 2022, Vol. 42
2022, Vol. 42文章信息
- 阮欧, 刘绥华, 陈芳, 罗杰, 胡海涛
- RUAN Ou, LIU Suihua, CHEN Fang, LUO Jie, HU Haitao
- 基于多源遥感的贵州草海国家级自然保护区黑颈鹤生境适宜性评价
- Habitat suitability evaluation of black-necked cranes based on multi-source remote sensing in Caohai National Nature Reserve, Guizhou
- 生态学报. 2022, 42(5): 1947-1957
- Acta Ecologica Sinica. 2022, 42(5): 1947-1957
- http://dx.doi.org/10.5846/stxb202010282765
-
文章历史
- 收稿日期: 2020-10-28
- 网络出版日期: 2021-11-17





2. 贵州省山地资源与环境遥感应用重点实验室, 贵阳 550025
2. Key Laboratory of Mountain Resources and Environmental Rensing Sensing, Guizou Normal University, Guiyang 550025, China
湿地生态系统可以为珍稀濒危物种提供必要的栖息地和觅食地[1]。近年来由于人为活动和全球气候变化的影响导致湿地生境的丧失与退化威胁到了水禽的生存和繁殖, 尤其是黑颈鹤(Grus nigricollis)等野生濒危鸟类[2]。因此, 了解黑颈鹤的生境要求并评估其生境质量可以更有效地开展湿地水鸟及其栖息地的保护和管理。
黑颈鹤是世界上15种鹤类中最晚被发现的鹤类, 也是唯一终生生活在高原的鹤类[3]。由于其数量较少, 被列入《世界自然保护联盟》(IUCN)易危种(VU), 是我国I级重点保护野生动物[4]。贵州草海国家级自然保护区是黑颈鹤重要的越冬地和迁徙中转站, 也是当前黑颈鹤越冬数量最多的自然保护区之一, 2015年草海自然保护区越冬黑颈鹤数量就已经达到了2000余只[5]。因此, 受到了众多学者的关注。当前, 国内外学者对物种生境适宜性的空间分布进行了大量的研究, 其中通过建立物种出现点与其周围生境特征之间相关性的经验物种分布模型已经成为了当前预测物种生境分布的研究热点。此类研究运用的预测和评估模型主要包括回归模型分析[6-7]、最大熵模型(MaxEnt)[8-9]、生态位因子分析(ENFA)[10]、机理模型等[11], 其中最大熵模型(MaxEnt)以其具有操作简单、对物种出现点样本数量要求低、预测结果较准确等特点被众多学者认为是预测和评价物种生境适宜性的最佳模型[12-13]。
在过去, 草海地区为了发展农业和旅游业, 开展了包括围湖造田、建造观景台及开发旅游项目等活动, 不可避免地破坏了当地的景观, 给保护区带来了更多的人为干扰。近年来, 这些现象在政府的努力下得到了明显改善, 湿地环境得到不断恢复, 越冬黑颈鹤的数量也在不断增加, 由于草海自然保护区靠近威宁县城, 同时也是著名的景区, 人为干扰依旧强烈, 研究当前越冬黑颈鹤生境适宜性对自然保护区今后保护和管理方案的提出同样具有重要意义;MaxEnt模型在预测物种生境适宜性分析时输入的环境变量通常包括自然、人文等因子等, 现有多数研究输入的环境因子都是各大科研机构提供的免费数据, 普遍存在分辨率低的问题, 虽然对大尺度范围的物种生境适宜性研究影响并不大, 但是在小尺度研究区的预测和评估上往往会导致较大的误差。因此, 在小尺度研究区物种生境适宜性的预测上需要精度更高的环境因子数据才能满足需求, 才能更加准确地预测物种的实际分布状况。本文通过多源遥感数据以及地形辅助数据建立影响黑颈鹤生境适宜性的9个分辨率为8m的环境变量, 运用MaxEnt模型预测和评估草海自然保护区黑颈鹤精细的栖息地与觅食地, 以为草海自然保护区越冬黑颈鹤的保护和管理提供科学参考。
1 材料与方法 1.1 研究区概况草海国家级自然保护区, 位于贵州省西部威宁县县城西南面, 介于东经104°10′—104°20′, 北纬26°47′—26°53′之间(图 1)。研究区湖面海拔2171.7m, 水域面积为25km2, 是黑颈鹤等228种鸟类的重要越冬地和迁徙中转站。草海自然保护区因其生物多样性、生态环境典型、物种丰富而吸引着大量黑颈鹤来此越冬。据文献记载(图 2), 1985—1986年在草海越冬的黑颈鹤仅为223只, 之后逐年上升, 到1997年, 草海黑颈鹤数量已达到512只, 2011—2012年为1300只, 2013年达到1450只以上, 2014年超过1700只, 2015年在开展冬季水鸟调查时, 在草海发现的黑颈鹤已达到2000余只, 成为目前黑颈鹤越冬数量最多自然保护区之一[5, 14]。

|
| 图 1 草海国家级自然保护区位置及黑颈鹤出现点示意图 Fig. 1 Schematic diagram of the location of Caohai National Nature Reserve and the appearance point of black-necked crane |
1.2 数据准备及环境因子生成 1.2.1 栖息地与觅食地出现点数据

2018年11—12月期间对草海地区黑颈鹤进行实地调查。在确定觅食地的位置时, 采用的方法是在不打扰黑颈鹤觅食的情况下站在位置较高的地方利用双目望远镜(放大10—60倍)观察黑颈鹤的觅食行为, 如果该点有觅食行为且在不受干扰的情况下觅食时间超过15min以上时, 将该点确定为觅食地, 并在黑颈鹤离开后用GPS对该点进行地理位置记录。对于栖息地的确定, 首先向草海自然保护区工作人员及周边居民了解黑颈鹤的栖息地大致分布情况, 然后在早上黑颈鹤外出觅食飞出栖息地后, 进入栖息地进行GPS定位, 对于无法进入但能在高地用望远镜观察到的地方采用高清遥感影像目测的方法在影像中确定地理位置。一共观察到觅食点和栖息点共112个, 为避免数据的过于拟合, 以100m为阈值对数据进行筛选, 剔除距离小于100m的点位。通过筛选最终选取草海地区黑颈鹤觅食地点位28个, 栖息地点位44个进行分析。
1.2.2 环境变量数据黑颈鹤并非随机选择栖息地生境, 影响其栖息地选择的环境因子主要包括水源、食物、干扰源和遮蔽物[3, 15]。根据生境适宜性的评价原则, 结合前人的研究和草海地区实际情况, 选取了距水域距离、距耕地距离、距建筑物距离、距道路距离、植被覆盖度、优势植被分布、水深、坡度、坡向等9个生境因子。实验流程如下: ①选取2019年11—12期间的高分1号(GF-1)、高分3号(GF-3)号遥感卫星数据协同对研究区土地利用分类, 并基于分类数据提取人为干扰源与水域;②利用GF-1多光谱数据计算植被盖度;③选取2019年10月14日的高分5号(GF-5)号采用混合像元分解的方法反演优势植被的覆盖度状况;④基于Landsat8 OLI与实地测量的水深数据采用回归模型反演草海水深;⑤ALOS 12.5m DEM用于计算草海地区的坡度、坡向。环境变量获取后, 通过ArcGIS10.2平台对所有环境因子统一范围裁剪, 并统一将投影设置为WGS_1984坐标, 分辨率重采样为8m, 以MaxEnt模型的输入格式ASC输出, 然后将黑颈鹤出现点数据与环境变量输入MaxEnt模型进行分析, 最后再将预测结果加载到ArcGIS10.2, 对生境适宜性进行等级划分、面积计算和可视化表达。各数据源详细的数据参数见表 1。
| 数据源Data source | 时间time | 参数Parameter | 获取方式Acquisition method |
| 高分一号Gaofen-1 | 2019年11月22日 | 多光谱4波段, 分辨率8m | 高分贵州分中心 |
| 高分三号Gaofen-3 | 2019年12月7日 | 全极化条带, 分辨率8m | |
| 高分五号Gaofen-5 | 2019年9月14日 | 高光谱330波段, 分辨率30m | |
| 陆地卫星8号Landsat8 OLI | 2018年10月8日 | 多光谱9波段, 分辨率30m | 地理空间数据云 |
| 数字高程模型Digital Elevation Model | 2010年 | 分辨率12.5m | 美国国家航空航天局官网 |
(1) 土地利用类型
土地利用分类采用经过预处理后的GF-1多光谱数据与C波段全局化GF-3合成孔径雷达数据协同利用支持向量机(SVM)的方法进行, 该方法已得到李鹏等人证实, 两者协同的分类总精度可达90%, 高于单独使用一种数据源的分类精度[16]。本文结合草海地区5种地物的光谱特征、纹理特征、各类指数特征以及极化散射特征等信息, 利用SVM方法将研究区分为林地、建筑物、草地、耕地以及水域5种地类, 通过对分类后的地类进行精度评价, 结果如图 3, 可以看出除了耕地和建筑外其他土地类型的生产者精度和用户精度都在90%以上, 分类精度整体较好。

|
| 图 3 土地利用分类精度 Fig. 3 Accuracy of land use classification |
(2) 植被盖度
本文采用植被指数转换模型计算研究区的植被覆盖度[17]。具体实现流程如下:
首先, 计算归一化植被指数。其表达式为:
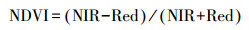
|
(1) |
式中, NIR为GF-1第4波段, Red为第3波段。
根据上述公式计算研究区的NDVI值, NDVI值服从正态随机分布, 其小于5%和大于95%的值不是植被覆盖程度的反映。选择NDVI=5%的值为NDVImin, NDVI=95%的值为NDVImax, 按照如下公式计算植被覆盖度[18]:

|
(2) |
植被覆盖度(VFC)值小于0或者大于1的区域通常是河流、道路、居民点等地类, 直接归为VFC=0, 既植被覆盖度为0。植被覆盖度VFC在(0, 1]区间的表示真实的植被覆盖度。
(3) 优势植被分布
通过2018年11月2日对草海的优势植被进行实地调查, 发现草海自然保护区的主要优势植被主要是水葱(Scirpus validus Vahl)、荆三稜(Scirpus yagara)等沙草科植被, 这些植被的块茎是黑颈鹤的主要食物来源[19]。通过对这些优势植被进行光谱采集和GPS定位, 利用GF-5号高光谱数据通过像元解混获取其分布状况及丰度。实现步骤是首先对GF-5号高光谱数据进行坏波段剔除、条纹去除、辐射定标、大气校正和正射校正等预处理, 最后利用基于线性光谱解混模型的全约束最小二乘法对优势植被进行丰度反演。
(4) 水深
草海自然保护区水深图是由2018年10月8日获取的Landsat 8多光谱数据与2018年11月2—3日实地测得的水深数据, 通过回归得到水深与影像之间关系而得, 通过对反演的水深与实测水深进行拟合分析, 其R2为0.61, 能一定程度上反应草海地区的水深。
(5) 地形因子
地形因子包括坡向、坡度因子。利用ArcGIS 10.2的表面分析工具对ALOS 12.5m的DEM进行处理生成坡向与坡度并以研究区的边界进行裁剪然后对其进行重采样, 分辨率与土地利用保持一致即8m。
(6) 距人为干扰和水域的距离
从草海保护区的土地覆盖图中提取建筑、耕地、道路和水域等因子, 通过ArcGIS 10.2空间分析工具中的欧氏距离计算研究区域内每个像素距建筑物、道路、耕地以及水域的最近距离, 各环境变量结果见图 4。

|
| 图 4 研究区各环境因子示意图 Fig. 4 Schematic diagram of environmental factors in the study area |
草海地区黑颈鹤的生境适宜性分析, 选用的是Philips和Dudík开发的MaxEnt软件(Version 3.4.1)进行的[9, 20], 将经过筛选后的28个觅食地点位、44个栖息地点位以及9个环境因子导入软件, 两种点位分别随机选取75%的点位作为模型的训练集, 其余25%作为测试点, 选择刀切法(Jackknife)来检测各变量的重要性, 模型中其余参数都保持默认设置, 已有研究表明保持默认设置也能较好的满足对结果的需求[21-22]。为了保证模型稳定性, 采用10次重复交叉验证的平均栖息地和觅食地的适宜性指数作为最终的结果。
模型运行结果的精度检验采用受使用者工作特征曲线(ROC曲线)下的面积值(AUC)进行, 理论上AUC的值是在0—1之间, 值越大代表模型的预测的结果精度越高[23]。一般认为, AUC值的评判标准为: 低于0.6的表明模型不能区分偏好生境和环境背景, 模型预测失败;0.6—0.7之间是代表效果较差;0.7—0.8表示效果一般;0.8—0.9代表效果较好;大于0.9表示效果优秀[13, 24]。
2 结果 2.1 模型预测结果评估从模型的ROC曲线可以看出(图 5), 模型10次重复计算后的测试集栖息地的AUC平均值为0.986, 觅食地的AUC平均值为0.948, 两者的AUC值均大于0.94, 表明模型的效果达到了极好水平, 预测结果较为合理可靠, 可以用于对草海自然保护区黑颈鹤栖息地与觅食地适宜性分布的预测分析。

|
| 图 5 栖息地与觅食地ROC曲线验证结果 Fig. 5 ROC curve of MaxEnt prediction |
各环境变量对模型的相对贡献率如图 6所示, 在栖息地模型预测中, 距耕地距离、距水域距离、水深及距建筑距离是影响黑颈鹤栖息的主要环境因子, 其中贡献率最大的是距耕地距离, 占比为43.80%, 紧随其后的距水域距离为25.50%、水深为13.00%、距建筑距离为6.20%, 这4个环境变量对模型的贡献率为88.50%。说明这4个环境因子对黑颈鹤的栖息地适宜性影响程度最大。而与优势植被、距道路距离、植被盖度、坡度和坡向等对模型的贡献率均在4%以下, 表明对黑颈鹤的栖息地适宜性影响较小。图 7栖息地环境因子响应曲线表明, 黑颈鹤偏好距离耕地200—300m、离水域较近、水深在0—20cm且远离人类居住500m左右的区域栖息。在觅食地的模型预测结果中, 距耕地距离、优势植被、距建筑物距离和水深对模型的相对贡献率较大, 占总贡献率的89.50%, 其中距耕地距离为38.00%、优势植被分布为23.50%、距建筑物距离为14.70%、水深为13.30%。贡献率最小的5个环境因子为植被盖度(1.40%)、道路(1.80%)、坡度(4.30%)、水域(1.90%)以及坡向(0.80%)。从图 7觅食地环境因子响应曲线可以看出, 黑颈鹤觅食地更喜欢选择在与建筑物有一定距离的耕地或者优势植被覆盖度较高、水深在0—20cm区域, 其中耕地遗留的农作物籽粒以及优势植被的种子与块茎可提供丰富的食物。

|
| 图 6 各环境因子对模型的相对贡献率 Fig. 6 Variation of the contributions of the main environmental factors to distribution of wintering habitats of black-necked cranes |

|
| 图 7 主要环境变量黑颈鹤出现概率响应曲线 Fig. 7 Probability response curve of main environmental variables black-necked crane |
通过ArcGIS 10.2的重分类功能采用自然断点法将草海自然保护区黑颈鹤的栖息地与觅食地分为最适宜区、次适宜区与不适宜区三种类型, 得到黑颈鹤栖息地与觅食地适宜性分布图(图 8)。从表 2可以看出最适宜黑颈鹤栖息的地区面积仅为2.163km2, 次适宜栖息地面积也只有4.241 km2, 不适宜栖息的地区面积为93.235km2, 占比达到了研究区总面积的93.573%, 而最适宜和较适宜的栖息地仅占6.427%。从图 8中可以看出, 最适宜区主要围绕草海水体岸边, 形状为细窄的条带状, 较适宜栖息的地区主要围绕最适宜的地区分布, 分布较为破碎;黑颈鹤觅食地最适宜和次适宜分布地与栖息地的分布状况基本相同, 都围绕着草海水体分布, 但面积范围扩展到了部分耕地, 次适宜区较为破碎。其中最适宜区面积为6.049 km2, 次适宜区面积为6.595 km2, 不适宜区面积为86.994 km2, 最适宜区与次适宜区在草海水域的东部和西部分布最为集中。

|
| 图 8 黑颈鹤栖息地与觅食地适宜性分布 Fig. 8 Suitability distribution of black-necked crane habitat and foraging place |
| 类型 Type |
面积及比例 Area and proportion |
最适宜区 Optimum area |
次适宜区 Sub-suitable area |
不适宜区 Inappropriate area |
| 栖息地Habitat | 面积/km2 | 2.163 | 4.241 | 93.235 |
| 比例/% | 2.171 | 4.256 | 93.573 | |
| 觅食地Foraging | 面积/km2 | 6.049 | 6.595 | 86.994 |
| 比例/% | 6.070 | 6.619 | 87.308 |
MaxEnt模型多被用于预测和评价物种的生境适宜性, 在当前的众多模型中被认为是效果最好的物种分布预测模型, 已广泛运用在濒危珍稀鸟类的生境适宜性预测研究中, 且都取得了较为理想的结果。例如, 那晓东等[25]利用数据MaxEnt模型绘制了典型湿地保护区内濒危水禽的精细尺度繁殖栖息地。成功地识别了研究区域内潜在的合适筑巢地点, 且与实地调查的筑巢位置相匹配;曹铭昌等[26]利用MaxEnt模型对盐城保护区丹顶鹤越冬生境变化进行了分析, 模型预测结果与保护区丹顶鹤同步调查结果基本保持一致。本研究展示了高分辨率多源数据, 包括高分系列遥感数据(GF-1高分辨率光学遥感、GF-3合成孔径雷达和GF-5高光谱遥感影像)、Landsat8多光谱遥感以及12.5m DEM等在黑颈鹤栖息地和觅食地生境预测中的效用, 虽然收集的草海自然保护区黑颈鹤的栖息点与觅食点样本并不多, 但由于多源遥感数据生成的环境因子的空间分辨率高, 从MaxEnt模型的预测结果来看最终的效果较为理想。预测结果的空间分布与武大伟等[27]近年在草海湿地观测的黑颈鹤觅食地和栖息地位置空间分布基本相符。
3.2 黑颈鹤生境适宜性分析水、食物、人为干扰源和遮蔽物被认为是影响黑颈鹤分布的主要环境因子[3, 15]。本研究表明, 在草海湿地黑颈鹤在对栖息地的选择中更偏好距离耕地200—300m、离水域较近、水深在0—20m且远离人类居住点500 m左右的区域, 对于觅食地的选择则更喜欢选择在与建筑物有一定距离的耕地或者优势植被覆盖度较高且水深在0—20cm的区域, 这与前人调查研究的结果是相符的。从本研究结果中可以看出草海地区黑颈鹤的栖息地与觅食地都表现出对耕地的高度依赖性, 相对贡献率都在38%以上, 这一结果并不意外, 因为耕地在这个研究区的占比非常大, 从前人对黑颈鹤生境的研究中也可以发现, 在云南大山包[28]、永善县[29]以及青藏高原的拉萨河谷[30]等地, 黑颈鹤经常出现在耕地中觅食, 耕地是黑颈鹤生境的重要组成部分, 这与本文研究结果一致。这一现象可能是因为耕地会遗留如马铃薯、玉米等残余作物, 已有研究表明玉米在冬季为黑颈鹤提供了80%的食物供应[31], 因此耕地提供的食物会比其他的环境因子更多。但也有一部分研究表明黑颈鹤对沼泽地表现出强烈的偏好[28], 出现不同观点的原因可能是由于在观察黑颈鹤觅食与栖息地的时间不同导致的, 11月至次年1月黑颈鹤的主要觅食地在耕地, 而2月—3月这期间黑颈鹤会通过整理羽毛、鸣叫和跳舞的行为吸引异性[32], 为即将到来的繁殖期做准备, 沼泽地为这些行为提供了最佳的环境[28]。另外2月—3月也为春耕期, 强烈的人为活动干扰会迫使黑颈鹤离开耕地而选择沼泽觅食。其次, 研究也发现20cm以下的浅水区在黑颈鹤的生境选择中起着重要的作用, 可能是因为浅水区可为黑颈鹤的栖息、觅食、社会行为和饮水提供条件, 这一现象与云南大山包黑颈鹤越冬行为的研究是一致的[33];另外, 人为干扰因子源也是影响黑颈鹤觅食地与栖息地选择的重要因素[34-35], 在青藏高原和云南大山包的黑颈鹤生境研究中, 远离道路和住宅区更有利于黑颈鹤的觅食, 但是从本文研究结果来看, 只有建筑这一人为干扰因子对黑颈鹤的生境选择贡献率稍大, 但也仅为6.20%和14.70%, 而距道路距离对黑颈鹤生境的影响并不大, 对栖息地与觅食地选择的贡献率都在2%以下, 造成这一现象的原因可能是因为草海附近建筑较多且分布较广, 同时农耕区作物的耕种和运输等人类活动主要依赖于道路, 当地在耕作区域修建了许多的道路来促进农业的发展, 使得耕地被路网分割为小块, 大多数的耕地与道路的直线距离都在100m以内, 使得觅食地分布必须在道路和建筑物附近, 没有其他选择。
3.3 黑颈鹤的保护建议了解物种的生境适宜性是对其进行保护和管理的基础, 水禽作为湿地生态系统中最具有代表性的动物类群, 灵敏反映着湿地生境质量的变化, 是湿地生态系统健康的指示物种[36]。研究发现, 黑颈鹤的栖息地与觅食地主要围绕湖泊周围分布, 耕地、优势植被分布、水源、水深等都是黑颈鹤生境选择的重要因素。由于草海地区主要以农业为主, 同时也是旅游胜地, 游客量大, 黑颈鹤的栖息地与觅食地距离人类活动的区域较近。在过去, 人们对湿地的价值认识比较淡泊, 保护意识不强, 围湖造田、湖内捕鱼和游客活动范围没有约束等现象普遍存在, 导致黑颈鹤的生境适宜性区域变得非常小, 选择在草海越冬的黑颈鹤数量较少。近年来, 在政府的重视下, 草海自然保护区开始坚守发展和生态两手抓, 在黑颈鹤栖息地周围实施湿地生态效益补偿、退耕还湖、拆除观景台等措施恢复湿地, 很大程度上改善了黑颈鹤的生境适宜性和生存空间, 黑颈鹤数量也在不断增长。虽然采取了一系列生态保护措施, 但当前旅游业越来越发达, 旅游人数不断增加, 提高旅游人员和当地居民的保护意识也很重要, 尤其是在越冬黑颈鹤数量越来越多的今天, 针对上述问题, 建议景区采取以下措施: 1)在宣传旅游的同时宣传保护黑颈鹤的重要性, 提高游客的保护意识;2)在景区划定指定的观鸟和拍摄地, 禁止游客私自进入黑颈鹤栖息地;3)加强对当地居民宣传教育和管理, 提高其对黑颈鹤的自觉保护意识;4)耕地收获遗留的作物是黑颈鹤越冬期间重要的食物来源, 保护区应鼓励当地居民多种植玉米等黑颈鹤喜爱农作物, 或者建立相应的食物源基地, 以保证黑颈鹤越冬期间的食物需求。
4 结论黑颈鹤是一种野生濒危鸟类, 其对栖息地和觅食地的选择都有特殊的要求, 如植被种类、水域、水深、植被覆盖和人为干扰等。本文采用高分辨率多源遥感影像与地形辅助数据结合通过MaxEnt模型预测草海自然保护区越冬黑颈鹤精细尺度的生境适宜性分布特征。结果表明: 运用高分辨率遥感生成的环境因子结合MaxEnt模型能较优秀的预测黑颈鹤精细的觅食地与栖息地, AUC值均大于0.948;距耕地距离、距水域距离、水深、距建筑距离是影响黑颈鹤栖息地主要环境因子, 而影响觅食地分布的主要变量则是距耕地距离、距建筑距离、优势植被和水深;研究区黑颈鹤栖息地与觅食地的最适宜区都较小, 面积分别为2.163km2和6.049 km2, 且都分布在草海湖体周围, 呈条带状分布。次适宜地面积也仅为4.241 km2与6.595 km2, 其分布多围绕最适宜区, 分布较为破碎。
| [1] |
Turner R K, van den Bergh J C J M, Söderqvist T, Barendregt A, van der Straaten J, Maltby E, van Ierland E C. Ecological-economic analysis of wetlands: scientific integration for management and policy. Ecological Economics, 2000, 35(1): 7-23. DOI:10.1016/S0921-8009(00)00164-6 |
| [2] |
Lu N, Jia C X, Lloyd H, Sun Y H. Species-specific habitat fragmentation assessment, considering the ecological niche requirements and dispersal capability. Biological Conservation, 2012, 152: 102-109. DOI:10.1016/j.biocon.2012.04.004 |
| [3] |
李筑眉, 李凤山. 黑颈鹤研究. 上海: 上海科技教育出版社, 2005.
|
| [4] |
汪松. 中国濒危动物红皮书-鸟类. 北京: 科学出版社, 1998.
|
| [5] |
冉景丞, 蒙文萍, 粟海军, 张明明. 草海湿地环境问题对黑颈鹤越冬影响及管理对策讨论. 野生动物学报, 2017, 38(1): 35-39. DOI:10.3969/j.issn.1000-0127.2017.01.006 |
| [6] |
Wilsey C B, Lawler J J, Cimprich D A. Performance of habitat suitability models for the endangered black-capped vireo built with remotely-sensed data. Remote Sensing of Environment, 2012, 119: 35-42. DOI:10.1016/j.rse.2011.12.002 |
| [7] |
Kramer-Schadt S, Kaiser T S, Frank K, Wiegand T. Analyzing the effect of stepping stones on target patch colonisation in structured landscapes for Eurasian Lynx. Landscape Ecology, 2011, 26(4): 501-513. DOI:10.1007/s10980-011-9576-4 |
| [8] |
Elith J, Phillips S J, Hastie T, Dudík M, Chee Y E, Yates C J. A statistical explanation of MaxEnt for ecologists. Diversity and Distributions, 2011, 17(1): 43-57. DOI:10.1111/j.1472-4642.2010.00725.x |
| [9] |
Phillips S J, Dudík M. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography, 2008, 31(2): 161-175. DOI:10.1111/j.0906-7590.2008.5203.x |
| [10] |
王学志, 徐卫华, 欧阳志云, 刘建国, 肖燚, 陈佑平, 赵联军, 黄俊忠. 生态位因子分析在大熊猫(Ailuropoda melanoleuca)生境评价中的应用. 生态学报, 2008, 28(2): 821-828. DOI:10.3321/j.issn:1000-0933.2008.02.045 |
| [11] |
欧阳志云, 刘建国, 肖寒, 谭迎春, 张和民. 卧龙自然保护区大熊猫生境评价. 生态学报, 2001, 21(11): 1869-1874. DOI:10.3321/j.issn:1000-0933.2001.11.018 |
| [12] |
Xu W H, Ouyang Z Y, Viña A, Zheng H, Liu J G, Xiao Y. Designing a conservation plan for protecting the habitat for giant pandas in the Qionglai mountain range, China. Diversity and Distributions, 2006, 12(5): 610-619. DOI:10.1111/j.1366-9516.2006.00236.x |
| [13] |
Phillips S J, Anderson R P, Schapire R E. Maximum entropy modeling of species geographic distributions. Ecological Modelling, 2006, 190(3/4): 231-259. |
| [14] |
李凤山, 刘文, 李振吉, 刘强, 吴建东, 洪省, 王汝斌, 史云勋. 贵州草海越冬水鸟数量及其近20年来的变化(英文). 动物学研究, 2014, 35(S1): 85-91. |
| [15] |
彭明春, 王崇云, 钟兴耀. 云南大山包黑颈鹤自然保护区综合科学考察研究. 北京: 科学出版社, 2013.
|
| [16] |
李鹏, 黎达辉, 李振洪, 王厚杰. 黄河三角洲地区GF-3雷达数据与Sentinel-2多光谱数据湿地协同分类研究. 武汉大学学报: 信息科学版, 2019, 44(11): 1641-1649. |
| [17] |
赵英时. 遥感应用分析原理与方法. 北京: 科学出版社, 2003.
|
| [18] |
李苗苗. 植被覆盖度的遥感估算方法研究[D]. 北京: 中国科学院研究生院(遥感应用研究所), 2003.
|
| [19] |
贵州科学院生物研究所. 草海科学考察报告. 贵阳: 贵州人民出版社, 1986.
|
| [20] |
Phillips S J, Dudk M, Schapire R E. Maxent software for modeling species niches and distributions (Version 3. 4.1). https://biodiversityinformatics.amnh.org/open_source/maxent, 2019.5.30.
|
| [21] |
刘振生, 高惠, 滕丽微, 苏云, 王晓勤, 孔芳毅. 基于MAXENT模型的贺兰山岩羊生境适宜性评价. 生态学报, 2013, 33(22): 7243-7249. |
| [22] |
Behdarvand N, Kaboli M, Ahmadi M, Nourani E, Mahini A S, Aghbolaghi M A. Spatial risk model and mitigation implications for wolf-human conflict in a highly modified agroecosystem in western Iran. Biological Conservation, 2014, 177: 156-164. DOI:10.1016/j.biocon.2014.06.024 |
| [23] |
Catry F X, Rego F C, Bação F L, Moreira F. Modeling and mapping wildfire ignition risk in Portugal. International Journal of Wildland Fire, 2009, 18(8): 921-931. DOI:10.1071/WF07123 |
| [24] |
Swets J. Measuring the accuracy of diagnostic systems. Science, 1988, 240(4857): 1285-1293. DOI:10.1126/science.3287615 |
| [25] |
Na X D, Zhou H T, Zang S Y, Wu C S, Li W L, Li M. Maximum Entropy modeling for habitat suitability assessment of Red-crowned crane. Ecological Indicators, 2018, 91: 439-446. DOI:10.1016/j.ecolind.2018.04.013 |
| [26] |
曹铭昌, 孙孝平, 乐志芳, 崔鹏, 龚溪, 徐海根. 基于MAXENT模型的丹顶鹤越冬生境变化分析: 以盐城保护区为例. 生态与农村环境学报, 2016, 32(6): 964-970. |
| [27] |
武大伟, 胡灿实, 张明明, 粟海军. 贵州草海湿地越冬黑颈鹤觅食地与农耕地的空间关系研究. 生态学报, 2021, 41(8): 3238-3247. |
| [28] |
Kong D J, Yang X J, Liu Q, Zhong X Y, Yang J X. Winter habitat selection by the Vulnerable black-necked crane Grus nigricollis in Yunnan, China: implications for determining effective conservation actions. Oryx, 2011, 45(2): 258-264. DOI:10.1017/S0030605310000888 |
| [29] |
Lu G Y, Yang X J. Black-necked Cranes (Grus nigricollis) wintering in Yongshan County, Yunnan and their conservation. Zoological Research, 2014, 35(S1): 143-150. |
| [30] |
王岐山, 李凤山. 中国鹤类研究. 昆明: 云南教育出版社, 2008.
|
| [31] |
Liu W, Jin Y Y, Wu Y J, Zhao C H, He X C, Wang B, Ran J H. Home range and habitat use of breeding black-necked cranes. Animals, 2020, 10(11): 1975. DOI:10.3390/ani10111975 |
| [32] |
孔德军, 杨晓君, 钟兴耀, 道美标, 朱勇. 云南大山包黑颈鹤日间越冬时间分配和活动节律. 动物学研究, 2008, 29(2): 195-202. |
| [33] |
孔德军. 云南大山包黑颈鹤(Grus nigricollis)越冬行为和保护研究[D]. 昆明: 中国科学院昆明动物研究所, 2008.
|
| [34] |
蒋政权, 李凤山, 冉江洪, 刘伟, 赵晨皓, 张波, 李华. 若尔盖湿地保护区黑颈鹤巢期及影响因子. 生态学报, 2017, 37(3): 1027-1034. |
| [35] |
Zhang L X, An B, Shu M L, Yang X J. Nest-site selection, reproductive ecology and shifts within core-use areas of Black-necked Cranes at the northern limit of the Tibetan Plateau. PeerJ, 2017, 5: e2939. DOI:10.7717/peerj.2939 |
| [36] |
方宇, 张学霞, 郭长庆. 若尔盖湿地黑颈鹤生境适宜性. 生态学杂志, 2020, 39(9): 3021-3031. |