 2022, Vol. 42
2022, Vol. 42文章信息
- 李萌, 谭许脉, 肖纳, 徐浩成, 莫雪青, 张文, 尤业明, 黄雪蔓
- LI Meng, TAN Xumai, XIAO Na, XU Haocheng, MO Xueqing, ZHANG Wen, YOU Yeming, HUANG Xueman
- 引种降香黄檀和顶果木对石漠化山区土壤磷素转化及其有效性的影响
- Effects of introducing Dalbergia odorifera and Acricarpus fraxinifolius on soil phosphorus transformation and availability in rocky desertification mountainous regions
- 生态学报. 2022, 42(4): 1594-1605
- Acta Ecologica Sinica. 2022, 42(4): 1594-1605
- http://dx.doi.org/10.5846/stxb202012223252
-
文章历史
- 收稿日期: 2020-12-22
- 网络出版日期: 2021-11-04





2. 广西友谊关森林生态系统定位观测研究站, 凭祥 532600
2. Guangxi Youyiguang Forest Ecosystem Research Station, Pingxiang 532600, China
土壤养分对植物的生长起着关键作用, 直接影响植物群落的组成、稳定和演替[1]。磷是植物生长发育的必需营养元素之一, 与植物体内的各种代谢过程密不可分[2]。植物所利用的磷元素主要来源于土壤, 但土壤中的磷素大多与其他物质相结合, 以化合状态存在, 很难被植物直接吸收利用[3]。土壤中无机磷的含量、存在形态及转化过程等直接影响磷的生物有效性[4], 因此如何提高土壤磷素利用率和加快磷素循环已成为国内外许多科学家广泛关注的热点。
土壤微生物是陆地生态系统中的重要组成部分, 也是生态系统功能的重要驱动者, 并且能敏锐地感知土壤中养分含量的变化[5-6]。土壤微生物生物量是土壤有机质中最为活跃的部分, 其含量高低可以表征土壤肥力的变化水平, 对维持土壤生态系统平衡有重要意义, 其变化能敏感反映出土壤养分有效性的变化和有效表征土壤有机物质的代谢强度, 并受土壤pH、温度、透气性等环境因子影响[7-10]。微生物量一般包括微生物量碳、微生物量氮和微生物量磷。
岩溶地区是地球表层系统的重要组成部分, 全球岩溶区面积约5.1亿km2, 大约占地球面积的10% [11]。岩溶区生态环境恶劣, 土壤贫瘠, 水土流失严重, 植被覆盖率低[12]。我国西南岩溶区是石漠化最为严重的区域, 而且石漠化的面积呈现增加的趋势, 且岩溶区生态系统功能急剧退化, 严重影响该区域经济社会以及环境的可持续发展。此外, 岩溶区成土过程缓慢, 土层浅薄, 生态系统土壤氮素储量远低于非岩溶区, 是典型的生态脆弱带[13-14]。有研究发现, 岩溶区退化生态系统恢复初期主要受氮限制, 演替后期主要受磷限制, 而在恢复中期, 则受氮、磷共同限制[15]。土壤养分循环相互耦合[16], 氮和磷作为生命元素驱动着其他养分元素的转化和周转, 是养分循环中的核心部分[17]。近年来, 国家先后启动实施一系列重大生态工程进行石漠化综合治理, 其中植被恢复作为一项重要的生态治理措施, 由于其不同恢复模式下植物种类及其配置的不同, 必然导致凋落物凋落规律、数量和组成, 以及林地环境等因素的不同, 从而对土壤磷素产生不同的影响[4, 15, 18]。虽然已有许多国内外学者利用不同方法和途径, 研究了有关不同的植物、恢复方式和土地利用类型等对岩溶区土壤磷素的影响[4, 15, 19], 但到目前为止, 对不同固氮树种及其种植模式下的石漠化山区土壤无机磷组分变化规律的认识还存在很大的不足, 这将影响我们对石漠化山区土壤养分退化成因及其发生机制、养分的维持及演变规律的认识。因此, 在石漠化山区植被恢复重建中开展关于引进优良固氮树种及其种植模式对其土壤磷有效性及无机磷组分变化的研究, 对于改善石漠化山区土壤质量, 缓解其土壤磷素对植被恢复的限制作用具有重要的现实意义。
本研究选取广西石漠化最为严重的马山县作为研究区域, 对研究石漠化区土壤肥力恢复及其维持机制具有较强的代表性。选择不同固氮树种及其种植模式恢复起来的纯林和混交林作为研究对象, 采用野外监测和室内分析试验相结合的方法, 重点探究不同固氮树种及种植模式对石漠化区表层土壤磷素转化和有效性的影响及其驱动因子。通过对上述问题的研究, 为定向调控岩溶区表层土壤磷循环过程, 长期提高该区土壤磷有效性的树种和种植模式的选择等方面提供重要的科学依据, 促进岩溶区生态、经济和社会可持续发展。
1 材料与方法 1.1 研究地概况研究区域位于广西壮族自治区南宁市马山县(23°24′—24°20′N, 107°41′ —108°29′E), 地处广西中部略偏西位置, 居红水河中段南岸, 大明山北麓。马山县的地貌以山区丘陵地形为主, 东西部为石山区, 西南部和中部为土岭丘陵。该地属于南亚热带季风型气候, 年均气温21.3℃, 年均降雨量1667.1 mm;夏季多暴雨, 易发生洪涝, 春秋两季雨水少, 易造成干旱;相对湿度76%, 年平均无霜期343 d。土壤类型主要以碳酸盐岩发育的石灰土为主, 呈弱碱性, 石粒含量高, 质地粘重, 土壤层薄。原生植被属于南亚热带季风常绿阔叶林, 但由于长期过度垦殖、放牧、樵采等, 原生性森林早已消失, 导致基岩裸露, 出现大面积石漠化景观。现在境内岩溶地貌发育, 生态环境脆弱, 是广西石漠化最严重的地区之一。
1.2 试验设计和土壤样品采集本研究的试验样地是依托中央财政林业科技推广示范资金跨区域重点推广示范项目“广西马山县石山(石灰岩)地区造林绿化优良速生树种栽培技术推广示范”所建的平台上进行。在对研究区的石漠化情况进行全面调查的基础上, 选择马山县白山镇民族村约50 hm2典型的石漠化区域作为植被恢复试验地, 该试验地稀疏分布的灌木主要有黄荆条、红背山麻杆、地桃花、黄花稔等;草本植物主要有小花露籽草、蔓生莠竹、卷柏、肾蕨和扇叶铁线蕨等, 无乔木分布。根据石漠化区因地制宜、耐干旱和热量变化等的树种选择原则, 于2011年2月选择石山优良造林的珍贵豆科树种降香黄檀(Dalbergia odorifera T. Chen)和豆科树种顶果木(Acrocarpus fraxinifolius Wight ex Arn. In Mag.)进行造林试验, 分别构建了降香黄檀纯林(The pure plantation of Dalbergia odorifera)、顶果木纯林(The pure plantation of Acricarpus fraxinifolius)和降香黄檀/顶果木混交林(The mixed plantation of Dalbergia odorifera and Acricarpus fraxinifolius)3种林分类型。同时以相邻的无植被恢复的石山灌丛作为对照(CK)。因顶果木是优良速生树种, 适当提高顶果木在混交林中的比例, 可以为珍贵树种降香黄檀后期生长过程中干型的塑造(如: 减少主干分杈)和生长环境的构建提供更有利的冠层结构, 并保障混交林中2个树种能够更好地生长。因此, 在混交林的树种比例设置上, 采取降香黄檀: 顶果木为3:7的混交模式。造林时, 降香黄檀的苗高约60—70 cm, 地径约为0.3—0.4 cm;顶果木的苗高约25—30 cm, 地径约为0.4—0.5 cm。各林分类型具有相似的立地条件(海拔、土层厚度、土壤类型)和抚育措施等, 林分建立后, 每年的6—8月对各样地进行林分调查, 记录样地内林木的树高、胸径和冠幅等。
根据随机区组设计原则, 本研究于2019年7月初(植被生长恢复8年后), 在整个试验区内随机选择5个实验区组, 区组之间的距离不低于200 m。其中, 每个区组内均包含4种不同立地类型(具体为对照组、降香黄檀纯林、顶果木纯林和降香黄檀/顶果木混交林), 区组内每种立地类型分别设置一个400 m2 (20 m×20 m)的样方。整个实验共设置了20个实验样方, 并对每个实验样方进行调查, 其基本概况详见表 1。于2019年8月对每个20 m×20 m样方进行土壤取样, 在每个样方内随机布设10个取样点, 去除表面的杂质后用土钻(直径10 cm)取0—10 cm深的表层土壤, 每个取样样方的土壤经充分混匀后作为一个土壤样品。所有土壤样品当天带回实验室后立即剔除沙石、植物根茎等杂质, 过2 mm土壤筛后分为两份。一份存放在4℃冰箱中用于测定土壤速效养分和微生物生物量碳、氮和磷;另一份自然风干后经进一步研磨, 全部过0.149 mm土壤筛, 用于测定土壤全量养分和pH值。
| 样地类型 Site types |
海拔 Altitude/m |
土层厚度 Soil thickness/cm |
土壤类型 Soil type |
密度 SD/(株/hm2) |
胸径 DBH/cm |
树高 Tree hight /m |
胸高断面积 BA/(m2/hm2) |
| 对照Control | 290—310 | 30—40 | 石灰土 | NA | NA | NA | NA |
| 降香黄檀纯林 The pure plantation of dalbergia odorifera |
290—310 | 30—40 | 石灰土 | 485±17 | 7.9±0.4 | 8.6±0.6 | 2.5±0.3 |
| 顶果木纯林 The pure plantation of acricarpus fraxinifolius |
290—310 | 30—40 | 石灰土 | 455±17 | 7.4±0.5 | 9.4±0.5 | 2.2±0.3 |
| 降香黄檀/顶果木混交林 The mixed plantation of dalbergia odorifera and Acricarpus fraxinifolius |
290—310 | 30—40 | 石灰土 | 465±19 | 7.8±0.4 | 10.1±0.6 | 2.4±0.3 |
| SD:密度Density;DBH:胸径Diameter at breast height;BA:胸高断面积Basal area. 数据=平均值±标准误,n=5 | |||||||
土壤理化性质的测定方法参照鲍士旦主编的《土壤农化分析》进行[20]。称取新鲜土壤在105℃下烘干至恒重, 根据其质量减少量计算, 获得土壤含水量;土壤pH值采用pH计测定(土∶水=1 ∶ 2.5, w/v);土壤有机质(SOM)采用K2Cr2O7-H2SO4氧化法测定;土壤全氮(TN)采用凯氏定氮法测定含量;称取相当于10 g干土的鲜土, 经过50 mL CaCl2溶液(0.01 mol/L) 浸提后, 在流动分析仪上测定其铵态氮(NH4+-N)和硝态氮(NO3--N)含量。土壤总磷(TP)采用硫酸消煮法提取, 钼锑抗比色法测定[21]。速效磷(AP)采用0.5 mol/L的NaHCO3溶液浸提30 min, 然后采用钼锑抗比色法测定[21]。
土壤微生物量碳、氮和磷(MBC、MBN和MBP)采用氯仿熏蒸法提取[22-23]。MBC和MBN用K2SO4(0.5 mol/L)浸提, 然后用TOC/TN同步分析仪(Multi N/C 3100)测定。MBP用NaHCO3(0.5 mol/L)浸提, 然后采用钼锑抗比色法测定[21]。
石灰性土壤无机磷的形态分级采用蒋柏藩和顾益初的方法[24-25]分为6级。Ca2-P采用NaHCO3(0. 25 mol/L)浸提;Ca8-P采用NH4OAc(0. 5 mol/L)浸提;Ca10-P采用1/2 H2SO4(0. 5 mol/L)浸提;Al-P采用NH4F(0. 5 mol/L)浸提;Fe-P采用NaOH(0. 1 mol/L)+ Na2CO3(0. 1 mol/L)浸提;O-P, 又称闭蓄态磷, 采用柠檬酸钠(0. 3 mol/L)+连二亚硫酸钠+ NaOH(0. 5 mol/L)浸提。有机磷=总磷-无机磷。
1.4 数据分析本研究采用土壤磷素活化系数(Phosphorus activation coefficient, 简称PAC)来衡量土壤全磷的有效性[4]。其中PAC的计算方式如下:
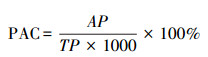
|
(1) |
式中, AP为速效磷(mg/kg);TP为总磷(g/kg)。
运用SPSS 24.0软件对数据进行单因素方差分析(one-way ANOVA), 采用最小显著差异法(LSD)比较均值之间的差异程度, 显著性水平全部设置为P<0.05。采用Canoco 4.5对不同林分的土壤无机磷组分和土壤性质进行冗余分析(RDA)。采用SigmaPlot 10.0进行绘图。
2 结果与分析 2.1 固氮树种对石漠化山区表层土壤性质的影响在石漠化山区引入固氮树种进行植被恢复8年后, SOM、NO3--N、AP、MBC、MBN和MBP含量均发生较大变化。其中, 降香黄檀/顶果木混交林、顶果木纯林和降香黄檀纯林的SOM含量与对照相比分别显著增加了53.7%、53.5%和49.9% (P<0.05)。降香黄檀/顶果木混交林土壤的NO3--N含量与降香黄檀纯林和对照相比较分别显著提高了85.8%和86.9% (P<0.05), 顶果木纯林土壤的NO3--N含量与降香黄檀纯林和对照相比较分别显著提高了58.7%和59.6% (P<0.05)。降香黄檀/顶果木混交林的AP含量比顶果木纯林、降香黄檀纯林和对照样地分别显著增加了62.3%, 154.7%和197.2% (P<0.05), 顶果木纯林的土壤AP含量比降香黄檀纯林和对照样地分别显著增加57.0%和83.2%(P<0.05)。与对照样地相比, 降香黄檀/顶果木混交林、顶果木纯林和降香黄檀纯林的土壤MBC含量分别显著提高了21.8%、16.6%和16.9% (P<0.05)。降香黄檀/顶果木混交林的土壤MBN含量与顶果木纯林、降香黄檀纯林和对照样地相比较分别显著提高了12.9%, 17.9%和17.6% (P<0.05)。降香黄檀/顶果木混交林的MBP含量比顶果木纯林、降香黄檀纯林和对照样地分别显著增加37.4%, 150.5%和175.9% (P<0.05), 顶果木纯林的土壤MBP含量比降香黄檀纯林和对照样地分别显著增加了82.4%和100.9%(P<0.05)(表 2)。
| 样地类型 Site types |
有机质SOM/ (g/kg) |
全氮TN/ (g/kg) |
铵态氮 NH4+-N/ (mg/kg) |
硝态氮 NO3--N/(mg/kg) |
总磷 TP/(g/kg) |
有效磷 AP/(mg/kg) |
| 对照Control | 39.8±3.9b | 4.5±0.9a | 14.3±2.1a | 18.3±2.1b | 0.3±0.0a | 6.1±0.6c |
| 降香黄檀纯林 The pure plantation of Dalbergia odorifera |
59.6±3.8a | 4.3±0.3a | 15.9±1.8a | 18.4±0.8b | 0.3±0.1a | 7.1±0.7c |
| 顶果木纯林 The pure plantation of Acricarpus fraxinifolius |
61.0±4.6a | 4.7±0.6a | 15.9±1.4a | 29.2±2.5a | 0.3±0.0a | 11.1±1.3b |
| 降香黄檀/顶果木混交林 The mixed plantation of dalbergia odorifera and acricarpus fraxinifolius |
61.1±4.1a | 4.3±0.5a | 18.4±2.1a | 34.2±3.7a | 0.3±0.1a | 18.0±2.6a |
| 样地类型 Site types |
有机磷 Po/(mg/kg) |
pH | 微生物量碳 MBC/(mg/kg) |
微生物量氮 MBN/(mg/kg) |
微生物量磷 MBP/(mg/kg) |
|
| 对照Control | 268.5±8.3a | 7.7±0.3a | 306.9±15.5b | 50.6±0.8b | 12.3±1.1c | |
| 降香黄檀纯林 The pure plantation of Dalbergia odorifera |
262.2±9.1a | 7.5±0.1a | 358.9±13.9a | 50.5±1.6b | 13.6±0.9c | |
| 顶果木纯林 The pure plantation of Acricarpus fraxinifolius |
255.5±9.5a | 7.6±0.2a | 357.8±5.6a | 52.8±1.4b | 24.7±1.9b | |
| 降香黄檀/顶果木混交林 The mixed plantation of dalbergia odorifera and acricarpus fraxinifolius |
243.1±5.5a | 7.0±0.2a | 373.9±10.4a | 59.6±1.3a | 33.9±1.4a | |
| SOM: 有机质Soil organic matter;TN: 全氮Total nitrogen;NH4+-N: 铵态氮Ammonium nitrogen;NO3--N: 硝态氮Nitrate nitrogen;TP: 总磷Total phosphorus;AP: 有效磷Available phosphorus;Po: 有机磷Organic phosphorus;MBC: 微生物量碳Soil microbial biomass carbon;MBN: 微生物量氮Soil microbial biomass nitrogen;MBP: 微生物量磷Soil microbial biomass phosphorus; 同列不同小写字母表示该指标在不同样地类型之间差异显著(P<0.05), 数据=平均值±标准误, n=5 | ||||||
土壤磷素活化系数(PAC), 即土壤有效磷与全磷的比值, 常被用来表示土壤全磷的有效性。PAC大于2%说明全磷中各形态的磷, 容易转化为有效磷, 供植物吸收利用, 反之则不利于全磷转化为有效磷。由图 1可知, 对照的PAC<2.0%, 而降香黄檀纯林、顶果木纯林和降香黄檀/顶果木混交林的PAC值均>2.0%, 分别为1.9%, 2.5%, 3.8%和6.7%。表明固氮树种的引入能有效提高石漠化山区土壤全磷的有效性, 其中以混交种植方式的效果最佳(P<0.05)。

|
| 图 1 不同样地类型中PAC的变化 Fig. 1 The changes of PAC in different site types PAC: 土壤磷素活化系数Phosphorus activation coefficient;CK: 对照Control; PD: 降香黄檀纯林The pure plantation of Dalbergia odorifera; PA: 顶果木纯林The pure plantation of Acricarpus fraxinifolius; MP: 降香黄檀/顶果木混交林The mixed plantation of Dalbergia odorifera and Acricarpus fraxinifolius; 不同小写字母代表差异显著(P<0.05), 数据=平均值±标准误, n=5 |
从图 2可以看出, NH4+-N、NO3--N、MBN和AP分别呈显著线性正相关关系(P<0.05), MBN与MBP呈极显著线性正相关关系(P<0.01), 而TN与TP和AP的关系不显著(P>0.05)。

|
| 图 2 土壤各种形态磷和氮之间的关系 Fig. 2 The relationship between phosphorus and nitrogen in various forms of soil |
由图 3可知, 降香黄檀/顶果木混交林、顶果木纯林和降香黄檀纯林的土壤Ca2-P含量与对照样地相比分别显著提高203.6%, 230.0%和105.4% (P<0.05);降香黄檀/顶果木混交林和顶果木纯林的土壤Ca2-P含量比降香黄檀纯林的分别显著提高47.8%和60.7% (P<0.05)。降香黄檀/顶果木混交林的土壤Ca8-P含量比顶果木纯林和对照样地分别显著提高28.8%和27.8% (P<0.05);顶果木纯林的土壤Ca8-P含量比降香黄檀纯林和对照样地分别显著增加30.6%和29.5% (P<0.05)。降香黄檀/顶果木混交林和顶果木纯林的土壤Ca10-P含量与对照样地相比分别显著增加49.0%和38.2% (P<0.05);然而, 3种固氮树种林分(降香黄檀/顶果木混交林, 顶果木纯林和降香黄檀纯林)的土壤Ca10-P含量两两之间差异不显著(P>0.05)。降香黄檀/顶果木混交林的土壤Al-P含量比降香黄檀纯林和对照样地分别显著增加760.7%和872.6% (P<0.05);顶果木纯林的土壤Al-P含量比降香黄檀纯林和对照样地分别显著提高599.4%和690.4% (P<0.05)。降香黄檀/顶果木混交林的土壤Fe-P含量比降香黄檀纯林和对照样地分别显著提高91.9%和116.3% (P<0.05);顶果木纯林的土壤Fe-P含量比降香黄檀纯林和对照样地分别显著提高80.1%和103.1% (P<0.05)。降香黄檀/顶果木混交林的土壤O-P含量比顶果木纯林、降香黄檀纯林和对照样地相比分别显著增加161.2%, 218.6%和517.0% (P<0.05);顶果木纯林的土壤O-P含量比降香黄檀纯林和对照样地相比分别显著增加46.1%和136.2% (P<0.05);对照样地与降香黄檀纯林的土壤O-P含量没有差异(P>0.05)。其中, 4种林分各无机磷形态含量大小顺序均为: Ca10-P>O-P>Al-P>Fe-P>Ca2-P>Ca8-P。

|
| 图 3 不同固氮树种及其种植模式对石漠化山区表层土壤无机磷组分的影响 Fig. 3 Effects of different N2-fixing species and planting patterns on inorganic phosphorus fractions in topsoil in karst areas 不同小写字母代表差异显著(P<0.05), 数据=平均值±标准误, n=5 |
以土壤各无机磷组分为因变量, 土壤基本性质为解释变量进行冗余分析(RDA)。结果表明, 模型筛选出的7个土壤基本性质共同解释了土壤无机磷组分变化的88% (表 3, 总解释率=条件效应的总和), 而通过蒙特卡洛置换检验发现, MBP、SOM和MBN是驱动土壤无机磷组分发生变化的最关键因子(P<0.05), 分别解释了土壤无机磷组分变化的74%(P=0.002, F=51.60), 5%(P=0.006, F=4.10)和4%(P=0.012, F=3.31)(表 3)。在RDA排序的双标图中, 第一主轴将对照与顶果木纯林和降香黄檀/顶果木混交林分开, 第二主轴将对照与降香黄檀纯林和顶果木纯林分开, 表明固氮树种的引入均能改变土壤无机磷组分的组成, 其中顶果木纯林和降香黄檀/顶果木混交林对无机磷组分的改变最为明显(图 4)。此外, 土壤无机磷组分与土壤基本性质存在显著的相关性, 除了pH与各土壤无机磷组分呈显著负相关外, 其余筛选出的土壤基本性质与各土壤无机磷组分均呈显著正相关(图 4)。
| 因子 Factor |
边际效应 Marginal effect |
条件效应 Conditional effect |
P | F |
| MBP | 0.741 | 0.74 | 0.002** | 51.60 |
| SOM | 0.394 | 0.05 | 0.006** | 4.10 |
| MBN | 0.242 | 0.04 | 0.012* | 3.31 |
| MBC | 0.234 | 0.01 | 0.230 | 1.41 |
| pH | 0.230 | 0.02 | 0.242 | 1.40 |
| NH4+-N | 0.185 | 0.01 | 0.296 | 1.24 |
| NO3--N | 0.231 | 0.01 | 0.562 | 0.69 |
| * P<0.05, ** P<0.01 | ||||

|
| 图 4 无机磷组分和各种土壤基本性质的冗余分析 Fig. 4 Redundancy analysis of inorganic phosphorus fractions and soil basic properties SOM: 有机质Soil organic matter;NH4+-N: 铵态氮Ammonium nitrogen;NO3--N: 硝态氮Nitrate nitrogen;MBC: 微生物量碳Soil microbial biomass carbon;MBN: 微生物量氮Soil microbial biomass nitrogen;MBP: 微生物量磷Soil microbial biomass phosphorus |
在岩溶区生态系统中, 土壤氮和磷的有效性是决定其地上生产力的关键因素, 是限制其植物生长发育的最主要因子之一[4]。Binkley等[26]发现, 在热带和亚热带地区通过改变植被类型, 土壤肥力和化学特性可能在短短数年内发生变化。李树斌等[27]也发现植被组成可能影响森林土壤的养分有效性。本研究结果表明, 石漠化山区引进固氮树种进行植被恢复8年后, 土壤的SOM、NO3--N、AP、MBC、MBN和MBP含量均显著增加, 土壤氮和磷的有效性得到有效提高(表 2)。发生以上变化的原因, 一方面可能是因为引进的固氮树种是乔木, 相比天然保留地(CK)稀疏的灌木和草本具有更多更发达的地下根系, 可以吸收土壤中更多的养分, 甚至从岩层更深处汲取营养供植物和微生物生长繁殖[28-29]。另一方面可能是固氮树种与固氮细菌结合, 能够持续为土壤固定大量的氮, 并提高土壤氮有效性, 除供植物自身生长需要外, 也为土壤微生物大量繁殖提供必需的养分, 进而提高土壤SOM、MBC、MBN和MBP含量[30]。也有研究表明植物需要从无机自然界中吸收自己所需要的养分来完成生命活动, 并以枯枝落叶等形式将各种营养元素又归还土壤, 使各种营养元素在不同深度的土壤中重新分布, 这就可能导致表层土壤的养分含量得到提高[31]。此外, 本研究中对照样地的SOM含量处于较高水平, 其主要原因可能包括: 一是从2011年造林进行植被恢复以来, 试验区就封山禁止人为的垦殖、放牧、樵采等活动, 对照样地的有机质经过8年积累得到提升;二是由于石山立地条件的具有特殊性, 地形较为复杂, 有很多坑洼、沟壑等, 也会使对照样地内形成特殊小生境, 导致其表层土壤有机质含量较高。本研究发现采用不同固氮树种或种植模式进行石漠化山区植被恢复对改善土壤养分的有效性有差异, 顶果木纯林和顶果木/降香黄檀混交林的土壤氮和磷有效性显著高于降香黄檀纯林, 这可能与不同林分类型其地上生产力、地下根系数量以及植物残体进行养分归还速度、质量和数量等不同有关[28, 32]。
土壤磷素活化系数(PAC)是衡量土壤TP有效性的重要指标[33]。PAC大于2.0%说明TP中各形态的磷容易转化为AP供植物吸收利用, 反之则不利于TP转化为AP[4]。本研究发现对照样地的PAC<2.0%, 降香黄檀纯林、顶果木纯林和降香黄檀/顶果木混交林的PAC>2.0%, 降香黄檀/顶果木混交林的PAC值最大(图 1), 说明石漠化山区引进固氮树种顶果木或降香黄檀进行植被恢复均能有效提高土壤磷的转化及其有效性, 且以顶果木和降香黄檀混交种植的效果最好。豆科植物由于具有固氮能力, 已被广泛用于改良土壤肥力和提高生态系统净初级生产力[34]。土壤中物质交换、能量流动和信息传递错综复杂, 其中氮元素与磷元素循环关系紧密[35-36]。有人认为, 固氮树种可以增强土壤氮有效性, 一方面使植物的生产力提高, 地面富含氮、磷元素的凋落物增加, 直接向土壤中释放可溶性氮、磷, 进行养分归还;另一方面影响土壤微生物群落的生物化学性质, 促进土壤磷酸酶活性进而使土壤磷有效性提高[2]。也有人认为固氮树种对土壤AP含量无显著影响, 原因是土壤氮有效性增强的同时, 土壤中磷矿化的作用也在增强, 迅速弥补了土壤中被植物吸收的AP, 所以土壤中的AP仍处于一个平衡状态[37]。而我们的研究发现NH4+-N、NO3--N和MBN与AP分别呈显著线性正相关关系, MBN与MBP呈极显著线性正相关关系(图 2), 表明引进固氮树种在增加石漠化山区土壤氮有效性的同时也增加了土壤磷的有效性。
3.2 固氮树种对石漠化山区表层土壤无机磷组分的影响土壤磷包括有机磷和无机磷两大类, 虽然土壤有机磷占土壤总磷的比例大多在20%—80%之间, 但是土壤无机磷是植物可利用磷的重要来源[38]。蒋柏潘和顾益初提出的石灰型土壤无机磷分级体系把土壤无机磷分为Ca2-P、Ca8-P、Ca10-P、Al-P、Fe-P和O-P六种, 可以准确地评价土壤AP库的大小、形态转换和磷元素的供应情况[24-25]。土壤无机磷中Ca2-P、Ca8-P和Al-P属于有效态无机磷, 很容易被植物吸收利用;Ca10-P、Fe-P和O-P属于缓效态无机磷, 较难被植物直接利用[39]。杨慧等[4]研究岩溶区不同的土地利用类型发现, 改变植被类型后土壤的各无机磷形态发生了一定的变化。我们研究也发现在石漠化山区引进固氮树种进行植被恢复后, 与对照相比各林分的土壤无机磷组分发生变化, 其中顶果木/降香黄檀混交林和顶果木纯林的各种无机磷的增加量均达到显著水平。在本研究中, 土壤各形态无机磷的含量排列顺序大致都遵循Ca10-P>O-P>Al-P>Fe-P> Ca2-P >Ca8-P。与对照相比, 引入固氮树种进行植被恢复8年后, 作为植物最有效磷源的Ca2-P大概增加了1—2倍;作为第二有效磷源的Ca8-P大概增加了0.27—0.30倍, Al-P大概增加了6—8倍;作为缓效态磷源的Ca10-P大概增加了0.38—0.49倍, Fe-P大概增加了1—1.2倍, O-P大概增加了1—5倍(图 3)。这说明了引进固氮树种后, 有利于有效态土壤无机磷的释放, 也积累了很多缓效态的土壤无机磷。蔡鑫淋等[40]研究岩溶区生态恢复中土壤无机磷形态特征表明林草间作地的无机磷含量高于荒草地, 这与我们的研究结果一致, 但土壤无机磷中O-P最多这一结果与我们的结果相反。化党领等[41]研究发现石灰型土壤中的无机磷主要以Ca-P为主与我们的研究结果一致。
影响土壤无机磷形态转化的原因有很多, 许多科学家已经对不同植被类型、不同土壤类型、不同土壤层次、不同施肥方式等对土壤无机磷组分的转化开展了大量研究[39, 41-43]。成土母质是土壤形成的基础, 是土壤中磷素含量和形态的最重要决定因素之一[40]。本研究所采集的土样均来自同一种石灰岩发育而成的土壤, 土样采集的不同在于样品所处的主要植被类型不同。齐雁冰等[44]指出土壤理化性质是影响土壤无机磷组分含量和形态变化的主要原因, 林德喜等[45]发现土壤有机质对无机磷形态变化起重要作用。微生物也可能是影响土壤无机磷形态和含量的重要因素。土壤微生物是连接地上植物群落和地下土壤养分交流的纽带, 参与各种土壤养分的循环[46], 而且对周围土壤环境的变化极为敏感, 可以及时调整MBC、MBN和MBP等的含量, 改变对土壤有机磷的矿化速率, 从而对土壤无机磷组分的形态和含量产生影响。通过对土壤无机磷组分与各种土壤基本性质进行冗余分析, 我们发现引进固氮树种进行植被恢复后各样地的无机磷形态和含量均发生了明显的变化, 土壤无机磷组分与土壤基本性质存在一定的相关性, 除了pH与各土壤无机磷组分呈显著负相关外, MBP、SOM、MBN、MBC、NH4+-N和NO3--N与各土壤无机磷组分均呈显著正相关(图 4), 而且MBP、SOM和MBN是影响石漠化山区土壤无机磷组分发生变化的关键因素(表 3)。这与Achat等[47]、Vincent等[48]、Huang和Spohn[49]的研究结果一致, 表明更高的土壤有机质含量可以增加微生物的生物量和活性, 并最终提高土壤磷的矿化率。本研究结果表明固氮树种及其种植模式对石漠化区土壤磷素转化及其有效性具有促进作用, 其中以降香黄檀和顶果木混交林的效果最为显著。而近期的一些研究发现非固氮树种及其他植被也可以在一定程度上改善石漠化区的土壤养分状况, 但不同的植物种类或恢复模式产生的作用效果存在较大差异[50-51]。
4 结论在石漠化山区引种珍贵固氮树种(降香黄檀和顶果木)进行植被恢复后, 一定程度上改善了该区表层土壤的养分状况, 使土壤无机磷组分发生了改变, 提高了土壤氮和磷的有效性, 其中以混交林的效果最为显著。土壤无机磷组分与土壤基本性质存在一定的相关性, MBP、SOM和MBN可能是影响石漠化山区土壤无机磷组分发生变化的最关键因素。本研究为珍贵固氮树种在石漠化生态恢复与重建中的应用提供理论依据和实践支持, 促进岩溶区生态、经济和社会可持续发展。
| [1] |
Wardle D A, Walker L R, Bardgett R D. Ecosystem properties and forest decline in contrasting long-term chronosequences. Science, 2004, 305(5683): 509-513. DOI:10.1126/science.1098778 |
| [2] |
冯婵莹, 郑成洋, 田地. 氮添加对森林植物磷含量的影响及其机制. 植物生态学报, 2019, 43(3): 185-196. |
| [3] |
章爱群, 贺立源, 赵会娥, 郭再华, 崔雪梅. 有机酸对土壤无机态磷转化和速效磷的影响. 生态学报, 2009, 29(8): 4061-4069. DOI:10.3321/j.issn:1000-0933.2009.08.006 |
| [4] |
杨慧, 曹建华, 孙蕾, 栾会妮, 侯彦林. 岩溶区不同土地利用类型土壤无机磷形态分布特征. 水土保持学报, 2010, 24(2): 135-140. |
| [5] |
Prescott C E, Grayston S J. Tree species influence on microbial communities in litter and soil: current knowledge and research needs. Forest Ecology and Management, 2013, 309: 19-27. DOI:10.1016/j.foreco.2013.02.034 |
| [6] |
Ushio M, Kitayama K, Balser T C. Tree species-mediated spatial patchiness of the composition of microbial community and physicochemical properties in the topsoils of a tropical montane forest. Soil Biology and Biochemistry, 2010, 42(9): 1588-1595. DOI:10.1016/j.soilbio.2010.05.035 |
| [7] |
Zhao X, Wang Q, Kakubari Y. Stand-scale spatial patterns of soil microbial biomass in natural cold-temperate beech forests along an elevation gradient. Soil Biology and Biochemistry, 2009, 41(7): 1466-1474. DOI:10.1016/j.soilbio.2009.03.028 |
| [8] |
Fanin N, Hättenschwiler S, Fromin N. Litter fingerprint on microbial biomass, activity, and community structure in the underlying soil. Plant and Soil, 2014, 379(1/2): 79-91. |
| [9] |
王宁, 罗佳琳, 赵亚慧, 李勇, 于建光. 不同麦秸还田模式对稻田土壤微生物活性和微生物群落组成的影响. 农业环境科学学报, 2020, 39(1): 125-133. |
| [10] |
孙瑞, 孙本华, 高明霞, 杨学云, 张树兰. 长期不同土地利用方式下土土壤微生物特性的变化. 植物营养与肥料学报, 2015, 21(3): 655-663. |
| [11] |
盛茂银, 刘洋, 熊康宁. 中国南方喀斯特石漠化演替过程中土壤理化性质的响应. 生态学报, 2013, 33(19): 6303-6313. |
| [12] |
Yan Y J, Dai Q H, Hu G, Jiao Q, Mei L N, Fu W B. Effects of vegetation type on the microbial characteristics of the fissure soil-plant systems in karst rocky desertification regions of SW China. Science of the Total Environment, 2020, 712: 136543. DOI:10.1016/j.scitotenv.2020.136543 |
| [13] |
朱晓锋, 陈洪松, 付智勇, 王克林, 张伟, 徐勤学, 方荣杰. 喀斯特灌丛坡地土壤-表层岩溶带产流及氮素流失特征. 应用生态学报, 2017, 28(7): 2197-2206. |
| [14] |
张伟, 王克林, 刘淑娟, 叶莹莹, 潘复静, 何寻阳. 喀斯特峰丛洼地植被演替过程中土壤养分的积累及影响因素. 应用生态学报, 2013, 24(7): 1801-1808. |
| [15] |
Zhang W, Zhao J, Pan F J, Li D J, Chen H S, Wang K L. Changes in nitrogen and phosphorus limitation during secondary succession in a karst region in Southwest China. Plant and Soil, 2015, 391(1): 77-91. |
| [16] |
Tian H Q, Chen G S, Zhang C, Melillo J M, Hall C A S. Pattern and variation of C: N: P ratios in China's soils: a synthesis of observational data. Biogeochemistry, 2010, 98(1/3): 139-151. |
| [17] |
Zhao F Z, Ren C J, Han X H, Yang G H, Wang J, Doughty R. Changes of soil microbial and enzyme activities are linked to soil C, N and P stoichiometry in afforested ecosystems. Forest Ecology and Management, 2018, 427: 289-295. DOI:10.1016/j.foreco.2018.06.011 |
| [18] |
Liu C C, Liu Y G, Guo K, Qiao X G, Zhao H W, Wang S J, Zhang L, Cai X L. Effects of nitrogen, phosphorus and potassium addition on the productivity of a karst grassland: Plant functional group and community perspectives. Ecological Engineering, 2018, 117: 84-95. DOI:10.1016/j.ecoleng.2018.04.008 |
| [19] |
罗攀, 陈浩, 肖孔操, 杨利琼, 文丽, 李德军. 地形、树种和土壤属性对喀斯特山区土壤胞外酶活性的影响. 环境科学, 2017, 38(6): 2577-2585. |
| [20] |
鲍士旦. 土壤农化分析(第三版). 北京: 中国农业出版社, 2000: 1-120.
|
| [21] |
Murphy J, Riley J P. A modified single solution method for the determination of phosphate in natural waters. Analytica Chimica Acta, 1962, 27: 31-36. DOI:10.1016/S0003-2670(00)88444-5 |
| [22] |
Jenkinson D S, Powlson D S. The effects of biocidal treatments on metabolism in soil-V: a method for measuring soil biomass. Soil Biology and Biochemistry, 1976, 8(3): 209-213. DOI:10.1016/0038-0717(76)90005-5 |
| [23] |
Brookes P C, Powlson D S, Jenkinson D S. Measurement of microbial biomass phosphorus in soil. Soil Biology and Biochemistry, 1982, 14(4): 319-329. DOI:10.1016/0038-0717(82)90001-3 |
| [24] |
蒋柏藩, 顾益初. 石灰性土壤无机磷分级体系的研究. 中国农业科学, 1989, 22(3): 58-66. DOI:10.3321/j.issn:0578-1752.1989.03.012 |
| [25] |
顾益初, 蒋柏藩. 石灰性土壤无机磷分级的测定方法. 土壤, 1990, 22(2): 101-102, 110-110. |
| [26] |
Binkley D, Giardina C, Bashkin M A. Soil phosphorus pools and supply under the influence of Eucalyptus saligna and nitrogen-fixing Albizia facaltaria. Forest Ecology and Management, 2000, 128(3): 241-247. DOI:10.1016/S0378-1127(99)00138-3 |
| [27] |
李树斌, 周丽丽, 陈宝英, 杨起帆, 丁国昌, 林思祖. 亚热带树种转换对林地土壤碳氮磷计量比的影响. 森林与环境学报, 2019, 39(6): 575-583. |
| [28] |
Zhang C, Liu GB, Xue S, Song Z L. Rhizosphere soil microbial activity under different vegetation types on the Loess Plateau, China. Geoderma, 2011, 161(3/4): 115-125. |
| [29] |
Feng J, Wu J J, Zhang Q, Zhang D D, Li Q X, Long C Y, Yang F, Chen Q, Cheng X L. Stimulation of nitrogen-hydrolyzing enzymes in soil aggregates mitigates nitrogen constraint for carbon sequestration following afforestation in subtropical China. Soil Biology and Biochemistry, 2018, 123: 136-144. DOI:10.1016/j.soilbio.2018.05.013 |
| [30] |
丁国昌, 万晓华, 杨起帆, 林群星, 黄志群. 亚热带树种转换对林地土壤微生物群落结构和功能的影响. 应用生态学报, 2017, 28(11): 3751-3758. |
| [31] |
Jobbágy E G, Jackson R B. The uplift of soil nutrients by plants: biogeochemical consequences across scales. Ecology, 2004, 85(9): 2380-2389. DOI:10.1890/03-0245 |
| [32] |
Horodecki P, Jagodziński A M. Tree species effects on litter decomposition in pure stands on afforested post-mining sites. Forest Ecology and Management, 2017, 406: 1-11. DOI:10.1016/j.foreco.2017.09.059 |
| [33] |
张英鹏, 陈清, 李彦, 李国生, 孙明, 刘兆辉. 不同磷水平对山东褐土耕层无机磷形态及磷有效性的影响. 中国农学通报, 2008, 24(7): 245-248. |
| [34] |
Zhao J, Wang X L, Wang X L, Fu S L. Legume-soil interactions: legume addition enhances the complexity of the soil food web. Plant and Soil, 2014, 385(1-2): 273-286. DOI:10.1007/s11104-014-2234-2 |
| [35] |
Vitousek P M, Porder S, Houlton B Z, Chadwick O A. Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions. Ecological Applications, 2010, 20(1): 5-15. DOI:10.1890/08-0127.1 |
| [36] |
Nasto M K, Alvarez-Clare S, Lekberg Y, Sullivan B W, Townsend A R, Cleveland C C. Interactions among nitrogen fixation and soil phosphorus acquisition strategies in lowland tropical rain forests. Ecology Letters, 2014, 17(10): 1282-1289. DOI:10.1111/ele.12335 |
| [37] |
Yang K, Zhu J J, Gu J C, Yu L Z, Wang Z Q. Changes in soil phosphorus fractions after 9 years of continuous nitrogen addition in a Larix gmelinii plantation. Annals of Forest Science, 2015, 72(4): 435-442. DOI:10.1007/s13595-014-0444-7 |
| [38] |
李新乐, 侯向阳, 穆怀彬, 李西良, 郭丰辉. 连续6年施磷肥对土壤磷素积累、形态转化及有效性的影响. 草业学报, 2015, 24(8): 218-224. |
| [39] |
曲均峰, 戴建军, 徐明岗, 李菊梅. 长期施肥对土壤磷素影响研究进展. 热带农业科学, 2009, 29(3): 75-80. DOI:10.3969/j.issn.1009-2196.2009.03.017 |
| [40] |
蔡鑫淋, 舒英格, 陈梦军. 喀斯特山区生态恢复中土壤剖面无机磷形态分布特征及其影响因素. 水土保持通报, 2020, 40(2): 107-114. |
| [41] |
化党领, 余长坤, 刘世亮, 刘芳, 介晓磊. 石灰性土壤不同土层磷形态研究. 中国农学通报, 2008, 24(9): 277-282. |
| [42] |
包先明, 崔宏. 退耕还湖后不同植被群落湿地土壤剖面磷素形态分布特征. 土壤, 2019, 51(1): 25-31. |
| [43] |
王艳玲, 王杰, 赵兰坡, 蔡洪光. 黑土无机磷形态及其有效性研究. 水土保持学报, 2004, 18(3): 85-89. DOI:10.3321/j.issn:1009-2242.2004.03.022 |
| [44] |
齐雁冰, 常庆瑞, 田康, 刘梦云, 刘京, 陈涛. 黄土丘陵沟壑区不同植被恢复模式土壤无机磷形态分布特征. 农业环境科学学报, 2013, 32(1): 56-62. |
| [45] |
林德喜, 范晓晖, 胡锋, 杨林章, 韩晓增. 长期施肥后简育湿润均腐土中磷素形态特征的研究. 土壤学报, 2006, 43(4): 605-610. DOI:10.3321/j.issn:0564-3929.2006.04.011 |
| [46] |
Huang X M, Liu S R, Wang H, Hu Z D, Li Z G, You Y M. Changes of soil microbial biomass carbon and community composition through mixing nitrogen-fixing species with Eucalyptus urophylla in subtropical China. Soil Biology and Biochemistry, 2014, 73: 42-48. DOI:10.1016/j.soilbio.2014.01.021 |
| [47] |
Achat D L, Bakker M R, Zeller B, Pellerin S, Bienaimé S, Morel C. Long-term organic phosphorus mineralization in Spodosols under forests and its relation to carbon and nitrogen mineralization. Soil Biology and Biochemistry, 2010, 42(9): 1479-1490. DOI:10.1016/j.soilbio.2010.05.020 |
| [48] |
Vincent A G, Turner B L, Tanner E V J. Soil organic phosphorus dynamics following perturbation of litter cycling in a tropical moist forest. European Journal of Soil Science, 2010, 61(1): 48-57. DOI:10.1111/j.1365-2389.2009.01200.x |
| [49] |
Huang W J, Spohn M. Effects of long-term litter manipulation on soil carbon, nitrogen, and phosphorus in a temperate deciduous forest. Soil Biology and Biochemistry, 2015, 83: 12-18. DOI:10.1016/j.soilbio.2015.01.011 |
| [50] |
陈志霞, 喻阳华, 吴银菇. 黔中石漠化区大理石矿山不同植被恢复对土壤养分及化学计量的影响. 贵州师范大学学报: 自然科学版, 2021, 39(1): 95-102. |
| [51] |
郑武扬, 王艳霞, 王月江, 冯刚刚. 喀斯特石漠化区不同植被治理模式对土壤质量的影响. 西部林业科学, 2020, 49(4): 41-47. |