 2022, Vol. 42
2022, Vol. 42文章信息
- 杨琪, 李书恒, 李家豪, 杜建峰, 王嘉川
- YANG Qi, LI Shuheng, LI Jiahao, DU Jianfeng, WANG Jiachuan
- 西安4种落叶乔木物候期对气候变化的响应
- Response of phenology phases of four deciduous trees to climate change in Xi'an
- 生态学报. 2022, 42(4): 1462-1473
- Acta Ecologica Sinica. 2022, 42(4): 1462-1473
- http://dx.doi.org/10.5846/stxb202011212992
-
文章历史
- 收稿日期: 2020-01-12
- 网络出版日期: 2021-11-04





2. 陕西省地表系统与环境承载力重点实验室, 西安 710127
2. Shaanxi Key Laboratory of Earth Surface System and Environmental Carrying Capacity, Xi'an 710127, China
物候学(Phenology)是研究自然界的植物(包括农作物)、动物和环境条件(气候、水文和土壤条件)的周期变化及其相互关系的科学[1-2]。植物物候现象不仅反映季节变化, 也是生态系统对全球气候变化的响应[3], 因此, 研究植物物候对理解植被-气候关系, 提高植被生产力、陆地生态系统碳储备及碳循环过程具有重要的意义[4-5]。
国外的物候研究起步较早, 在物候与气候变化的响应研究方面已经取得了显著的成果, 观测资料表明, 欧洲、北美、地中海[6-8]等地的木本植物物候随着温度的变化呈现春季物候提前, 秋季物候期延后, 物候生长季延长的现象。在我国, 严应存等[9]研究发现青藏高原气候变暖是影响青稞物候期变化的主导因素;Chen等[10]研究发现, 在温带地区温度每升高1℃可能会导致生长季节提前2.8d, 结束日期延迟2.1d。这与我国其他地区[11-17]的物候变化趋势相一致, 普遍呈现春季物候提前, 秋季延迟的现象。也有学者发现, 在某些地区, 植物物候对降水的敏感度更高, 尤其是在干旱情况[18-20], Xin等[21]研究发现降水对干旱半干旱区域植被返青期的影响大于气温;陈效逑等[22]发现我国温带旱柳叶变色始期和落叶末期的发生日期受到前期气温和降水量的综合影响;肖芳等[23]研究内蒙古草原物候发现前2个月降水量对大针茅开花期的影响较大, 降水量的增加也会使得大针茅黄枯期推迟。除此之外, 日照对植物物候变化具有同样重要的作用[24-25]。Keller等[26]研究奥地利的33种高山植物花期发现, 54%的植物开花期对光周期比较敏感;郑景云等[27]研究发现日照是影响晚夏植物花粉传播的关键气候因子。
关于西安地区植物物候变化的影响因素分析, 目前主要探讨了温度、降水等气候因素对植物物候期变化的影响, 较多关注于物候期与单一气象因子的响应程度分析, 如白洁等[28]分析了1963-2007年西安木本植物物候期与气温和降水的关系。此外现阶段的研究多以春季物候为主, 王焕炯等[29]利用积温模型探究了气候波动对西安39种木本植物展叶始期的影响。陶泽兴等[30]比较了同一气候区内西安和宝鸡两地植物始花期对温度变化的敏感性。而秋季物候作为影响植被生长季长度的另一重要节点, 在西安物候研究中较少提及[28, 31], 且研究时间较早。因此, 本文利用分段回归和趋势倾向率等方法, 从物候突变的角度分析毛白杨(Populus tomentosa)、杜梨(Pyrus betulifolia)、七叶树(Aesculus chinensis)和灯台树(Bothrocaryum controversum)4种落叶乔木展叶盛期和叶全变色期的生长趋势, 探讨了气温、降水和日照时数与物候期的响应关系, 并通过偏最小二乘回归(PLS)判断气候变量对物候期的综合影响。由于植被对气候变化的响应具有滞后效应[32], 气候因子对物候的影响也会随着时间的变化而变化[33], 因此简单的进行物候期与气候因子的相关性分析, 可能会掩盖一些现象。PLS兼具主成分分析和多元回归分析的优点, 考虑了自变量之间的相关性, 克服了预测变量相关导致的多元共线性问题[34], 能够识别气候因子影响物候的主要时段, 为深刻认识西安地区植物物候期与气候因子的相互作用机制提供科学基础。
1 研究区概况西安地处关中平原中部, 北濒渭河, 南临秦岭, 位于107.40°E-109.49°E和33.42°N-34.45°N之间, 属于暖温带半湿润大陆性季风气候, 四季分明, 雨热同期, 多年(1951-2018年)平均气温13.9℃, 年均降水量567mm。冬季低温少雨, 夏季高温多雨, 降水多集中在7-9月(约占全年降水约50%)。研究发现, 从1951年至今, 西安气温呈现出显著的增加趋势[35], 并且随着人口扩张和城市建筑面积的增加, 西安的城市热岛效应也在不断增加[36]。而城市热岛效应通过温度变化直接影响植物物候, 是城市植物物候变化最主要的影响因子[37]。因此, 西安是研究气候变化对物候响应的理想区域。
本文物候观测地点位于西安市老植物园内。该园创建于1959年, 是解放后全国重点建设的八大植物园之一, 占地面积20hm2, 收集保存植物3400余种, 积累了丰富的物候观测资料。
2 数据和方法 2.1 物候数据本文所使用的物候数据来源于陕西省西安市植物园, 选取了观测时间长、连续性好、物候现象强的毛白杨、杜梨、七叶树和灯台树四种乔木为研究对象。
为了研究物候变化特征及其与气候变化的响应关系, 选择了展叶盛期、叶全变色期分别作为春季和秋季的物候指示期。由于物候观测存在缺测年份, 对于记录中断的1987、1989和1993年采取剔除原则, 不做处理, 对于缺少个别的物候期的植物, 根据前后记录数据进行插值处理来保证物候数据的完整性。同时物候日期采用Julian日换算方法[1]进行换算, 即自元月1日至该日期的日数为Julian日, 从而得到物候期的时间序列。
2.2 气象数据本研究气象数据采用西安气象站的气象数据, 主要包括1951-2018年的日平均气温、降水量、日照时数数据, 其数据来源于中国气象数据科学共享服务网(www.geodata.cn)和全国温室数据系统(http://data.sheshiyuanyi.com/WeatherData/)。
2.3 研究方法 2.3.1 分段回归分段回归被广泛应用于植物物候转折[38-39], 已有研究表明, 植物物候不仅在物种间、地区间存在差异, 在年际间也略有差异[13, 40]。因此, 本文采用分段回归的方法分析西安地区植物物候转折。

|
(1) |
式中, x表示时间序列的年份, y是响应变量(年均植物物候期参数), β0、β1、β2是回归系数;β0为截距, β1与β1+β2分别为转折前后物候变化趋势;ε表示误差项;当检验显著性水平达到0.1时, 转折点达到了变异程度, α称为变异点。同时, 采用F检验检验分段线性回归的显著性。
2.3.2 偏最小二乘回归法偏最小二乘回归法(Partial least regression, PLS)最先产生于化学领域, 近年来被应用于植被变化归因[41]和植物物候变化研究[42-43]。偏最小二乘回归分析主要输出变量重要性值(Variable importance Index, VIP)和标准化的相关系数两个结果, 一般认为, VIP值越大其解释意义越显著, VIP>1说明自变量对因变量具有显著的解释意义;0.8 < VIP < 1具有中等程度解释意义;VIP < 0.8基本不具备解释意义[44]。计算公式如下[45]:
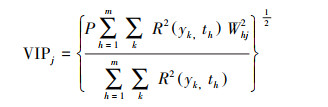
|
(2) |
式中, p为自变量个数;m为提取的成分个数;k为因变量;th为自变量的第h个成分;R2(yk, th)为yk与th间相关系数的平方;wkj2为自变量xj对构造th成分的贡献权重。文中将展叶盛期和叶全变色期作为响应变量, 四季的平均气温、降水和日照时数作为独立变量, 采用PLS法研究西安气候因子对物候期的综合影响。
3 结果与分析 3.1 1979-2018年西安气候因子变化特征从图 1可以看出, 1979-2018年西安地区的年均温以0.6℃/10a的速率呈显著的波动上升状态, 约上升了2.34℃, 温度起伏范围在12.7-15.8℃之间, 其中1984年均温最低, 为12.7℃, 2016年达到峰值, 为15.8℃;降水与气温变化刚好相反, 以11.88mm/10a的速率呈微弱波动下降趋势, 约减少了73.32mm, 降水范围在408.8-903.2mm之间;年累计日照时数以116.82h/10a的速率呈显著增加趋势。总体来看, 西安地区气候呈现暖干化的变化趋势, 其中春季的变化最为明显。

|
| 图 1 1979-2018年西安均温、降水和日照时数变化 Fig. 1 The change of annual temperature, precipitation and sunshine duration in Xi′an during 1979-2018 |
从图 2可以看出, 4种植物的平均物候期变化趋势一致, 春季物候期提前2.9d/10a(P < 0.01), 秋季物候平均推迟4.5d/10a(P < 0.01)。即物候生长期延长, 叶全变色期物候比展叶盛期物候对气候变化的响应更为敏感, 秋季叶全变色期变化速率和推迟天数对生长季延长的贡献大于春季展叶盛期, 这与赖小红等[46]发现重庆主城区的植物生长季延长主要依赖于秋季物候的推迟相一致。可能是因为城市热岛效应在秋冬季节更为强烈, 冬季气温偏高导致叶全变色期物候变化幅度增大, 延迟植物进入休眠期, 从而使得秋季物候期推迟。

|
| 图 2 1979-2018年物候期变化 Fig. 2 The change in the phenology period during 1979-2018 |
4种乔木展叶盛期发生在第73-115天, 叶全变色期发生在第280-336天。展叶盛期, 4种乔木均呈极显著提前趋势, 七叶树的变化速率最大, 为3.2d/10a(P < 0.01)。叶全变色期, 杜梨物候变化最为明显, 推迟5.7d/10a(P < 0.01), 灯台树的物候变化最小, 推迟2.9d/10a(P < 0.01)。从统计结果可看出, 4种乔木中灯台树对气候变化的敏感性反应最弱, 生长期延长5.6d/10a。
3.2.2 物候分段西安木本植物展叶盛期和叶全变色期于1982年发生转折(图 3), 已有研究表明全国年均气温转折时间为1984年[47], 秦岭地区的物候始期突变发生在1985年[14]。资料显示, 1981-2010年西安站气温上升速率加快, 同时随着城市发展进程加快, 1981-2010年也是西安城市热岛的一个明显加强期[35], 热岛效应通过扰乱气流的动态过程使得形成区别于森林生态系统的城市生态系统[48], 城市物候研究存在“热岛误差”效应[49], 这可能是物候期转折点略早于全国均温转折和秦岭地区物候转折的原因。

|
| 图 3 1979-2018年物候期变化与曲线斜率 Fig. 3 The change in the phenology period and curve slope during 1979-2018 |
由于转折前期物候序列较短, 本文仅分析转折后物候变化趋势。可以看出(图 4), 4个树种展叶盛期显著提前, 平均提前3.8d/10a(P < 0.01), 叶全变色期平均推迟5.0d/10a(< 0.01), 与1979-2018年整个物候变化序列相比, 物候期变化速率和显著性增加, 植被生长季长度明显延长。其中, 毛白杨与杜梨展叶期相差天数由1981年的20d缩短为2018年的14d, 叶全变色期由1981年的12d缩短为8d, 表明物候变化与气候的阶段性特征是一致的。

|
| 图 4 1979-2018年物候期变化 Fig. 4 The change in the phenology period during 1979-2018 |
研究区木本植物物候期与气候因子的相关分析结果如图 5所示。由图 5可以看出, 展叶盛期与春季气温表现为极显著负相关, 即气温越高, 植物生长发育所需的积温达到时间越早, 从而春季物候期起始时间越早。4种植物展叶盛期与气温的相关性表现不尽相同。毛白杨、七叶树展叶盛期与全年的气温关系都极为密切, 呈极显著负相关;杜梨主要受春季和夏季的气温控制, 相关系数分别为-0.529和-0.424(P < 0.01);灯台树展叶盛期与春季、夏季、冬季的气温极显著相关, 相关系数在-0.672--0.458(P < 0.01)之间。展叶盛期与气温的相关性分析发现, 不同树种物候发生期对气温的需求不同, 春季物候期不仅受春季气温的影响, 同时前期气温也将对物候发生产生一定的作用。

|
| 图 5 物候与气候因子的关系 Fig. 5 The relationship between phenology and climate factors |
叶全变色期与秋季气温表现为显著正相关, 即温度升高会使得叶全变色期物候指标推迟, 这可能与秋季降水减少, 升温使得植被光合作用增强, 从而减少叶绿素退化, 延迟进入生长季末期有关[50]。其中, 毛白杨、杜梨叶全变色期与春季和秋季的气温极显著正相关;灯台树叶全变色期与夏季、秋季气温正相关性最好, 分别为0.447和0.559(P < 0.01)。
相关分析结果表明毛白杨和杜梨物候变化不受降水控制, 七叶树和灯台树与降水的相关性各不相同, 其中, 七叶树展叶盛期与冬季降水呈显著负相关关系, 可能是因为冬季降水较多, 不利于植物休眠, 降水对七叶树展叶盛期的影响存在滞后效应;灯台树叶变色期与夏季的降水呈显著负相关相关, 相关系数为-0.342(P < 0.05)。
图 5可以看出, 展叶盛期与春季的日照时数呈现负相关关系, 即春季降水减少, 日照时数增多, 植物光合作用增强, 刺激植被生长发芽, 使得物候期提前;杜梨展叶盛期与春季、冬季日照时数显著相关, 相关系数分别为-0.446、-0.426(P < 0.01);七叶树与灯台树展叶期受春季、秋季日照的控制。
秋季物候期与日照时数主要呈正相关关系, 即日照时数减少, 植物蒸散发变缓, 利于植物体内有机质的累积和能量输送, 使得植物生长季延长, 秋季物候期推迟。毛白杨叶全变色期与春季日照呈极显著正相关关系, 相关系数为0.438(P < 0.01);七叶树叶全变色期与冬季日照极显著相关, 相关系数为0.507(P < 0.01);灯台树叶全变色期则主要受夏季日照影响, 杜梨与日照时数的相关性不显著。
3.3.2 气候因子对物候期的综合影响效应由偏最小二乘回归所得的图 6可知, 4种植被春季物候与气温和日照呈负相关, 说明春季气温上升, 日照时数增加, 蒸发增强, 形成水分胁迫, 光合作用减弱, 使得植物展叶盛期出现提前趋势;降水对于春季物候的影响不强, 春季物候主要受前一年冬季的降水影响, 与之表现出负相关关系, 这说明冬季降水的增加可能会使来年春季物候期提前。

|
| 图 6 物候期与气候因子的偏最小二乘回归系数 Fig. 6 Partial Least Squares Regression Coefficient of Phenological Period and Climate Factors |
不同于展叶盛期物候期, 气温与秋季物候基本呈现正相关关系, 植物秋季物候主要受当季的气温影响, 均为显著正相关;杜梨秋季物候还受夏季温度影响, 说明气温对物候期的影响存在滞后效应;降水对于叶全变色期的影响规律性不强, 模型系数正负变化较大, 其中灯台树与夏秋两季的降水呈负相关关系, 说明物候发生期前降水的减少会使得秋季物候期推迟, 这可能是因为西安地区四季分明, 雨热同期, 降水增加使得土壤含水量增加, 通过影响植被羟化作用来增强植被的光合作用, 促使树木加速生长, 从而缩短生长季, 反之使得植被秋季物候期推迟[51]。
4种乔木展叶盛期的标准化偏最小二乘回归模型:

|
4种乔木叶全变色期的标准化偏最小二乘回归模型:
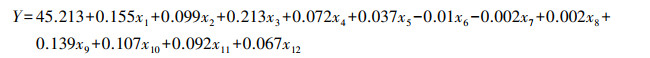
|
式中, x1-x12依次表示春季均温、夏季均温、秋季均温、冬季均温、春季日照时数、夏季日照时数、秋季日照时数、冬季日照时数、春季降水、夏季降水、秋季降水、冬季降水。
图 7和表 1为展叶盛期PLS结果。VIP直方图显示, 春季均温在植物展叶盛期起主导控制作用, 各因子的VIP值大小排序为春季气温>夏季气温>冬季气温>春季日照时数>秋季气温>秋季日照时数>冬季日照时数>夏季日照时数>冬季降水>秋季降水>春季降水>夏季降水, 即影响因子对物候始期的解释能力依次是: 春季气温>夏季气温>冬季气温>春季日照时数>秋季气温>秋季日照时数>冬季日照时数>夏季日照时数>冬季降水>秋季降水>春季降水>夏季降水。

|
| 图 7 气候因子对展叶盛期综合影响的PLS分析结果 Fig. 7 Comprehensive impact of the climate factors in the leaf expansion peak period according to PLS analysis |
| 展叶盛期VIP Blooming period VIP |
毛白杨 Populus tomentosa |
杜梨 Pyrus betulifolia |
七叶树 Aesculus chinensis |
灯台树 Bothrocaryum controversum |
| 春季T | 1.7485* | 1.50984* | 1.7686* | 1.75932* |
| 夏季T | 1.23267 | 1.21691 | 1.43397 | 1.20896 |
| 秋季T | 1.11267 | 0.870659 | 1.29133 | 1.07964 |
| 冬季T | 1.28686 | 1.12485 | 1.05824 | 1.20253 |
| 春季P | 0.458702 | 0.171076 | 0.309475 | 0.404419 |
| 夏季P | 0.34083 | 0.659197 | 0.47302 | 0.070344 |
| 秋季P | 0.657485 | 0.600935 | 0.632898 | 0.710782 |
| 冬季P | 0.591038 | 0.628694 | 0.758816 | 0.74 |
| 春季R | 1.2071 | 1.27129 | 0.984233 | 1.11531 |
| 夏季R | 0.663704 | 0.925119 | 0.791346 | 0.630831 |
| 秋季R | 0.946247 | 1.02398 | 1.04048 | 1.20258 |
| 冬季R | 0.792512 | 1.21467 | 0.234461 | 0.72259 |
| *表示最大值;T: 气温Temperature;P: 降水Precipitation;R: 日照时数Sunshine hours; PLS:偏最小二乘回归Partial Least Squares; VIP: 变量重要性值Variable importance index | ||||
图 8和表 2表明秋季均温对4种乔木叶全变色期物候变化的影响最大, 各因子的VIP值大小排序为秋季气温>春季气温>春季日照>夏季日照>夏季气温>秋季日照>冬季气温>冬季日照>春季降水>夏季降水>冬季降水>秋季降水, 即影响因子对物候末期的解释能力依次是秋季气温>春季气温>春季日照>夏季日照>夏季气温>秋季日照>冬季气温>冬季日照>春季降水>夏季降水>冬季降水>秋季降水, 但气候因子对不同树种叶全变色期的解释能力有所差别, 其中冬季和春季的日照时数对七叶树秋季物候的解释能力大于秋季气温, 这可能与七叶树喜温暖湿润及光照充足的生态环境有关。

|
| 图 8 气候因子对叶全变色期综合影响的PLS分析结果 Fig. 8 Comprehensive impact of the climate factors in the total leaf discoloration period according to PLS analysis |
| 叶全变色期VIP Whole leaf discoloration period VIP |
毛白杨 Populus tomentosa |
杜梨 Pyrus betulifolia |
七叶树 Aesculus chinensis |
灯台树 Bothrocaryum controversum |
| 春季T | 1.48756 | 1.94863 | 1.17652 | 0.690199 |
| 夏季T | 0.941242 | 0.719723 | 0.334189 | 1.4099 |
| 秋季T | 1.92495* | 2.26581* | 1.35158 | 1.73186* |
| 冬季T | 1.07613 | 0.323353 | 0.093486 | 1.01651 |
| 春季P | 0.111334 | 0.276637 | 0.078193 | 0.636519 |
| 夏季P | 0.06561 | 0.785829 | 0.216803 | 0.987303 |
| 秋季P | 0.235434 | 0.528095 | 0.33834 | 0.832341 |
| 冬季P | 0.401166 | 0.022319 | 0.144191 | 0.057774 |
| 春季R | 1.45852 | 1.20952 | 1.59619 | 0.360625 |
| 夏季R | 0.702753 | 0.087078 | 1.11841 | 1.62626 |
| 秋季R | 0.933316 | 0.027742 | 1.32693 | 0.782304 |
| 冬季R | 0.558962 | 0.040712 | 1.70904* | 0.200867 |
| *表示最大值 | ||||
总体而言, 影响因子对物候期的解释能力排序为: 气温>日照时数>降水, 即气温对植物物候指标的影响大于降水和日照时数, 春季和秋季气温的升高对促使乔木展叶盛期提前, 叶全变色期推迟的主导控制作用最为显著, 当季气温是影响物候发生期变化最重要的因素。
4 结论与讨论 4.1 结论(1) 在过去的39年, 西安地区气候呈暖干化趋势, 年平均气温和日照时数呈显著波动上升趋势, 年降水量变化趋势与之相反, 呈微弱下降趋势。
(2) 近39年, 4种落叶乔木均表现为春季物候期提前, 提前速率为2.9d/10a, 秋季物候期推迟, 推迟速率为4.5d/10a, 生长季长度延长趋势。叶全变色期物候比展叶盛期物候对气候变化的响应更为敏感, 秋季叶变色期变化速率和推迟天数对生长季延长的贡献大于春季展叶盛期。
(3) 西安木本植物展叶盛期和叶全变色期于1982年发生转折, 转折后, 物候特征发生了显著变化, 与整体趋势相比, 展叶盛期提前速率加快, 提前速率为3.8d/10a, 叶全变色期延后, 延后速率为5.0d/10a;在树种差异方面, 展叶盛期毛白杨提前速率最大, 为4.5d/10a, 叶全变色期七叶树的推迟速率最大, 为6.1d/10a。4种树木中, 灯台树对气候变化的敏感性最弱, 由此可见, 选取代用指标评价区域气候环境变化时应考虑不同树种的生理特征和种间差异。
(4) 4种乔木春季物候期与气温、日照时数呈负相关关系, 气温越高、日照时数越长, 春季物候前提前;乔木秋季物候期与气温、日照时数呈正相关关系, 气温升高, 日照时数增多, 秋季物候期延后;降水对4种乔木的影响作用均不显著。
(5) 4种落叶乔木展叶盛期与叶全变色期均受同期气温、日照时数、降水的综合影响。影响因子对物候期的解释能力排序为: 气温>日照时数>降水。气候因子对春季物候和秋季物候的影响解释量存在差异, 物候发生期当季气温是影响植物物候变化的主导因素。
4.2 讨论本文研究发现, 随着气候变暖的加速, 西安地区落叶乔木物候也随之变化, 春季物候期提前, 秋季物候期推后, 生长期延长, 与学者们[15, 23, 52]关于北半球中高纬度地区物候变化研究结果一致。
本文相关性结果表明, 气温是制约西安落叶乔木物候期变化的关键气候因子, 温度升高促进植物体内酶的活性, 延长植物发育过程[53]。展叶盛期与春季气温呈极显著负相关, 叶全变色期与秋季气温呈显著正相关, 春季气温回升越快, 展叶盛期发生越早, 秋季气温高使得叶片褐变推迟, 这与其它木本植物的研究结果相一致[54-56]。研究发现, 4种落叶乔木展叶盛期与上年冬季和当年春季的气温呈显著负相关, 说明气候因子对植物物候的影响具有滞后作用, 植物物候受前期和当季气候因子的共同作用。毛白杨、杜梨、灯台树3种乔木叶全变色期主要受秋季气温的控制, 七叶树叶全变色期对日照更为敏感, 植物体内具有光敏色素, 缩短光周期会诱导温带植物进入休眠状态[57], 日照时数越短, 植物体内水分充足, 树叶变色越晚, 这与河西走廊东部旱柳[58], 科尔沁沙地小叶杨和榆树研究结果相似[59];已有研究表明, 物候期与不同时期的降水表现出不同的相关性, 未表现出明显的规律性[60-61], 这与本文研究的降水对不同树种的物候变化的影响相一致。表明物候变化不仅受气温的影响, 同时也受降水和日照的影响, 是多种气候因素共同作用的结果, 且各个物候期对气候因子的响应也存在差异。
植物物候变化是多因素综合影响的结果, 本文仅着重分析了气候因子中的气温、降水和日照时数的影响, 在未来的研究中还应关注湿度、土壤温度、蒸散发、CO2等其他气候因子以及人为因素、地形要素、土壤类型、水文环境和植物自身的遗传因素、生理特征、种间差异等。
| [1] |
竺可桢, 宛敏渭. 物候学. 北京: 科学出版社, 1973.
|
| [2] |
Franklin J F. Phenology and seasonality modeling. Forest Science, 1975, 21(2): 211-212. |
| [3] |
翟佳, 袁凤辉, 吴家兵. 植物物候变化研究进展. 生态学杂志, 2015, 34(11): 3237-3243. |
| [4] |
Richardson A D, Keenan T F, Migliavacca M, Ryu Y, Sonnentag O, Toomey M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agricultural and Forest Meteorology, 2013, 169: 156-173. DOI:10.1016/j.agrformet.2012.09.012 |
| [5] |
Donnelly A, Jones M B, Sweeney J. A review of indicators of climate change for use in Ireland. International Journal of Biometeorology, 2004, 49(1): 1-12. |
| [6] |
Cui T F, Martz L, Lamb E G, Zhao L, Guo X L. Comparison of grassland phenology derived from MODIS satellite and PhenoCam near-surface remote sensing in North America. Canadian Journal of Remote Sensing, 2019, 45(5): 707-722. DOI:10.1080/07038992.2019.1674643 |
| [7] |
Chmielewski F M, Rötzer T. Response of tree phenology to climate change across Europe. Agricultural and Forest Meteorology, 2001, 108(2): 101-112. DOI:10.1016/S0168-1923(01)00233-7 |
| [8] |
Peñuelas J, Filella I. Herbaria century record of increasing eutrophication in Spanish terrestrial ecosystems. Global Change Biology, 2001, 7(4): 427-433. DOI:10.1046/j.1365-2486.2001.00421.x |
| [9] |
严应存, 赵全宁, 王喆, 豆青芳. 青海省门源县1980-2015年青稞物候期变化趋势及其驱动因素. 生态学报, 2018, 38(4): 1264-1271. |
| [10] |
Chen X Q, Xu L. Phenological responses of Ulmus pumila (Siberian Elm) to climate change in the temperate zone of China. International Journal of Biometeorology, 2012, 56(4): 695-706. DOI:10.1007/s00484-011-0471-0 |
| [11] |
葛全胜, 郑景云, 张学霞, 郝志新. 过去40年中国气候与物候的变化研究. 自然科学进展, 2003, 13(10): 1048-1053. DOI:10.3321/j.issn:1002-008X.2003.10.007 |
| [12] |
杨光, 宋戈, 韦振锋, 刘晗. 基于时序指数西北植被物候时空变化特征. 水土保持研究, 2015, 22(6): 213-218. |
| [13] |
马晓芳, 陈思宇, 邓婕, 冯琦胜, 黄晓东. 青藏高原植被物候监测及其对气候变化的响应. 草业学报, 2016, 25(1): 13-21. |
| [14] |
邓晨晖, 白红英, 翟丹平, 高山, 黄晓月, 孟清, 贺映娜. 气候变化背景下1964-2015年秦岭植物物候变化. 生态学报, 2017, 37(23): 7882-7893. |
| [15] |
李净, 刘红兵, 李彩云, 李龙. 基于GIMMS 3g NDVI的近30年中国北部植被生长季始期变化研究. 地理科学, 2017, 37(4): 620-629. |
| [16] |
李丹, 吴秀芹, 张靖宙, 于洋. 西南喀斯特断陷盆地植被物候动态变化及其与气候因子的响应. 水土保持研究, 2020, 27(6): 168-173. |
| [17] |
何宝忠, 丁建丽, 李焕, 刘博华, 陈文倩. 新疆植被物候时空变化特征. 生态学报, 2018, 38(6): 2139-2155. |
| [18] |
Gong Y, Staudhammer C L, Wiesner S, Starr G, Zhang Y L. Characterizing growing season length of subtropical coniferous forests with a phenological model. Forests, 2021, 12(1): 95. DOI:10.3390/f12010095 |
| [19] |
Agossou C, Kang S. Climatic factors controlling interannual variability of the onset of vegetation phenology in the northern Sub-Saharan Africa from 1988 to 2013. African Journal of Ecology, 2020, 58(2): 299-308. DOI:10.1111/aje.12699 |
| [20] |
Fu Y H, Zhou X C, Li X X, Zhang Y R, Geng X J, Hao F H, Zhang X, Hanninen H, Guo Y H, De Boeck H J. Decreasing control of precipitation on grassland spring phenology in temperate China. Global Ecology and Biogeography, 2020, 30(2): 490-499. |
| [21] |
Xin Q C, Broich M, Zhu P, Gong P. Modeling grassland spring onset across the Western United States using climate variables and MODIS-derived phenology metrics. Remote Sensing of Environment, 2015, 161: 63-77. DOI:10.1016/j.rse.2015.02.003 |
| [22] |
陈效逑, 庞程, 徐琳, 李静, 张晴华, 尉杨平. 中国温带旱柳物候期对气候变化的时空响应. 生态学报, 2015, 35(11): 3625-3635. |
| [23] |
肖芳, 桑婧, 王海梅. 气候变化对内蒙古鄂温克旗典型草原植物物候的影响. 生态学报, 2020, 40(8): 2784-2792. |
| [24] |
Schaber J, Badeck F W. Physiology-based phenology models for forest tree species in Germany. International Journal of biometeorology, 2003, 47(4): 193-201. DOI:10.1007/s00484-003-0171-5 |
| [25] |
王连喜, 陈怀亮, 李琪, 余卫东. 植物物候与气候研究进展. 生态学报, 2010, 30(2): 447-454. |
| [26] |
Keller F, Körner C. The role of photoperiodism in Alpine plant development. Arctic, Antarctic, and Alpine Research, 2003, 35(3): 361-368. DOI:10.1657/1523-0430(2003)035[0361:TROPIA]2.0.CO;2 |
| [27] |
郑景云, 葛全胜, 郝志新. 气候增暖对我国近40年植物物候变化的影响. 科学通报, 2002, 47(20): 1582-1587. DOI:10.3321/j.issn:0023-074X.2002.20.013 |
| [28] |
白洁, 葛全胜, 戴君虎, 王英. 西安木本植物物候与气候要素的关系. 植物生态学报, 2010, 34(11): 1274-1282. DOI:10.3773/j.issn.1005-264x.2010.11.004 |
| [29] |
王焕炯, 陶泽兴, 葛全胜. 气候波动对西安39种木本植物展叶始期及其积温需求的影响. 植物生态学报, 2019, 43(10): 877-888. DOI:10.17521/cjpe.2019.0178 |
| [30] |
陶泽兴, 葛全胜, 徐韵佳, 王焕炯. 西安和宝鸡木本植物花期物候变化及温度敏感度对比. 生态学报, 2020, 40(11): 3666-3676. |
| [31] |
王传海, 吴飞倩, 李淑娟, 赵国强. 西安植物园木本植物近十余年物候变化的特征分析. 中国农业气象, 2006, 27(4): 261-264. DOI:10.3969/j.issn.1000-6362.2006.04.001 |
| [32] |
Garonna I, De Jong R, De Wit A J W, Mücher C A, Schmid B, Schaepman M E. Strong contribution of autumn phenology to changes in satellite-derived growing season length estimates across Europe (1982-2011). Global Change Biology, 2014, 20(11): 3457-3470. DOI:10.1111/gcb.12625 |
| [33] |
Park H, Jeong S J, Ho C H, Kim J, Brown M E, Schaepman M E. Nonlinear response of vegetation green-up to local temperature variations in temperate and boreal forests in the Northern Hemisphere. Remote Sensing of Environment, 2015, 165: 100-108. DOI:10.1016/j.rse.2015.04.030 |
| [34] |
Guo L, Dai J H, Ranjitkar S, Xu J C, Luedeling E. Response of chestnut phenology in China to climate variation and change. Agricultural and Forest Meteorology, 2013, 180: 164-172. DOI:10.1016/j.agrformet.2013.06.004 |
| [35] |
金丽娜, 曲静, 翟园, 张弘. 西安近63年气候变化特征综合分析. 陕西气象, 2014(3): 17-20. DOI:10.3969/j.issn.1006-4354.2014.03.006 |
| [36] |
张文静, 吴素良, 郝丽, 王琦, 胡琳. 西安城市热岛效应变化特征分析. 陕西气象, 2019(1): 18-21. DOI:10.3969/j.issn.1006-4354.2019.01.003 |
| [37] |
Roetzer T, Wittenzeller M, Haeckel H, Nekovar J. Phenology in central Europe-differences and trends of spring phenophases in urban and rural areas. International Journal of Biometeorology, 2000, 44(2): 60-66. DOI:10.1007/s004840000062 |
| [38] |
Zhang G L, Zhang Y J, Dong J W, Xiao X M. Green-up dates in the Tibetan Plateau have continuously advanced from 1982 to 2011. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(11): 4309-4314. DOI:10.1073/pnas.1210423110 |
| [39] |
Wang X H, Piao S L, Ciais P, Li J S, Friedlingstein P, Koven C, Chen A P. Spring temperature change and its implication in the change of vegetation growth in North America from 1982 to 2006. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(4): 1240-1245. DOI:10.1073/pnas.1014425108 |
| [40] |
Cleland E E, Chuine I, Menzel A, Mooney H A, Schwartz M D. Shifting plant phenology in response to global change. Trends in Ecology & Evolution, 2007, 22(7): 357-365. |
| [41] |
侯美亭, 胡伟, 乔海龙, 李伟光, 延晓冬. 偏最小二乘(PLS)回归方法在中国东部植被变化归因研究中的应用. 自然资源学报, 2015, 30(3): 409-422. |
| [42] |
邓晨晖, 白红英, 高山, 黄晓月, 孟清, 赵婷, 张扬, 苏凯, 郭少壮. 1964-2015年气候因子对秦岭地区植物物候的综合影响效应. 地理学报, 2018, 73(5): 917-931. |
| [43] |
Yu H Y, Luedeling E, Xu J C. Winter and spring warming result in delayed spring phenology on the Tibetan Plateau. Proceedings of the National Academy of Sciences of Sciences of the United States of America, 2010, 107(51): 22151-22156. |
| [44] |
Pérez-Enciso M, Tenenhaus M. Prediction of clinical outcome with microarray data: a partial least squares discriminant analysis (PLS-DA) approach. Human genetics, 2003, 112(5/6): 581-592. |
| [45] |
Johnson R A, Wichern D W. Applied Multivariate Statistical Analysis. New Jersey: Prentice Hall, 2002.
|
| [46] |
赖小红, 李名扬, 刘聪, 钟雨航, 林立, 王海洋. 植物物候对重庆主城区热岛效应的响应. 生态学报, 2019, 39(19): 7025-7034. |
| [47] |
Ding Y H, Ren G Y, Zhao Z C, Xu Y, Luo Y, Li Q P, Zhang J. Detection, causes and projection of climate change over China: an overview of recent progress. Advances in Atmospheric Sciences, 2007, 24(6): 954-971. DOI:10.1007/s00376-007-0954-4 |
| [48] |
Lakatos L. Connection between phenological phases and urban heat island in Debrecen and Szeged, Hungary. Acta Climatologica et Chorologica, 2003, 36/37: 79-88. |
| [49] |
许格希, 裴顺祥, 郭泉水, 牛树奎. 城市热岛效应对气候变暖和植物物候的影响. 世界林业研究, 2011, 24(6): 12-17. |
| [50] |
Liu Q, Fu Y H, Zeng Z Z, Huang M T, Li X R, Piao S L. Temperature, precipitation, and insolation effects on autumn vegetation phenology in temperate China. Global Change Biology, 2016, 22(2): 644-655. DOI:10.1111/gcb.13081 |
| [51] |
季劲钧, 黄玫, 刘青. 气候变化对中国中纬度半干旱草原生产力影响机理的模拟研究. 气象学报, 2005, 63(3): 257-266. |
| [52] |
邓晨晖, 白红英, 马新萍, 黄晓月, 赵婷. 2000-2017年秦岭山地植被物候变化特征及其南北差异. 生态学报, 2021, 41(3): 1068-1080. |
| [53] |
李荣平, 周广胜, 张慧玲. 植物物候研究进展. 应用生态学报, 2006, 17(3): 541-544. DOI:10.3321/j.issn:1001-9332.2006.03.037 |
| [54] |
Kelsey K C, Pedersen S H, Leffler A J, Sexton J O, Feng M, Welker J M. Winter snow and spring temperature have differential effects on vegetation phenology and productivity across Arctic plant communities. Global Change Biology, 2021, 27(8): 1572-1586. DOI:10.1111/gcb.15505 |
| [55] |
Qiao C C, Shen S, Cheng C X, Wu J X, Jia D, Song C Q. Vegetation phenology in the Qilian Mountains and its response to temperature from 1982 to 2014. Remote Sensing, 2021, 13(2): 286. DOI:10.3390/rs13020286 |
| [56] |
卓莉, 张子彦, 雷小雨, 李秋萍, 陶海燕. 基于蒙特卡洛生存分析探究东北森林物候的影响因素. 地理学报, 2019, 74(3): 490-503. |
| [57] |
Hänninen H. Modeling bud dormancy release in trees from cool and temperate regions. Acta Forest Fennica, 1990(213): 7660. |
| [58] |
魏育国, 蒋菊芳, 刘明春, 梁从虎. 河西走廊东部木本植物物候对气候变化的响应. 干旱区研究, 2012, 29(1): 109-114. |
| [59] |
杨丽萍, 代海燕, 陈素华, 秦艳, 吕迪波, 郭晓丽. 气候变化对科尔沁沙地木本植物物候期的影响. 干旱区研究, 2017, 34(3): 518-523. |
| [60] |
Shen M G, Piao S, Dorji T, Liu Q, Cong N, Chen X Q, An S, Wang S P, Wang T, Zhang G X. Plant phenological responses to climate change on the Tibetan Plateau: research status and challenges. National Science Review, 2015, 2(4): 454-467. DOI:10.1093/nsr/nwv058 |
| [61] |
孔冬冬, 张强, 黄文琳, 顾西辉. 1982-2013年青藏高原植被物候变化及气象因素影响. 地理学报, 2017, 72(1): 39-52. |