 2022, Vol. 42
2022, Vol. 42文章信息
- 方青慧, 杨晶, 张彩军, 张倩, 苏军虎
- FANG Qinghui, YANG Jing, ZHANG Caijun, ZHANG Qian, SU Junhu
- 放牧管理模式对高原鼢鼠(Eospalax baileyi)鼠丘形态特征的影响
- Effects of grazing management regimes on mound morphological characteristics of plateau zokors (Eospalax baileyi)
- 生态学报. 2022, 42(4): 1619-1628
- Acta Ecologica Sinica. 2022, 42(4): 1619-1628
- http://dx.doi.org/10.5846/stxb202012253273
-
文章历史
- 收稿日期: 2020-12-25
- 网络出版日期: 2021-11-04





2. 甘肃农业大学-新西兰梅西大学草地生物多样性研究中心, 兰州 730070;
3. 甘肃省祁连山草原生态系统野外科学观测研究站, 武威 733200
2. Gansu Agricultural University-Massey University Research Centre for Grassland Biodiversity, Gansu Agricultural University, Lanzhou 730070, China;
3. Gansu Qilianshan Grassland Ecosystem Observation and Research Station, Wuwei 733200, China
放牧是草地的主要利用方式之一, 对草地的演替和草地生产力有显著影响[1]。放牧过程中家畜采食、践踏和排泄会影响草地及其功能的发挥, 不同时间的扰动及草地植物、微生物和动物等各个组分间的相互作用, 也会对草地产生显著不同的影响。然而, 家畜放牧除对土壤理化性质、植被特性和草地生产力等有着显著影响外[2], 对啮齿动物的分布、生活空间和繁殖特性等也有影响[3]。家畜的踩踏和采食可使啮齿动物喜食杂草比例上升, 其食物资源的分布结构发生改变[4]。同时, 较低的覆盖水平会使小型哺乳动物的种类增加, 成为草地管理的一个棘手问题[5], 而啮齿动物的推土造丘活动是其对草地影响的主要因素, 会影响到草地土壤特性及植物群落结构的改变, 甚至会出现草地秃斑, 造成严重的水土流失等。
放牧管理模式作为控制放牧强度、维持家畜数量和草地生态系统平衡的利用体系指标[2], 对草地群落结构、草地生产力、土壤结构和养分性质等产生了不同的影响[6]。针对放牧强度和放牧频率的草原管理模式, 已经有禁牧、自由放牧和轮牧等不同模式[7], 其中禁牧是草地管理最常用及草地恢复最快最有效的手段, 也是促进草地生态系统中物质循环和能量流动, 调节食草动物与植物之间关系的有效方法, 但长期禁牧不利于地上植被的继续恢复, 甚至会导致草地生产力降低等[1]。通过对家畜放牧强度与放牧时间上的调整, 近年来又产生了传统放牧和生长季休牧等不同的放牧管理模式[8], 而确定合理的放牧管理模式和放牧时期更有利于草地健康发展和草地保护。因此, 全面评价不同放牧管理模式对草地的影响, 并采用适宜的管理措施是草地可持续利用的关键。
高原鼢鼠(Eospalax baileyi)是青藏高原独有的地下啮齿动物[9], 其在取食、交配、构筑巢窝和洞道挖掘等活动中不断啃食牧草, 并向地表堆土形成鼠丘, 过多的鼠丘会覆盖植物, 造成植物死亡, 在一定程度上也造成了水土流失, 影响着草地生态系统的结构和功能[10-11]。鼠丘的表面积大小及其土壤性质的变化也是对草地产生影响的关键, 高原鼢鼠推土形成的新鼠丘属于次生裸地[12], 在水力和风力长期侵蚀及家畜践踏的过程中其形状发生改变, 形成的鼠丘斑块面积增大[13]。由于鼠丘斑块中心与边缘的理化性质不同[14], 其差异导致斑块的边缘和中心演替程度不同, 对草地产生的影响不同。除此之外, 家畜的踩踏也会使土壤结构发生改变, 土壤变紧实, 增加高原鼢鼠在挖掘过程中的能量消耗[15], 这些因素都会影响高原鼢鼠的造丘活动, 鼠丘的不同特征也直接关系到其影响的性质(危害性)及其大小。目前仅见鼠丘分布格局[16]、鼠丘演替过程中植被及其土壤理化性质的变化和鼠丘密度等的研究[17-18]。而阐明不同放牧管理模式下高寒草甸高原鼢鼠鼠丘的变化特征, 对评价高原鼢鼠的危害和影响及草地管理利用具有重要意义。基于此, 本研究于5月、8月和10月测定禁牧、生长季休牧、传统放牧和连续放牧4种放牧管理模式下高原鼢鼠当季鼠丘(采样时近2个月内形成)的半径、高度、表面积和体积, 同时结合其土壤紧实度、水分、容重和地上、地下生物量的不同变化, 旨在明确放牧对高原鼢鼠鼠丘形态特征的影响, 为高原鼢鼠影响评价、鼠害防治及草地管理提供理论依据。
1 样地概况及研究方法 1.1 样地概况试验样地位于甘肃省武威市天祝藏族自治县抓喜秀龙镇马营沟附近(37°11′N, 102°32′E), 海拔为2700—3100m, 昼夜温差较大, 空气较为稀薄, 太阳辐射强, 气候寒冷潮湿, 年均降水量416mm, 主要集中于7—9月份, 年蒸发量为1592mm, 年均气温-0.1℃, ≥0℃的年积温1380℃, 无绝对无霜期。优势种主要有矮嵩草(Kobresia humilis)、垂穗披碱草(Elymus nutans)和苔草(Carex humilis)等, 坡地有灌丛分布;动物以啮齿类为主, 有高原鼢鼠和达乌尔黄鼠(Spermophilus dauricus)等近10种, 其中高原鼢鼠是当地的优势种之一[19]。
1.2 试验设计与取样方法本试验选择4种放牧管理模式的高寒草甸样地, 包括禁牧(NG)样地(样地面积为7 hm2, 自2010年起全年禁牧, 无放牧)、生长季休牧(RG)样地(样地面积为18hm2, 管理模式大于10年, 每年在4月20日—9月20日间休牧, 其余时间自由放牧, 放牧率为4.86羊hm-2 a-1、传统放牧(TG)样地(样地面积为18 hm2, 管理模式大于10年, 每年在6月20日—9月20日间休牧, 其余时间自由放牧, 放牧率为6.08羊hm-2 a-1)和连续放牧(CG)(样地面积为2hm2, 管理模式大于20年, 全年自由放牧, 放牧率为8.57羊hm-2 a-1)[8]。
分别于2019年5月下旬, 8月中旬和10月下旬在试验样地内选取25个当季(即5月、8月和10月不同时期鼠丘是采样时近2个月内形成的)形成的新鼠丘(不足25个新鼠丘则全部选取), 用卷尺测得其最大直径、最小直径及其高度, 以鼠丘的俯视图最大直径和最小直径的均值作圆[20], 利用用球缺公式计算其体积(V)和表面积(S)大小。
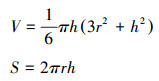
|
式中, π为圆周率, r为鼠丘半径, h为鼠丘高度。
分别于这3个时期在各试验样地选取9个样点, 在每个样点附近挖取一个30cm的剖面, 以10cm为间隔, 用体积为100cm3的环刀取土壤样品, 用铝盒烘干法测定其水分和容重;取样后用SC-900土壤紧实度仪在选取的9个样点附近任选5个点测定每个样地的土壤紧实度, 每2.5cm记录一次, 测0—30cm的紧实度, 求其平均值[21]。最后, 在每个样地内按照“W”型选取9个样点, 清除地表杂物, 每个样点建立20cm×20cm的样方, 分0—10, 10—20, 20—30cm取生物量, 将土样装入60目尼龙网袋, 抖落散土, 并用清水冲洗干净, 并在70℃下烘至恒重, 称其干重, 其中在8月植物生长旺盛时期进行地上生物量的测定, 在每个样地内设9个样方, 按照禾本科、莎草科、豆科和杂草类4大功能群分类, 齐地刈割地上植物, 将其装入信封, 在恒温70℃下烘干称重[22]。
1.3 数据分析用Excel 2016进行数据统计并制作表格;用SPSS 23.0软件对各指标数据进行One-way ANOVA分析, 并对鼠丘表面积和体积与植物各功能群地上生物量之间进行相关性分析;应用R语言软件Vegan软件包基于线性模型对鼠丘特征与土壤因子(土壤水分、土壤容重和土壤紧实度)及地下生物量之间的关系进行冗余分析(Redundancy analysis, RDA), 所有排序的显著性均由Permutation test(置换999次)进行检验[23];用GraphPad Prism8进行图片绘制。
2 结果与分析 2.1 放牧管理模式对植物地上、地下生物量的影响放牧管理模式显著影响了植物地上生物量(表 1)。与NG相比, 禾本科植物和豆科植物的地上生物量在RG、TG和CG样地均显著降低(P<0.05), 其大小依次为NG>RG>TG>CG;莎草科植物的地上生物量在RG和CG样地分别较NG样地降低了73.86%和84.62%, 在TG样地增加了38.30%, 其中在TG样地显著高于NG、RG和CG样地(P<0.05);放牧管理模式也显著影响了杂草类植物的地上生物量, 其中TG和CG样地分别较NG样地增加了20.32%和8.95%, 而在RG样地降低了15.65%(P<0.05)。
| 月份 Months |
处理 Treatments |
地下生物量Underground biomass | 地上生物量Aboveground biomass | ||||||
| 0—10cm | 10—20cm | 20—30cm | GRA | CYP | LEG | MIS | |||
| 5 | NG | 2321.83±180.43a | 905.31±84.06a | 409.34±4.49a | — | — | — | — | |
| RG | 1329.94±48.43c | 762.63±18.92a | 445.01±64.81a | — | — | — | — | ||
| TG | 1844.59±35.30b | 886.63±87.42a | 419.53±36.78a | — | — | — | — | ||
| CG | 1222.93±114.89c | 387.26±51.55b | 122.29±19.29b | — | — | — | — | ||
| 8 | NG | 4875.75±145.64a | 669.33±58.05a | 449.92±16.75a | 193.86±2.81a | 67.80±2.33b | 83.91±2.79a | 112.62±7.17b | |
| RG | 2726.92±221.90b | 636.00±116.08a | 180.92±67.88b | 168.98±4.35b | 17.72±2.49c | 69.03±3.48b | 95.00±7.11c | ||
| TG | 2758.42±1234.45b | 553.33±125.35a | 333.83±102.23ab | 137.98±9.09c | 93.77±4.19a | 59.56±2.31c | 135.50±0.47a | ||
| CG | 2631.75±324.41b | 352.42±49.10b | 197.42±5.28b | 27.54±1.10d | 10.43±0.51c | 6.07±0.09d | 122.70±1.26ab | ||
| 10 | NG | 2895.75±442.64a | 522.67±84.79a | 312.92±119.68a | — | — | — | — | |
| RG | 2716.83±305.56a | 538.83±70.02a | 362.50±39.26a | — | — | — | — | ||
| TG | 2780.08±289.40a | 527.58±65.69a | 390.75±143.23a | — | — | — | — | ||
| CG | 2210.67±72.02b | 351.33±31.26b | 234.25±24.91b | — | — | — | — | ||
| NG: 禁牧No grazing;RG: 生长季休牧Growing season rest grazing;TG: 传统放牧Traditional grazing;CG: 连续放牧Continuous grazing;GRA: 禾本科Gramineae;CYP: 莎草科Cyperaceae;LEG: 豆科Leguminosae;MIS: 杂草类Miscellaneous;—: 无数据No data;不同的字母表示同一季节不同放牧管理模式间的差异性显著(P<0.05) | |||||||||
放牧管理模式显著影响了植物地下生物量的变化, 除8月10—20cm土层的地下生物量外, 5月、8月和10月0—20cm土层的地下生物量均为NG>TG>RG>CG, 其中在0—10cm土层, 5月和8月NG样地的地下生物量在显著大于RG、TG和CG样地的地下生物量(P<0.05), 而10月CG样地的地下生物量显著小于NG、RG和TG样地的地下生物量(P<0.05), 10—20cm土层和20—30cm(5月和10月)土层的地下生物量在CG样地均显著小于其它3种放牧管理模式(P<0.05)。8月RG、TG和CG样地20—30cm土层的地下生物量较NG分别减少了59.79%、25.80%和56.12%, 其中在RG和CG样地与NG样地存在显著差异(P<0.05)(表 1)。
2.2 放牧管理模式对土壤因子的影响放牧管理模式显著影响了土壤水分含量的变化, 5月RG、TG和CG样地的土壤水分分别较NG样地增加了11.65%、8.43%和19.35%;8月和10月CG样地的土壤水分显著大于NG、RG和CG(P<0.05);5月、8月和10月土壤容重大小依次为CG>TG>RG>NG, 其中5月CG样地的土壤容重较NG样地显著增加(P<0.05), 8月RG、TG和CG样地分别较NG样地分别增加了8.33%、10.42%和29.17%(5月), 2.94%、11.76%和91.18%(8月)及8.82%, 14.71%和61.76%(10月), 而只有CG样地的土壤容重较NG样地变化显著(P<0.05);5月RG、TG和CG样地的土壤紧实度较NG样地均显著增加(P<0.05), 8月和10月4种放牧管理模式下土壤紧实度的大小依次为CG>TG>RG>NG, 其中在CG样地显著高于其它样地(P<0.05)(图 1)。

|
| 图 1 不同放牧管理模式下土壤因子的变化 Fig. 1 The change of different grazing management remiges on soil factors |
不同放牧管理模式下高原鼢鼠的鼠丘形态特征变化存有显著差异, 5月放牧管理模式对高原鼢鼠鼠丘半径的影响不显著, 但8月和10月CG样地的鼠丘半径显著小于其它放牧管理模式下鼠丘半径(P<0.05);RG、TG和CG样地的鼠丘体积在5月较NG样地分别降低了20.09%、14.62%和31.72%, 其中在NG和CG样地存在显著差异(P<0.05), 8月鼠丘体积在RG和CG样地显著小于NG和TG样地(P<0.05), 10月鼠丘体积在NG、RG、TG和CG样地依次减小且在CG样地较其它样地变化显著(P<0.05);5月鼠丘高度在NG、RG、TG和CG样地依次减小, 其中CG和NG样地存在显著差异(P<0.05), 8月RG和CG样地的鼠丘高度较NG和TG样地的鼠丘高度显著减小(P<0.05), 10月CG样地鼠丘高度显著小于其它样地的鼠丘高度(P<0.05);鼠丘表面积在5月和8月的大小依次为NG>TG>RG>CG, 其中在CG样地显著小于NG样地, 而10月在NG、RG、TG和CG样地依次减小, 其中在CG样地显著小于其它样地(P<0.05)(图 2)。

|
| 图 2 不同放牧管理模式下高原鼢鼠鼠丘形态特征 Fig. 2 The mound morphological characteristics of mound in the plateau zokor under different grazing management regimes |
以土壤水分、土壤容重、土壤紧实度和地下生物量作为解释变量, 鼠丘体积和表面积作为响应变量进行RDA分析(图 3)。NG样地的地下生物量对鼠丘体积的影响较大, CT样地各土壤因子和地下生物量对鼠丘体积和表面积的影响均较小, RG样地的土壤水分、土壤容重和地下生物量对鼠丘表面积和体积的影响较大, CG样地的土壤水分、容重和地下生物量对鼠丘表面积影响较大, 而土壤紧实度对鼠丘体积影响较大。

|
| 图 3 鼠丘表面积和体积与土壤因子、地下生物量的RDA排序图 Fig. 3 RDA ranking between surface area and volumes of mounds and soil factors, underground biomass MV: 鼠丘体积, Volumes of mounds;MSA: 鼠丘表面积, Surface area of mounds;SM: 土壤水分, Soil moisture;SBD: 土壤容重, Soil bulk density;SC: 土壤紧实度, UGB: 地下生物量, Underground biomass; RDA: 冗余分析Redundancy analysis |
不同放牧管理模式下高原鼢鼠鼠丘大小与土壤因子、地下生物量RDA结果(表 2)表明, NG样地土壤因子及地下生物量只能解释9.38%的鼠丘大小变化, 2轴累计解释99.99%的土壤因子及地下生物量与鼠丘特征之间的关系;生长季休牧第1排序轴和第2排序轴分别解释39.87%和1.42%的鼠丘大小变化, 土壤水分、土壤容重、土壤紧实度和地下生物量只能解释41.29%的鼠丘特征变化, 还有58.71%是其不能解释的。第1排序轴和第2排序轴解释分别解释96.57%和3.43%的土壤因子及地下生物量与鼠丘特征之间的关系, 2轴累计解释100.00%;传统放牧样地RDA分析的2轴累计解释10.04%的鼠丘大小变化, 第1排序轴和第2排序轴分别解释97.10%和2.89%的土壤因子及地下生物量与鼠丘特征之间的关系;连续放牧样地RDA分析的2轴累计解释27.84%的鼠丘大小变化, 第1排序轴和第2排序轴分别解释94.73%和5.26%的土壤因子及地下生物量与鼠丘特征之间的关系, 由此可说明土壤因子及地下生物量的变化并不能完全解释鼠丘大小的变化, 但排序轴能很好地反映土壤因子及地下生物量与鼠丘的大小之间的变化关系。生长季休牧下蒙特卡罗置换检验结果显著(P<0.05), 禁牧、传统放牧和连续放牧下蒙特卡罗置换检验结果不显著(P>0.05)。
| 参数 Parameter |
禁牧 No grazing |
生长季休牧 Rest grazing in growing season |
传统放牧 Traditional grazing |
连续放牧 Continuous grazing |
|||||||
| RDA1 | RDA2 | RDA1 | RDA2 | RDA1 | RDA2 | RDA1 | RDA2 | ||||
| 特征值Eigenvalues | 0.16 | 0.02 | 0.80 | 0.03 | 0.20 | 0.01 | 0.53 | 0.03 | |||
| 鼠丘大小变化百分比/% Percentage change in mound size |
8.21 | 1.17 | 39.87 | 1.42 | 9.75 | 0.29 | 26.37 | 1.47 | |||
| 鼠丘大小变化累积百分比/% Cumulative percentage change in mound size |
8.21 | 9.38 | 39.87 | 41.29 | 9.75 | 10.04 | 26.37 | 27.84 | |||
| 土壤因子、地下生物量-鼠丘大小变化百分比/% Soil factors, underground biomass-Percentage change in mound size |
87.54 | 12.45 | 96.57 | 3.42 | 97.10 | 2.89 | 94.73 | 5.26 | |||
| 土壤因子、地下生物量-鼠丘大小累积变化百分比/% Soil factors, underground biomass-cumulative percentage change in mound size |
87.54 | 99.99 | 96.57 | 100.00 | 97.10 | 99.99 | 94.73 | 99.99 | |||
| 蒙特卡罗置换检验Monte Carlo replacement test | P=0.700 | P=0.018 | P=0.659 | P=0.107 | |||||||
| RDA: 冗余分析, Redundancy analysis | |||||||||||
在各土壤因子及地下生物量与鼠丘特征的RDA排序相关系数中, 放牧管理模式影响下土壤水分、土壤容重、土壤紧实度和地下生物量对鼠丘特征的影响不同, 按照决定系数r2排序, 禁牧样地的结果为土壤紧实度>土壤容重>土壤水分>地下生物量, 其中土壤容重对鼠丘特征的影响显著(P<0.05), 土壤紧实度对鼠丘特征的影响极显著(P<0.01);生长季休牧和传统放牧样地的结果均为地下生物量>土壤紧实度>土壤水分>土壤容重, 其中土壤紧实度对鼠丘特征的影响显著(P<0.05), 地下生物量对鼠丘特征的影响极显著(P<0.01);传统放牧样地各因子对鼠丘特征的影响均不显著(P>0.05)。连续放牧样地的结果为土壤紧实度>地下生物量>土壤水分>土壤容重, 其中土壤紧实度, 地下生物量和土壤水分对鼠丘特征的影响显著(P<0.05)(表 3)。
| 环境因子 Environmental factors |
禁牧 No grazing |
生长季休牧 Rest grazing in growing season |
传统放牧 Traditional grazing |
连续放牧 Continuous grazing |
|||||||
| r2 | P | r2 | P | r2 | P | r2 | P | ||||
| 土壤水分Soil moisture | 0.2124 | 0.059 | 0.0199 | 0.808 | 0.0451 | 0.568 | 0.2288 | 0.038 | |||
| 土壤容重Soil bulk density | 0.2191 | 0.044 | 0.0125 | 0.876 | 0.0396 | 0.639 | 0.1911 | 0.080 | |||
| 地下生物量Underground biomass | 0.0756 | 0.360 | 0.4956 | 0.006 | 0.0674 | 0.433 | 0.2446 | 0.039 | |||
| 土壤紧实度Soil compactness | 0.4630 | 0.001 | 0.2742 | 0.017 | 0.0534 | 0.499 | 0.3173 | 0.010 | |||
相关性分析结果表明, NG样地鼠丘体积和表面积与莎草科植物的地上生物量呈极显著正相关关系(P<0.01);RG样地鼠丘体积和表面积与豆科植物的地上生物量呈极显著正相关关系(P<0.01), 鼠丘表面积与莎草科植物的地上生物量呈显著负相关关系(P<0.05);TG样地鼠丘体积和表面积与禾本科和豆科植物的地上生物量呈极显著负相关关系(P<0.01);CG样地的鼠丘体积和表面积与莎草科植物的地上生物量呈极显著正相关关系(P<0.01), 鼠丘体积与禾本科和豆科植物的地上生物量呈显著负相关关系(P<0.05)(表 4)。
| 功能群 Functional group |
鼠丘体积Volumes of mounds | 鼠丘表面积Surface area of mounds | |||||||
| NG | RG | TG | CG | NG | RG | TG | CG | ||
| GRA | 0.255 | -0.135 | -0.994** | -0.871* | 0.160 | -0.042 | -0.970** | -0.789 | |
| CYP | 0.965** | -0.791 | 0.696 | 0.984** | 0.935** | -0.844* | 0.593 | 1.000** | |
| LEG | -0.704 | 0.997** | -0.994** | -0.871* | -0.632 | 0.986** | -0.970** | -0.789 | |
| MIS | 0.710 | -0.135 | 0.593 | 0.010 | 0.775 | -0.042 | 0.696 | -0.137 | |
| **表示0.01水平上的相关性显著, *表示0.05水平上的相关性显著 | |||||||||
放牧显著影响了高原鼢鼠的微生境特征。从植被地上、地下生物量的变化上可看出不同放牧管理模式下高原鼢鼠食物资源的丰度变化, 而其土壤水分、容重和紧实度是高原鼢鼠取食活动的能量消耗, 代表了获取食物的代价等。与NG相比, 其它放牧管理模式下禾本科和豆科植物的地上生物量显著减少(P<0.05), 而杂草类植物(TG管理模式除外)的地上生物量显著增加(P<0.05);0—20cm层的地下生物量(除8月10—20cm层的地下生物量)在NG→CG管理模式下依次减少, 该结果与前人的研究结果相似[24]。放牧家畜的采食和践踏能够使植物变低矮, 影响植物的地上和地下部分生长发育, 同时也使其物种组成发生变化, 优质牧草减少, 杂草种类和数量增加[4], 加之放牧管理模式是在放牧时间和空间上的调整, 而RG、TG和CG均会在植物返青时期进行放牧, 此时植物刚生长露出地面, 光合强度较弱, 而家畜啃食和践踏破坏了草地植物的地上部分, 进而影响植物的生长发育和草地恢复, 导致放牧后植物的生物量变小[25]。本研究发现, NG→CG管理模式下土壤容重和紧实度依次增加, 土壤水分在CG模式下显著高于NG模式, 家畜的践踏是土壤结构发生改变, 增加了土壤的紧实程度, 植物的组成和类型发生改变, 而高密度和细根构型的草毡表层会增加土壤的紧实状况[26]。与本研究不同的是, 有研究发现放牧会降低土壤水分含量[27], 因为土壤水分会受降水量、土壤紧实度和植被类型等多个因素的影响, 而草地土壤生态系统本身也具备一定的复杂性和弹性, 这使得不同的放牧压力对草地土壤特性的影响不同, 也与放牧压力、放牧家畜的种类与结构、草地生态系统的类型有关[28]。
3.2 放牧管理模式对高原鼢鼠鼠丘形态特征的影响放牧显著影响了高原鼢鼠鼠丘的形态特征。RG、TG和CG管理模式下高原鼢鼠的鼠丘半径、表面积和体积较NG均变小。这与放牧家畜践踏增加了土壤紧实度、土壤容重和水分, 减少了植物地下生物量有关, 放牧导致了高原鼢鼠的食物资源变少, 挖掘难度加大, 能量消耗增加, 向地表的推土量变少, 鼠丘变小[29]。有研究发现, 高原鼢鼠比较喜欢土壤松软而杂草较多的环境[30], 放牧过程中家畜的践踏和采食使得优质牧草(禾本科)减少, 杂草增多, 使高原鼢鼠的食物结构发生改变[29], 进一步导致高原鼢鼠的分布发生改变, 其对资源的竞争也是导致高原鼢鼠鼠丘的形态特征出现差异的原因之一。本研究中5月、8月和10月鼠丘的半径、体积和表面积均在NG样地最大, CG样地最小且差异显著(P<0.05)。放牧管理模式对鼠丘形态特征的影响存在着季节性差异, 这可能是高原鼢鼠在不同季节的挖掘目的不同所致[18], 5月高原鼢鼠刚经历过越冬且处于繁殖期, 需要更多的能量补充, 此时食物资源的分布和数量及配偶的分布是影响高原鼢鼠挖掘的主要因素[31], 而0—20cm土层地下生物量与鼠丘的形态特征具有一致的变化规律也对此进行了更好的诠释。8月高原鼢鼠处于育幼时期, 其活动范围相对较小, 而且充足的雨水和较高的温度使得植物生长比较茂盛, 根系发达, 食物资源充足[18];10月高原鼢鼠由于受高寒气候的影响, 草原植被逐渐进入枯黄期, 高原鼢鼠为储备长达5个月越冬期的食物, 因此食物(地下生物量)是影响该季节高原鼢鼠挖掘的主要因素[20]。总之, 家畜的践踏和干扰及季节的更替引起土壤结构、土壤性质和植物物种组成发生改变, 会进一步导致高原鼢鼠在挖掘过程中的耗能和代价增加, 食物资源和分布结构的改变, 是导致其鼠丘形态特征变化的原因之一。然而, 在高原鼢鼠掘洞造丘过程中, 还受到放牧家畜的卧息[15]、天敌、种内和种间的竞争[32]及自身的生物学特性[33]等多个因素的影响。鼢鼠本身的生活方式也可能导致高原鼢鼠鼠丘的形态特征发生变化, 如群居生活的高原鼢鼠鼠丘可能较独居生活的鼢鼠鼠丘大。另外, 高寒草甸植物地上部分能够拦截和蓄积降雨, 也能够抵抗风力, 植物地下根系具有固结土壤和增加土壤入渗的作用[34], 而本研究所选取的均为2个月以内新鼠丘, 植被盖度低且土质松软, 易受家畜的践踏和风雨侵蚀等因素的影响[12], 导致其形态特征发生改变。
3.3 放牧管理模式对鼠丘、环境因子及地上、地下生物量关系的影响高原鼢鼠是青藏高原上的优势物种之一, 常年营地下洞道生活, 其挖掘洞穴及推土造丘与土壤环境和植被特征等的变化息息相关[9]。相关性分析结果表明, NG和CG样地的莎草科植物、RG样地的豆科植物及TG样地的和本科和豆科植物的地上生物量显著影响了高原鼢鼠鼠丘形态特征, 由此可说明不同放牧管理模式下植物的演替、分布和组成结构对高原鼢鼠的洞道挖掘和推土造丘具有显著影响。采用冗余分析法分析高原鼢鼠鼠丘表面积和体积与土壤因子及地下生物量之间的关系发现, 2个排序轴几乎能够全部解释土壤因子、地下生物量与鼠丘形态特征之间的关系, 说明不同放牧管理措施导致土壤紧实度、土壤水分、土壤容重和地下生物量的变化是影响高原鼢鼠推土造丘的关键因子。同时, 本研究还发现放牧管理模式影响下各土壤因子及地下生物量对鼠丘特征的影响程度不同, NG和CG样地土壤紧实度对鼠丘特征的影响最大, RG和TG样地的地下生物量对鼠丘特征的影响最大, 这可能是放牧制度是放牧在时间和空间上的调整[17], 土壤紧实度和地下生物量是影响高原鼢鼠鼠丘形态特征的两个主要因素。而NG样地植物生长茂盛, 地下根系充足, 高原鼢鼠能够获取足量的植物, 因而地下生物量对高原鼢鼠挖掘及推土造丘活动的影响相对较小;CG样地属于全年放牧, 放牧时间较长, 其由于家畜践踏, 使得其土壤变得紧实, 增加了高原鼢鼠在挖掘过程中的能量消耗, 使得其挖掘难度增加[18], 而RG和TG样地的放牧时间分别集中在10月到次年5月和7月, 此时高原鼢鼠正在进行食物储备和繁殖育幼时期, 需要大量的食物储备, 此时进行放牧, 由于家畜的践踏和啃食, 使得植被遭到一定的破坏, 对高原鼢鼠的觅食活动造成影响。由此进一步可以说明高原鼢鼠的挖掘行为与其采食、储食及土壤性状的变化存在一定的关联性[35]。另外, 前人研究发现, 高原鼢鼠的鼠丘特征的变化与除与其采食、储食等有关外, 还与高原鼢鼠的繁殖特性、年龄结构和性别比例等有关联[18, 31], 但本研究只考虑了放牧下鼠丘大小与土壤因子及植被地下生物量之间的关系, 并未考虑其它环境和高原鼢鼠的繁殖特性、年龄结构和性别比例等其它因素的影响, 在今后的研究过程中, 我们会着重考虑这些因素对高原鼢鼠的影响。
4 结论放牧管理模式对高原鼢鼠鼠丘有显著影响, 且具有季节性差异。与禁牧相比, 生长季休牧、传统放牧和连续放牧样地的鼠丘半径、高度、表面积和体积有变小趋势, 其中连续放牧较其它样地变化显著(P<0.05);冗余分析结合蒙特卡罗置换检验结果发现, 土壤水分、容重、紧实度和地下生物量几乎能够全部解释了鼠丘特征, 放牧管理模式下土壤紧实度和地下生物量对高原鼢鼠鼠丘的特征具有显著影响。
| [1] |
Courtois D R, Perryman B L, Hussein H S. Vegetation change after 65 years of grazing and grazing exclusion. Rangeland Ecology & Management, 2004, 57(6): 574-582. |
| [2] |
彭祺, 王宁. 不同放牧制度对草地植被的影响. 农业科学研究, 2005, 26(1): 27-30. DOI:10.3969/j.issn.1673-0747.2005.01.007 |
| [3] |
Wu R X, Wei X T, Liu K S, Zhong M Y, Liu Y H, Pan D, Shao X Q. Nutrient uptake and allocation by plants in recent mounds created by subterranean rodent, plateau zokor Eospalax baileyi. Polish Journal of Ecology, 2017, 65(1): 132-143. DOI:10.3161/15052249PJE2017.65.1.012 |
| [4] |
刘伟, 周立, 王溪. 不同放牧强度对植物及啮齿动物作用的研究. 生态学报, 1999, 19(3): 376-382. DOI:10.3321/j.issn:1000-0933.1999.03.015 |
| [5] |
苏军虎, 南志标, 纪维红. 家畜放牧对草地啮齿动物影响的研究进展. 草业学报, 2016, 25(11): 136-148. DOI:10.11686/cyxb2015587 |
| [6] |
武晓东, 阿娟, 付和平, 金珩. 人为不同干扰条件下荒漠啮齿动物群落格局的动态特征——动物与植物群落的典型相关分析. 生态学报, 2008, 28(12): 5999-6017. DOI:10.3321/j.issn:1000-0933.2008.12.028 |
| [7] |
樊才睿, 李畅游, 孙标, 史小红, 赵胜男, 付尧, 王静洁. 不同放牧制度草场产流产沙过程模拟试验. 水土保持学报, 2016, 30(1): 47-53. |
| [8] |
李文, 曹文侠, 刘皓栋, 李小龙, 徐长林, 师尚礼, 冯今, 周传猛. 不同放牧管理模式对高寒草甸草原土壤呼吸特征的影响. 草业学报, 2015, 24(10): 22-32. DOI:10.11686/cyxb2015152 |
| [9] |
鲍根生, 王宏生, 曾辉, 卡着才让, 马志贵, 刘生财. 不同形成时间高原鼢鼠鼠丘土壤养分分配规律. 生态学报, 2016, 36(7): 1824-1831. |
| [10] |
Zhang Y M, Zhang Z B, Liu J J. Burrowing rodents as ecosystem engineers: the ecology and management of plateau zokors Myospalax fontanierii in alpine meadow ecosystems on the Tibetan Plateau. Mammal Review, 2003, 33(3/4): 284-294. |
| [11] |
王玉琴, 鲍根生, 王宏生, 曾辉, 李继鹏. 不同放牧制度对高原鼢鼠活动区高寒草地群落结构影响. 草地学报, 2018, 26(1): 134-141. |
| [12] |
胡媛, 查轩, 黄少燕. 重要水源地东圳库区土地利用与水土流失特征空间关系分析. 水土保持研究, 2012, 19(2): 53-56. |
| [13] |
刘江, 徐先英, 张荣娟, 丁爱强, 付全贵, 赵鹏. 人工梭梭林大沙鼠(Rhombomys opimus)土丘斑块与鼠洞塌陷斑块破碎度及点格局. 中国沙漠, 2018, 38(3): 610-618. |
| [14] |
邓云飞. 苦竹+牛鞭草模式中草地斑块的土壤养分动态及牧草的生长特性[D]. 雅安: 四川农业大学, 2011.
|
| [15] |
Torre I, Díaz M, Martínez-Padilla J, Bonal R, Viñuela J, Fargallo J A. Cattle grazing, raptor abundance and small mammal communities in Mediterranean grasslands. Basic and Applied Ecology, 2007, 8(6): 565-575. DOI:10.1016/j.baae.2006.09.016 |
| [16] |
何俊龄, 张金沙, 杨莹博, 邓汉刚, 魏进招, 张卫国. 高原鼢鼠土丘空间格局及主要特征研究. 草业学报, 2006, 15(1): 107-112. DOI:10.3321/j.issn:1004-5759.2006.01.018 |
| [17] |
张倩, 杨晶, 姚宝辉, 蔡志远, 孙小妹, 王缠, 郭怀亮, 谭宇尘, 苏军虎. 放牧管理模式对高寒草甸鼢鼠鼠丘群落演替的影响. 生态学报, 2020, 40(8): 2802-2811. |
| [18] |
王大伟, 郭永旺, 刘祖华, 杨文超, 张德勇, 王福美, 李宁, 刘晓辉. 川西北草原高原鼢鼠种群季节性特征与分布特点. 植物保护, 2013, 39(4): 124-128. DOI:10.3969/j.issn.0529-1542.2013.04.026 |
| [19] |
杨晶, 张倩, 姚宝辉, 蔡志远, 王缠, 郭怀亮, 苏军虎. 高原鼢鼠扰动对高寒草甸土壤理化特性及植物生物量的影响. 草地学报, 2020, 28(2): 492-499. |
| [20] |
王权业, 樊乃昌. 高原鼢鼠(Myospalax baileyi)的挖掘活动及其种群数量统计方法的探讨. 兽类学报, 1987(4): 283-290. |
| [21] |
鲍士旦. 土壤农化分析. 北京: 中国农业出版社, 1999: 42-50.
|
| [22] |
Tilman D, Reich P B, Knops J M H. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature, 2006, 441(7093): 629-632. DOI:10.1038/nature04742 |
| [23] |
赖江山, 米湘成. 基于Vegan软件包的生态学数据排序分析//第九届全国生物多样性保护与持续利用研讨会论文集. 厦门: 国际生物多样性计划中国委员会, 中国科学院生命科学与生物技术局, 2010: 332-343.
|
| [24] |
许宏斌, 辛晓平, 宝音陶格涛, 闫瑞瑞, 王旭, 陈宝瑞, 金东艳, 姚静, 刘志英. 放牧对呼伦贝尔羊草草甸草原生物量分布的影响. 草地学报, 2020, 28(3): 768-774. |
| [25] |
崔树娟, 布仁巴音, 朱小雪, 白玲, 汪诗平. 不同季节适度放牧对高寒草甸植物群落特征的影响. 西北植物学报, 2014, 34(2): 349-357. |
| [26] |
樊博, 林丽, 曹广民, 柯浔, 李以康, 杜岩功, 郭小伟, 李茜, 钱大文, 兰玉婷, 周春丽. 不同演替状态下高寒草甸土壤物理性质与植物根系的相互关系. 生态学报, 2020, 40(7): 2300-2309. |
| [27] |
靳茗茗, 徐增让, 成升魁. 藏北高寒草地植被和土壤对不同放牧强度的响应. 生态学报, 2020, 40(23): 8753-8762. |
| [28] |
张成霞, 南志标. 放牧对草地土壤理化特性影响的研究进展. 草业学报, 2010, 19(4): 204-211. |
| [29] |
张兴禄, 李广. 高原鼠兔和高原鼢鼠在高寒草甸生态系统的作用. 草业科学, 2015, 32(5): 816-822. |
| [30] |
于海玲, 樊江文, 李愈哲, 史文娇. 高原鼢鼠干扰对三江源区高寒草甸群落特征的影响. 应用生态学报, 2018, 29(6): 1902-1910. |
| [31] |
王莹, 魏代红, 张卫国, 卫万荣. 高原鼢鼠地上土丘性状季节性差异. 草业科学, 2018, 35(11): 2738-2743. DOI:10.11829/j.issn.1001-0629.2017-0668 |
| [32] |
田富, 程云湘, 周国利, 马青青, 申波, 曲久, 常生华. 高原鼠兔洞穴密度与高寒草甸植物群落结构以及土壤因子的关系. 草业科学, 2019, 36(4): 1094-1104. |
| [33] |
姬程鹏. 祁连山东段高原鼢鼠暖季活动节律及巢域面积变化研究[D]. 兰州: 甘肃农业大学, 2017.
|
| [34] |
刘忆轩, 李多才, 侯扶江. 甘肃马鹿春秋季放牧对高寒草原土壤理化性质的影响. 草业科学, 2019, 36(2): 273-283. |
| [35] |
鲁庆彬, 张阳, 周材权. 秦岭鼢鼠的洞穴选择与危害防控. 生态学报, 2011, 31(7): 1993-2001. |