 2022, Vol. 42
2022, Vol. 42文章信息
- 刘亚玲, 信忠保, 李宗善, 买尔当·克依木, 闫腾飞
- LIU Yaling, XIN Zhongbao, LI Zongshan, Maierdang Keyimu, YAN Tengfei
- 河北坝上樟子松人工林径向生长及其对气候因素的响应
- Response of radial growth of Pinus sylvestris var. mongolica to climate factors in Bashang area of Hebei Province
- 生态学报. 2022, 42(5): 1830-1840
- Acta Ecologica Sinica. 2022, 42(5): 1830-1840
- http://dx.doi.org/10.5846/stxb202010272750
-
文章历史
- 收稿日期: 2020-01-02
- 网络出版日期: 2021-11-17
- 修订日期: 2021-11-17





2. 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085;
3. 张北县林业局, 张北 076450
2. State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China;
3. Zhangbei Forestry Bureau, Zhangbei 076450, China
随着人类文明的进步和科技的发展, 人为温室气体排放量不断增加, 导致全球温度快速上升, 气候变化成为世界上最关注的科学热点之一[1]。森林作为地球上最大的陆地生态系统, 是地球系统物质循环和能量流动中极为重要的一环[2-3], 在维持全球的生态平衡中扮演着不可或缺的角色。全球气候变暖导致干旱的发生强度和发生频率在逐年增加, 导致部分森林树种出现衰退的现象, 生态系统的结构和功能发生了巨大变化[4-5]。生态弹性是评估树木遭受干扰后的抵抗能力和恢复到初始状态的能力[6]。生态弹性可以准确评估森林对当前和过去气候变化的反应, 有助于预测未来森林对气候变化的适应能力[7]。树木年轮以其定年准确, 分辨率高等优点已广泛应用于树木对干扰和环境胁迫的响应研究[8]。通过研究樟子松的生态弹性, 可以预测樟子松人工林对气候变化的适应性, 为樟子松的经营管理提供理论依据。
河北坝上地区干旱少雨, 植被稀少, 土壤沙化严重, 生态环境脆弱。为改善当地生态环境, 20世纪70年代末营造了以小叶杨为主的人工防护林, 20世纪90年代初补植了以樟子松为主的人工防护林。该区防护林是三北防护林工程的重要组成部分, 同时是京津风沙源的重要生态屏障, 经过40 a的建设其生态效益和经济效益显著[9]。但自2002年开始, 该地区小叶杨防护林逐步出现退化、枯死现象, 危及当地生态平衡, 樟子松防护林的优势逐渐凸显[10], 在河北坝上地区衰退杨树林的更新改造中, 不再使用小叶杨作为主要造林树种, 而普遍选用耐寒耐旱的樟子松。目前我国樟子松的树木年轮学研究主要集中在内蒙古呼伦贝尔沙地和毛乌素沙地、黑龙江大兴安岭和帽儿山、辽宁章古台以及河北塞罕坝地区。大兴安岭和帽儿山等地区樟子松生长主要受温度的影响[11-13]; 呼伦贝尔沙地、毛乌素沙地、章古台及塞罕坝等地区樟子松径向生长主要受降水的限制, 并且章古台地区樟子松出现了生长衰退的现象[14-16]。河北坝上地区樟子松径向生长对气候响应的研究未见报道, 因此我们研究了河北坝上地区树龄最大的樟子松林, 研究其径向生长过程、限制其生长的主要气候因素以及遭遇干旱后樟子松的动态变化, 对评估樟子松在河北坝上地区的生长适宜性具有重要代表性和前瞻性。
樟子松(Pinus sylvestris var. mongolica)原产于中国大兴安岭、红花尔基和呼伦贝尔沙地, 为浅根性树种, 其耐寒耐旱性强, 可在-40--50 ℃的低温中生长, 对水分要求不严格, 并且因其抗逆性强, 寿命长, 有很好的生态和经济价值而被广泛应用于三北防护林的造林工程中。河北坝上地区在20世纪90年代初种植的樟子松人工林目前没有出现衰退林相, 本研究以河北坝上地区樟子松防护林为研究对象, 运用树木年轮气候学原理及方法, 建立樟子松差值年表、BAI年表。旨在揭示樟子松径向生长特征, 明确限制樟子松在该地区生长的主要气候因素, 以及樟子松在河北坝上地区的适宜性。研究结果对于提高森林生态防护作用非常重要, 为未来衰退杨树林的更新改造和樟子松人工林的栽植和经营管理提供科学依据。
1 研究区与研究方法 1.1 研究区概况采样点位于河北省张家口市张北县波罗素林场内(41.336°N, 114.879°E), 是河北坝上樟子松人工林的主要分布区(图 1)。张北县地处内蒙古东南缘, 地势东南高西北低, 东南部为坝上高原区, 西北部为丘陵区, 中部属平原区, 丘陵洼地交错分布。研究区属该县中部平原地区, 海拔1370-1390 m, 相对高差20 m。气候为中温带大陆性季风气候, 四季分明, 夏季炎热多雨, 冬季寒冷干燥。年平均气温为4.0 ℃(1992-2019年), 最冷月(1月)的均温为-14.4 ℃, 最热月(7月)的均温为19.7 ℃, 极端高温为25.3 ℃, 极端低温为-19.7 ℃; 年均降水量为390.1 mm, 6-9月降水量占全年降水量的75%, 年蒸发量1700-1800 mm, 为降水量的4-5倍(图 2)。1992年以来平均气温(0.34 ℃/10a)、最高气温(0.35 ℃/10a)和最低气温(0.21 ℃/10a)均上升迅速; 降水量呈下降趋势, 最大降水量(1995年)533.9 mm, 最小降水量(1997年)245.2 mm; 帕默尔干旱指数下降明显(-0.30/10a); 多年平均相对湿度为56.9%(图 3)。年均日照时数2897.8 h, 年均7级以上大风日数30 d左右。主要土壤类型为栗钙土类。乔木主要包括樟子松(Pinus sylvestris var. mongolica)、小叶杨(Populus simonii)、山杨(Populus davidiana)、云杉(Picea asperata)等; 灌木主要包括柠条(Caragana korshinskii)、沙棘(Hippophae rhamnoides)、小红柳(Salix microstachya)等; 草本植物丰富, 主要分为盐渍化滩地蒿类、低洼地草甸类以及坡梁地禾本类[17]。该地区樟子松为20世纪90年代种植的人工纯林, 造林时使用的是2年生裸苗, 初植密度为1300株/hm2, 后期抚育管理措施缺乏, 现存密度为950株/hm2, 行距为4 m, 株距为3 m, 研究区内樟子松树龄基本一致。

|
| 图 1 河北坝上地区树木年轮采样点及气象站分布图 Fig. 1 Location map of tree-ring sample site and meteorological station in Northwestern of Hebei, China |

|
| 图 2 1992-2019年河北坝上地区温度降水季节分布 Fig. 2 Seasonal distribution of temperature and precipitation in Northwestern of Hebei, China (1992-2019 a) |

|
| 图 3 河北坝上地区气候因素年际变化趋势(1992-2019年) Fig. 3 Climatic changing trend of Northwestern of Hebei, China from 1992 to 2019 |
在河北坝上地区波罗素林场内选取40株长势良好、树龄较大、受病虫干扰小的樟子松, 在胸高处(1.3 m)用生长锥沿东西方向钻取树芯, 每棵树钻取2-3个树芯, 共取样80个。采集的树芯根据国际上树轮样品标准处理过程进行预处理。在实验室内将采集的树芯固定风干[18], 用不同粗细的干砂纸(400目、800目和1200目)依次打磨到样芯光滑且亮, 并且在电子显微镜下能看清樟子松的年轮。利用骨架图法交叉定年后, 用年轮分析仪Win DENDROTM(Regent instrument inc. Canada)测量樟子松树轮的原始宽度, 其精度为0.001 mm。用COFECHA程序对交叉定年和测量结果进行检验, 剔除相关性差的样芯, 最后余下57根样芯进行年表建立[19]。
由于样本树龄较短, 研究主要集中在树木生长的高频变化上, 故应去除树木随年龄增长的生长趋势和其他非一致性扰动导致的非气候信号, 而突出其中的气候信号[20], 利用ARSTAN程序建立年表, 去趋势方法为负指数函数, 该方法可减少树间竞争、地形、地貌等因素对年表指数拟合的影响, 最终得到樟子松标准年表(Standard chronology, STD)、自回归年表(Arstan chronology, ARS)和差值年表(Residual chronology, RES)。本研究选用RES进行树木生长与气候间的相关分析, RES是在STD的基础上, 考虑到森林内树与树的竞争以及可能存在的人类活动导致的树轮宽度序列的低频变化, 以及时间序列的自回归模式对STD进行拟合并再次标准化, 去掉树木个体特有的前期生理条件对后期生长造成的连续性影响而建立的一种年表, 它只含有群体共有的高频变化。
1.3 胸高断面积增量当树轮宽度相对稳定或变窄时, 树木生长量可能继续增大[21], 为了保留气候条件对树木生长影响的信息, 本文使用BAI来评估研究区内樟子松人工林的生长速率和生长趋势。树木胸高断面积增量:
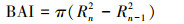
|
式中, Rn是n年的树盘半径; Rn-1是n-1年的树轮半径。
1.4 气象数据获取本研究采用的气候要素数据来源于张北气象站(114.42°E, 41.09°N, 海拔为1393.3 m), 时间跨度为1992-2019年。气候要素为月均温度、月均最低温度、月均最高温度、月均相对湿度和降水量, 以上数据从中国气象数据网(http://data.cma.cn/)获取。由于温度和降水等综合作用对土壤有效含水量的影响明显, 本研究选用帕默尔干旱指数(PDSI)反映樟子松生长与土壤水分的关系, 其数据来源于荷兰皇家气象局服务网(http://climexp.knmi.nl/start.cgi)中的PDSI网格点数据集[22](114.50°-115.00°E, 41.00°-41.50°N), 时间跨度为1992-2017年。考虑到树木生长的连贯性, 当年的生长必定受到前一年气候的影响, 在进行气候响应分析时选用上一年6月到当年10月共17个月的气象数据。
1.5 数据处理该地区位于半干旱地区, 干旱缺水是其主要气候特征, 在干旱年份樟子松生长受到严重影响, 樟子松径向生长显著下降, 结合樟子松差值年表和BAI年表与PDSI年际波动数据, 研究樟子松生长遭遇干旱时其生态弹性。选定樟子松3个生长低点年份2000年(PDSI=-1.92)、2007年(PDSI=-3.34)和2014年(PDSI=-1.54)来研究樟子松的抵抗力、恢复力和弹性力。
利用Lloret等[23]的公式来进行樟子松抵抗力、恢复力和弹性力的计算:
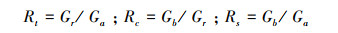
|
式中, Rt、Rc和Rs分别表示树木的抵抗力、恢复力和弹性力; Gr表示生长显著下降时数值; Ga和Gb分别表示生长显著下降前3年和后3年的平均值。Rt>0.75说明树木对干旱具有较高的抵抗力; Rc>1.25说明发生干旱事件后树木仍有较好的恢复力。
本文利用Excel 2013软件分析河北坝上地区樟子松的径向生长过程, 揭示该地区樟子松的生长状况。使用Dendroclim 2002国际树木年轮程序[24]分析年表与各气候数据间的相关性, 并用Origin 2018绘制樟子松差值年表、BAI年表以及相关图表的制作。
2 研究结果 2.1 樟子松年表统计特征本研究建立了樟子松标准年表和差值年表, 年表质量的高低决定着研究结果是否准确, 利用标准差、平均敏感度、序列间相关系数、信噪比、样本总体代表性等来评价年表质量(表 1)。标准差反映年表的上下波动, 标准差越大表明年表所包含的气候信息越多, 对气候越敏感[25], 樟子松标准年表和差值年表的标准差分别为0.358和0.308。一介自相关系数反映上一年气候状况对当年年轮宽度生长的影响强弱, 其值分别为0.295和0.063, 表明樟子松径向生长受上年气候条件的影响[26]。樟子松序列间相关系数分别为0.579和0.607, 表示不同树芯间的年轮宽度变化具有较好的一致性, 数据质量较好[20]。平均敏感度分别为0.304和0.324, 信噪比分别为12.364和13.906, 主要代表气候的短期变化或高频变化, 平均敏感度大于0.15以及信噪比大于4代表年表质量较高[27-28]。样本总体代表性分别为0.925和0.933, 是指所采集样芯对整个区域的代表程度, 通常样本总体代表性大于0.85表明年表质量较高[29]。综上所述, 樟子松差值年表质量较好, 以下运用差值年表来进行气候响应分析。樟子松差值年表的起始年份样本量大于5根, 可以较好地反映径向生长年际波动(图 4)。
| 年表类型 Type |
标准差 SD |
平均敏感度 MS |
一阶自相关 AR1 |
样芯间相关系数 Rbt |
样本总体代表性 EPS |
信噪比 SNR |
| 标准年表STD | 0.358 | 0.304 | 0.295 | 0.579 | 0.925 | 12.364 |
| 差值年表RES | 0.308 | 0.324 | 0.063 | 0.607 | 0.933 | 13.906 |
| STD: 标准年表Standard chronology; RES: 差值年表Residual chronology; MS: 平均敏感度Mean sensitivity; SD: 标准差Standard deviation; AR1:一阶自相关First-order autocorrelation; Rbt: 样芯间相关系数Mean inter-series correlation; SNR: 信噪比Signal-to-noise ratio; EPS: 样本总体代表性Express population siginal | ||||||

|
| 图 4 河北坝上地区樟子松差值年表 Fig. 4 Residual chronology of Pinus sylvestris var. mongolica forest in Northwest of Hebei, China |
河北坝上地区樟子松生长随树龄增加呈现在波动中下降趋势, 在各个生长阶段的径向生长速率差距显著(图 5), 在1-10 a生长速率较快为(4.81±0.63)mm/a; 11-20 a樟子松生长速率逐渐下降, 平均生长速率为(2.95±0.87)mm/a, 相对1-10 a生长速率下降了38.7%;21-28 a樟子松径向生长处于平稳状态, 其平均生长速率为(2.20±0.43)mm/a, 相对1-10 a生长速率下降54.3%。

|
| 图 5 河北坝上地区樟子松树轮宽度径向生长特征、树轮宽度和胸高断面积增量(BAI)年际变化 Fig. 5 Ring-width and BAI variation of Pinus sylvestris var. mongolica of Hebei, China |
河北坝上地区樟子松树轮宽度年际变化显示(图 5), 樟子松树轮宽度在1992-2000年期间为生长平稳期, 平均生长速率为(4.49±1.15)mm/a; 在2001-2010年期间呈现下降趋势, 平均生长速率为(3.62±1.39)mm/a, 较前期生长速率下降了19.3%;2011-2019年期间樟子松生长呈平稳特征, 平均生长速率为(2.21±0.68)mm/a, 较前期生长速率下降了50.8%。胸高断面积生长量(BAI)是每年树轮增加的面积, 相比与年轮宽度, 它能够更好地反映树木的生长速率和生长趋势[30]。樟子松生长期间BAI年际波动较大(图 5), 1992-2000年樟子松BAI一直呈上升趋势, 平均生长速率为(5.09±2.26)cm2/a; 2001-2019年期间呈平稳特征, 平均生长率为(10.46±0.67)cm2/a, 较前期生长速率增加了105.5%, 表明樟子松表现出稳定、旺盛的生产力状态, 尚未出现生产力衰退现象。
2.3 樟子松RES与气候因素的关系河北坝上地区樟子松径向生长受温度和降水两者共同的影响。樟子松RES与温度因素多呈负相关, 樟子松径向生长与上一年9、10、12月及当年7月份的平均温度和最高温度呈显著负相关, 其中最高温度对樟子松生长影响更为显著(P < 0.05, 图 6)。樟子松径向生长与降水和相对湿度多呈正相关, 樟子松树轮宽度与上一年9月及当年5、7月份的降水和相对湿度呈显著正相关(P < 0.05, 图 7)。帕默尔干旱指数在干旱区的研究中广泛应用, 它是根据降水、温度以及土壤含水量数据进行计算, 代表土壤水分的有效状况, 其值越小表明有效土壤水分越低。樟子松径向生长与帕默尔干旱指数多呈正相关, 其中与上一年9月-当年10月份的帕默尔干旱指数呈显著正相关, 该地区樟子松遭受干旱胁迫(P < 0.05, 图 7)。

|
| 图 6 河北坝上樟子松差值年表与温度因素的相关关系 Fig. 6 Correlation coefficients of Pinus sylvestris var. mongolica chronology between temperature factors *代表P < 0.05, 月份前的“-”表示上一年, 如-6表示上一年6月份 |

|
| 图 7 樟子松差值年表与降水、相对湿度和PDSI的相关关系 Fig. 7 Correlation coefficients of Pinus sylvestris var. mongolica chronology between precipitation relative humidity and PDSI *代表P < 0.05, 月份前的“-”表示上一年, 如-6表示上一年6月份 |
该地区气候暖干化趋势明显, 樟子松生长极易遭受严重干旱胁迫, 在干旱缺水年份樟子松生长受到严重影响, 结合樟子松差值年表和BAI年表与PDSI年际波动数据, 研究樟子松生长显著下降时其生态弹性。选出2000年、2007年和2014年来分析樟子松生长显著下降时的抵抗力、恢复力和弹性力(图 8)。通过樟子松差值年表研究樟子松的生态弹性, 发现樟子松随树龄增加, 抵抗力由0.82下降到0.55, 在2000年樟子松具有较高的抵抗力; 樟子松的恢复力呈上升趋势, 2014年达到了1.91, 恢复力较强; 樟子松的弹性力呈现稳定特征。通过樟子松BAI年表研究樟子松的生态弹性, 发现樟子松随树龄增加, 抵抗力同样在下降, 由0.93下降到0.52, 仅在2000年具有较高的抵抗力; 樟子松的恢复力呈上升趋势, 2014年达到了1.72;樟子松的弹性力较为稳定(图 9)。

|
| 图 8 樟子松差值年表和BAI年表与PDSI的关系 Fig. 8 Correlation of Pinus sylvestris var. mongolica chronologies between PDSI |

|
| 图 9 樟子松的抵抗力、恢复力和弹性力 Fig. 9 Resistance, resilience and recovery of Pinus sylvestris var. mongolica |
樟子松在2000、2007和2014年遭遇干旱时生长下降严重。樟子松树轮宽度在2000年时达到生长低点, 2000年当地干旱指数达到-1.92, 处于轻微干旱等级, 干旱胁迫导致樟子松径向生长下降。干旱胁迫对树木年轮宽度影响显著, 树木在面对严重的干旱时会表现出应激症状, 如叶片过早的发黄、脱落, 树木处于较脆弱的状态, 碳水化合物储量变少, 树木的高度、年轮宽度等因此受到限制[31]。2007年和2014年均为干旱年, 尤其2007年达到严重干旱程度, 樟子松树轮宽度在这两个时期达到较窄水平。2000年树轮宽度有所上升, 在第二年或后几年出现显著的下降, 表明樟子松对气候的响应具有明显的“滞后性”。树木在受到极端干旱胁迫时, 当年的响应可能不会十分明显, 在极端干旱的次年或之后几年形成窄轮[31]。《中华人民共和国林业行业标准主要树种龄级与龄组划分》(LY/T2908-2017)中将樟子松的生长划分为五个阶段: 1-30 a为幼龄期, 31-50 a为中龄期, 51-60 a为近熟期, 61-80 a为成熟期, 80 a之后为过熟期。河北坝上地区樟子松树轮宽度随树龄的增大, 径向生长速率呈下降趋势, 在21-28 a时处于平稳期, 樟子松的生长阶段较国家行标的生长阶段划分提前, 当地樟子松可能出现衰退的现象。
干旱胁迫影响樟子松BAI增长速率, 樟子松BAI在1992-2000年期间增长迅速, 在2007年遭遇严重干旱事件后其增长速率下降至近几年最低值, 其后樟子松BAI增长逐渐恢复, 说明樟子松遭遇干旱后有一定的恢复机能, 可以较好的适应河北坝上地区干旱的气候条件。当地樟子松的树龄较小, 仅有20-30 a, 理论上樟子松的最大树龄可达350年左右[32], 当地樟子松BAI年际增长量变化趋势不能完全反映当地樟子松是否出现衰退现象, 还需要对当地樟子松进行连续观测。
3.2 樟子松差值年表与气候因子关系特征河北坝上地区樟子松年内树干径向生长呈现季节性规律, 主要划分为4个阶段: 4月初到5月中旬的春季萌芽期、5月中旬到7月中旬的夏季生长期、7月中旬到10月中旬的秋冬交替期以及10月中旬到下年4月初的冬季休眠期[33]。河北坝上地区樟子松径向生长主要与上一年生长季末期(9、10月)、休眠期(12月)和当年生长季(7月)的月平均最高温度呈显著负相关, 高温显著抑制樟子松的径向生长。上年生长季末期和休眠期温度越高, 导致蒸发作用增强, 水分减少, 在第二年进入生长季后没有充足的水分, 抑制樟子松的光合速率, 樟子松径向生长受到限制[34]。生长季内温度过高, 树木蒸腾作用和呼吸作用均加强, 光合作用减弱, 消耗掉过多的养分, 樟子松径向生长速率减缓, 易形成窄轮[35-36]。
樟子松差值年表与上年生长季末(9月)及当年生长季(5、7月)降水呈显著正相关, 其中受上年降水的影响更为显著, 说明樟子松径向生长与降水存在“滞后效应”, 上年生长季后期降水可以为根系层补充充足的水分, 为促进当年树木的生长起着重要作用[37]。5、7月樟子松进入生长期, 气温逐渐增加, 蒸腾作用加强, 导致土壤水分不足, 树木生长受到限制, 有效的降水可以增加土壤水分, 为樟子松的正常生长提供必要的水分条件。相对湿度是受温度和降水共同调控的气候因子[38], 影响樟子松的生长, 樟子松生长主要受当年生长季(5、7、8月)的影响。植物生长空气湿度在50%-80%为宜, 而河北坝上地区相对湿度在56%左右, 属于半干旱气候, 当树木周围环境中的相对湿度增高时, 会促进植物气孔张开, CO2进入叶肉细胞, 促进植物的光合作用, 有助于樟子松的径向生长。
PDSI反映干旱严重程度, 具有较好时空可比较性, 不但综合了水分亏缺量和持续时间因子对干旱程度的影响, 并充分考虑了干旱持续时间, 是评估干旱程度的较好指标[39-40]。河北坝上地区樟子松径向生长与上一年9月至当年10月PDSI呈显著正相关。干旱程度增大会抑制樟子松的生长, 当樟子松遭遇干旱胁迫时, 叶片气孔关闭会阻碍树木与外界进行气体交换, 减缓光合作用, 树木体内碳水化合物缺乏, 呼吸作用和新陈代谢会消耗体内营养物质, 使其处于饥饿的状态, 极易形成窄轮[41]。
3.3 河北坝上樟子松的生态弹性本研究中随樟子松树龄的增加, 樟子松的抵抗力呈现下降趋势, 而恢复力出现上升趋势, 在遭遇干旱时树木的恢复力和抵抗力存在一种平衡关系, 抵抗力较弱的区域在干旱后会有较强的恢复力[42]。近30年来, 河北坝上地区温度上升明显, 有效土壤水分利用率呈降低的趋势(图 3), 该地区易发生干旱事件, 如果水分可用性的变化超过阈值, 光合作用减弱, 导致碳水化合物减少, 抵抗力减弱, 树木无法抵抗干旱将面临生长下降的风险, 在抵抗力弱的地区干旱极易引发树木生长的衰退[43]。此外, 如果发生持续的干旱, 它们可能持续影响树木的生长, 导致树木生长下降的风险增加[44]。当抵抗力减小到一定程度后, 其恢复力可能会减小, 在评估樟子松在河北坝上生长适应性时要综合评估抵抗力和恢复力以及影响它们的因素, 充分考虑降水和土壤水分对樟子松的影响[45]。
4 结论河北坝上地区樟子松差值年表有效长度为1992-2019年, 根据其生长过程将其划分为3个阶段, 1-10 a的快速生长期, 11-20 a的生长下降期以及21-28 a的生长平稳期。樟子松胸高断面积增量在2000年后呈现平稳特征, 樟子松对河北坝上地区的干旱环境有一定的适应性。樟子松径向生长主要与平均温度和最高温度呈显著负相关, 其中最高温度对其影响最显著。樟子松径向生长主要与上一年9月和当年生长季(5、7月)的降水显著正相关, 适量的降水促进河北坝上地区樟子松的生长。干旱胁迫会抑制樟子松的径向生长, 干旱指数越小说明干旱程度越严重, 樟子松与多月份(上一年9月-当年10月)的干旱指数呈显著正相关。随着气候变暖, 当地暖干化特征显著, 樟子松的抵抗力下降, 恢复力上升, 若持续发生干旱, 樟子松生长下降的风险增加, 易引发树木的衰退。
| [1] |
IPCC. Summary for policymakers//Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt K B, Tignor M, Miller H L, eds. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2013.
|
| [2] |
Olsson P O, Jönsson A M, Eklundh L. A new invasive insect in Sweden-Physokermes inopinatus: tracing forest damage with satellite based remote sensing. Forest Ecology and Management, 2012, 285: 29-37. DOI:10.1016/j.foreco.2012.08.003 |
| [3] |
陆旭. 吉林省人工林樟子松径向生长及其空间气候响应研究[D]. 沈阳: 沈阳农业大学, 2018.
|
| [4] |
Goswami B N, Venugopal V, Sengupta D, Madhusoodanan M S, Xavier P K. Increasing trend of extreme rain events over India in a warming environment. Science, 2006, 314(5804): 1442-1445. DOI:10.1126/science.1132027 |
| [5] |
Zhu L J, Cooper D J, Yang J W, Zhang X, Wang X C. Rapid warming induces the contrasting growth of Yezo spruce (Picea jezoensis var. microsperma) at two elevation gradient sites of northeast China. Dendrochronologia, 2018, 50: 52-63. DOI:10.1016/j.dendro.2018.05.002 |
| [6] |
Thompson I, Mackey B, McNulty S, Mosseler A. Forest resilience, biodiversity, and climate change: a synthesis of the biodiversity/resilience/stability relationship in forest ecosystems. Canada: Secretariat of the Convention on Biological Diversity, Montreal, 2009.
|
| [7] |
申佳艳, 李帅锋, 黄小波, 王绍武, 苏建荣. 金沙江流域不同海拔处云南松生态弹性及生长衰退过程. 林业科学, 2020, 56(6): 1-11. |
| [8] |
Helman D, Lensky I M, Yakir D, Osem Y. Forests growing under dry conditions have higher hydrological resilience to drought than do more humid forests. Global Change Biology, 2017, 23(7): 2801-2817. DOI:10.1111/gcb.13551 |
| [9] |
郑春雅, 许中旗, 马长明, 孙守家, 闫腾飞. 冀西北坝上地区杨树防护林退化的影响因素. 林业资源管理, 2018(1): 9-15, 147-147. |
| [10] |
杜海峰. 河北坝上杨树退化的原因及改造技术求索. 环境与发展, 2019, 31(7): 195-195, 197-197. |
| [11] |
王晓春, 宋来萍, 张远东. 大兴安岭北部樟子松树木生长与气候因子的关系. 植物生态学报, 2011, 35(3): 294-302. |
| [12] |
徐静, 郭滨德, 孙洪志. 帽儿山地区不同种源樟子松树轮对气候因子的响应. 林业科学研究, 2016, 29(4): 581-586. DOI:10.3969/j.issn.1001-1498.2016.04.018 |
| [13] |
李俊霞, 白学平, 张先亮, 常永兴, 陆旭, 赵学鹏, 陈振举. 大兴安岭林区南、北部天然樟子松生长对气候变化的响应差异. 生态学报, 2017, 37(21): 7232-7241. |
| [14] |
尚建勋, 时忠杰, 高吉喜, 徐丽宏, 吕世海, 冯朝阳, 王鲁秀. 呼伦贝尔沙地樟子松年轮生长对气候变化的响应. 生态学报, 2012, 32(4): 73-80. DOI:10.3969/j.issn.1673-1182.2012.04.017 |
| [15] |
李露露, 李丽光, 陈振举, 周永斌, 张先亮, 白学平, 常永兴, 肖建强. 辽宁省人工林樟子松径向生长对水热梯度变化的响应. 生态学报, 2015, 35(13): 4508-4517. |
| [16] |
雷帅, 张劲松, 孟平, 何子兵, 程顺, 孙守家. 中国北部不同地点樟子松人工林径向生长对气候响应的差异. 生态学报, 2020, 40(13): 4479-4492. |
| [17] |
苏王新, 李卓, 陈书琴, 李海防. 河北坝上地区植被覆盖演化特征及其风险评估. 干旱区研究, 2018, 35(3): 686-694. |
| [18] |
Stokes M A, Smiley T L. An Introduction to Tree-Ring Dating. Chicago: University of Chicago Press, 1968.
|
| [19] |
Holmes R L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 1983, 43: 51-67. |
| [20] |
Brubaker L B. Tree rings and climate. Ecology, 1977, 58(6): 1400-1401. DOI:10.2307/1935092 |
| [21] |
Cook E R. A Time Series Analysis Approach to Tree-Ring Standardization[D]. Tucson, AZ, USA: The University of Arizona, 1985.
|
| [22] |
Harris I, Jones P D, Osborn T J, Lister D H. Updated high-resolution grids of monthly climatic observations-the CRU TS3.10 Dataset. International Journal of Climatology, 2014, 34(3): 623-642. DOI:10.1002/joc.3711 |
| [23] |
Lloret F, Keeling E G, Sala A. Components of tree resilience: effects of successive low-growth episodes in old ponderosa pine forests. Oikos, 2011, 120(12): 1909-1920. DOI:10.1111/j.1600-0706.2011.19372.x |
| [24] |
Biondi F, Waikul K. DENDROCLIM2002: a C++ program for statistical calibration of climate signals in tree-ring chronologies. Computers & Geosciences, 2004, 30(3): 303-311. |
| [25] |
Fritts H C. Tree Rings and Climate. London: Academic Press, 1976.
|
| [26] |
孙铂, 雷莺, 郭玲霞, 刘宇峰, 王倩. 秦岭通天河地区树轮年表的建立及特征分析. 咸阳师范学院学报, 2019, 34(4): 58-60. DOI:10.3969/j.issn.1672-2914.2019.04.015 |
| [27] |
Savva Y, Oleksyn J, Reich P B, Tjoelker M G, Vaganov E A, Modrzynski J. Interannual growth response of Norway spruce to climate along an altitudinal gradient in the Tatra Mountains, Poland. Trees, 2006, 20(6): 735-746. DOI:10.1007/s00468-006-0088-9 |
| [28] |
Smith K T. An organismal view of dendrochronology. Dendrochronologia, 2008, 26(3): 185-193. DOI:10.1016/j.dendro.2008.06.002 |
| [29] |
Wigley T M L, Briffa K R, Jones P D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. Journal of Applied Meteorology and Climatology, 1984, 23(2): 201-213. DOI:10.1175/1520-0450(1984)023<0201:OTAVOC>2.0.CO;2 |
| [30] |
刘敏. 中国东北红松生长对气候变化的响应及其动态研究[D]. 哈尔滨: 东北林业大学, 2017.
|
| [31] |
梁慧敏, 魏江生, 贺敏, 周梅, 赵鹏武. 大兴安岭南段华北落叶松人工林径向生长对气候变化的响应. 温带林业研究, 2019, 2(3): 31-36, 62-62. DOI:10.3969/j.issn.2096-4900.2019.03.006 |
| [32] |
宋来萍. 大兴安岭樟子松年轮气候响应及年龄影响探讨[D]. 哈尔滨: 东北林业大学, 2011.
|
| [33] |
季倩雯, 郑成洋, 张磊, 曾发旭. 河北塞罕坝樟子松径向生长动态变化及其与气象因子的关系. 植物生态学报, 2020, 44(3): 257-265. |
| [34] |
常永兴, 陈振举, 张先亮, 白学平, 赵学鹏, 李俊霞, 陆旭. 气候变暖下大兴安岭落叶松径向生长对温度的响应. 植物生态学报, 2017, 41(3): 279-289. |
| [35] |
王婷, 于丹, 李江风, 马克平. 树木年轮宽度与气候变化关系研究进展. 植物生态学报, 2003, 27(1): 23-33. DOI:10.3321/j.issn:1005-264X.2003.01.004 |
| [36] |
贺敏, 魏江生, 石亮, 周梅, 赵鹏武. 大兴安岭南段山杨径向生长和死亡对区域气候变化的响应. 生态学杂志, 2018, 37(11): 3237-3244. |
| [37] |
韦景树, 李宗善, 焦磊, 陈维梁, 伍星, 王晓春, 王帅. 黄土高原羊圈沟小流域人工物种和自然物种径向生长对气候变化的响应差异. 生态学报, 2018, 38(22): 8040-8050. |
| [38] |
韩金生, 赵慧颖, 朱良军, 张远东, 李宗善, 王晓春. 小兴安岭蒙古栎和黄菠萝径向生长对气候变化的响应比较. 应用生态学报, 2019, 30(7): 2218-2230. |
| [39] |
Alley W M. The palmer drought severity index: limitations and assumptions. Journal of Applied Meteorology and Climatology, 1984, 23(7): 1100-1109. DOI:10.1175/1520-0450(1984)023<1100:TPDSIL>2.0.CO;2 |
| [40] |
卫捷, 陶诗言, 张庆云. Palmer干旱指数在华北干旱分析中的应用. 地理学报, 2003, 58(S1): 91-99. |
| [41] |
McDowell N, Pockman W T, Allen C D, Breshears D D, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams D G, Yepez E A. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought?. New Phytologist, 2008, 178(4): 719-739. DOI:10.1111/j.1469-8137.2008.02436.x |
| [42] |
Gazol A, Camarero J J, Vicente-Serrano S M, Sánchez-Salguero R, Gutiérrez E, de Luis M, Sangüesa-Barreda G, Novak K, Rozas V, Tíscar P A, Linares J C, Martín-Hernández N, del Castillo E M, Ribas M, García-González I, Silla F, Camisón A, Génova M, Olano J M, Longares L A, Hevia A, Tomás-Burguera M, Galván J D. Forest resilience to drought varies across biomes. Global Change Biology, 2018, 24(5): 2143-2158. DOI:10.1111/gcb.14082 |
| [43] |
Xu C Y, Liu H Y, Anenkhonov O A, Korolyuk A Y, Sandanov D V, Balsanova L D, Naidanov B B, Wu X C. Long-term forest resilience to climate change indicated by mortality, regeneration, and growth in semiarid southern Siberia. Global Change Biology, 2017, 23(6): 2370-2382. DOI:10.1111/gcb.13582 |
| [44] |
Allen C R, Angeler D G, Cumming G S, Folke C, Twidwell D, Uden D R. Quantifying spatial resilience. Journal of Applied Ecology, 2016, 53(3): 625-635. DOI:10.1111/1365-2664.12634 |
| [45] |
Fang O Y, Zhang Q B. Tree resilience to drought increases in the Tibetan Plateau. Global Change Biology, 2019, 25(1): 245-253. DOI:10.1111/gcb.14470 |