 2022, Vol. 42
2022, Vol. 42文章信息
- 赵阳, 曹家豪, 李波, 齐瑞, 刘婷, 陈学龙, 高本强, 赵艳丽, 刘锦乾, 曹秀文
- ZHAO Yang, CAO Jiahao, LI Bo, QI Rui, LIU Ting, CHEN Xuelong, GAO Benqiang, ZHAO Yanli, LIU Jinqian, CAO Xiuwen
- 洮河自然保护区大峪沟林区紫果云杉群落木本植物种群生态位特征
- Niche of woody plant populations of Picea purpurea community in Dayugou forest area, Taohe Nature Reserve, Gansu Province
- 生态学报. 2022, 42(5): 1865-1875
- Acta Ecologica Sinica. 2022, 42(5): 1865-1875
- http://dx.doi.org/10.5846/stxb202102070395
-
文章历史
- 收稿日期: 2021-02-07
- 网络出版日期: 2021-11-17





2. 甘肃白龙江森林生态系统定位观测研究站, 舟曲 746300
2. GanSu Bailongjiang National Forest Ecosystem Research Station, Zhouqu 746300, China
生态位(niche)概念自被Grinnell[1]引入生态学研究领域以来, 有关其定义和内涵一直存在诸多争议, 截至目前尚未形成统一共识[2-3], 因此生态位研究一直是生态学的重要课题。100多年来, 生态位理论在不断丰富和发展中已成为解释自然群落中物种共存与竞争机制的基本理论[4-5], 被应用到种群、群落以及生态系统等复杂组织水平上。研究森林群落优势种群生态位能客观地反映种群对资源的利用能力及其在群落或生态系统中的功能和地位[6-7], 还可以量化物种之间、物种与环境之间的相互关系[8], 反映群落结构与功能、生物多样性、物种进化以及群落演替的方向等[9-10]。这对群落生物多样性保护, 揭示种群生存策略和机制[11]及预测群落发展趋势具有重要意义。
洮河自然保护区位于洮河上游, 地处青藏高原东部边缘的甘南山区与黄土高原过渡区, 属青藏高原湿润气候与高原大陆性季风气候交汇带[12]。独特的地理环境和气候条件孕育出丰富多样的生态类型, 作为黄河的一级支流, 洮河上游的高原湿地、草原、森林等共同构成生态屏障维持着洮河乃至黄河上游区域的生态平衡, 森林作为最顶级的生态类型, 发挥着第一屏障作用。以紫果云杉(Picea purpurea)为主要建群种的原始林群落在本区域中高海拔有着大面积分布, 经过长期的适应与自然选择已形成完整的生态系统结构, 在物种多样性保护、局域生态安全维护等方面发挥着极为重要的作用。截止目前, 本区域有关紫果云杉的研究多存在于种群结构[12-13]和育苗等方面, 但其群落组成与结构如何, 生存状态如何, 种间竞争与共存关系如何, 相关研究尚不多见。为此, 本研究以紫果云杉群落为对象, 通过分析木本植物种群生态位特征, 研究群落组成与结构、物种多样性、种间关系以及物种与群落环境的关系, 揭示种群对资源和空间的利用能力、群落生存状态与发展趋势, 为种群和群落的保护提供理论依据。
1 研究区与研究方法 1.1 研究区概况研究区位于洮河国家级自然保护区大峪沟林区。大峪沟林区位于甘肃省甘南州卓尼县木耳镇大峪沟(103°33′-104°02′E, 34°10′-34°52′N), 面积约10.5万hm2, 林区湿润多雨, 光照充足, 四季不明, 小气候多样。海拔2500 m-4920 m, 相对高差大, 为典型的高原山地地貌。年均日照2266 h, 年均温2.3℃, 无霜期105 d, 年均降水640 mm, 河流纵横, 水资源丰富, 土壤主要为棕、褐色森林土[14]。植被类型多样, 阳坡主要为灌丛、高山草甸等, 阴坡、河谷地带主要是以云杉(Picea asperata)、冷杉(Abies fabri)等为建群种的原始暗针叶林群落, 森林结构相对完整, 覆盖率50.74%, 林下天然更新良好[15]。主要树种有紫果云杉、岷江冷杉(Abies faxoniana)、桦木(Betula)、柏木(Cupressus)等, 森林质量总体较高, 野生动植物资源丰富。
1.2 样地设置2020年7-8月, 在对大峪沟林区进行了全面踏查的基础上, 采用典型抽样原则设置50 m×50 m的紫果云杉样地8块(表 1), 逐一测量并记录样地内胸径大于3 cm的全部乔木树种的胸径、树高、冠幅, 乔木层郁闭度等指标。并在每个样地设置3个10 m×10 m的灌木样方, 记录各样方内的灌木种类、数量、地径、高度、盖度。各样地用GPS定位, 记录海拔、坡度、坡向等立地因子。
| 样地号 No. |
海拔 Elevation/m |
坡向 Direction |
坡位 Slope position |
密度 Density /(株/hm2) |
平均胸径 Mean DBH/cm |
平均树高 Average height/m |
郁闭度 Canopy density/% |
灌层盖度 Coverage /% |
最大树龄 Most old/a |
| 1 | 2915 | 半阴坡 | 下 | 925 | 18.87±0.81 | 13.64 | 63.60 | 82.50 | 116 |
| 2 | 2880 | 半阴坡 | 下 | 1275 | 19.13±1.33 | 12.43 | 75.35 | 64.25 | 106 |
| 3 | 3100 | 半阴坡 | 中下 | 1509 | 12.08±0.56 | 10.07 | 65.60 | 82.30 | 98 |
| 4 | 2955 | 半阴坡 | 下 | 2210 | 14.88±1.23 | 10.96 | 75.65 | 78.55 | 110 |
| 5 | 3028 | 阴坡 | 中下 | 2153 | 15.95±1.50 | 11.80 | 70.50 | 68.50 | 105 |
| 6 | 2961 | 阴坡 | 下 | 2325 | 14.38±1.02 | 10.39 | 68.50 | 82.65 | 166 |
| 7 | 3024 | 阴坡 | 下 | 1973 | 21.25±3.60 | 15.70 | 68.85 | 70.25 | 143 |
| 8 | 3041 | 阴坡 | 下 | 2125 | 21.48±3.34 | 15.89 | 71.50 | 72.60 | 110 |
重要值取物种在每个样地的平均值, 计算方法参见文献[16-17], 频数为该物种出现的样地、样方数。
1.3.2 生态位以样地代表多种资源的综合状态[16], 用Levins指数BL[17]和Shannon指数BS[6]计算生态位宽度:
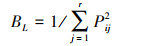
|
(1) |
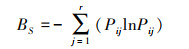
|
(2) |
式中, Bi为第i物种的生态位宽度, Pij为第i物种在第j个资源水平上的重要值占该物种在所有资源水平上重要值总和的比例, Pij = nij/∑nij, nij为物种i在第j资源位上的重要值, r为样方数。
生态位总宽度[18]B:
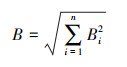
|
(3) |
式中, Bi为物种i的生态位宽度, n为物种数。
生态位相似[19]Cik:
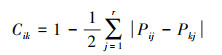
|
(4) |
式中, Cik为物种i和k的生态位相似性, Cik∈[0, 1], Pij和Pkj分别为物种i和k在资源位j上的重要值。
生态位重叠[20]Oik:
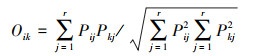
|
(5) |
式中, Oik为物种i和物种k的生态位重叠指数, 当Oik = 1时, 物种i和k在所有资源位中分布完全相同, 即物种i与k生态位完全重叠; 当Oik = 0时, 两个物种无共同资源状态, 生态位完全不重叠。
采用Excel2010和SPSS21.0软件对数据进行统计、计算和分析。
2 结果与分析 2.1 物种组成与重要值特征紫果云杉群落乔木层共有树种8种(表 2), 隶属于4科6属, 重要值大小为紫果云杉>岷江冷杉>祁连圆柏>红桦>糙皮桦>秦林柳>云杉>刺柏, 紫果云杉重要值远大于其它树种, 为该群落的建群种, 岷江冷杉为次优势种。灌木层共有植物50种, 隶属于16科25属(表 3), 重要值大于2的有22种, 其中最大的为峨眉蔷薇(16.690), 其次为唐古特忍冬(14.103)、蓝靛果(12.174)和葱皮忍冬(10.567)等。乔木树种在灌木层均有幼苗存在, 其中紫果云杉与岷江冷杉的幼苗重要值较大, 分别为4.872和4.912。总体来看, 灌木层蔷薇科物种最多(14种), 其重要值总和为45.32, 小于忍冬科(11种)的61.64, 表明忍冬科为灌木层第一优势科, 其次为蔷薇科, 茶藨子科(也叫醋栗科)(6种)以9.20的重要值位居第3。
| 编号 No. |
种名 Species |
重要值 Importance value/% |
变异系数 Coefficient variation |
编号 No. |
种名 Species |
重要值 Importance value/% |
变异系数 Coefficient variation |
|
| 1 | 紫果云杉Picea purpurea | 36.929 | 0.245 | 5 | 糙皮桦Betula utilis | 6.641 | 1.406 | |
| 2 | 岷江冷杉Abies faxoniana | 20.050 | 0.735 | 6 | 秦岭柳Salix qinlingica | 3.909 | 1.335 | |
| 3 | 祁连圆柏Sabina przewalskii | 14.417 | 0.616 | 7 | 云杉Picea asperata | 2.325 | 1.879 | |
| 4 | 红桦Betula albo-sinensis | 13.957 | 0.630 | 8 | 刺柏Juniperus formosana | 1.770 | 1.823 |
| 编号 No. |
种名 Species |
重要值 Importance value/% |
变异系数 Coefficient variation |
编号 No. |
种名 Species |
重要值 Importance value/% |
变异系数 Coefficient variation |
|
| 1 | 峨眉蔷薇Rosa omeiensis | 16.690 | 0.999 | 26 | 毛花忍冬Lonicera trichosantha | 1.700 | 0.983 | |
| 2 | 唐古特忍冬Lonicera tangutica | 14.103 | 0.577 | 27 | 陇塞忍冬Lonicera tangutica | 1.361 | 0.866 | |
| 3 | 蓝靛果Lonicera caerulea | 12.174 | 0.497 | 28 | 花楸Sorbus pohuashanensis | 1.338 | 0.866 | |
| 4 | 葱皮忍冬Lonicera ferdinandii | 10.567 | 0.480 | 29 | 刚毛忍冬Lonicera hispida | 0.846 | 0.990 | |
| 5 | 银露梅Potentilla glabra | 8.535 | 0.726 | 30 | 华西箭竹Fargesia nitida | 0.833 | 0.935 | |
| 6 | 华西忍冬Lonicera webbiana | 7.880 | 0.654 | 31 | 云杉Picea asperata | 0.709 | 1.738 | |
| 7 | 六道木Abelia biflora | 6.769 | 0.810 | 32 | 华西蔷薇Rosa moyesii | 0.668 | 1.047 | |
| 8 | 扁刺蔷薇Rosa sweginzowii | 5.134 | 0.801 | 33 | 平枝栒子Cotoneaster horizontalis | 0.599 | 0.893 | |
| 9 | 岷江冷杉Abies faxoniana | 4.912 | 1.028 | 34 | 宝兴茶藨子Ribes moupinense | 0.530 | 1.000 | |
| 10 | 紫果云杉Picea purpurea | 4.872 | 0.418 | 35 | 四川忍冬Lonicera szechuanica | 0.508 | 0.869 | |
| 11 | 祁连圆柏Sabina przewalskii | 4.031 | 0.843 | 36 | 杜鹃Rhododendron simsii | 0.430 | 1.000 | |
| 12 | 桦木Betula | 3.895 | 0.832 | 37 | 刺柏Juniperus formosana | 0.344 | 2.646 | |
| 13 | 陕甘花楸Sorbus koehneana | 3.640 | 0.608 | 38 | 虎榛子Ostryopsis davidiana | 0.335 | 0.935 | |
| 14 | 欧亚绣线菊Spiraea media | 3.591 | 0.991 | 39 | 灰栒子Cotoneaster acutifolius | 0.296 | 1.000 | |
| 15 | 冰川茶藨子Ribes glaciale | 3.384 | 0.767 | 40 | 卫矛Euonymus alatus | 0.293 | 0.935 | |
| 16 | 红脉忍冬Lonicera nervosa | 3.076 | 0.751 | 41 | 瘤糖茶藨子Ribes himalense | 0.293 | 1.000 | |
| 17 | 糖茶藨子Ribes himalense | 2.915 | 0.866 | 42 | 金露梅Potentilla fruticosa | 0.261 | 0.935 | |
| 18 | 甘肃小檗Berberis kansuensis | 2.780 | 0.850 | 43 | 白毛山梅花Philadelphus schrenkii | 0.258 | 1.000 | |
| 19 | 忍冬Lonicera japonica | 2.655 | 1.235 | 44 | 臭樱Maddenia hypoleuca | 0.240 | 0.935 | |
| 20 | 华北绣线菊Spiraea fritschiana | 2.422 | 0.827 | 45 | 胡枝子Lespedeza bicolor | 0.216 | 1.000 | |
| 21 | 栓翅卫矛Euonymus phellomanus | 2.192 | 1.327 | 46 | 刺五加Acanthopanax senticosus | 0.205 | 0.866 | |
| 22 | 秦岭柳Salix alfredi | 2.152 | 0.922 | 47 | 瑞香Daphne odora | 0.143 | 1.000 | |
| 23 | 细枝绣线菊Spiraea myrtilloides | 1.879 | 0.880 | 48 | 山茱萸Cornus officinalis | 0.057 | 0.935 | |
| 24 | 黑水柳Salix heishuiensis | 1.835 | 0.816 | 49 | 五加Acanthopanax gracilistylus | 0.050 | 1.000 | |
| 25 | 茶藨子Ribes nigrum | 1.826 | 0.761 | 50 | 悬钩子Rubus corchorifolius | 0.032 | 1.000 |
如表 4所示, 乔木层重要值最大的紫果云杉其生态位宽度(BL、BS)也最大, 为7.548和2.050, 其次, 红桦、祁连圆柏和岷江冷杉三者生态位宽度差异不大, 云杉和刺柏生态位宽度最小。前4种优势树种BL和BS生态位宽度大小均为紫果云杉>祁连圆柏>红桦>岷江冷杉, 而后4种弱势种BL和BS生态位宽度大小出现差异, 可见不同物种Levins指数和Shannon指数大小变化不同, 但总体趋势一致。
| 编号 No. |
种名 Species |
频数 Frequency |
生态位宽度 Niche breadth |
编号 No. |
种名 Species |
频数 Frequency |
生态位宽度 Niche breadth |
|||
| BL | Bs | BL | BS | |||||||
| 1 | 紫果云杉Picea purpurea | 8 | 7.548 | 2.050 | 5 | 糙皮桦Betula utilis | 4 | 2.687 | 1.173 | |
| 2 | 岷江冷杉Abies faxoniana | 7 | 5.695 | 1.761 | 6 | 秦岭柳Salix qinlingica | 4 | 2.876 | 1.078 | |
| 3 | 祁连圆柏Sabina przewalskii | 8 | 5.801 | 1.930 | 7 | 云杉Picea asperata | 1 | 1.766 | 0.625 | |
| 4 | 红桦Betula albo-sinensis | 7 | 5.727 | 1.835 | 8 | 刺柏Juniperus formosana | 1 | 1.851 | 0.699 | |
灌木层生态位宽度(BL、BS)最大的为紫果云杉幼苗(6.807、1.979)(表 5), 其次为忍冬科的蓝靛果(6.414、1.958)、葱皮忍冬(6.249、1.885)、唐古特忍冬(6.002、1.892)和华西忍冬(5.601、1.860), 蔷薇科生态位宽度最大的3个种为陕甘花楸(5.468、1.787)、银露梅(4.795、1.717)和扁刺蔷薇(4.402、1.701), 这8个种除扁刺蔷薇BS外, 生态位宽度均大于重要值最大的峨眉蔷薇(4.005、1.707)。茶藨子科生态位宽度最大的3个种为冰川茶藨子(4.133、1.525)、茶藨子(3.458、1.291)和糖茶藨子(3.123、1.340)。在3个优势科中, 忍冬科11种植物的生态位总宽度(13.654、4.466)大于蔷薇科14种植物(11.183、4.079)及茶藨子科6种植物的生态位总宽度(6.464、2.406), 再次表明忍冬科为灌木层第一优势科。此外, 桦木(包括红桦和糙皮桦)(4.728、1.662)、祁连圆柏(4.675、1.625)及岷江冷杉(3.891、1.489)的幼苗也有较大的生态位宽度。生态位宽度BL和BS最小值为1和0的有16种, 如虎榛子、臭樱等, 表示该物种在所有样方中仅出现1次, 为该群落的偶见种。
| 编号 No. |
种名 Species |
频数 Frequency |
生态位宽度 Niche breadth |
编号 No. |
种名 Species |
频数 Frequency |
生态位宽度 Niche breadth |
|||
| BL | BS | BL | BS | |||||||
| 1 | 峨眉蔷薇Rosa omeiensis | 24 | 4.005 | 1.707 | 26 | 毛花忍冬Lonicera trichosantha | 2 | 1.571 | 0.674 | |
| 2 | 唐古特忍冬Lonicera tangutica | 24 | 6.002 | 1.892 | 27 | 陇塞忍冬Lonicera tangutica | 2 | 1.999 | 0.693 | |
| 3 | 蓝靛果Lonicera caerulea | 24 | 6.414 | 1.958 | 28 | 花楸Sorbus pohuashanensis | 2 | 1.996 | 0.692 | |
| 4 | 葱皮忍冬Lonicera ferdinandii | 23 | 6.249 | 1.885 | 29 | 刚毛忍冬Lonicera hispida | 3 | 1.541 | 0.606 | |
| 5 | 银露梅Potentilla glabra | 21 | 4.795 | 1.711 | 30 | 华西箭竹Fargesia nitida | 1 | 1.000 | 0.000 | |
| 6 | 华西忍冬Lonicera webbiana | 24 | 5.601 | 1.860 | 31 | 云杉Picea asperata | 2 | 1.989 | 0.690 | |
| 7 | 六道木Abelia biflora | 15 | 3.446 | 1.349 | 32 | 华西蔷薇Rosa moyesii | 2 | 1.331 | 0.415 | |
| 8 | 扁刺蔷薇Rosa sweginzowii | 22 | 4.402 | 1.701 | 33 | 平枝栒子Cotoneaster horizontalis | 2 | 1.680 | 0.595 | |
| 9 | 岷江冷杉Abies faxoniana | 20 | 3.891 | 1.489 | 34 | 宝兴茶藨子Ribes moupinense | 1 | 1.000 | 0.000 | |
| 10 | 紫果云杉Picea purpurea | 24 | 6.808 | 1.979 | 35 | 四川忍冬Lonicera szechuanica | 2 | 1.965 | 0.684 | |
| 11 | 祁连圆柏Sabina przewalskii | 21 | 4.675 | 1.625 | 36 | 杜鹃Rhododendron simsii | 1 | 1.000 | 0.000 | |
| 12 | 桦木Betula | 18 | 4.728 | 1.662 | 37 | 刺柏Juniperus formosana | 1 | 1.000 | 0.000 | |
| 13 | 陕甘花楸Sorbus koehneana | 21 | 5.468 | 1.787 | 38 | 虎榛子Ostryopsis davidiana | 1 | 1.000 | 0.000 | |
| 14 | 欧亚绣线菊Spiraea media | 9 | 2.535 | 0.997 | 39 | 灰栒子Cotoneaster acutifolius | 1 | 1.000 | 0.000 | |
| 15 | 冰川茶藨子Ribes glaciale | 18 | 4.133 | 1.525 | 40 | 卫矛Euonymus alatus | 1 | 1.000 | 0.000 | |
| 16 | 红脉忍冬Lonicera nervosa | 12 | 3.542 | 1.315 | 41 | 瘤糖茶藨子Ribes himalense | 1 | 1.000 | 0.000 | |
| 17 | 糖茶藨子Ribes himalense | 12 | 3.123 | 1.340 | 42 | 金露梅Potentilla fruticosa | 1 | 1.000 | 0.000 | |
| 18 | 甘肃小檗Berberis kansuensis | 12 | 3.212 | 1.334 | 43 | 白毛山梅花Philadelphus schrenkii | 1 | 1.000 | 0.000 | |
| 19 | 忍冬Lonicera japonica | 3 | 1.311 | 0.451 | 44 | 臭樱Maddenia hypoleuca | 1 | 1.000 | 0.000 | |
| 20 | 华北绣线菊Spiraea fritschiana | 6 | 3.799 | 1.517 | 45 | 胡枝子Lespedeza bicolor | 1 | 1.000 | 0.000 | |
| 21 | 栓翅卫矛Euonymus phellomanus | 5 | 2.440 | 1.163 | 46 | 刺五加Acanthopanax senticosus | 2 | 1.996 | 0.692 | |
| 22 | 秦岭柳Salix alfredi | 3 | 2.842 | 1.069 | 47 | 瑞香Daphne odora | 1 | 1.000 | 0.000 | |
| 23 | 细枝绣线菊Spiraea myrtilloides | 2 | 1.823 | 0.644 | 48 | 山茱萸Cornus officinalis | 1 | 1.000 | 0.000 | |
| 24 | 黑水柳Salix heishuiensis | 3 | 2.708 | 1.042 | 49 | 五加Acanthopanax gracilistylus | 1 | 1.000 | 0.000 | |
| 25 | 茶藨子Ribes nigrum | 4 | 3.458 | 1.291 | 50 | 悬钩子Rubus corchorifolius | 1 | 1.000 | 0.000 | |
综上可知, 重要值大的物种生态位宽度往往较大, 但也存在一定差异。如在乔木层, 岷江冷杉重要值大于祁连圆柏, 但生态位宽度却小于祁连圆柏。灌木层中峨眉蔷薇重要值最大, 但生态位宽度却仅为第12, 紫果云杉、岷江冷杉的幼苗重要值分别为第10和第9, 但生态位宽度却为第1和第10。结合表 2、3和4、5可以看出, 生态位宽度除与重要值有关外, 还与物种频数及重要值变异系数有关, 即物种频数越大、重要值变异系数越小, 其生态位宽度越大。
2.3 生态位相似紫果云杉群落8种乔木树种共构成28个种对(表 6), 生态位相似平均值为0.390, 相似性大于0.6的有7对, 占总对数的25.0%, 前4个优势种之间生态位相似平均值大于后4个弱势种。相似性最大的种对为岷江冷杉与红桦(0.722), 其次为紫果云杉与祁连圆柏(0.717)、紫果云杉与红桦(0.715), 弱势种秦岭柳与云杉、糙皮桦与刺柏的相似性也在0.6以上。相似性最低的为岷江冷杉与刺柏(0.073), 其次为岷江冷杉与云杉、秦岭柳, 红桦与云杉等。
灌木层50种植物共构成1225个种对, 生态位相似平均值为0.217, 总体相似性较低。其中相似性小于0.6的有1141对, 占总对数的93.14%, 绝大多数种对相似性介于0-0.4之间, 包括相似性为0的365对。相似性大于0.6的有84对, 仅占6.86%, 其中0.6-0.8的59对, 0.8-0.9的7对, 0.9-1.0的2对, 相似性为1的有16对。相似性大的如刚毛忍冬和毛花忍冬(0.911)、云杉与花楸(0.978)等以及相似性为1的种对多出现在非优势种之间。前22个优势或相对优势种构成的231个种对(表 7)生态位相似平均值为0.452, 相似性大于0.6的有46对, 占19.91%, 其中0.6-0.8的43对, 0.8以上3对, 相似性最大的为蓝靛果与葱皮忍冬(0.815), 其次为唐古特忍冬与银露梅(0.806)、葱皮忍冬与扁刺蔷薇(0.800)。
| 编号 No. |
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 |
| 2 | 0.684 | ||||||||||||||||||||
| 3 | 0.552 | 0.582 | |||||||||||||||||||
| 4 | 0.579 | 0.631 | 0.815 | ||||||||||||||||||
| 5 | 0.518 | 0.806 | 0.505 | 0.548 | |||||||||||||||||
| 6 | 0.609 | 0.566 | 0.568 | 0.553 | 0.540 | ||||||||||||||||
| 7 | 0.653 | 0.596 | 0.322 | 0.370 | 0.667 | 0.598 | |||||||||||||||
| 8 | 0.778 | 0.610 | 0.694 | 0.800 | 0.479 | 0.470 | 0.509 | ||||||||||||||
| 9 | 0.661 | 0.555 | 0.544 | 0.510 | 0.423 | 0.605 | 0.626 | 0.640 | |||||||||||||
| 10 | 0.630 | 0.766 | 0.735 | 0.746 | 0.725 | 0.584 | 0.478 | 0.701 | 0.501 | ||||||||||||
| 11 | 0.441 | 0.661 | 0.538 | 0.582 | 0.672 | 0.430 | 0.428 | 0.472 | 0.302 | 0.736 | |||||||||||
| 12 | 0.481 | 0.614 | 0.479 | 0.436 | 0.667 | 0.688 | 0.509 | 0.348 | 0.391 | 0.556 | 0.405 | ||||||||||
| 13 | 0.489 | 0.645 | 0.738 | 0.719 | 0.534 | 0.615 | 0.408 | 0.637 | 0.609 | 0.729 | 0.534 | 0.458 | |||||||||
| 14 | 0.218 | 0.323 | 0.424 | 0.387 | 0.290 | 0.219 | 0.186 | 0.300 | 0.053 | 0.441 | 0.592 | 0.157 | 0.335 | ||||||||
| 15 | 0.490 | 0.699 | 0.586 | 0.559 | 0.621 | 0.400 | 0.385 | 0.541 | 0.676 | 0.616 | 0.431 | 0.535 | 0.558 | 0.097 | |||||||
| 16 | 0.267 | 0.383 | 0.434 | 0.496 | 0.310 | 0.547 | 0.260 | 0.302 | 0.404 | 0.310 | 0.450 | 0.401 | 0.556 | 0.347 | 0.306 | ||||||
| 17 | 0.420 | 0.428 | 0.568 | 0.387 | 0.329 | 0.524 | 0.378 | 0.378 | 0.586 | 0.453 | 0.266 | 0.358 | 0.573 | 0.266 | 0.457 | 0.349 | |||||
| 18 | 0.318 | 0.540 | 0.491 | 0.507 | 0.479 | 0.259 | 0.229 | 0.474 | 0.238 | 0.593 | 0.536 | 0.353 | 0.507 | 0.567 | 0.505 | 0.186 | 0.225 | ||||
| 19 | 0.572 | 0.362 | 0.153 | 0.207 | 0.290 | 0.220 | 0.442 | 0.404 | 0.334 | 0.315 | 0.315 | 0.154 | 0.115 | 0.115 | 0.215 | 0.020 | 0.067 | 0.187 | |||
| 20 | 0.735 | 0.501 | 0.555 | 0.550 | 0.430 | 0.555 | 0.488 | 0.665 | 0.687 | 0.532 | 0.378 | 0.427 | 0.346 | 0.124 | 0.560 | 0.289 | 0.360 | 0.295 | 0.493 | ||
| 21 | 0.662 | 0.604 | 0.367 | 0.463 | 0.454 | 0.303 | 0.434 | 0.632 | 0.499 | 0.480 | 0.422 | 0.348 | 0.443 | 0.077 | 0.494 | 0.249 | 0.184 | 0.405 | 0.615 | 0.485 | |
| 22 | 0.218 | 0.323 | 0.424 | 0.387 | 0.476 | 0.277 | 0.401 | 0.300 | 0.053 | 0.451 | 0.565 | 0.299 | 0.335 | 0.785 | 0.097 | 0.227 | 0.266 | 0.537 | 0.115 | 0.124 | 0.077 |
| 物种编号同表 3 Species number saw Table 3 | |||||||||||||||||||||
综上可知, 紫果云杉群落乔木层各物种之间生态位均有相似性, 且相似性大于灌木层。生态位宽度大的物种间生态位相似性一般较大, 但非优势种之间也可见较大的生态位相似性。灌木层前22个优势或相对优势种之间未出现相似性为0或1的种对。相似性为0, 表明两物种未在同一资源位中出现, 相似性为1, 表示两物种在同一资源位出现的频度相同, 此外同科或同属的物种之间可见较大的生态位相似性。
2.4 生态位重叠紫果云杉群落8种乔木树种间均存在生态位重叠(表 8), 重叠均值为0.462, 重叠值大于0.6的有9对, 占总对数的32.14%, 其中大于0.8的有4对, 最大为糙皮桦和刺柏(0.890), 其次为糙皮桦和祁连圆柏(0.851)、祁连圆柏和刺柏(0.846)、秦岭柳和云杉(0.845), 但这几个种对中除祁连圆柏外, 其它均为弱势种。前4个优势种构成的6个种对生态位重叠值均在0.5以上, 其重叠平均值大于后4个弱势种。
灌木层1225个种对生态位重叠平均值为0.298, 总体重叠程度较低, 重叠值小于0.6的有974对, 占总对数的79.51%, 绝大多数介于0-0.4之间, 其中365个种对(占29.80%)重叠值为0, 表现出生态位分化现象。重叠值大于0.6的有251对, 占比20.49%, 其中0.6-0.8的180对, 0.8-0.9的34对, 0.9-1.0的21对, 有16个种对重叠值为1, 表现出生态趋同现象。50种灌木仅有峨眉蔷薇、唐古特忍冬、蓝靛果、华西忍冬和紫果云杉幼苗这5个种与其它所有种均有生态位重叠, 其中紫果云杉幼苗与其它种重叠的均值最大, 为0.504。前22个种构成的231个种对(表 9)生态位重叠均值为0.539, 其中重叠值小于0.6的有121对, 占比52.38%, 未出现重叠值为0或1的种对。0.6以上110对, 占比47.62%, 其中0.8-0.9的21对, 0.9-1.0的6对, 重叠值最大的为忍冬和栓翅卫矛(0.923), 其次为峨眉蔷薇和忍冬(0.923)、峨眉蔷薇与扁刺蔷薇(0.912)等。
| 编号 No. |
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 |
| 2 | 0.783 | ||||||||||||||||||||
| 3 | 0.565 | 0.677 | |||||||||||||||||||
| 4 | 0.703 | 0.717 | 0.892 | ||||||||||||||||||
| 5 | 0.628 | 0.882 | 0.536 | 0.484 | |||||||||||||||||
| 6 | 0.607 | 0.655 | 0.639 | 0.598 | 0.652 | ||||||||||||||||
| 7 | 0.816 | 0.748 | 0.400 | 0.431 | 0.819 | 0.738 | |||||||||||||||
| 8 | 0.912 | 0.748 | 0.712 | 0.875 | 0.491 | 0.483 | 0.600 | ||||||||||||||
| 9 | 0.839 | 0.716 | 0.663 | 0.635 | 0.484 | 0.684 | 0.653 | 0.758 | |||||||||||||
| 10 | 0.741 | 0.880 | 0.863 | 0.840 | 0.815 | 0.637 | 0.665 | 0.809 | 0.630 | ||||||||||||
| 11 | 0.563 | 0.801 | 0.658 | 0.706 | 0.815 | 0.437 | 0.590 | 0.607 | 0.347 | 0.816 | |||||||||||
| 12 | 0.393 | 0.728 | 0.524 | 0.407 | 0.831 | 0.793 | 0.598 | 0.269 | 0.432 | 0.674 | 0.510 | ||||||||||
| 13 | 0.543 | 0.748 | 0.832 | 0.796 | 0.496 | 0.677 | 0.458 | 0.675 | 0.722 | 0.744 | 0.553 | 0.516 | |||||||||
| 14 | 0.201 | 0.368 | 0.687 | 0.671 | 0.360 | 0.229 | 0.199 | 0.455 | 0.061 | 0.641 | 0.723 | 0.159 | 0.545 | ||||||||
| 15 | 0.598 | 0.829 | 0.679 | 0.597 | 0.633 | 0.471 | 0.393 | 0.601 | 0.747 | 0.723 | 0.508 | 0.639 | 0.665 | 0.115 | |||||||
| 16 | 0.280 | 0.520 | 0.564 | 0.617 | 0.366 | 0.653 | 0.288 | 0.318 | 0.429 | 0.382 | 0.532 | 0.448 | 0.695 | 0.380 | 0.383 | ||||||
| 17 | 0.379 | 0.419 | 0.779 | 0.479 | 0.353 | 0.633 | 0.354 | 0.375 | 0.727 | 0.545 | 0.222 | 0.469 | 0.676 | 0.268 | 0.585 | 0.373 | |||||
| 18 | 0.370 | 0.615 | 0.642 | 0.711 | 0.463 | 0.230 | 0.220 | 0.635 | 0.216 | 0.782 | 0.617 | 0.383 | 0.637 | 0.716 | 0.484 | 0.198 | 0.176 | ||||
| 19 | 0.910 | 0.615 | 0.239 | 0.473 | 0.476 | 0.274 | 0.690 | 0.803 | 0.617 | 0.521 | 0.446 | 0.131 | 0.227 | 0.030 | 0.415 | 0.010 | 0.016 | 0.253 | |||
| 20 | 0.905 | 0.686 | 0.600 | 0.687 | 0.554 | 0.550 | 0.621 | 0.817 | 0.790 | 0.692 | 0.489 | 0.423 | 0.398 | 0.094 | 0.683 | 0.243 | 0.429 | 0.273 | 0.807 | ||
| 21 | 0.901 | 0.771 | 0.378 | 0.617 | 0.520 | 0.358 | 0.632 | 0.876 | 0.698 | 0.636 | 0.492 | 0.280 | 0.480 | 0.093 | 0.621 | 0.182 | 0.096 | 0.482 | 0.923 | 0.780 | |
| 22 | 0.255 | 0.423 | 0.601 | 0.541 | 0.574 | 0.383 | 0.438 | 0.382 | 0.040 | 0.697 | 0.731 | 0.388 | 0.441 | 0.906 | 0.075 | 0.249 | 0.263 | 0.659 | 0.080 | 0.123 | 0.083 |
| 物种编号同表 3 Species number saw Table 3 | |||||||||||||||||||||
综上可知, 紫果云杉乔木层普遍存在生态位重叠, 且重叠程度大于灌木层。灌木层表现出生态位分化和生态趋同特征, 表明物种间可能存在着激烈的竞争。生态位宽度较大的物种之间重叠程度一般较大, 且与其它物种重叠的概率高、重叠程度大, 非优势种间、非优势种与优势种之间也可见较大的生态位重叠, 同科或同属的物种间生态位重叠往往较大。
3 结论与讨论 3.1 物种重要值与生态位宽度重要值表示植物在群落中的优势程度[17], 生态位宽度表示植物对环境的适应和对资源的利用能力[21]。本研究表明, 植物重要值越大生态位宽度也往往越大, 但也存在一定差异。在乔木层, 紫果云杉重要值和生态位宽度均远大于其它种, 占绝对优势, 但作为次优势种, 岷江冷杉的频数与生态位宽度却均小于祁连圆柏和红桦, 资源利用和适应能力均不如二者。灌木层生态位宽度最大的紫果云杉幼苗重要值仅为第9, 而重要值最大的峨眉蔷薇和唐古特忍冬的生态位宽度则为第12和第4。可见, 生态位宽度与重要值无明确相关规律[5], 其值大小还与重要值在资源位中分布的均匀程度有关[18], 即物种在各资源位分布越均匀, 重要值变异系数越小, 生态位宽度越大[22]。紫果云杉在灌木层虽无明显优势, 但所有样方均有其幼苗存在, 且分布均匀变异系数最小(表 3), 因此生态适应能力最强, 而峨眉蔷薇、唐古特忍冬等虽然所有样方也均有出现, 但其变异系数较大, 因此生态位宽度小于紫果云杉幼苗。
生态位宽度不仅与物种自身生态适应性有关, 也与其分布范围密切相关, 并受竞争和环境因子共同影响[5]。在灌木层, 重要值和生态位宽度均较大的物种多属忍冬科和蔷薇科, 且此二科所有种的重要值总和及生态位总宽度均大于其它所有种之和。大量研究表明, 忍冬科植物根系发达生根力强, 喜阳, 耐阴、耐寒, 适应性极强; 蔷薇科植物多喜光, 亦耐半阴, 较耐寒, 萌蘖性强, 适生范围广; 茶藨子科植物多耐寒, 喜湿润环境。本研究区紫果云杉群落乔木层郁闭度不高, 林隙较大, 高异质性的林下环境更有利于忍冬科和蔷薇科物种的生长。因此, 紫果云杉群落灌木层是以蓝靛果、葱皮忍冬、唐古特忍冬等忍冬科植物和以银露梅、峨眉蔷薇等蔷薇科植物为主体, 茶藨子科及其它科(种)为辅构成的物种多样和结构稳定的群落。
3.2 生态位相似生态位相似反映物种对资源利用的相似程度, 生态位宽度大的物种之间生态位相似性一般较大[19], 但也有研究认为, 生态位相似与生态位宽度无明显关联[17]。本研究中, 乔木层紫果云杉与3个优势种及其相互之间生态位相似性大于后4个弱势种, 但作为弱势种, 云杉与秦岭柳、刺柏与糙皮桦的相似性也大于0.6。灌木层紫果云杉幼苗与大多优势种有较大的生态位相似性, 与非优势种的相似性则普遍较低。相似性较大的蓝靛果与葱皮忍冬、唐古特忍冬与银露梅等均有较大的生态位宽度, 但相似性大于0.8的9个种对有6个种对出现在非优势种之间、非优势种与优势种之间, 包括相似性大于0.9的刚毛忍冬与毛花忍冬、花楸与云杉幼苗, 可见, 生态位宽度较小的物种间也可出现高度的生态位相似性。本研究认为, 生态位相似受物种生态特性及生境共同影响, 在同一群落相同层次, 生态特性相近的物种因资源需求相似常占有相似的生态位, 因此, 同科或同属物种间常有较高的生态位相似性, 如金露梅与平枝栒子等。
调查研究发现, 本研究区紫果云杉群落多分布于中下坡位, 并有轻、中度干扰存在。乔木层林木高大, 受干扰影响小, 灌木层受到干扰时, 生态位宽度小的物种常有消失的风险, 但由于种间生态位相似性, 一个物种的消失会得到与其生态位高度相似的另一物种的补充, 因此生态位相似还反映着群落的稳定性[23], 而弱势种或偶见种的消失通常不影响群落结构。紫果云杉幼苗、峨眉蔷薇、唐古特忍冬、蓝靛果和华西忍冬的生态位宽度大且与所有种均有生态位相似性, 因此, 任何物种的消失都可能由这5个种来补充, 这有利于紫果云杉种群的发展和群落稳定。相似性为0或1的种对多出现在非优势种之间, 表现出生态位相斥和生态趋同现象, 这可能是由于物种在长期共存中, 竞争及干扰引起了生态位分化和对资源需求的互补或重叠[24], 也可能是该研究区并不完全反映所有种的生态位特征。
3.3 生态位重叠生态位重叠是两个或多个物种对同一资源或多个资源共同利用的量化表达[21, 25], 通常生态位宽度大的物种分布均匀, 资源利用能力强[6, 26], 与其它物种重叠概率高、重叠程度大[27]。本研究中紫果云杉与其它7个乔木树种生态位重叠的均值最大, 其次为祁连圆柏与红桦, 但弱势种秦岭柳和云杉的生态位重叠值也达0.8以上。在灌木层, 仅生态位宽度最大的前5个种与其它所有种有重叠, 其中紫果云杉幼苗与其它种重叠的均值最大, 其次为唐古特忍冬、华西忍冬等。生态位宽度较大的前12个种与其它种重叠均值(0.356-0.504)大于16个偶见种与其它种重叠的均值(0.117-0.281), 但生态位宽度较小的华西蔷薇与宝兴茶藨子, 平枝荀子与金露梅等重叠值也达0.9以上, 可见, 生态位重叠值大小与生态位宽度无明确的相关规律[28]。本研究认为, 弱势种群适应性弱, 易受排挤, 再加上生境的异质性, 当其在某个适宜生境集中生长时也会出现较大的生态位重叠[17], 因此生态位重叠是受物种生态特性和生境共同影响[29]。同科或同属的物种间由于趋于相同的生活型及相似的生态特性常有较大生态位重叠[30], 如扁刺蔷薇和峨眉蔷薇等。
生态位重叠反映着不同物种在资源共享时存在的竞争及相互促进的关系[31], 因此其值的大小并不完全代表种间竞争程度。紫果云杉群落乔木层普遍存在生态位重叠, 且重叠均值较大, 灌木层前22个优势种间生态位重叠均值也较大, 但所有种的生态位重叠均值仅为0.298, 总体重叠程度较低。生态位重叠值越大, 预示着占据共同生态空间的物种间潜在的竞争越大[32-33]。调查研究发现, 紫果云杉群落乔木层垂直分层明显, 物种多样性低, 数量少, 空间资源充足, 因此, 生态位重叠主要反映物种间的共存状态与竞争发生可能性, 以及具有相似生境需求的物种间的相互促进关系[31], 如桦木等喜阳树种的存在会为紫果云杉的更新提供有利条件。而灌木层植物多达50种, 物种多样性较高, 且多在适宜的生境下集中分布, 如林窗等, 因此, 必会产生因资源不足引起的竞争排斥现象, 因此生态位重叠在反应物种共存的同时主要反映种间竞争程度, 365个种对出现生态位分化及16个种对表现出生态趋同进一步证明了竞争的存在。
经本研究可知, 紫果云杉作为建群种占据着群落大部分空间, 主导着群落演替, 岷江冷杉和祁连圆柏等为伴生种, 共同影响着群落生存状态, 桦木等阳性树种与紫果云杉等针叶树种形成生态互补和相互促进的关系。紫果云杉、岷江冷杉等幼苗耐阴性强, 更新初期需要荫蔽环境[12], 林内广泛分布的忍冬、蔷薇等优势种(科)在与其幼苗竞争的同时也为其更新提供了有利条件。乔、灌木层共有物种及其分布状况反映着物种的扩散能力[34], 紫果云杉在乔、灌木层均有最大的生态位宽度, 生态适应性和资源利用能力最强, 具有进一步扩散的可能或正处于扩散阶段。
致谢: 感谢洮河林区和青海黄南州麦秀林场工作人员给予的帮助。| [1] |
Grinnell J. The niche-relationships of the California Thrasher. The Auk, 1917, 34(4): 427-433. DOI:10.2307/4072271 |
| [2] |
彭文俊, 王晓鸣. 生态位概念和内涵的发展及其在生态学中的定位. 应用生态学报, 2016, 27(1): 327-334. |
| [3] |
牛克昌, 刘怿宁, 沈泽昊, 何芳良, 方精云. 群落构建的中性理论和生态位理论. 生物多样性, 2009, 17(6): 579-593. |
| [4] |
储诚进, 王酉石, 刘宇, 蒋林, 何芳良. 物种共存理论研究进展. 生物多样性, 2017, 25(4): 345-354. |
| [5] |
陈玉凯, 杨琦, 莫燕妮, 杨小波, 李东海, 洪小江. 海南岛霸王岭国家重点保护植物的生态位研究. 植物生态学报, 2014, 38(6): 576-584. |
| [6] |
林勇, 艾训儒, 姚兰, 黄伟, 陈斯. 木林子自然保护区不同群落类型主要优势种群的生态位研究. 自然资源学报, 2017, 32(2): 223-234. |
| [7] |
董雪, 李永华, 张正国, 李思瑶, 包岩峰, 郝玉光, 姚斌. 甘肃酒泉荒漠戈壁灌木群落优势物种生态位特征. 中国沙漠, 2020, 40(4): 138-145. |
| [8] |
王刚, 赵松岭, 张鹏云, 陈庆诚. 关于生态位定义的探讨及生态位重叠计测公式改进的研究. 生态学报, 1984, 4(2): 119-127. |
| [9] |
Dolédec S, Chessel D, Gimaret-Carpentier C. Niche separation in community analysis: a new method. Ecology, 2000, 81(10): 2914-2927. DOI:10.1890/0012-9658(2000)081[2914:NSICAA]2.0.CO;2 |
| [10] |
李登武, 张文辉, 任争争. 黄土沟壑区狼牙刺群落优势种群生态位研究. 应用生态学报, 2005, 16(12): 2231-2235. DOI:10.3321/j.issn:1001-9332.2005.12.002 |
| [11] |
Müller R, Nowicki C, Barthlott W, Ibisch P L. Biodiversity and endemism mapping as a tool for regional conservation planning-case study of the Pleurothallidinae(Orchidaceae)of the Andean rain forests in Bolivia. Biodiversity&Conservation, 2003, 12(10): 2005-2024. |
| [12] |
赵阳, 刘锦乾, 陈学龙, 杨萌萌, 曹家豪, 齐瑞, 曹秀文. 洮河上游紫果云杉种群结构特征. 植物生态学报, 2020, 44(3): 266-276. |
| [13] |
赵阳, 曹秀文, 李波, 齐瑞, 曹家豪, 陈学龙, 杨萌萌, 陈林生. 甘肃南部林区4种天然林种群结构特征. 林业科学, 2020, 56(9): 21-29. |
| [14] |
邱书志, 王伟, 丁骞, 杨永林, 汪淑筠, 王少明, 闫毓斌, 赵亮生. 洮河林区森林生态系统服务功能及价值评估. 中南林业科技大学学报, 2018, 38(2): 97-102. |
| [15] |
许幼霞, 周旭, 赵娟, 周秋文, 韦小茶. 半湿润半干旱过渡区洮河流域植被盖度变化特征. 水土保持通报, 2016, 36(6): 308-314. |
| [16] |
伏捷, 孙才志, 张华, 何红, 祝业平, 王颖. 辽东山地老秃顶子古冰缘地貌植物种群生态位特征. 地理研究, 2018, 37(4): 731-741. |
| [17] |
刘润红, 常斌, 荣春艳, 姜勇, 杨瑞岸, 刘星童, 曾惠帆, 傅桂焕. 漓江河岸带枫杨群落主要木本植物种群生态位. 应用生态学报, 2018, 29(12): 3917-3926. |
| [18] |
吴会峰, 宋丽娟, 杜峰, 郝文芳. 黄土丘陵区草地主要种群生态位及其环境解释. 应用生态学报, 2017, 28(11): 3494-3504. |
| [19] |
铁军, 李燕芬, 金山, 陈艳彬, 张桂萍, 茹文明. 山西历山国家级自然保护区猕猴栖息地优势树种生态位特征. 生态学杂志, 2015, 34(3): 634-641. |
| [20] |
管杰然, 商天其, 伊力塔, 叶诺楠, 余树全. 天目山常绿落叶阔叶混交林优势种生物量变化及群落演替特征. 生态学报, 2017, 37(20): 6761-6772. |
| [21] |
刘加珍, 陈亚宁, 张元明. 塔里木河中游主要植物种群的生态特征分析. 地理研究, 2003, 22(5): 663-670. DOI:10.3321/j.issn:1000-0585.2003.05.015 |
| [22] |
陈俊华, 刘兴良, 何飞, 刘世荣. 卧龙巴朗山川滇高山栎灌丛主要木本植物种群生态位特征. 林业科学, 2010, 46(3): 23-28. |
| [23] |
张伟, 王万林, 胡玉昆, 尹伟, 公延明, 房飞, 阿德列提·艾列吾塔力甫. 新疆伊犁荒漠草原恢复措施下植物群落生态位特征分析. 中国生态农业学报, 2011, 19(2): 358-362. |
| [24] |
郝建锋, 李艳, 齐锦秋, 裴曾莉, 黄雨佳, 蒋倩, 陈亚. 人为干扰对碧峰峡栲树次生林群落物种多样性及其优势种群生态位的影响. 生态学报, 2016, 36(23): 7678-7688. |
| [25] |
牛慧慧, 陈辉, 付阳, 杨祎, 张斯琦, 张博雄. 柴达木盆地东部荒漠植物生态位特征. 生态学报, 2019, 39(8): 2862-2871. |
| [26] |
张继强, 陈文业, 谈嫣蓉, 刘冬皓, 袁海峰, 王斌杰, 刘鸿源, 陈旭. 甘肃敦煌西湖湿地芦苇盐化草甸植物群落生态位特征研究. 南京林业大学学报: 自然科学版, 2019, 43(2): 191-196. |
| [27] |
班卫强, 严成, 尹林克, 杨美琳, 王忠臣. 古尔班通古特沙漠南缘不同立地条件植物多样性和优势种群生态位特征研究. 中国沙漠, 2012, 32(6): 1632-1638. |
| [28] |
陈艳瑞, 尹林克. 人工防风固沙林演替中群落组成和优势种群生态位变化特征. 植物生态学报, 2008, 32(5): 1126-1133. DOI:10.3773/j.issn.1005-264x.2008.05.017 |
| [29] |
郭平平, 税伟, 江聪, 简小枚, 朱粟锋, 张永永, 冯洁, 陈毅萍. 退化天坑倒石坡林下优势物种生态位特征. 应用生态学报, 2019, 30(11): 3635-3645. |
| [30] |
董雪, 辛智鸣, 段瑞兵, 黄雅茹, 菅凯敏, 马海峰. 乌兰布和沙漠典型灌木群落多样性及其生态位. 干旱区研究, 2020, 37(4): 1009-1017. |
| [31] |
王立龙, 王广林, 黄永杰, 李晶, 刘登义. 黄山濒危植物小花木兰生态位与年龄结构研究. 生态学报, 2006, 26(6): 1862-1871. DOI:10.3321/j.issn:1000-0933.2006.06.030 |
| [32] |
潘高, 张合平, 潘登. 湖南青羊湖国家森林公园针阔混交林主要种群生态位特征. 中南林业科技大学学报, 2017, 37(2): 69-75. |
| [33] |
林思祖, 黄世国, 洪伟, 黄宝龙, 俞新妥. 杉阔混交林主要种群多维生态位特征. 生态学报, 2002, 22(6): 962-968. DOI:10.3321/j.issn:1000-0933.2002.06.025 |
| [34] |
刘海丰, 薛达元, 桑卫国. 暖温带森林功能发育过程中的物种扩散和生态位分化. 科学通报, 2014, 59(24): 2359-2366. |