 2022, Vol. 42
2022, Vol. 42文章信息
- 杨镒如, 张茗珊, 张凌楠, 路强强, 洪艺雪, 刘晓宏
- YANG Yiru, ZHANG Mingshan, ZHANG Lingnan, LU Qiangqiang, HONG Yixue, LIU Xiaohong
- 秦岭中西部油松径向生长对气候因子的响应差异研究
- Different responses of radial growth of Pinus tabuliformis to climate in the middle and western Qinling Mountains
- 生态学报. 2022, 42(4): 1474-1486
- Acta Ecologica Sinica. 2022, 42(4): 1474-1486
- http://dx.doi.org/10.5846/stxb202102060392
-
文章历史
- 收稿日期: 2021-02-06
- 网络出版日期: 2021-11-04





2. 陕西省西安植物园(陕西省植物研究所), 西安 710061;
3. 中国科学院西北生态环境资源研究院冰冻圈科学国家重点实验室, 兰州 730000
2. Xi'an Botanical Garden of Shaanxi Province (Institute of Botany of Shaanxi Province), Xi'an 710061, China;
3. State Key Laboratory of Cryospheric Sciences, Northwest Institute of Eco-Environmental Resources, Chinese Academy of Sciences, Lanzhou 730000, China
自20世纪80年代以来, 全球变暖已成不争的事实[1]。有研究表明, 全球气候变暖会导致干旱事件频发[2-3], 威胁树木的生长, 甚至直接或间接引发森林死亡[4]。目前不仅在干旱、半干旱区, 在温带甚至是热带地区也出现了干旱加剧导致森林树木生长衰退和死亡的现象[5-7]。因此, 分析气候变化背景下树木径向生长对气候因子及干旱事件的响应机制, 对准确预测未来气候变暖进一步加剧对树木生长的影响至关重要。秦岭是我国重要的南北地理分界线和气候分界线[8], 也是树木生长对气候变化响应的敏感区和脆弱区[9]。在气候暖干化背景下, 秦岭地区的气候要素变化及其可能产生的影响备受学者关注[10-11]。树木年轮因其连续性好、定年准确、分辨率高、容易获取等特点, 已被广泛用于指示树木的径向生长[12]。油松(Pinus tabuliformis)作为一种在秦岭中低海拔地区分布广泛的针叶树, 因其对气候因子响应敏感, 其年轮数据已被广泛用于树木年轮学的研究中。例如华亚伟等利用秦岭中部油松树轮样芯重建了陕西省镇安县过去近165年3-4月平均最高气温[13], 宋慧明等利用油松树轮样芯重建了秦岭北坡石门山地区过去近400年5-7月平均温度[14]。
秦岭地区地形复杂, 南北气候特征差异明显, 在该地区已开展的树轮生态学研究中发现, 限制树木径向生长的关键气候因子具有明显的空间差异性。例如Shi等在秦岭东部伏牛山的研究发现, 油松径向生长与当年5-6月平均最高气温显著负相关, 与当年5月的降水显著正相关[15];Sun等在秦岭石门山和华山对油松及华山松(Pinus armandii)的研究发现, 油松生长主要受生长季平均最高气温的限制, 华山松主要受4-6月降水的限制[16]。Qin等在秦岭中部太白山对太白红杉(Larix chinensis)的研究发现, 树木径向生长与当年6月平均气温以及前一年6月和当年4月降水显著相关, 且与降水的相关性强于与温度的相关性[17]。有研究在综述秦岭地区已有的树木年轮学研究后认为, 在秦岭以北的地区气候偏干旱, 水分是限制树木径向生长的主要因子, 同时高温引起的土壤水分胁迫也对树木生长有显著抑制作用, 因此树木多表现为与降水因子显著正相关, 与气温因子显著负相关;而在秦岭分水岭和南坡地区, 由于东亚夏季风能够带来充沛的降水, 因而树木径向生长多与气温因子相关性显著, 而与降水因子关系不明显[18]。
人们在关注气候变暖导致气温升高对树木生长产生影响的同时, 也更加关注干旱事件对树木生长的影响, 因为相较于气温逐渐升高带来的影响, 干旱事件的发生对陆地植被光合作用和生长状况的影响更加明显[19]。干旱事件不仅使林冠光合作用减弱, 并且会对树木的生长造成长期、滞后的影响[20]。抵抗力、恢复力等指标已被广泛用于评估树木应对干旱事件能力的研究当中, 并且这些指标会随分布区域、气候环境的差异而有所不同[21]。例如Antonio等对地中海白松(Pinus halepensis)的研究发现, 在西班牙东部干旱地区的树木比在湿润地区的树木具有更差的抵抗力, 且该研究区树木的抵抗力随干旱程度的增加而下降[22];而Cavin等对欧洲山毛榉(Fagus sylvatica)的研究发现, 生长在该物种分布区干旱边缘的树木表现出更强的抵抗力[23]。在全球气候变化背景下, 确定不同干湿区域树木对干旱事件的抵抗力和恢复力, 明晰其对干旱事件的响应策略, 对于制定正确的森林经营方法以进行森林管理和保护具有重要意义。
本研究以秦岭中西部南北坡的油松年轮样品为研究对象, 基于树轮宽度, 建立标准年表(STD), 计算树木径向生长抵抗力、恢复力、生长减少量等指标, 并结合气温、降水及帕默尔干旱指数(PDSI)等气候资料, 旨在: (1)分析秦岭中西部南北坡油松径向生长对气候因子的响应差异;(2)探究在不同干湿环境下油松应对干旱事件能力的差异, 为评估全球气候变化背景下秦岭地区油松群落动态变化提供科学依据。
1 材料与方法 1.1 研究区概况秦岭位于中国东部季风区, 海拔大致在1500-3000 m之间, 北坡山势陡峻, 南坡较缓, 是暖温带和北亚热带的分界线[24], 夏季温暖湿润, 冬季寒冷干旱。由于秦岭山脉对气流的阻隔效应明显, 致使其南北气候差异较大, 年降水量整体表现为南坡多于北坡, 年均温呈现由东南向西北递减、南麓明显高于北麓的特征[24]。本文涉及的4个采样点分别位于秦岭中部和西部的南北坡两侧(图 1), 石门山(SMS)位于秦岭西部北坡, 张家庄(ZJZ)位于秦岭西部南坡, 南五台(NWT)位于秦岭中部北坡, 木王山(MWS)位于秦岭中部南坡。秦岭具有明显的植被垂直更替, 以MWS为例, 海拔1500 m左右的植被林以油松、栎类为优势树种, 海拔1500 m以上以秦岭冷杉(Abies chensiensis)、高山杜鹃(Rhododendron lapponicum)、铁杉(Tsuga chinensis)为主要优势树种, 混交有华山松、山杨(Populus davidiana)、漆树(Toxicodendron vernicifluum)、云杉(Picea asperata)、白桦(Betula platyphylla)等树种。油松是秦岭地区分布广泛的优势针叶树种, 且年轮样芯具有分辨率高、对主控因子响应积极等特点, 因此选其作为本文的研究对象。

|
| 图 1 树轮采样点示意图 Fig. 1 Tree-ring sampling sites map |
研究区气候受到我国东南季风和西南季风的影响。4个采样点附近气象站数据显示年平均降水量在521-839 mm之间, 季节分布不均, 且主要集中在7-9月。天水、凤县、长安以及镇安气象站数据显示, 在1960-2017年间, 4个气象站点最热月(7月)平均气温在21-26.7℃之间, 最冷月(1月)平均气温在-4.2-3℃之间(图 2)。通过对4个站点1991-2000年气温数据的趋势检验发现, 凤县年平均气温和5-7月均温呈显著上升趋势, 天水和镇安年平均气温呈显著上升趋势(图 3)。

|
| 图 2 研究区1960-2017月平均最高气温、平均气温、平均最低气温及总降水 Fig. 2 Monthly mean maximum temperature, mean temperature, mean minimum temperature and mean precipitation in the study area from 1960 to 2017 |

|
| 图 3 研究区1960-2017年平均最高气温、平均气温、平均最低气温及总降水 Fig. 3 Changes of annual mean maximum temperature, mean temperature, mean minimum temperature and total precipitation in the study area from 1960 to 2017 |
2017及2018年7月分别在秦岭中西部南北坡的石门山(SMS)、张家庄(ZJZ)、南五台(NWT)、木王山(MWS)共4处设立采样点开展油松树芯样品采集工作(表 1)。样点选取在具有一定倾角的坡地或崖边地带, 以保证树木生长对气候变化的响应较为敏感。使用生长锥在每棵树胸径(距地面1.3 m)处相互垂直的两个方向上钻取2根样芯, 编号后放入纸管封存。
| 类型Type | 石门山 | 张家庄 | 南五台 | 木王山 |
| 纬度Latitude | 34.44°N | 34.14°N | 33.95°N | 33.23°N |
| 经度Longitude | 106.14°E | 106.52°E | 108.58°E | 108.36°E |
| 海拔Altitude/m | 1994 | 1400 | 1600 | 2065 |
| 样本量Sample depth | 53 | 46 | 73 | 72 |
| 公共区间Common interval | 1863-2017 | 1929-2017 | 1844-2018 | 1874-2017 |
| 标准差Standard deviation(SD) | 0.377 | 0.303 | 0.201 | 0.170 |
| 信噪比Signal to noise ratio(SNR)/% | 13.019 | 39.485 | 16.962 | 25.100 |
| 平均敏感度Mean sensitivity(MS) | 0.223 | 0.296 | 0.136 | 0.138 |
| 所有样芯平均相关系数Mean correlation coefficient among all series(r1) | 0.310 | 0.473 | 0.200 | 0.206 |
| 树间平均相关系数Mean correlation coefficient between trees(r2) | 0.669 | 0.774 | 0.661 | 0.576 |
| 树内平均相关系数Mean correlation coefficient within trees(r3) | 0.301 | 0.467 | 0.181 | 0.195 |
| 样本群体表达信号Express population signal (EPS) | 0.929 | 0.975 | 0.944 | 0.962 |
| 一阶自相关系数Auto-correlation order(ac1) | 0.657 | 0.190 | 0.623 | 0.527 |
| 第一主成分所占方差量Variance in the first principal comment (pc1) | 0.356 | 0.519 | 0.234 | 0.291 |
| *代表在0.05水平显著相关; **代表在0.01水平显著相关 | ||||
将样品带回实验室进行固定、风干、打磨等预处理, 利用显微镜进行目视交叉定年, 利用精度为0.01 mm的LINTAB 6.0年轮宽度测量仪测量轮宽, 而后利用COFFECHA程序对定年及测量结果进行检验, 剔除质量较差、无法正常交叉定年的样品序列, 最终保留SMS处27棵油松53根样芯、ZJZ处23棵油松46根样芯、NWT处37棵油松73根样芯以及MWS处36棵油松72根样芯进入主序列(表 1)。
运用ARSTAN程序里的负指数函数, 对树轮宽度数据进行去趋势和标准化, 建立标准年表、差值年表及自回归年表, 通过比较, 最终确定采用标准年表(STD)进行后续研究。采用子样本信号强度SSS>0.85的年份作为年表的起始年, 最终得到4个样点处年表的可靠区间分别为1863-2017年、1929-2017年、1844-2018年和1874-2017年(图 4)。

|
| 图 4 各采样点标准年表(虚线), 单根树芯的标准年表(灰线)及样本量(点线), SSS为子样本信号强度 Fig. 4 Standard chronology of the site(the dash lines), standard chronology of each tree-ring core(the gray lines)and sample depth(the scatter lines)at each sampling site(SSS is subsample signal strength) |
从中国气象数据网(http://data.cma.cn/)分别获取4个采样点附近的天水气象站(34.35°N, 105.45°E, 海拔1141.6 m)、凤县气象站(33.56°N, 106.33°E, 海拔984 m)、长安气象站(34.05°N, 108.53°E, 海拔445 m)、镇安气象站(33.26°N, 109.09°E, 海拔693.7 m)1962-2017年的气象数据, 包括月降水量、月平均气温、月平均最高气温、月平均最低气温(图 2、图 3)。从荷兰皇家气象研究所数据共享网(http://climexp.knmi.nl)获取各样点所在0.5°×0.5°网格的帕默尔干旱指数(PDSI)。
采用1962-2017年4个样点油松STD年表与所获气象数据进行分析。考虑到树木径向生长对气候的响应存在“滞后效应”, 即树木径向生长状况不仅与当年的气候因子有关, 也受前一年气候的影响[25], 且秦岭树木的生长季通常在4-9月[26], 因此采用各样点前一年6月到当年9月的月平均气温、月平均最高气温、月平均最低气温、月降水量以及PDSI数据, 分别与该样点油松STD年表做Pearson相关分析, 以此描述树木径向生长状况对气候因子的响应。
1.4 干旱发生年及其前后树木径向生长变化各样点STD年表与该地PDSI做Pearson相关分析结果表明, 在SMS、ZJZ、NWT树木径向生长与各月PDSI均呈显著正相关, 但MWS树木与PDSI相关性极弱(图 5), 因此选取前三个样点开展树木径向生长对干旱事件响应的进一步分析。

|
| 图 5 各采样点标准年表与月气候因子在1962-2017年间的相关系数 Fig. 5 Correlation coefficients of the standard chronology of each sampling point and the monthly climate factors during 1962-2017 虚线代表 0.05水平的显著水平, p6-p12表示前一年6-12月, c1-c9表示当年1-9月; PDSI:帕默尔干旱指数Palmer drought severity index |
为分析干旱事件对树木径向生长的影响, 将PDSI小于-3的年份定义为干旱年[27]。计算STD年表在1963-2017的平均值与标准差, 求得平均值与1.5倍标准差的差值, 定义STD年表中小于这一差值的值为生长极低值。通过比较发现5月PDSI与STD年表两序列同步性较高, 且2000年的PDSI极低值与树木径向生长极低值相对应。因此选择2000年为目标干旱年, 进一步分析树木应对此干旱事件的能力。
计算树木径向生长对干旱事件响应的两个基础指标: 抵抗力(Resistance)和恢复力(Recovery), 分别表征树木在干旱事件发生后保持生长的能力以及恢复生长的能力。此外根据Schwarz等[28]提出的新评估方法计算恢复期(Recovery period)、总生长减少量(Total growth reduction)、平均生长减少量(Average growth reduction)以及平均恢复速率(Average recovery rate)4个指标。其中, 恢复期为树木生长在干旱发生后达到干旱前生长水平所需的时间;总生长减少量为在恢复期内的每一年生长降低的累积值, 即因干旱造成的增量总损失;平均生长减少量为总生长减少量除以恢复期, 量化了恢复期内平均每年树木生长受干旱影响的程度;平均恢复速率为干旱年生长减少的速率(等于1减抵抗力)除以恢复期, 表示恢复期内平均每年生长减少的速率。假设在任意抵抗力下树木的恢复弹力(Resilience)均等于1, 将其定义为完全恢复弹力标准线, 表示在干旱事件发生前后树木的生长可以完全恢复;通过比较树木生长的实际抵抗力、恢复力关系与完全恢复弹力标准线, 以探究树木实际恢复弹力。上述计算公式如下:
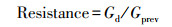
|
(1) |
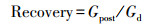
|
(2) |
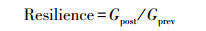
|
(3) |
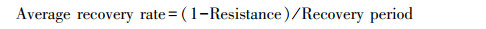
|
(4) |
式中, Gd表示干旱年的STD年表值, Gprev和Gpost分别表示干旱事件发生前5年和后5年STD年表的算术平均值。
2 结果与分析 2.1 树轮年表及参数由表 1可知, 4个样地标准年表统计特征值之间差异较为明显。信噪比(SNR)在ZJZ最高为39.485, 其次为MWS(25.1)、NWT(16.962)、SMS(13.091), 各年表SNR均处于较高水平, 说明样本包含丰富的环境信息;样本总体代表性(EPS)均大于0.85, 表明样本所含信号基本能代表总体特征;ZJZ处树间平均相关系数(r3)最大, 其次是SMS、MWS、NWT, 表明在ZJZ、SMS树木径向生长能够更好地反映气候变化的一致性。西部SMS、ZJZ的标准差(SD)和平均敏感度(MS)均大于中部NWT、MWS, 总体而言, 西部地区树木包含更多气候信息且径向生长对气候变化的敏感度更高[29]。
对4个样地的一阶差年表进行Pearson相关, 结果发现SMS和ZJZ、SMS和NWT、NWT和MWS的一阶差年表之间均呈现0.01水平上的极显著相关;ZJZ与NWT、MWS的一阶差年表之间也具有0.05水平上的显著相关(表 2), 表明样点间树木生长的年际高频变化有较高的一致性(图 4)。
| SMS | ZJZ | NWT | MWS | |
| SMS | 1 | 0.578** | 0.592** | 0.246 |
| ZJZ | 1 | 0.287* | 0.272* | |
| NWT | 1 | 0.462** | ||
| MWS | 1 | |||
| STD:标准年表Standard chronology;SMS:石门山;ZJZ: 张家庄;NWT:南五台;MWS:木王山; | ||||
各样地STD年表与气候因子的相关关系分析结果表明(图 5), SMS树木径向生长与前一年7月的平均气温、平均最高气温、平均最低气温均显著负相关(r介于-0.396--0.286), 与前一年9月的平均最高气温显著负相关(r=-0.294), 与当年5-7月的平均气温和平均最高气温显著负相关(r介于-0.471--0.412);ZJZ树木径向生长与前一年7-9月平均气温和平均最高气温显著负相关(r介于-0.439--0.291), 与当年4月的平均最高气温、当年5-7月的平均气温和平均最高气温显著负相关(r介于-0.468--0.280);NWT树木径向生长与前一年7-9月的平均气温、前一年7-8月的平均最低气温、前一年8-9月的平均最高气温均呈显著负相关(r介于-0.395--0.278), 与当年5月的平均气温呈显著负相关(r=-0.402);MWS树木径向生长与前一年8月的平均气温呈显著负相关(r=-0.370), 与当年2月的平均最高气温和平均最低气温、当年3-4月的平均气温呈显著正相关(r介于0.295-0.351)。
SMS树木径向生长与当年5月和7月降水呈显著正相关(r介于0.314-0.369), ZJZ树木径向生长与前一年9月和当年5月降水呈显著正相关(r介于0.316-0.372), SMS、ZJZ年表与全年各月PDSI均呈正相关(r介于0.206-0.572), 尤其以当年5-7月最为显著(r介于0.468-0.526);NWT树木径向生长与前一年9月和当年5月降水显著负相关(r介于0.276-0.348), 与各月PDSI均呈正相关(r介于0.248-0.469), 尤其以前一年9月和当年5-6月最为显著(r介于0.422-0.469);MWS年表与当年4月降水显著正相关(r=0.32), 与PDSI未呈现显著相关关系。
2.3 树木径向生长对干旱事件的响应研究结果表明3个样地树木对干旱的抵抗力表现为在NWT最强, 在SMS最弱(即NWT>ZJZ>SMS);总生长减少量表现为在SMS最高, 在NWT最低(即SMS>ZJZ>NWT);平均减少量在SMS和ZJZ处较为一致且均高于NWT;3个指标综合表明NWT树木径向生长对干旱的抵抗能力最强, 其次为ZJZ和SMS(图 6)。树木对干旱事件的恢复力在SMS与ZJZ较为一致, 均小于NWT;平均恢复速率则表现为ZJZ和NWT较为一致, 均高于SMS, 因此树木对干旱事件的恢复能力总体表现为SMS弱于NWT和ZJZ(图 6)。

|
| 图 6 SMS、ZJZ、NWT单根STD年表的抵抗力、总生长减少量、平均生长减少量、恢复力以及平均恢复速率 Fig. 6 Resistance, total growth reduction, average growth reduction, resistance, and average recovery rate of each single STD chronology at SMS, ZJZ, and NWT |
通过比较干旱年(2000年)树木径向生长的实际恢复弹力与完全恢复弹力标准线发现(图 7), 在SMS对于抵抗力低于0.54的样芯年表, 其实际恢复弹力低于完全恢复弹力标准线, 并且随着抵抗力减弱, 实际恢复弹力逐渐远离完全恢复弹力标准线;在ZJZ和NWT, 年表的实际恢复弹力在完全恢复弹力标准线上方, 表明这两点的树木恢复弹力均强于1, 即干旱后树木能完全恢复到干旱发生之前的状态, 甚至比之前的状态更好;因此树木对干旱事件的恢复弹力同样表现为SMS弱于NWT和ZJZ。

|
| 图 7 SMS、ZJZ、NWT树木对2000年干旱事件完全恢复弹力标准线和实际恢复弹力 Fig. 7 The full resilience line and the actual resilience about drought event in 2000 at SMS, ZJZ, and NWT |
秦岭地区由于地势复杂, 气候差异大, 年降水量整体表现为南坡多于北坡, 年均温由东南向西北递减, 因此势必会造成限制其南北树木径向生长的关键气候因子有所差异。在秦岭北部为暖温带季风区, 较南部更加干旱, 升温容易引起土壤水分胁迫, 从而对树木生长产生不利影响。研究结果表明在SMS、ZJZ及NWT 3个位于西部及北坡的样点, 树木径向生长与前一年生长季中后期、当年生长季早期及中期的气温呈显著负相关关系。这可能是由于生长季中后期温度升高导致油松为减少水分蒸腾而关闭气孔, 光合速率下降, 有机物的合成及储存减少, 不利于当年树木的生长, 甚至影响下一年树木的生长[30]。同时夏季气温升高通常会引起干旱事件, 使木质部导水管道栓塞, 降低树木对水分的获取能力, 造成树木生长下降[31-33]。此现象在我国西北干旱区也较为普遍, 例如侯迎等在崆峒山对油松的研究表明径向生长与夏季均温呈显著负相关关系[34], 在青藏高原东缘的研究中也发现冷杉的径向生长与前一年8月温度显著负相关[35]。生长季初期平均最高气温升高可能导致土壤水分蒸发加剧, 同时树木蒸腾作用增强会进一步加快土壤水分的消耗, 春季高温导致的干旱加剧会对树木生长产生明显的抑制作用, 故表现为树木径向生长与生长季早期温度呈显著负相关[36]。Shi等在东秦岭的研究发现油松径向生长与当年5-6月平均最高气温显著负相关[15]。Sun等在秦岭北部的研究同样表明油松生长受5月平均最高气温的限制[16]。秦岭西部及北部3个样点STD年表均与当年生长季初期降水显著正相关, 这是由于在较干旱的样点, 生长季初期气温开始上升, 若降水不足则容易造成严重的水分胁迫[37], 此时充足的降水能够提高土壤的含水量, 为树木形成层启动提供必要的水分条件。在秦岭伏牛山的研究中也表现出当年5月降水对油松生长有显著促进作用[15]。ZJZ、NWT年表与前一年生长季中后期降水显著正相关, 这是由于前一年降水量充沛, 土壤含水量较高, 有利于土壤水源的涵养, 甚至可以延长生长季从而使树木积累更多的营养物质, 为来年树木形成层活动提供良好条件[37-39]。
秦岭南坡受东亚季风的影响, 气候相对北坡更加湿润, 树木生长的主要限制因子为温度[18], 降水对树木生长的限制作用不显著, 故而秦岭中部南坡样点(MWS)树木径向生长与当年初春休眠期的气温表现为显著正相关。休眠期较高的温度能够减轻冻害, 促进积雪融化, 为生长季开始时树木生长提供充足的水分, 甚至能够提前生长季开始时间[40-41]。这与刘洪滨等在秦岭南坡的研究结论一致[42]。Dang等人的研究也表明早春温度是控制大陆气候区亚高山环境的主要限制因子[43]。MWS油松与生长季末期平均最低气温及平均气温表现为显著正相关, 这可能是因为在油松生长季末期平均气温升高能够有效延长生长季的结束时间[44]。MWS年表与生长季早期降水显著负相关, 初春积雪融化, 土壤含水量充足, 过多的降水会造成云量过多, 阻碍光合作用, 这也印证了4月气温与径向生长的显著正相关关系。Chen等在秦岭西部舟曲县对岷江冷杉的研究也得出了相似的结论[45]。
3.2 秦岭中西部南北坡油松径向生长对干旱事件响应差异研究通过分析2000年干旱事件前后树木径向生长状况变化发现, 与秦岭南坡较为湿润的ZJZ相比, 北坡SMS树木对干旱的抵抗能力较差, 且恢复能力和恢复弹力也更差;而相较位于中部北坡的NWT, 西部北坡较干旱的SMS树木同样表现出应对干旱事件抵抗能力、恢复能力及恢复弹力更差的特点。这可能是因为NWT位于秦岭中部, 气温和降水条件均优于西部, 更利于树木生长。同时, NWT树木对气候变化的敏感度低于SMS、ZJZ树木, 因此受干旱事件产生的影响也相对较小, 具有较强的抵抗力。NWT树木在干旱发生后的生长减少量低, 且气候条件适宜, 更利于树木的生长, 因此该处树木对干旱的恢复能力同样强于其它两样点, 甚至很快可以恢复至干旱前的生长水平。传统研究发现, 抵抗力较高的树木通常恢复力较低, 两者之间通常呈现相反的关系, 反映了树木应对干旱事件时抵抗能力和恢复能力之间的一种权衡, 例如在对北美和欧洲100多个树种树木的年轮研究发现, 干旱环境下树木径向生长对干旱的抵抗力和恢复力之间通常呈显著负相关[24]。但有学者提出, 这种抵抗力与恢复力的权衡可能是由于传统的计算方法所导致的, 并不具有显著的实际意义[28], 因此本研究引用新的参数评估树木对干旱的抵抗能力和恢复能力, 结果发现这种权衡现象并非在所有地区都存在。
树木应对干旱事件的能力会因区域干湿差异而有所不同, 此现象在其他地区的研究中也有相应发现。例如有研究发现, 西班牙东部干旱地区的地中海白松(Aleppo pine)比湿润地区树木的抵抗力更差[22];在对祁连山青海云杉的研究中发现, 西部较干旱地区树木的抵抗力同样比东部较湿润地区更差, 且树木从干旱中恢复的能力主要取决于该地的降水量[46], 均与本文研究结果一致。Gazol等对北美和欧洲2000多个样点的100多种树木的研究中发现, 湿润区域的温带阔叶林对干旱有更强的抵抗力, 而干旱半干旱区域的针叶林的抵抗力则更差[21], 同样印证了这一结论。然而, 也有研究得出相反的结论, 对欧洲山毛榉的研究发现, 生长在该物种分布区干旱边缘的树木抵抗力更强[23], 这说明并非所有树木的抵抗力都表现为在湿润地区更强, 因此在不同环境下分析树木径向生长应对干旱事件的能力和策略需要更深入的分析。抵抗力与恢复弹力的指标对树木径向生长应对干旱事件时树木的长期存活非常重要, 有研究表明, 对于恢复弹力较差的树木, 在干旱发生后常会出现生长严重下降乃至死亡的现象。DeSoto等利用超过3500棵树的存活和已死亡树木的树轮数据库分析发现, 与存活树木相比, 在缺水期间死亡的同树种树木对干旱事件的抵抗力较弱;对于被子植物, 较低的抵抗力往往使得树木在干旱事件中有较高的死亡风险[47]。因此在未来对研究区树木的管理与保护工作中, 应更加关注西部北坡树木生长状况, 中部树木则无需进行过多的人工干预。
4 结论秦岭西部及北坡油松径向生长均受到前一年生长季中后期及当年生长季早中期高温的制约, 且会受到生长季初期降水不足的影响。中部南坡样点的油松则与休眠期和当年生长季末期气温显著正相关, 并受当年生长季早期降水不足的制约。总而言之, 生长季气温升高所引发的水分匮缺, 是研究区西部和北坡油松径向生长受限制的主要原因;且中部南坡树木受温度因子和水分的制约要明显弱于其他3个样点。西部及北坡的3个样点树木径向生长均受到2000年干旱事件的影响, 通过对干旱事件发生前后各样点油松生长状况的分析发现, 秦岭西部北坡的油松相比西部南坡及中部北坡对干旱具有更差的抵抗力及恢复力, 这体现了干旱地区树木生长对气候变化的响应更为敏感的特征。本研究比较了秦岭中西部不同干湿环境下油松对气候因子响应及应对干旱事件能力的差异, 可为气候变暖背景下该区域油松生长的管理及森林生态系统的保护提供理论依据。
致谢: 感谢张家庄林场工作人员在树轮样品采集中给予的帮助。| [1] |
葛全胜, 王芳, 王绍武, 程邦波. 对全球变暖认识的七个问题的确定与不确定性. 中国人口·资源与环境, 2014, 24(1): 1-6. |
| [2] |
Vicente-Serrano S M, Camarero J J, Azorin-Molina C. Diverse responses of forest growth to drought time-scales in the Northern Hemisphere. Global Ecology and Biogeography, 2014, 23(9): 1019-1030. DOI:10.1111/geb.12183 |
| [3] |
Dai A G. Increasing drought under global warming in observations and models. Nature Climate Change, 2013, 3(1): 52-58. DOI:10.1038/nclimate1633 |
| [4] |
贾飞飞, 孙翠洋, 孙红月, 李鑫. 昌岭山两个优势树种径向生长对气候变化的响应. 生态学报, 2019, 39(17): 6332-6340. |
| [5] |
Xu X T, Medvigy D, Trugman A T, Guan K, Good S P, Rodriguez-Iturbe I. Tree cover shows strong sensitivity to precipitation variability across the global tropics. Global Ecology and Biogeography, 2018, 27(4): 450-460. DOI:10.1111/geb.12707 |
| [6] |
Gazol A, Camarero J J, Vicente-Serrano S M, Sánchez-Salguero R, Gutiérrez E, de Luis M, Sangüesa-Barreda G, Novak K, Rozas V, Tíscar P A, Linares J C, Martín-Hernández N, del Castillo E M, Ribas M, García-González I, Silla F, Camisón A, Génova M, Olano J M, Longares L A, Hevia A, Tomás-Burguera M, Galván J D. Forest resilience to drought varies across biomes. Global Change Biology, 2018, 24(5): 2143-2158. DOI:10.1111/gcb.14082 |
| [7] |
Schwalm C R, Anderegg W R L, Michalak A M, Fisher J B, Biondi F, Koch G, Litvak M, Ogle K, Shaw J D, Wolf A, Huntzinger D N, Schaefer K, Cook R, Wei Y X, Fang Y Y, Hayes D, Huang M Y, Jain A, Tian H Q. Global patterns of drought recovery. Nature, 2017, 548(7666): 202-205. DOI:10.1038/nature23021 |
| [8] |
康慕谊, 朱源. 秦岭山地生态分界线的论证. 生态学报, 2007, 27(7): 2774-2784. DOI:10.3321/j.issn:1000-0933.2007.07.014 |
| [9] |
周旗, 卞娟娟, 郑景云. 秦岭南北1951-2009年的气温与热量资源变化. 地理学报, 2011, 66(9): 1211-1218. |
| [10] |
延军平, 郑宇. 秦岭南北地区环境变化响应比较研究. 地理研究, 2001, 20(5): 576-582. DOI:10.3321/j.issn:1000-0585.2001.05.007 |
| [11] |
丁敏, 黄春长. 秦岭北麓平原对全球变暖的响应及其对果业发展的影响研究. 自然资源学报, 2003, 18(2): 235-239. DOI:10.3321/j.issn:1000-3037.2003.02.017 |
| [12] |
Herman F R. Tree rings and climate. Forest Science, 1978, 24(3): 409. |
| [13] |
华亚伟, 张红娟, 刘康. 基于油松树轮重建陕西省镇安县165年以来3-4月平均最高气温. 应用生态学报, 2020, 31(2): 381-387. |
| [14] |
Song H M, Liu Y, Li Q, Gao N, Ma Y Y, Zhang Y H. Tree-Ring based May-July temperature reconstruction since AD 1630 on the western Loess Plateau, China. PLoS One, 2014, 9(4): e93504. DOI:10.1371/journal.pone.0093504 |
| [15] |
Shi J F, Li J B, Cook E R, Zhang X Y, Lu H Y. Growth response of Pinus tabulaeformis to climate along an elevation gradient in the eastern Qinling Mountains, central China. Climate Research, 2012, 53(2): 157-167. DOI:10.3354/cr01098 |
| [16] |
Sun C F, Liu Y. Climate response of tree radial growth at different timescales in the Qinling Mountains. PLoS One, 2016, 11(8): e0160938. DOI:10.1371/journal.pone.0160938 |
| [17] |
Jin Q, Bai H Y, Su K, Li S H, Wang J. The response of tree-ring growth to climate at upper timberline of southern aspect of Mt. Taibai. MATEC Web of Conferences, 2016, 49: 05003. DOI:10.1051/matecconf/20164905003 |
| [18] |
王红桃, 包光. 秦岭及周边地区树木径向生长与气候因子的关系研究进展. 亚热带资源与环境学报, 2016, 11(1): 52-58. DOI:10.3969/j.issn.1673-7105.2016.01.008 |
| [19] |
Barbeta A, Mejía-Chang M, Ogaya R, Voltas J, Dawson T E, Peñuelas J. The combined effects of a long-term experimental drought and an extreme drought on the use of plant-water sources in a Mediterranean forest. Global Change Biology, 2015, 21(3): 1213-1225. DOI:10.1111/gcb.12785 |
| [20] |
Anderegg, W R L, Flint A, Huang C Y, Flint L, Berry J A, Davis F W, Sperry J S, Field C B. Tree mortality predicted from drought-induced vascular damage. Nature Geoscience, 2015, 8(5): 367-371. DOI:10.1038/ngeo2400 |
| [21] |
Gazol A, Camarero J J, Anderegg W R L, Vicente-Serrano S M. Impacts of droughts on the growth resilience of Northern Hemisphere forests. Global Ecology and Biogeography, 2017, 26(2): 166-176. DOI:10.1111/geb.12526 |
| [22] |
Gazol A, Ribas M, Gutiérrez E, Camarero J J. Aleppo pine forests from across Spain show drought-induced growth decline and partial recovery. Agricultural and Forest Meteorology, 2017, 232: 186-194. DOI:10.1016/j.agrformet.2016.08.014 |
| [23] |
Cavin L, Jump A S. Highest drought sensitivity and lowest resistance to growth suppression are found in the range core of the tree Fagus sylvatica L. not the equatorial range edge. Global Change Biology, 2017, 23(1): 362-379. DOI:10.1111/gcb.13366 |
| [24] |
傅抱璞, 李兆元. 秦岭山地的气候特点. 陕西气象, 1983(1): 1-11. |
| [25] |
张贇, 尹定财, 张卫国, 岳海涛, 杜杰次丹, 李秋平, 杨荣, 田昆. 普达措国家公园2个针叶树种径向生长对温度和降水的响应. 生态学报, 2018, 38(15): 5383-5392. |
| [26] |
吴祥定, 邵雪梅. 中国秦岭地区树木年轮密度对气候响应的初步分析. 应用气象学报, 1994, 5(2): 253-256. |
| [27] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB/20481-2017气象干旱等级. 北京: 中国标准出版社, 2017.
|
| [28] |
Schwarz J, Skiadaresis G, Kohler M, Kunz J, Schnabel F, Vitali V, Bauhus J. Quantifying growth responses of trees to drought-a critique of commonly used resilience indices and recommendations for future studies. Current Forestry Reports, 2020, 6(3): 185-200. DOI:10.1007/s40725-020-00119-2 |
| [29] |
Yamaguchi D K. Methods of dendrochronology, applications in the environmental sciences. Arctic and Alpine Research, 1991, 23(1): 120. |
| [30] |
陈力, 吴绍洪, 戴尔阜. 长白山红松和落叶松树轮宽度年表特征. 地理研究, 2011, 30(6): 1147-1155. |
| [31] |
Silva L C R, Anand M. Probing for the influence of atmospheric CO2 and climate change on forest ecosystems across biomes. Global Ecology and Biogeography, 2013, 22(1): 83-92. DOI:10.1111/j.1466-8238.2012.00783.x |
| [32] |
Subedi N, Sharma M. Climate-diameter growth relationships of black spruce and jack pine trees in boreal Ontario, Canada. Global Change Biology, 2013, 19(2): 505-516. DOI:10.1111/gcb.12033 |
| [33] |
Deslauriers A, Beaulieu M, Balducci L, Giovannelli A, Gagnon M J, Rossi S. Impact of warming and drought on carbon balance related to wood formation in black spruce. Annals of Botany, 2014, 114(2): 335-345. DOI:10.1093/aob/mcu111 |
| [34] |
侯迎, 王乃昂, 李钢, 郑芳. 利用树轮资料重建1751-2005年崆峒山地区夏季温度变化. 气候变化研究进展, 2007, 3(3): 172-176. DOI:10.3969/j.issn.1673-1719.2007.03.010 |
| [35] |
彭剑峰, 勾晓华, 陈发虎, 刘普幸, 张永香, 张永, 田沁花. 阿尼玛卿山不同海拔祁连圆柏树轮宽度年表特征对比分析. 冰川冻土, 2006, 28(5): 713-721. DOI:10.3969/j.issn.1000-0240.2006.05.014 |
| [36] |
宋慧明, 刘禹, 梅若晨, 赵伯阳, Payomrat P, 张欣佳. 甘肃竺尼山油松树轮宽度气候响应. 地球环境学报, 2017, 8(2): 119-126. |
| [37] |
高尚玉, 鲁瑞洁, 强明瑞, 哈斯, 张登山, 陈原, 夏虹. 140年来腾格里沙漠南缘树木年轮记录的降水量变化. 科学通报, 2006, 51(3): 326-331. DOI:10.3321/j.issn:0023-074X.2006.03.014 |
| [38] |
鲁瑞洁, 夏虹. 腾格里沙漠南缘油松树轮宽度变化及其对气候因子的响应. 中国沙漠, 2006, 26(3): 399-402. DOI:10.3321/j.issn:1000-694X.2006.03.015 |
| [39] |
王亚军, 高尚玉, 马玉贞, 鲁瑞洁, 桑艳礼, 蒙红卫. 基于树轮重建的宁夏河东沙地西部公元1899年来年降水量的变化. 干旱区地理, 2010, 33(3): 377-384. |
| [40] |
Liu Y, Linderholm H W, Song H M, Cai Q F, Tian Q H, Sun J Y, Chen D L, Simelton E, Seftigen K, Tian H, Wang R Y, Bao G, An Z S. Temperature variations recorded in Pinus tabulaeformis tree rings from the southern and northern slopes of the central Qinling Mountains, central China. Boreas, 2009, 38(2): 285-291. DOI:10.1111/j.1502-3885.2008.00065.x |
| [41] |
李双双, 芦佳玉, 延军平, 刘宪锋, 孔锋, 王娟. 1970-2015年秦岭南北气温时空变化及其气候分界意义. 地理学报, 2018, 73(1): 13-24. |
| [42] |
刘洪滨, 邵雪梅. 秦岭南坡佛坪1789年以来1-4月平均温度重建. 应用气象学报, 2003, 14(2): 188-196. DOI:10.3969/j.issn.1001-7313.2003.02.006 |
| [43] |
Dang H S, Jiang M X, Zhang Q F, Zhang Y J. Growth responses of subalpine fir (Abies fargesii) to climate variability in the Qinling Mountain, China. Forest Ecology and Management, 2007, 240(1/3): 143-150. |
| [44] |
牛震敏, 基于树木年轮的川西高原气候变化历史重建[D]. 兰州: 兰州大学, 2016.
|
| [45] |
Chen F, Yuan Y J, Wei W S, Yu S L, Shang H M, Zhang T W, Zhang R B, Wang H Q, Qin L. Tree-ring based temperature reconstruction for the west Qinling Mountains (China): linkages to the High Asia, solar activity and Pacific-Atlantic Ocean. Geochronometria, 2014, 41(3): 234-244. DOI:10.2478/s13386-013-0159-9 |
| [46] |
Zhang L N, Li H, Ran Y L, Wang K Y, Zeng X M, Liu X H. Regional and local moisture gradients drive the resistance to and recovery from drought of Picea crassifolia Kom. in the Qilian Mountains, Northwest China. Forests, 2019, 10(9): 817. DOI:10.3390/f10090817 |
| [47] |
DeSoto L, Cailleret M, Sterck F, Jansen S, Kramer K, Robert E M R, Aakala T, Amoroso M M, Bigler C, Camarero J J, Čufar K, Gea-Izquierdo G, Gillner S, Haavik L J, Hereş A M, Kane J M, Kharuk V I, Kitzberger T, Klein T, Levanič T, Linares J C, Mäkinen H, Oberhuber W, Papadopoulos A, Rohner B, Sangüesa-Barreda G, Stojanovic D B, Suárez M L, Villalba R, Martínez-Vilalta J. Low growth resilience to drought is related to future mortality risk in trees. Nature Communications, 2020, 11(1): 545. DOI:10.1038/s41467-020-14300-5 |