H. Fungal and bacterial enzyme activities in Alnus nepalensis D. Don. European Journal of Soil Biology, 2001, 37(3): 175-180. DOI:10.1016/S1164-5563(01)01082-2 |
| [32] |
方向京, 李贵祥, 孟广涛, 柴勇, 和丽萍, 张正海, 李宁云. 珠江源头区占马地小流域不同用地水土保持作用研究. 中国水土保持科学, 2007, 5(4): 22-26. DOI:10.3969/j.issn.1672-3007.2007.04.005 |
| [33] |
李贵祥, 孟广涛, 方向京, 和丽萍, 张正海, 柴勇. 珠江源头区占马地小流域植被优化重建模式及效益研究. 水土保持研究, 2007, 14(5): 384-386. |
| [34] |
赵芳. 飞播马尾松林林下植被特征及其影响因子研究[D]. 南昌: 江西农业大学, 2015.
|
| [35] |
Nelson D W, Sommers L E. Total carbon, organic carbon, and organic matter//Sparks D L. Methods of Soil Analysis. Madison: SSSA and ASA, 1996: 961-1010.
|
| [36] |
栾慧, 刘红, 高一娜, 张立群, 王美玉, 郭维士. 土壤速效氮测定方法的改进. 湖南农业科学, 2017, 000(003): 88-89, 93. |
| [37] |
鲍士旦. 土壤农化分析. 北京: 中国农业出版社, 2000: 138.
|
| [38] |
吕晓男, 陆允甫, 王人潮. 土壤肥力综合评价初步研究. 浙江大学学报(农业与生命科学版), 1999(4): 38-42. |
| [39] |
赵川, 和丽萍, 李贵祥, 柴勇, 邵金平. 昆阳磷矿废弃地植被恢复对土壤质量的影响及评价. 西部林业科学, 2018, 47(2): 106-111. |
| [40] |
肖永林. 编制云南省一元立木材积表技术总结. 云南林业调查规划, 1981(4): 1-7. |
| [41] |
Giese L A B, Aust W M, Kolka R K, Trettin C C. Biomass and carbon pools of disturbed riparian forests. Forest Ecology and Management, 2003, 180(1/3): 493-508. |
| [42] |
Cech P G, Venterink H O, Edwards P J. N and P cycling in Tanzanian humid savanna: influence of herbivores, fire, and N2-fixation. Ecosystems, 2010, 13(7): 1079-1096. DOI:10.1007/s10021-010-9375-9 |
| [43] |
Macedo M O, Resende A S, Garcia P C, Boddey R M, Jantalia C P, Urquiaga S, Campello E F C, Franco A A. Changes in soil C and N stocks and nutrient dynamics 13 years after recovery of degraded land using leguminous nitrogen-fixing trees. Forest Ecology and Management, 2008, 255(5/6): 1516-1524. |
| [44] |
Sharma R, Sharma E, Purohit A N. Cardamom, mandarin and nitrogen-fixing trees in agroforestry systems in India's Himalayan region. Ⅱ. Soil nutrient dynamics. Agroforestry Systems, 1996, 35(3): 255-268. DOI:10.1007/BF00044457 |
| [45] |
Sitters J, Edwards P J, Venterink H O. Increases of soil C, N, and P pools along an Acacia tree density gradient and their effects on trees and grasses. Ecosystems, 2013, 16(2): 347-357. DOI:10.1007/s10021-012-9621-4 |
| [46] |
Mitchell J S, Ruess R W. N2 fixing alder (Alnus viridis spp. fruticosa) effects on soil properties across a secondary successional chronosequence in interior Alaska. Biogeochemistry, 2009, 95(2/3): 215-229. |
| [47] |
Orozco-Aceves M, Standish R J, Tibbett M. Soil conditioning and plant-soil feedbacks in a modified forest ecosystem are soil-context dependent. Plant and Soil, 2015, 390(1/2): 183-194. |
| [48] |
Boring L R, Swank W T, Waide J B, Henderson G S. Sources, fates, and impacts of nitrogen inputs to terrestrial ecosystems: review and synthesis. Biogeochemistry, 1988, 6(2): 119-159. |
| [49] |
Vitousek P M, Cassman K, Cleveland C, Crews T, Field C B, Grimm N B, Howarth R W, Marino R, Martinelli L, Rastetter E B, Sprent J I. Towards an ecological understanding of biological nitrogen fixation. Biogeochemistry, 2002, 57-58(1): 1-45. |
| [50] |
Sharma G, Sharma R, Sharma E, Singh K K. Performance of an age series of Alnus-Cardamom plantations in the Sikkim Himalaya: nutrient dynamics. Annals of Botany, 2002, 89(3): 273-282. DOI:10.1093/aob/mcf036 |
| [51] |
Barbier S, Gosselin F, Balandier P. Influence of tree species on understory vegetation diversity and mechanisms involved-A critical review for temperate and boreal forests. Forest Ecology and Management, 2008, 254(1): 1-15. DOI:10.1016/j.foreco.2007.09.038 |
| [52] |
McConnaughay K D M, Coleman J S. Biomass allocation in plants: ontogeny or optimality? A test along three resource gradients. Ecology, 1999, 80(8): 2581-2593. DOI:10.1890/0012-9658(1999)080[2581:BAIPOO]2.0.CO;2 |
| [53] |
Raich J W. Aboveground productivity and soil respiration in three Hawaiian rain forests. Forest Ecology and Management, 1998, 107(1/3): 309-318. |
| [54] |
Cech P G, Kuster T, Edwards P J, Venterink H O. Effects of herbivory, Fire and N2-fixation on nutrient limitation in a humid African savanna. Ecosystems, 2008, 11(6): 991-1004. DOI:10.1007/s10021-008-9175-7 |